
Low-Dose Empagliflozin Improves Systolic Heart Function after Myocardial Infarction in Rats: Regulation of MMP9, NHE1, and SERCA2a

 and
and
Abstract
:1. Introduction
2. Results
2.1. Short-Term Hemodynamic Effects of Empagliflozin
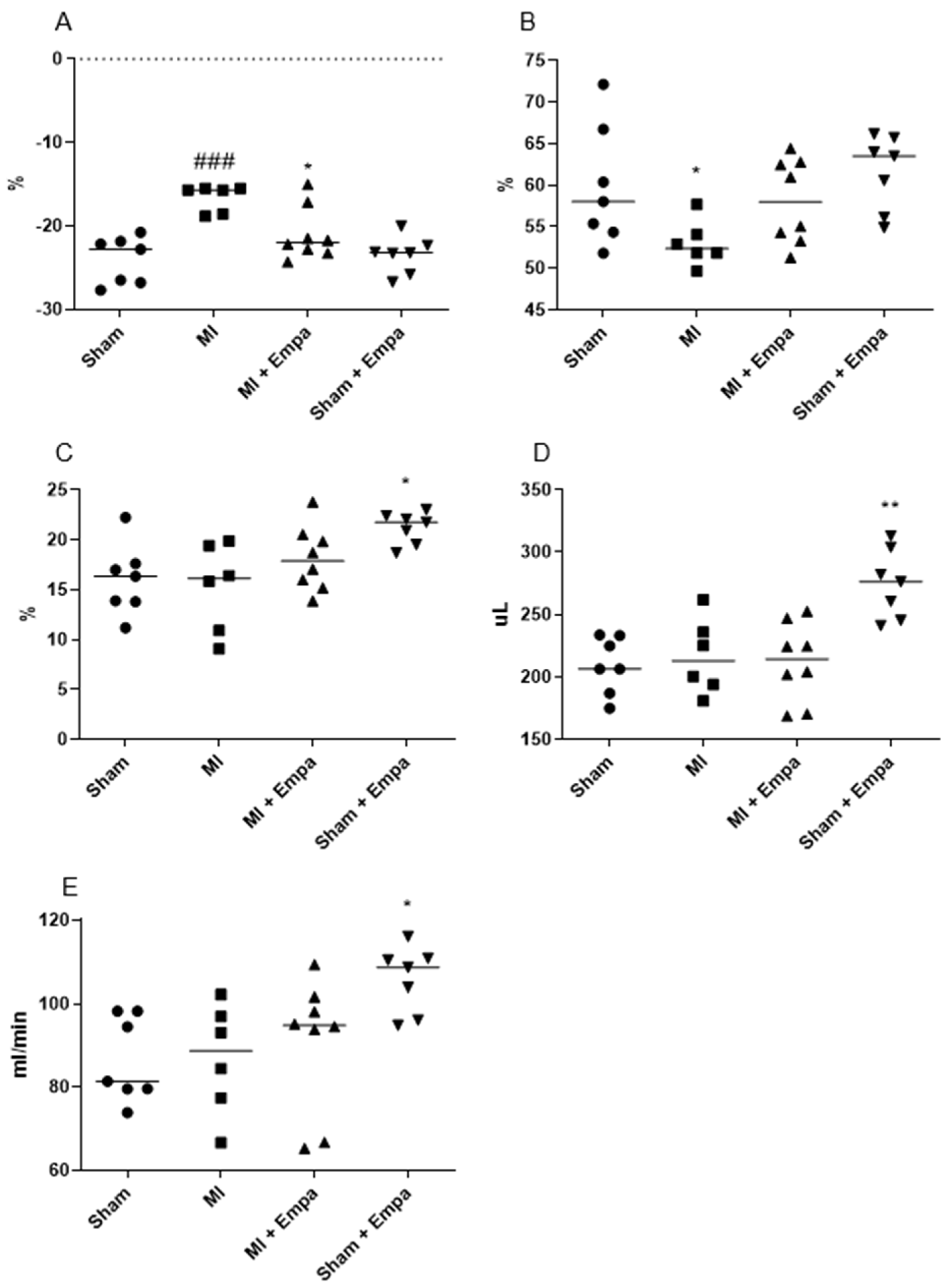
2.2. Hemodynamic Evaluation One Week after Myocardial Infarction
2.3. Regulation of Cardiac Transporters
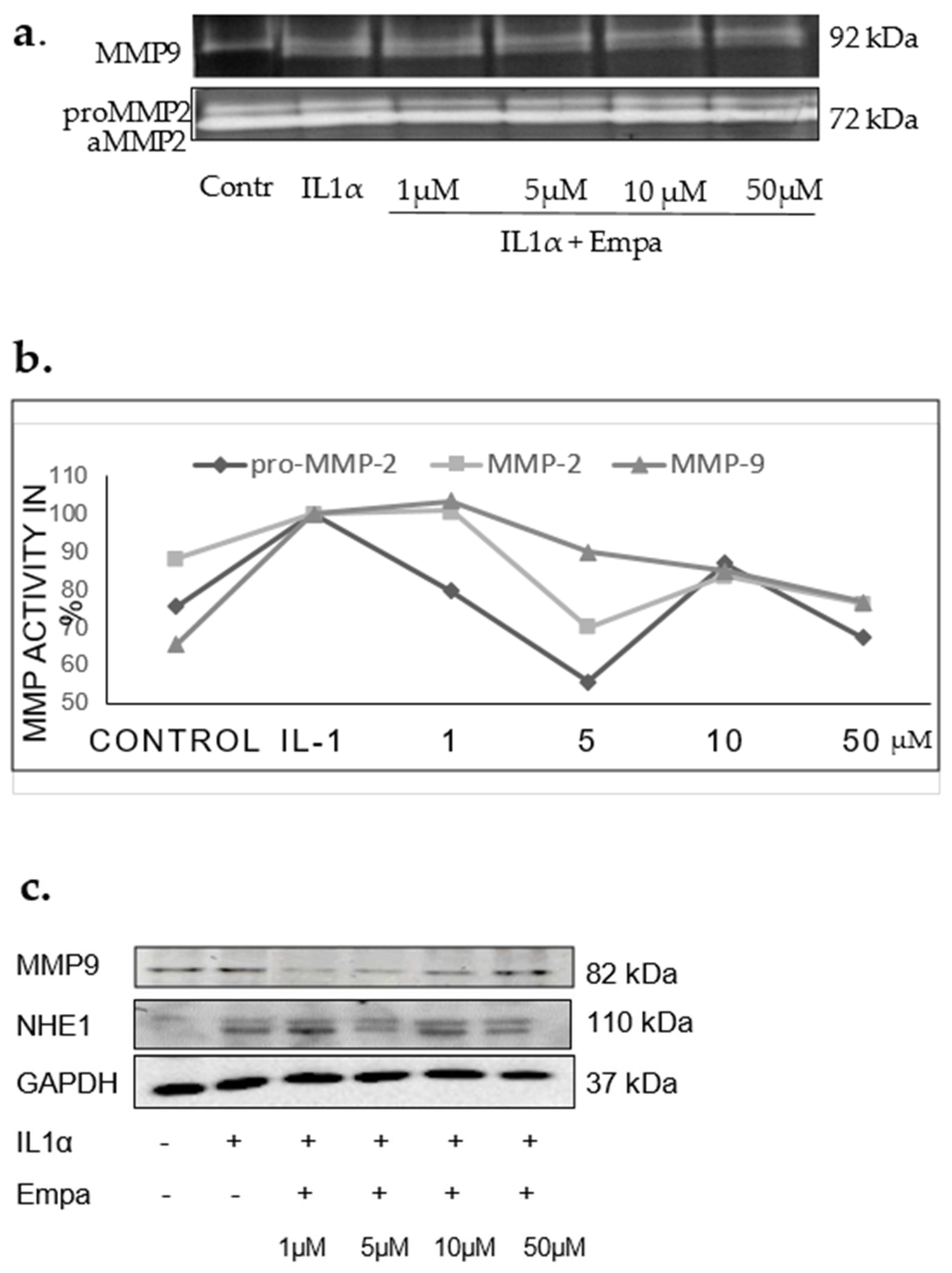
2.4. Regulation of MMP2, MMP9, and TIMP1
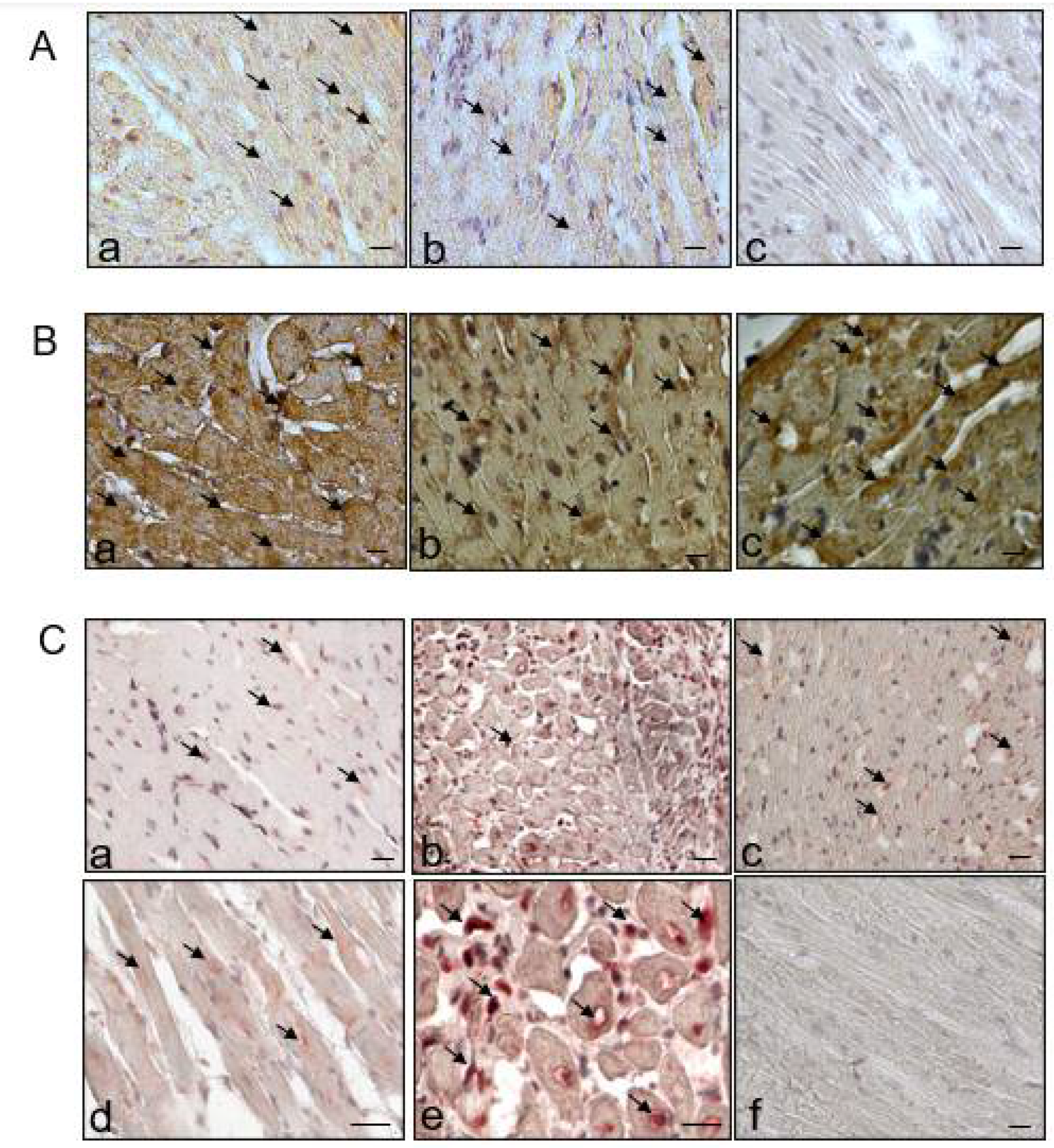
2.5. Regulation of Fibrosis and Necroptosis
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals
4.3. Experimental Protocol
4.4. Analysis of Hemodynamic Parameters
4.5. Glucose Levels
4.6. Immunoblotting Analysis
4.7. Immunohistochemistry
4.8. Cell Culture
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EF | ejection fraction |
| FS | fractional shortening |
| GLS | Global Longitudinal Strain |
| IL-1α | Interleukin 1 alpha |
| max dP/dt | maximal rate of rise of left ventricular pressure |
| min dP/dt | minimal rate of rise of left ventricular pressure |
| MLKL | mixed lineage kinase domain-like |
| MMP | matrix metalloproteinase |
| NBC | sodium bicarbonate cotransporter 1 |
| NCX | Na+/Ca2+ exchanger |
| NHE1 | sodium hydrogen exchanger 1 |
| SERCA2a | Sarco(endo)plasmic reticulum Ca2-ATPase |
| SGLT2 | Selective sodium-glucose cotransporter 2 |
| SV | stroke volume |
| TGF β1 | transforming growth factor beta1 |
| TIMP1 | tissue inhibitor of matrix metalloproteinase 1 |
References
- Zinman, B.; Lachin, J.M.; Inzucchi, S.E. Empagliflozin, cardiovascular outcomes, and mortality in Type 2 Diabetes. N. Engl. J. Med. 2016, 374, 1094. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.; Ghosal, S. Sodium-glucose cotransporter-2 inhibitors (SGLT-2i) reduce hospitalization for heart failure only and have no effect on atherosclerotic cardiovascular events: A meta-analysis. Diabetes Ther. 2019, 10, 891–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahaffey, K.W.; Neal, B.; Perkovic, V.; de Zeeuw, D.; Fulcher, G.; Erondu, N.; Shaw, W.; Fabbrini, E.; Sun, T.; Li, Q.; et al. Canagliflozin for primary and secondary prevention of cardiovascular events: Results from the canvas program (canagliflozin cardiovascular assessment study). Circulation 2018, 137, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiviott, S.D.; Raz, I.; Bonaca, M.P.; Mosenzon, O.; Kato, E.T.; Cahn, A.; Silverman, M.G.; Zelniker, T.A.; Kuder, J.F.; Murphy, S.A.; et al. Dapagliflozin and Cardiovascular Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2019, 380, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Norhammar, A.; Bodegård, J.; Nyström, T.; Thuresson, M.; Nathanson, D.; Eriksson, J.W. Dapagliflozin and cardiovascular mortality and disease outcomes in a population with type 2 diabetes similar to that of the DECLARE-TIMI 58 trial: A nationwide observational study. Diabetes Obes. Metabol. 2019, 21, 1136–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, A.; Vickneson, K.; Singh, J.S. SGLT2-inhibitors; more than just glycosuria and diuresis. Heart Fail Rev. 2021, 26, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Byrne, N.J.; Parajuli, N.; Levasseur, J.L.; Boisvenue, J.; Beker, D.L.; Masson, G.; Fedak, P.W.; Verma, S.; Dyck, J.R. Empagliflozin prevents worsening of cardiac function in an experimental model of pressure overload-induced heart failure. JACC Basic Transl. Sci. 2017, 2, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Connelly, K.A.; Zhang, Y.; Visram, A.; Advani, A.; Batchu, S.N.; Desjardins, J.F.; Thai, K.; Gilbert, R.E. Empagliflozin improves diastolic function in a nondiabetic rodent model of heart failure with preserved ejection fraction. JACC Basic Transl. Sci. 2019, 4, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Pabel, S.; Wagner, S.; Bollenberg, H.; Bengel, P.; Kovacs, A.; Schach, C.; Tirilomis, P.; Mustroph, J.; Renner, A.; Gummert, J.; et al. Empagliflozin directly improves diastolic function in human heart failure. Eur. J. Heart Fail. 2018, 20, 1690–1700. [Google Scholar] [CrossRef] [PubMed]
- Santos-Gallego, C.G.; Requena-Ibanez, J.A.; San Antonio, R.; Ishikawa, K.; Watanabe, S.; Picatoste, B.; Flores, E.; Garcia-Ropero, A.; Sanz, J.; Hajjar, R.J.; et al. Empagliflozin ameliorates adverse left ventricular remodeling in nondiabetic heart failure by enhancing myocardial energetics. J. Am. Coll. Cardiol. 2019, 73, 1931–1944. [Google Scholar] [CrossRef]
- Krasnova, M.; Kulikov, A.; Okovityi, S.; Ivkin, D.; Karpov, A.; Kaschina, E.; Smirnov, A. Comparative efficacy of empagliflozin and drugs of baseline therapy in post-infarct heart failure in normoglycemic rats. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Lim, V.G.; Bell, R.M.; Arjun, S.; Kolatsi-Joannou, M.; Long, D.A.; Yellon, D.M. SGLT2 inhibitor, canagliflozin, attenuates myocardial infarction in the diabetic and nondiabetic heart. JACC Basic Transl Sci. 2019, 4, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Miki, T.; Kuno, A.; Mizuno, M.; Sato, T.; Tanno, M.; Yano, T.; Nakata, K.; Kimura, Y.; Abe, K.; et al. Empagliflozin, an SGLT2 inhibitor, reduced the mortality rate after acute myocardial infarction with modification of cardiac metabolomes and antioxidants in diabetic rats. J. Pharmacol. Exp. Ther. 2019, 368, 524–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurista, S.R.; Silljé, H.H.; Oberdorf-Maass, S.U.; Schouten, E.M.; Pavez Giani, M.G.; Hillebrands, J.L.; van Goor, H.; van Veldhuisen, D.J.; de Boer, R.A.; Westenbrink, B.D. Sodium-glucose co-transporter 2 inhibition with empagliflozin improves cardiac function in non-diabetic rats with left ventricular dysfunction after myocardial infarction. Eur. J. Heart Fail. 2019, 21, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Connelly, K.A.; Zhang, Y.; Desjardins, J.F.; Nghiem, L.; Visram, A.; Batchu, S.N.; Yerra, V.G.; Kabir, G.; Thai, K.; Advani, A.; et al. Load-independent effects of empagliflozin contribute to improved cardiac function in experimental heart failure with reduced ejection fraction. Cardiovasc. Diabetol. 2020, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, M.; Xu, J.; Xu, B.; Kang, L. Empagliflozin prevents from early cardiac injury post myocardial infarction in non-diabetic mice. Eur. J. Pharm. Sci. 2021, 161, 105788. [Google Scholar] [CrossRef] [PubMed]
- Sayour, A.A.; Celeng, C.; Oláh, A.; Ruppert, M.; Merkely, B.; Radovits, T. Sodium-glucose cotransporter 2 inhibitors reduce myocardial infarct size in preclinical animal models of myocardial ischaemia-reperfusion injury: A meta-analysis. Diabetologia 2021, 64, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Li, T.; Wang, Y.; Chang, Y.; Cheng, Y.; Lu, Y.; Liu, X.; Xu, L.; Li, X.; Yu, X.; et al. Empagliflozin prevents cardiomyopathy via sGC-cGMP-PKG pathway in type 2 diabetes mice. Clin. Sci. 2019, 133, 1705–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, M.A.; Chakraborty, P.; Si, D.; Du, B.; Massé, S.; Lai, P.F.; Ha, A.C.; Nanthakumar, K. Anti-arrhythmic and inotropic effects of empagliflozin following myocardial ischemia. Life Sci. 2021, 1, 119440. [Google Scholar] [CrossRef]
- Peng, X.; Li, L.; Zhang, M.; Zhao, Q.; Wu, K.; Bai, R.; Ruan, Y.; Liu, N. Sodium-glucose cotransporter 2 inhibitors potentially prevent atrial fibrillation by ameliorating ion handling and mitochondrial dysfunction. Front. Physiol. 2020, 4, 912. [Google Scholar] [CrossRef]
- Wright, E.M.; Loo, D.D.; Hirayama, B.A. Biology of human sodium glucose transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrhovac, I.; Eror, D.B.; Klessen, D.; Burger, C.; Breljak, D.; Kraus, O.; Radović, N.; Jadrijević, S.; Aleksic, I.; Walles, T.; et al. Localizations of Na(+)-D-glucose cotransporters SGLT1 and SGLT2 in human kidney and of SGLT1 in human small intestine, liver, lung, and heart. Pflug. Arch. 2015, 467, 1881–1898. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; McGaffin, K.R.; Pastor-Soler, N.M.; Ahmad, F. SGLT1 is a novel cardiac glucose transporter that is perturbed in disease states. Cardiovasc. Res. 2009, 84, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippatos, T.D.; Liontos, A.; Papakitsou, I.; Elisaf, M.S. SGLT2 inhibitors and cardioprotection: A matter of debate and multiple hypotheses. Postgrad. Med. 2019, 131, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Chilton, R.; Tikkanen, I.; Cannon, C.P.; Crowe, S.; Woerle, H.J.; Broedl, U.C.; Johansen, O.E. Effects of empagliflozin on blood pressure and markers of arterial stiffness and vascular resistance in patients with type 2 diabetes. Diabetes Obes. Metabol. 2015, 17, 1180–1193. [Google Scholar] [CrossRef] [PubMed]
- Tikkanen, I.; Narko, K.; Zeller, C.; Green, A.; Salsali, A.; Broedl, U.C.; Woerle, H.J. Empagliflozin reduces blood pressure in patients with type 2 diabetes and hypertension. Diabetes Care 2015, 38, 420–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolic, M.; Zivkovic, V.; Jovic, J.J.; Sretenovic, J.; Davidovic, G.; Simovic, S.; Djokovic, D.; Muric, N.; Bolevich, S.; Jakovljevic, V. SGLT2 inhibitors: A focus on cardiac benefits and potential mechanisms. Heart Fail Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Rawat, S.; Ho, K.L.; Wagg, C.S.; Zhang, L.; Teoh, H.; Dyck, J.E.; Uddin, G.M.; Oudit, G.Y.; Mayoux, E.; et al. Empagliflozin increases cardiac energy production in diabetes: Novel translational insights into the heart failure benefits of SGLT2 Inhibitors. JACC Basic Transl. Sci. 2018, 3, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Byrne, N.J.; Matsumura, N.; Maayah, Z.H.; Ferdaoussi, M.; Takahara, S.; Darwesh, A.M.; Levasseur, J.L.; Jahng, J.W.; Vos, D.; Parajuli, N.; et al. Empagliflozin blunts worsening cardiac dysfunction associated with reduced NLRP3 (nucleotide-binding domain-like receptor protein 3) inflammasome activation in heart failure. Circ. Heart Fail. 2020, 13, e006277. [Google Scholar] [CrossRef]
- Li, X.; Lu, Q.; Qiu, Y.; do Carmo, J.M.; Wang, Z.; da Silva, A.A.; Mouton, A.; Omoto, A.C.; Hall, M.E.; Li, J.; et al. Direct cardiac actions of the sodium glucose co-transporter 2 inhibitor empagliflozin improve myocardial oxidative phosphorylation and attenuate pressure-overload heart failure. J. Am. Heart Assoc. 2021, 10, e018298. [Google Scholar] [CrossRef] [PubMed]
- Mustroph, J.; Wagemann, O.; Lücht, C.M.; Trum, M.; Hammer, K.P.; Sag, C.M.; Lebek, S.; Tarnowski, D.; Reinders, J.; Perbellini, F.; et al. Empagliflozin reduces Ca/calmodulin-dependent kinase II activity in isolated ventricular cardiomyocytes. ESC Heart Fail. 2018, 5, 642–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baartscheer, A.; Schumacher, C.A.; Wüst, R.C.; Fiolet, J.W.; Stienen, G.J.; Coronel, R.; Zuurbier, C.J. Empagliflozin decreases myocardial cytoplasmic Na. Diabetologia 2017, 60, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uthman, L.; Nederlof, R.; Eerbeek, O.; Baartscheer, A.; Schumacher, C.; Buchholtz, N.; Hollmann, M.W.; Coronel, R.; Weber, N.C.; Zuurbier, C.J. Delayed ischaemic contracture onset by empagliflozin associates with NHE1 inhibition and is dependent on insulin in isolated mouse hearts. Cardiovasc. Res. 2019, 115, 1533–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neef, S.; Maier, L.S. Novel aspects of excitation-contraction coupling in heart failure. Basic Res. Cardiol. 2013, 108, 360. [Google Scholar] [CrossRef] [PubMed]
- Røe, Å.T.; Ruud, M.; Espe, E.K.; Manfra, O.; Longobardi, S.; Aronsen, J.M.; Nordén, E.S.; Husebye, T.; Kolstad, T.R.S.; Cataliotti, A.; et al. Regional diastolic dysfunction in post-infarction heart failure: Role of local mechanical load and SERCA expression. Cardiovasc. Res. 2019, 115, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Ottolia, M.; Torres, N.; Bridge, J.H.; Philipson, K.D.; Goldhaber, J.I. Na/Ca exchange and contraction of the heart. J. Mol. Cell. Cardiol. 2013, 61, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardo, A.A.; Bernardo, C.M.; Espiritu, D.J.; Arruda, J.A. The sodium bicarbonate cotransporter: Structure, function, and regulation. Semin. Nephrol. 2006, 26, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Ten Hove, M.; Nederhoff, M.G.; van Echteld, C.J. Relative contributions of Na+/H+ exchange and Na+/HCO3- cotransport to ischemic Nai+ overload in isolated rat hearts. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H287–H292. [Google Scholar] [CrossRef] [PubMed]
- Taves, J.; Rastedt, D.; Canine, J.; Mork, D.; Wallert, M.A.; Provost, J.J. Sodium hydrogen exchanger and phospholipase D are required for alpha1-adrenergic receptor stimulation of metalloproteinase-9 and cellular invasion in CCL39 fibroblasts. Arch. Biochem. Biophys. 2008, 477, 60–66. [Google Scholar] [CrossRef]
- Yang, C.C.; Chen, Y.T.; Wallace, C.G.; Chen, K.H.; Cheng, B.C.; Sung, P.H.; Li, Y.C.; Ko, S.F.; Chang, H.W.; Yip, H.K. Early administration of empagliflozin preserved heart function in cardiorenal syndrome in rat. Biomed. Pharmacother. 2019, 109, 658–670. [Google Scholar] [CrossRef]
- Baumgartner, M.; Patel, H.; Barber, D.L. Na(+)/H(+) exchanger NHE1 as plasma membrane scaffold in the assembly of signaling complexes. Am. J. Physiol. Cell. Physiol. 2004, 287, C844–C850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putney, L.K.; Barber, D.L. Expression profile of genes regulated by activity of the Na-H exchanger NHE1. BMC Genom. 2004, 5, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Giordano, F.J.; Hilal-Dandan, R.; Choi, D.J.; Rockman, H.A.; McDonough, P.M.; Bluhm, W.F.; Meyer, M.; Sayen, M.R.; Swanson, E.; et al. Overexpression of the rat sarcoplasmic reticulum Ca2+ ATPase gene in the heart of transgenic mice accelerates calcium transients and cardiac relaxation. J. Clin. Investig. 1997, 100, 380–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prathipati, P.; Metreveli, N.; Nandi, S.S.; Tyagi, S.C.; Mishra, P.K. Ablation of matrix metalloproteinase-9 prevents cardiomyocytes contractile dysfunction in diabetics. Front. Physiol. 2016, 7, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pop, C.; Berce, C.; Ghibu, S.; Pop, A.; Kiss, B.; Irimie, A.; Popa, Ș.T.; Cismaru, G.; Loghin, F.; An, M.Ș. Validation and characterization of a heart failure animal model. Farmacia 2016, 64, 435–443. [Google Scholar]
- Kaschina, E.; Grzesiak, A.; Li, J.; Foryst-Ludwig, A.; Timm, M.; Rompe, F.; Sommerfeld, M.; Kemnitz, U.R.; Curato, C.; Namsolleck, P.; et al. Angiotensin II type 2 receptor stimulation: A novel option of therapeutic interference with the renin-angiotensin system in myocardial infarction? Circulation 2008, 118, 2523–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyhoff, N.; Lohr, D.; Foryst-Ludwig, A.; Klopfleisch, R.; Brix, S.; Grune, J.; Thiele, A.; Erfinanda, L.; Tabuchi, A.; Kuebler, W.M.; et al. Characterization of myocardial microstructure and function in an experimental model of isolated subendocardial damage. Hypertension 2019, 74, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Slavic, S.; Lauer, D.; Sommerfeld, M.; Kemnitz, U.R.; Grzesiak, A.; Trappiel, M.; Thöne-Reineke, C.; Baulmann, J.; Paulis, L.; Kappert, K.; et al. Cannabinoid receptor 1 inhibition improves cardiac function and remodelling after myocardial infarction and in experimental metabolic syndrome. J. Mol. Med. 2013, 91, 811–823. [Google Scholar] [CrossRef] [PubMed]
- Lauer, D.; Slavic, S.; Sommerfeld, M.; Thöne-Reineke, C.; Sharkovska, Y.; Hallberg, A.; Dahlöf, B.; Kintscher, U.; Unger, T.; Steckelings, U.M.; et al. Angiotensin type 2 receptor stimulation ameliorates left ventricular fibrosis and dysfunction via regulation of tissue inhibitor of matrix metalloproteinase 1/matrix metalloproteinase 9 axis and transforming growth factor β1 in the rat heart. Hypertension 2014, 63, e60–e67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vosgerau, U.; Lauer, D.; Unger, T.; Kaschina, E. Cleaved high molecular weight kininogen, a novel factor in the regulation of matrix metalloproteinases in vascular smooth muscle cells. Biochem. Pharmacol. 2010, 79, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham | Sham + Empa | MI + Vehicle | MI + Empa | |
|---|---|---|---|---|
| Heart rate (bpm) | 413 ± 21 | 386 ± 14 | 401 ± 19 | 426 ± 34 |
| EF (%) | 59.9 ± 7.2 | 61.6 ± 4.5 | 53.0 ± 2.7 | 58.0 ± 5.1 |
| GLS (%) | −24.1 ± 2.8 | −23.5 ± 2.2 | −16.6 ± 1.6 ††† | −21.9 ± 3.2 * |
| FS (%) | 16.0 ± 3.5 | 21.2 ± 1.6 † | 15.3 ± 4.4 | 18.1 ± 3.2 |
| SV (uL) | 209.5 ± 22.8 | 274.8 ± 27.6 †† | 216.4 ± 30.2 | 211.7 ± 31.5 |
| CO (mL/min) | 86.5 ± 10.2 | 105.8 ± 7.9 † | 86.8 ± 13.3 | 90.6 ± 16 |
| LVIDd (mm) | 6.6 ± 0.3 | 7.3 ± 0.2 | 6.9 ± 0.5 | 6.9 ± 0.6 |
| LVIDs (mm) | 3.8 ± 0.6 | 3.8 ± 0.6 | 3.6 ± 0.7 | 3.7 ± 0.9 |
| E/A Ratio | 1.71 ± 0.24 | 1.58 ± 0.14 | 1.84 ± 0.31 | 1.69 ± 0.25 |
| IVRT (mm/s) | 0.05120 ± 0.00743 | 0.05123 ± 0.00508 | 0.05715 ± 0.00841 | 0.04585 ± 0.00810 |
| IVCT (mm/s) | 0.04754 ± 0.01319 | 0.04417 ± 0.01057 | 0.04616 ± 0.00893 | 0.03804 ± 0.00965 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goerg, J.; Sommerfeld, M.; Greiner, B.; Lauer, D.; Seckin, Y.; Kulikov, A.; Ivkin, D.; Kintscher, U.; Okovityi, S.; Kaschina, E. Low-Dose Empagliflozin Improves Systolic Heart Function after Myocardial Infarction in Rats: Regulation of MMP9, NHE1, and SERCA2a. Int. J. Mol. Sci. 2021, 22, 5437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115437
Goerg J, Sommerfeld M, Greiner B, Lauer D, Seckin Y, Kulikov A, Ivkin D, Kintscher U, Okovityi S, Kaschina E. Low-Dose Empagliflozin Improves Systolic Heart Function after Myocardial Infarction in Rats: Regulation of MMP9, NHE1, and SERCA2a. International Journal of Molecular Sciences. 2021; 22(11):5437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115437
Chicago/Turabian StyleGoerg, Jana, Manuela Sommerfeld, Bettina Greiner, Dilyara Lauer, Yasemin Seckin, Alexander Kulikov, Dmitry Ivkin, Ulrich Kintscher, Sergey Okovityi, and Elena Kaschina. 2021. "Low-Dose Empagliflozin Improves Systolic Heart Function after Myocardial Infarction in Rats: Regulation of MMP9, NHE1, and SERCA2a" International Journal of Molecular Sciences 22, no. 11: 5437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115437