
Tenascin-C in Heart Diseases—The Role of Inflammation

1
Department of Pathology and Matrix Biology, Mie University Graduate School of Medicine, Tsu, Mie 514-8507, Japan
2
Mie University Research Center for Matrix Biology, Mie University Graduate School of Medicine, Tsu, Mie 514-8507, Japan
Int. J. Mol. Sci. 2021, 22(11), 5828; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115828
Submission received: 4 May 2021
/
Revised: 25 May 2021
/
Accepted: 27 May 2021
/
Published: 29 May 2021
(This article belongs to the Special Issue Matricellular Proteins: Modifiers of Cell Behavior in Development, Disease & Tissue Remodeling)
{kind=link}
{kind=link}
Abstract
:Tenascin-C (TNC) is a large extracellular matrix (ECM) glycoprotein and an original member of the matricellular protein family. TNC is transiently expressed in the heart during embryonic development, but is rarely detected in normal adults; however, its expression is strongly up-regulated with inflammation. Although neither TNC-knockout nor -overexpressing mice show a distinct phenotype, disease models using genetically engineered mice combined with in vitro experiments have revealed multiple significant roles for TNC in responses to injury and myocardial repair, particularly in the regulation of inflammation. In most cases, TNC appears to deteriorate adverse ventricular remodeling by aggravating inflammation/fibrosis. Furthermore, accumulating clinical evidence has shown that high TNC levels predict adverse ventricular remodeling and a poor prognosis in patients with various heart diseases. Since the importance of inflammation has attracted attention in the pathophysiology of heart diseases, this review will focus on the roles of TNC in various types of inflammatory reactions, such as myocardial infarction, hypertensive fibrosis, myocarditis caused by viral infection or autoimmunity, and dilated cardiomyopathy. The utility of TNC as a biomarker for the stratification of myocardial disease conditions and the selection of appropriate therapies will also be discussed from a clinical viewpoint.
1. Introduction
Tenascins are a family of large extracellular matrix (ECM) glycoproteins comprising 4 members: tenascin-C, R, X, and W [1,2,3,4,5]. Tenascin-C (TNC) was the first tenascin to be identified [6] and is the most characterized member of this family (reviewed in [1,7,8,9]). It is also an original member of the ‘matricellular protein’ family together with thrombosondin-1 and SPARC (secreted protein acidic and rich in cysteine; osteonectin) [10,11]. Matricellular proteins are a growing group of non-structural ECM proteins with the following common unique properties: (1) they are strongly up-regulated during embryonic development and tissue remodeling after injury; (2) they serve as biological mediators of cell function by binding to various cell surface receptors, other ECM molecules, or growth factors; and (3) they often induce de-adhesion or counter-adhesion.
One of the characteristics of TNC as a typical matricellular protein is that it is transiently expressed at specific sites during embryogenesis, weakly expressed in normal adults, and up-regulated under pathological conditions in various tissues. This spatiotemporally limited expression pattern is particularly prominent in the heart (reviewed in [12,13]). Accumulating evidence has shown diverse and important roles for TNC in responses to injury as well as cancer stroma [14,15], particularly in the regulation of inflammation [16,17]; however, neither the knockout nor overexpression of TNC induces a distinct phenotype during heart development [18,19].
The significant role of inflammation in the pathophysiological processes of various heart diseases has been attracting attention [20,21] since elevated levels of tumor necrosis factor (TNF) were reported in patients with heart failure for the first time in 1990 [22]. Various infectious pathogens or environmental injuries, such as ischemia and hemodynamic overloading, induce different types of inflammatory/immune responses. Inflammation is a response that facilitates adaptation to abnormal conditions and restores homeostasis and cardiovascular function. However, dysregulated inflammation leads to collateral myocardial damage that ultimately results in progressive ventricular dysfunction and adverse ventricular remodeling.
This review will primarily focus on the roles of TNC in various inflammatory reactions in heart diseases, including myocardial infarction (MI), pressure overload fibrotic hearts, and myocarditis/dilated cardiomyopathy (DCM) from a clinical viewpoint.
2. General Features of TNC in the Heart
TNC is transiently expressed during embryonic heart development and is rarely detected in normal adults, but is re-expressed in diseased hearts (reviewed in [12,13]).
Precardiac mesodermal cells, a special population of cardiomyocytes in embryonic hearts [18], and several cell lines of cardiomyocytes [23,24] have the potential to produce TNC. However, a major source of TNC in the adult pathological heart is residential interstitial fibroblasts [25,26,27]. A number of cardiac remodeling factors, including proinflammatory cytokines/chemokines, growth factors, angiotensin II [28], and endothelin I [29], up-regulate the expression of TNC in cardiac fibroblasts [12,30]. Multiple signaling cascades, including TGF-β/Smad 3/4, Toll-like receptor 4 (TLR4)/NFkB, PDGF/phosphoinositide 3-kinase/Akt, and PDGF/MAPK, may modulate the expression of TNC [2,31,32,33].
3. Embryonic Heart Development and TNC
The transient expression of TNC is often associated with cell migration and epithelial–mesenchymal/mesenchymal–epithelial transition (EMT/MET) in the following important steps of cardiac morphogenesis: (1) the differentiation of cardiomyocytes from the mesoderm, (2) cushion tissue and valve formation, and (3) coronary vascular development.
Precardiac mesodermal cells in the first heart field (FHH) express TNC in association with MET; however, its expression is quickly down-regulated when cells differentiate to express sarcomeric proteins. Cardiomyocytes originating from the secondary heart field in the outflow tract maintain the expression of TNC during the looping and shortening of the primitive heart tube [18].
During the complex morphogenesis of the four-chambered heart, endocardial cells, which form cushion tissue to develop the cardiac septa and valves, express TNC in association with EMT [18]. TNC is expressed at important steps during coronary vasculogenesis. Progenitor cells of the coronary vessels originate from the proepicardial organ (PEO) (reviewed in [34,35]) and migrate along the surface of the primitive heart tube to form the epicardium, which undergoes EMT to give rise to the coronary vasculature and interstitial fibroblasts. TNC is expressed in cells in PEO and its expression is associated with epicardial EMT [18].
A primitive vascular network forms to cover the surface of the ventricle, which is then remodeled and connected to the aorta to form the proximal region of coronary arteries [36,37,38]. During this process, TNC appears to guide developing vascular channels due to its spatial relationship: the vascular network is not formed in the TNC-positive outflow tract and eventually reaches the base of the aorta with the shortening of the TNC-positive outflow tract. TNC is also up-regulated with the maturation of coronary arteries and promotes the recruitment of vascular mural cells through PDGF-B-B/PDGF receptor β [39,40,41]. Although this strictly limited spatio-temporal expression suggests essential roles for TNC during embryonic heart development, its deletion or overexpression does not induce distinct changes in cardiogenesis [18,19].
Two different lines of TNC-deficient mice have been generated [42,43] and neither show a distinct morphological phenotype. However, careful analyses demonstrated that a deficiency of TNC may induce minor differences in the organogenesis of the prostate gland [44] and lungs [45,46], behavioral abnormalities [47,48,49], and pulmonary functional dysfunction in adulthood [50]. In comparisons with wild-type mice, no prominent differences were detected in cardiac morphology, histology, and function or in blood pressure in adult TNC-null mice [23,51,52,53,54,55,56,57] or in TNC-overexpressing mice [19].
4. Myocardial Infarction
4.1. Acute Myocardial Infarction and TNC
MI is the death of a part of the myocardium due to prolonged severe ischemia caused by coronary arterial occlusion, which induces typical acute sterile inflammation and subsequent myocardial repair.
In the early stage after infarction, at approximately 1–3 days, matrix metalloproteinases (MMPs) are activated and generate the fragmentation of ECM proteins to form a provisional matrix, which serves as a scaffold for cell adhesion. At approximately 3–7 days, inflammatory cells, primarily macrophages, actively remove necrotic myocytes (the inflammatory phase). Between 1–2 weeks, the damaged area is progressively replaced by the ingrowth of highly vascularized granulation tissue. Inflammatory cells are cleared, followed by the migration and proliferation of fibroblasts/myofibroblasts and vascular cells (the proliferative phase). The density of myofibroblasts is reduced and various matrix molecules in the infarct area interact and form mature collagen fibers, leading to scar tissue (the maturation phase) [13].
The expression of TNC becomes detectable within 24 h of the onset of infarction in experimental animal models, peaks on days 3–5, and disappears by day 28 [25,54,58]. Serum levels of TNC in human patients reflect this time course of expression [59]. TNC is exclusively expressed at the border zone between an infarct lesion and the intact myocardium [25,54,57,60,61].
The timing and location of the expression of TNC suggest that it plays special roles in the inflammatory phase to the early proliferative phase after MI. Several roles have been described for TNC during tissue repair after MI.
As a typical “de-adhesion” molecule (reviewed in [7,62]), TNC may loosen the adhesion of surviving cardiomyocytes from the surrounding connective tissue. In the normal heart, cardiomyocytes firmly adhere to the surrounding connective tissue via a costamere adhesion complex [63,64,65]. Although TNC does not break this strong adhesion by itself [25], it may free cardiomyocytes by promoting the expression of MMP-2 and MMP-9 [23], thereby facilitating the movement of cells for rearrangement. TNC simultaneously adheres weakly to cardiomyocytes, similar to a ‘post it’ [25,30], and may protect against anoikis.
TNC may also protect cardiomyocytes as a shock absorber. The border zone may be exposed to pathological mechanical stress due to a difference in the physical properties of the intact myocardium and necrotic tissue [66]. An adaptive and protective role for TNC in destructive mechanical stress through its elastic properties [67,68] has been suggested in aortic tissue [69].
The most well-documented role of TNC may be in the regulation of immune/inflammatory reactions in various pathologies, including cancer stroma [14,15]. In the inflammatory phase after MI, cardiomyocyte necrosis results in the release of the molecular signals, damage-associated molecular patterns (DAMPs) [70,71,72], which trigger inflammation-driven fibrotic responses [73,74]. TNC functions as a DAMP and is one of the ligands for TLR4 [72,75,76]. TNC induces the synthesis of proinflammatory cytokines through
TLR4 [55,57,75,76,77,78,79] and up-regulates the expression of MMPs in various types of cells, including macrophages [77,80].
Monocytes/macrophages are the main players in post-infarct inflammation [81]. A heterogeneous population of macrophages have been shown to infiltrate infarcted lesions [82,83,84,85,86], and are broadly classified as M1 or M2 macrophages [87]. M1 macrophages are associated with pro-inflammatory responses and are referred to as “classically activated macrophages”, while M2 macrophages, or “alternatively activated” macrophages, exhibit an anti-inflammatory and pro-regenerative phenotype [88]. On days 1–3 post-MI, M1-like macrophages primarily infiltrate the injured myocardium, drive inflammation, and remove damaged tissue. On approximately days 5–7 post-MI, infiltrating macrophages predominantly exhibit the M2-like phenotype [57,89]. A well-controlled switch from M1 to M2 is necessary for the appropriate resolution of inflammation and the facilitation of tissue healing [90].
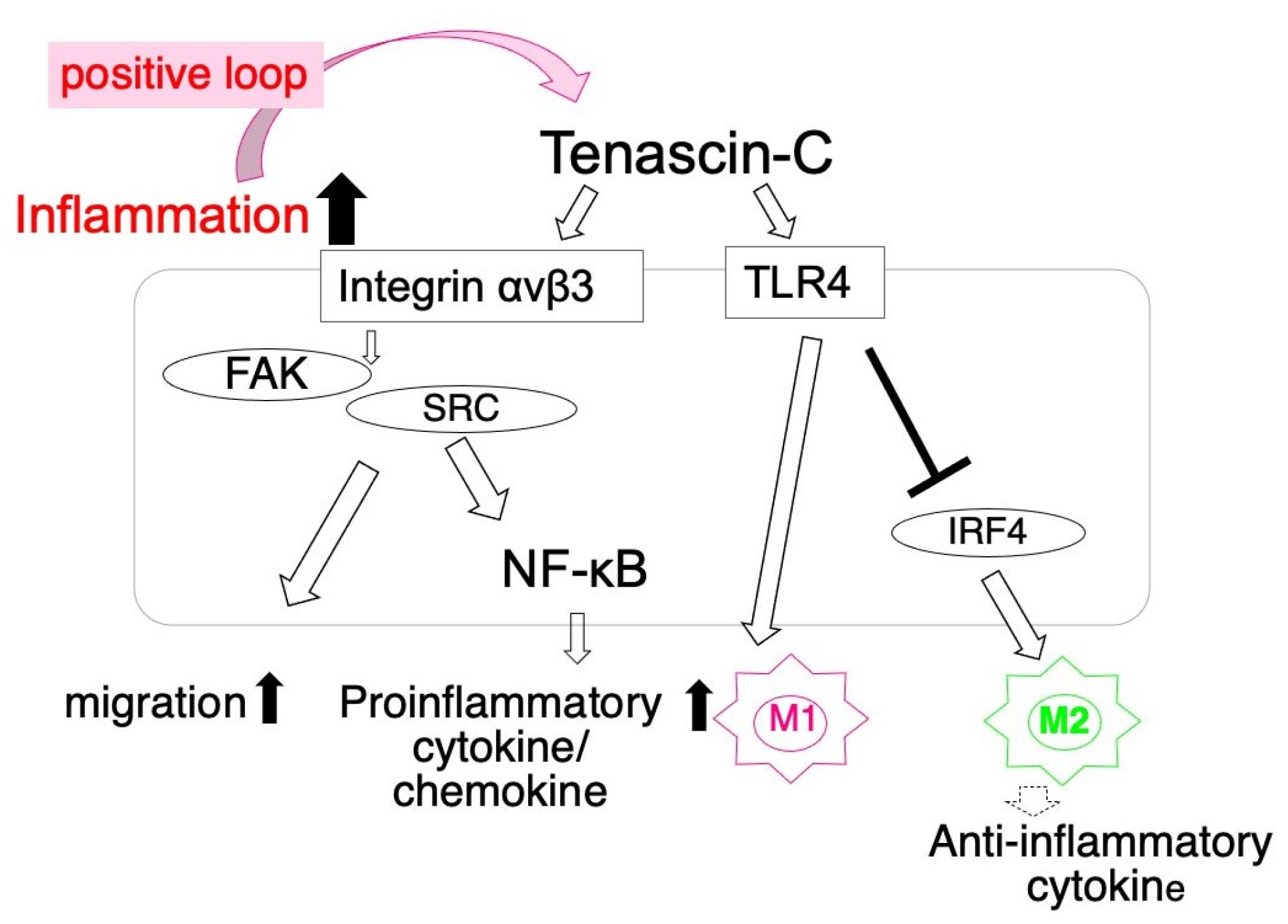
TNC enhances the pro-inflammatory phenotype of macrophages. Its deletion inhibited the expansion of M1, but enhanced that of M2 in a MI mouse model. In vitro, TNC/TLR4 enhanced the M1 macrophage polarization of bone marrow-derived macrophages, but inhibited the up-regulation of the M2 macrophage marker by suppressing interferon regulatory factor 4 (IRF4) [57]. TNC also accelerated the migration and up-regulated the synthesis of proinflammatory cytokines/chemokines via integrin αvβ3 of peritoneal macrophages [56]. Furthermore, proinflammatory cytokines up-regulated the expression of TNC, which may amplify inflammatory responses by creating a positive feedback loop [13,91] (Figure 1). TNC also induced the activation of inflammasomes through TRL4 in epicardial cells [92]. The overexpression of TNC in the mouse heart was shown to up-regulate the expression of proinflammatory cytokines/chemokines and increase mortality rates during the acute stage after MI [19]. Collectively, these findings indicate that TNC enhances proinflammatory responses during the acute stage after infarction.
Although the TNC/TRL4 axis drives inflammation, it also plays an important role in tissue repair [93]. Myocardial repair is primarily dependent on the recruitment and activation of fibroblasts/myofibroblasts because of the limited regenerative ability of cardiomyocytes. Myofibroblasts are specialized fibroblasts that express smooth muscle cell-specific proteins, such as α-smooth muscle actin (SMA), generate contract forces to minimize injured sites, and synthesize collagen to replace the tissue defect [94].
TNC may facilitate the recruitment of myofibroblasts in various tissues [95]. In a MI model of TNC knockout mice, the appearance of myofibroblasts in the injured heart was later than in wild-type mice [61]. In vitro, TNC accelerated the migration of fibroblasts, their conversion to myofibroblasts, as well as contractile force generation and collagen synthesis by fibroblasts [39,61,92,93], at least partly via the integrin αvβ1/TGF-β/smad2/3 cascade [96]. In terms of fibroblasts/myofibroblasts, TNC promoted myocardial repair by facilitating healing activities.
4.2. Post-Infarct Ventricular Remodeling and TNC
The most important clinical issue in patients with MI is post-infarct ventricular remodeling, which refers to the progressive dilatation of ventricles associated with systolic dysfunction weeks or months after infarction.
Previous clinical studies suggested that TNC contributes to the progression of adverse ventricular remodeling. MI patients with high serum levels of TNC in the acute stage were found to be at an increased risk of ventricular dilatation in the chronic stage and had a poor long-term prognosis [59,97]. Mouse model experiments revealed that post-infarct remodeling was attenuated in TNC knockout mice [51,54,57], suggesting that TNC is a harmful molecule that has a negative impact on ventricular remodeling. Dilatation of the ventricular chamber in the acute stage after infarction is explained by the rearrangement (slippage) of cardiomyocytes and degradation of the interstitial matrix by MMPs (reviewed in [98,99]). As a de-adhesion molecule, TNC may augment slippage [25,30]. Furthermore, the over-activation and inappropriate resolution of inflammation by the proinflammatory functions of TNC may also augment tissue destruction. However, difficulties are associated with elucidating the mechanisms by which TNC exacerbates the progression of post-infarct ventricular remodeling in the chronic stage because its expression is limited in the acute stage. A mouse model of post-infarct ventricular remodeling demonstrated that wild-type mice showed significantly poorer function and greater dilatation of the ventricles 3 months after MI than TNC-KO mice; however, the expression of TNC in the wild type had disappeared by this time point. Furthermore, no significant differences were observed in IL-6, TNF-α, or CCL2 levels between wild-type and TNC-KO mice at this time point [69]. Ventricular remodeling is a complex mechanism. The dysregulated process from inflammation to tissue repair during the acute stage may result in physically weak scar tissue at the healed stage, and TNC may at least partly, but significantly, contribute to adverse post-infarct ventricular remodeling.
5. Hypertensive Cardiac Fibrosis and TNC
Cardiac fibrosis is defined as the excessive deposition of collagen fibers in the myocardium and is observed in virtually all pathological hearts [86,100], such as MI, pressure overload [101], diabetes mellitus [102], hypertrophic cardiomyopathy [103], and DCM [104,105].
It is generally classified into 2 types: replacement/reparative and reactive fibrosis [106]. Scar formation after MI, discussed in the previous section, is a typical example of replacement/reparative fibrosis. In contrast, reactive fibrosis describes an increase in perivascular and interstitial collagen fibers in the absence of the significant loss of cardiomyocytes. This type of fibrosis is often observed in pressure or volume overload, aging, and diabetes mellitus. However, it is practically difficult to distinguish between reactive interstitial reactive fibrosis and replacement fibrosis caused by small infarct lesion low-flow ischemia in the advanced stages of myocardial diseases. In any case, similar molecular mechanisms may mediate fibrosis.
Previous findings suggested that ‘low-grade (smoldering) chronic inflammation’ is involved in the progression of reactive fibrosis in the pressure-overloaded hypertrophic heart [107,108,109]. This type of inflammation is characterized by the recruitment of immune cells, particularly macrophages, at the perivascular area. One of the well-studied neurohumoral cascades that link mechanical stress and inflammation/fibrosis is the renin/aldosterone/angiotensin system (RAAS). An angiotensin II (ANG II) or aldosterone infusion model as well as a transverse aortic constriction (TAC) model are often used to investigate the cellular and molecular mechanisms underlying reactive fibrosis.
In the ANG II-induced hypertensive hearts of mice, collagen fibers at the perivascular area increase, extend between individual myocytes, and are associated with macrophage infiltration. Interstitial cells synthesize TNC at perivascular lesions, which is then deposited at the vicinity of macrophages [28,56]. TNC stimulates macrophages in a paracrine manner via the integrin αVβ3/FAK/Src/Nuclear Factor-κB axis to facilitate the migration and expression of proinflammatory cytokines, including IL-6. IL-6 is also known as a pro-fibrotic cytokine that stimulates fibroblasts to synthesize collagen, leading to fibrosis [56]. Proinflammatory cytokines released from macrophages up-regulate the synthesis of TNC by cardiac fibroblasts in a paracrine manner, such that TNC may facilitate fibrosis by promoting inflammation with a positive feedback loop (Figure 1). Furthermore, TNC may directly stimulate fibroblasts via integrin αVβ1/TGFβ/smad2/3 [96] and PDGF-A-B/PDGF-receptor β [28] to augment reactive fibrosis.
In support of this schema of inflammation/fibrosis, a previous study using BALB/c mice with the knockout of TNC showed that ANG-II induced inflammation and fibrosis were less severe than in wild-type mice [56]. The genetic background is important in immune responses. BALB/c mice favor Th2 responses, whereas C57BL/6 mice show a preference for Th1 responses [110]. This difference may influence inflammatory/fibrotic responses in disease models and regulatory mechanisms. However, similar findings were observed in the TAC model using C57BL/6 mice with the knockout of TNC, suggesting proinflammatory and profibrotic roles for TNC [23,52,111]. In contrast, fibrosis was exacerbated by enhanced inflammation in the TAC model using C57BL/6 mice with the knockout of TNC [53].
This may be explained by the context dependency of TNC; it sometimes exhibits opposing functions depending on the experimental design, which contributes to the difficulties associated with clarifying the functions of this molecule.
6. Myocarditis
6.1. Acute Myocarditis and TNC
Myocarditis is an inflammatory condition in the myocardium that is histopathologically defined by inflammatory cell infiltration adjacent to cardiomyocytes and is classified as follows based on the predominant type of infiltrating cell: lymphocytic, eosinophilic, giant cell, or granulomatous [112].Lymphocytic myocarditis is the most common type and is characterized by the predominant infiltration of CD4- and CD8-positive T lymphocytes associated with CD68-positive macrophages [113]. However, the cell profile of infiltrates may vary according to the stage of disease [114].
Myocarditis may be caused by various infectious agents, exposure to drug treatments, and immune disorders [115]. Immune checkpoint inhibitor-related myocarditis has recently been attracting increasing attention in onco-cardiology [116,117].
Viral infection is a major cause of acute lymphocytic myocarditis in the developed world [118]. Cardiotropic viruses, such as coxsackievirus B3 (CVB3) and adenoviruses, have been implicated as a trigger of myocarditis. However, vasculotropic viruses, such as parvovirus B19, and lymphotropic viruses, including human herpesvirus 6, in patients with myocarditis are now being more frequently reported [115,119]. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) may cause cardiomyocyte injury via immune responses or macro/microvascular thrombi rather than by direct virus-mediated cytotoxicity [115,119]; however, the classic type of acute lymphocytic myocarditis is rare [120,121].
Mouse models of CVB3 infection have been used to investigate the pathogenesis of viral myocarditis [122,123].Within 3–4 days of inoculation, CVB3 enters cardiomyocytes by binding to the coxsackievirus-adenovirus receptor and directly causes cytotoxicity.
Cell injury activates innate immunity via pattern recognition receptors, such as TLRs, and various proinflammatory cytokines are released. Antigen-specific responses in adaptive immune cells, mostly T cells, then eliminate the virus and infected cells for up to 14 days (reviewed in [124]). Furthermore, as a potential consequence of expanding self-antigen exposure from virus-infected cardiomyocytes or epitope cross-reactivity, CVB3 infection may increase the number of autoreactive CD4+ T cells for multiple antigens, which contributes to the development of myocarditis [125,126].
The myosin heavy chain α isoform (MyHC-α) is a major cardiac autoantigen. MyHC-α-reactive T-cell numbers were previously reported to be markedly increased in myocarditis patients [127]. MyHC-α immunization with immune adjuvants or an injection of MyHC-α-loaded dendritic cells (DCs) induced autoimmune myocarditis in mice [128,129]. Therefore, virus-triggered immune reactions may be the principle cause of cardiomyocyte injury rather than direct virus-mediated cell injury [115]; however, the pathophysiology of myocarditis may vary depending on the causative virus.
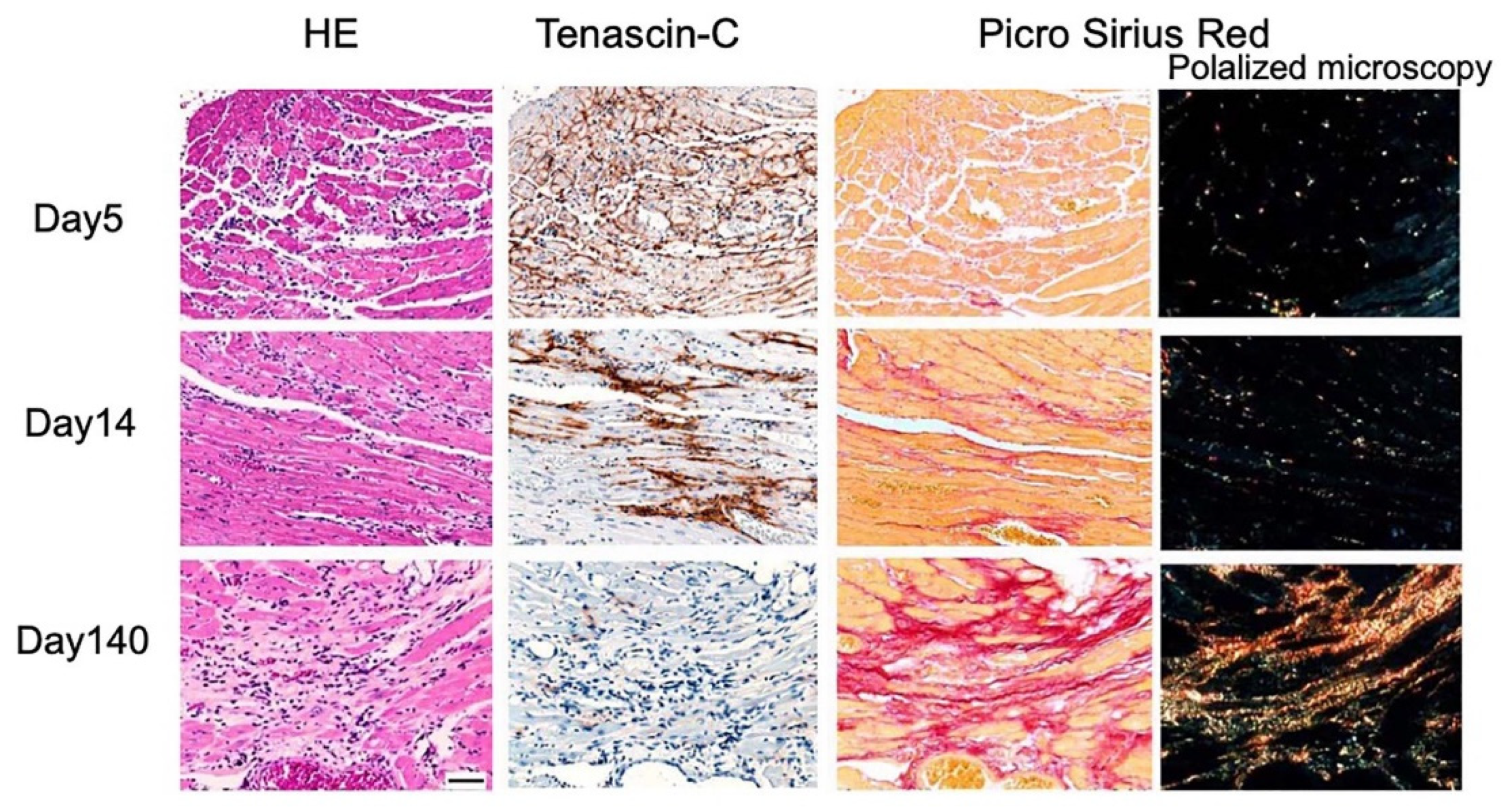
In a mouse CVB3 myocarditis model, the expression of TNC was detected in the early stage after the inoculation of the virus (Figure 2).
Recent studies suggested the involvement of TNC in the pathophysiological mechanism of viral infection. Mills et al. showed that a viral infection may directly induce the expression of TNC [130]. A synthetic viral mimic of human rhinoviruses induced TNC and TN-C-rich small extracellular vesicles, such as exosomes released from bronchial epithelial cells, via a stimulation with TLR3.
Viral infection-induced TNC may be involved in defense mechanisms against viral infection. TNC in breast milk binds to the HIV-1 envelope via fibrinogen-like globe (fbg) and fibronectin-type III (fn) domains, which neutralize HIV [131,132,133]. In contrast, recent findings indicated that TNC was delivered to the cells of distant organs in COVID-19 patients through plasma exosomes and may trigger pro-inflammatory cytokine signaling as a contributory factor to the cytokine storm [134]. Furthermore, TNC may modify the disease state of viral infection by interacting with other pathogens. For example, TNC binds to Streptococcus pyogenes (group A Streptococcus; GAS) and may increase GAS colonization, leading to GAS-influenza-A superinfection [135].
In mouse CVB3 myocarditis, the expression of TNC persisted during active inflammation after the elimination of the virus from the tissue [136] and disappeared in the healed phase (Figure 2), which was similar to that observed in an experimental mouse autoimmune myocarditis model [26]. The up-regulated expression of TNC at focal active sites of inflammation has been reported in endomyocardial biopsy specimens from human myocarditis patients [27], including immune checkpoint inhibitor-induced myocarditis [137]. Cross bone marrow transplantation experiments in mice revealed that the major source of TNC was residential fibroblasts in the heart and not bone marrow-derived inflammatory cells [13,111].
Locally synthesized TNC plays an important role in regulating immune/inflammatory responses in myocarditis. TNC may trigger inflammation [75] as a DAMP molecule and amplify the inflammatory responses of macrophages by creating a positive feedback loop [13,91], as discussed in an earlier section. Furthermore, TNC activates DCs to generate pathogenic autoreactive T cells and forms an important link between innate and acquired immunity [55,138]. DCs stimulated by TNC have been shown to produce various proinflammatory cytokines, including IL-6, which, in turn, induce naïve CD4+ T cells to differentiate into Th17 cells. Th17 cells are closely associated with autoimmunity and play a major role in myocarditis [139,140].
TNC also exerts diverse effects on lymphocytes (reviewed in [141]). It has been shown to support lymphocyte rolling [142], lymphoid progenitor cell homing, and peripheral T-cell migration [143,144], and also promotes T-cell activation and polarization [75,145,146,147,148], but inhibits the proliferation of CD4+ T cells [141].
The deletion of TNC may attenuate the severity of myocarditis by reducing Th17 cell infiltration in the mouse heart [55], suggesting that TNC adversely affects the status of myocarditis; however, it may also have an immunosuppressive function depending on both the inflammatory context and splicoform expressed [15,149].
6.2. Chronic Myocarditis, Inflammatory DCM(Idcm), Heart Failure and TNC
Acute inflammation is a protective response to infection and is typically self-limiting. However, the failure to clear viruses from the heart or dysregulation of the immune system may result in chronic myocarditis. Although the mechanisms underlying the transition from acute to chronic inflammation currently remain unclear, TNC may play a role in the persistence of inflammation [75]. The expression of TNC has been detected in a mouse model of chronic myocarditis induced by recombinant Bacille Calmette-Guérin (BCG) expressing a cardiac autoantigen [150] as well as in endomyocardial biopsy specimens from patients with chronic myocarditis [151]. Prolonged inflammation in chronic myocarditis induces progressive tissue destruction, eventually resulting in the clinical phenotypes of DCM with severe heart failure. DCM is a clinically defined heterogeneous group of myocardial diseases with left ventricular dilatation and contractile dysfunction [152]. DCM patients often have genetic mutations [153], although some cases may be attributed to chronic myocarditis [113]. Chronic myocarditis needs to be distinguished from other types of DCM because immunosuppressive agents may control disease progression in chronic myocarditis and eliminate the need for heart transplantation, whereas gene mutations cannot be treated by immunosuppression.
In addition to recognizing the importance of inflammation in the progression of heart failure, the term ‘iDCM’ is often used for a subgroup of DCM associated with inflammation. ‘Inflammation’ is detected in endomyocardial biopsy samples from iDCM patients, who have a poorer prognosis than DCM patients without inflammation [154,155]. However, it is important to note that iDCM and chronic myocarditis may not be synonymous. In several cases of DCM, gene mutations in the compositional elements of cardiomyocytes may contribute to vulnerability to stress in the failing heart, resulting in cell injury or death, which triggers innate immune responses. Although inflammation is an essential response during tissue repair, inflammatory mediators released during reactive inflammation in DCM hearts may further aggravate heart failure, resulting in a vicious cycle of inflammation [113]. Therefore, iDCM may be part of this pathological condition and the type of inflammation may differ from that of narrowly defined myocarditis predominantly mediated by acquired immunity, as discussed above. TNC was previously shown to be expressed in iDCM hearts and was associated with worse LV remodeling and long-term outcomes in DCM [156]. Since TNC amplifies inflammatory responses by creating several positive feedback loops in various pathophysiologies, it may enhance the vicious cycle in iDCM.
7. Clinical Application of TNC
TNC has potential as a diagnostic marker and target for the molecular imaging of inflammation based on its specific expression, which may facilitate the differentiation of myocarditis from DCM.
Immunostaining for TNC is useful for a precise diagnosis of active inflammation in biopsy samples of the myocardium from myocarditis and DCM patients [27,156,157]. A combination with molecular imaging for TNC may reduce the sampling errors of endomyocardial biopsy by clarifying the target site for biopsy in the heart. To date, several TNC-specific monoclonal antibodies or antibody fragments (scFv) have been developed [158,159] and are undergoing evaluations for inflammation imaging in the heart [160,161,162,163]. Furthermore, serum/plasma TNC may be a practical diagnostic biomarker. Elevated levels of TNC in the blood may reflect the up-regulated expression of TNC in the inflammatory lesions of DCM and predict worse left ventricular remodeling and long-term outcomes [156,164,165,166].
Similarly, TNC is applicable as a biomarker to detect acute and chronic rejection after human cardiac transplantation in association with inflammation [167].
A recent study reported that serum TNC may predict an increased risk of cardiovascular disease in patients with type 2 diabetes associated with chronic low-grade inflammation [168]. It is also important to note that TNC may also be useful for evaluating the disease condition of heart failure with a preserved ejection fraction (HFpEF) [169]. HFpEF is a leading cause of morbidity and mortality throughout the industrialized world. Although HFpEF is a heterogeneous group of heart diseases, obesity and type 2 diabetes mellitus are comorbidities in most patients. A recent study proposed that HFpEF is a multisystem disorder involving up-regulated immune and inflammatory signaling [170].
Serum TNC may be useful for the management of patients with arrythmia because fibrosis is considered to play a central role in stabilizing the re-entrant drivers that maintain arrhythmia [171]. TNC levels were found to be higher in patients with malignant arrythmia than in those with benign arrythmia [172]. In patients with atrial fibrillation, TNC levels in the left and right atria correlated with the severity of atrial dilatation [173].
An increasing number of studies have examined the clinical utility of serum TNC levels in patients with various heart diseases (reviewed in [149,174,175]), such as non-compaction/hypertrabeculation [176], hypertrophic cardiomyopathy [177], heart failure with ischemic heart disease, Kawasaki disease [178], rheumatic heart disease, and congenital heart disease in pediatric patients [179,180].
Many studies demonstrated that high serum TNC levels predicted adverse ventricular remodeling and increased the risk of death and major adverse cardiovascular events, suggesting that TNC is not only a marker for tissue damage and inflammation, it is also a ‘remodeling’ marker. In contrast, decreased TNC levels may be useful for evaluating reverse remodeling [181]. A recent study demonstrated that decreased TNC levels reflect the effectiveness of intravenous immunoglobulin therapy for Kawasaki disease and are useful as an indicator for assessing whether inflammation has decreased [182].
Moreover, the combination of serum TNC levels and plasma BNP levels is a stronger predictor than either biomarker alone in AMI and DCM [97,165]. This may be explained as follows: while BNP is secreted from stressed cardiomyocytes, TNC is released by activated interstitial fibroblasts. Interstitial fibroblasts may play a major role in the reconstruction of the injured heart because the regenerative ability of cardiomyocytes is very limited. Therefore, a combination of the two biomarkers may contribute to a more precise assessment of the whole heart by evaluating cardiomyocyte stress and the interstitial repair reaction.
Other matricellular proteins, such as galectin-3 [183], periostin [184], and osteopontin [185,186] have also been proposed as cardiovascular biomarkers. Galectin-3 is recommended by the 2017 Guidelines of the American Heart Association for the risk stratification and prognostic evaluation of patients with heart failure [187]. These matricellular proteins are up-regulated by tissue injury and inflammation and show a similar expression pattern to that of TNC. They sometimes localize with TNC and may interact. Periostin [188] and galectin-3 [189] directly bind to TNC, appear to regulate the expression levels of each other [33], and play a role in the regulation of inflammation and fibrosis. Furthermore, osteopontin and TNC have been shown to share some receptors and sometimes exert opposite effects [190]. They may regulate tissue inflammatory fibrotic responses by cooperating or counterbalancing each other. Therefore, each biomarker may reflect different cellular activities during myocardial tissue remodeling. The combination of these matricellular biomarkers may be useful for the more precise stratification of myocardial disease conditions and selection of appropriate therapies.
8. Conclusions
TNC may be a feasible diagnostic biomarker and target for the molecular imaging of inflammation because of its specific expression associated with inflammation, which may contribute to the stratification of patients with heart diseases and the selection of appropriate therapy.
Experimental studies using TNC-knockout and -overexpressing mice suggest that TNC is a noxious molecule in the heart because it aggravates inflammation/fibrosis in most cases. However, TNC has diverse functions and, thus, may exert both harmful and protective effects in the heart. The inhibition of the proinflammatory functions of TNC with appropriate timing may provide novel strategies that prevent the progression of heart failure.
Funding
This work was funded in part by grants from JSPS KAKENHI (Grant Number JP19H03442), AMED (Grant Number 20ek0109476h0001) and Japan Heart Foundation Research Grant on Dilated Cardiomyopathy.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Tucker, R.P.; Drabikowski, K.; Hess, J.F.; Ferralli, J.; Chiquet-Ehrismann, R.; Adams, J.C. Phylogenetic analysis of the tenascin gene family: Evidence of origin early in the chordate lineage. BMC Evol. Biol. 2006, 6, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiovaro, F.; Chiquet-Ehrismann, R.; Chiquet, M. Transcriptional regulation of tenascin genes. Cell Adh. Migr. 2015, 9, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Rathjen, F.G.; Hodge, R. Early Days of Tenascin-R Research: Two Approaches Discovered and Shed Light on Tenascin-R. Front. Immunol. 2020, 11, 612482. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Tenascin-X-Discovery and Early Research. Front. Immunol. 2020, 11, 612497. [Google Scholar] [CrossRef]
- Degen, M.; Scherberich, A.; Tucker, R.P. Tenascin-W: Discovery, Evolution, and Future Prospects. Front. Immunol. 2020, 11, 623305. [Google Scholar] [CrossRef] [PubMed]
- Chiquet-Ehrismann, R.; Mackie, E.J.; Pearson, C.A.; Sakakura, T. Tenascin: An extracellular matrix protein involved in tissue interactions during fetal development and oncogenesis. Cell 1986, 47, 131–139. [Google Scholar] [CrossRef]
- Chiquet-Ehrismann, R.; Tucker, R.P. Tenascins and the importance of adhesion modulation. Cold Spring Harb. Perspect. Biol. 2011, 3, a004960. [Google Scholar] [CrossRef] [Green Version]
- Chiquet, M. Tenascin-C: From Discovery to Structure-Function Relationships. Front. Immunol. 2020, 11, 611789. [Google Scholar] [CrossRef]
- Sakakura, T. Serendipity; Close Encounter of Tenascin C. Front. Immunol. 2020, 11, 620182. [Google Scholar]
- Bornstein, P. Matricellular proteins: An overview. J. Cell Commun. Signal. 2009, 3, 163–165. [Google Scholar] [CrossRef] [Green Version]
- Bornstein, P.; Sage, E.H. Matricellular proteins: Extracellular modulators of cell function. Curr. Opin. Cell Biol. 2002, 14, 608–616. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K. Tenascin-C in cardiovascular tissue remodeling: From development to inflammation and repair. Circ. J. 2012, 76, 2513–2520. [Google Scholar] [CrossRef] [Green Version]
- Imanaka-Yoshida, K.; Tawara, I.; Yoshida, T. Tenascin-C in cardiac disease: A sophisticated controller of inflammation, repair, and fibrosis. Am. J. Physiol. Cell Physiol. 2020, 319, C781–C796. [Google Scholar] [CrossRef] [PubMed]
- Deligne, C.; Murdamoothoo, D.; Gammage, A.N.; Gschwandtner, M.; Erne, W.; Loustau, T.; Marzeda, A.M.; Carapito, R.; Paul, N.; Velazquez-Quesada, I.; et al. Matrix-Targeting Immunotherapy Controls Tumor Growth and Spread by Switching Macrophage Phenotype. Cancer Immunol. Res. 2020, 8, 368–382. [Google Scholar] [CrossRef]
- Spenle, C.; Loustau, T.; Murdamoothoo, D.; Erne, W.; Beghelli-de la Forest Divonne, S.; Veber, R.; Petti, L.; Bourdely, P.; Morgelin, M.; Brauchle, E.M.; et al. Tenascin-C Orchestrates an Immune-Suppressive Tumor Microenvironment in Oral Squamous Cell Carcinoma. Cancer Immunol. Res. 2020, 8, 1122–1138. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Ospelt, C.; Gay, S.; Midwood, K.S. Location, location, location: How the tissue microenvironment affects inflammation in RA. Nat. Rev. Rheumatol. 2021, 17, 195–212. [Google Scholar] [CrossRef]
- Deligne, C.; Midwood, K.S. Macrophages and Extracellular Matrix in Breast Cancer: Partners in Crime or Protective Allies? Front. Oncol. 2021, 11, 620773. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K.; Matsumoto, K.; Hara, M.; Sakakura, T.; Yoshida, T. The dynamic expression of tenascin-C and tenascin-X during early heart development in the mouse. Differentiation 2003, 71, 291–298. [Google Scholar] [CrossRef]
- Yonebayashi, S.; Tajiri, K.; Hara, M.; Saito, H.; Suzuki, N.; Sakai, S.; Kimura, T.; Sato, A.; Sekimoto, A.; Fujita, S.; et al. Generation of Transgenic Mice that Conditionally Overexpress Tenascin-C. Front. Immunol. 2021, 12, 620541. [Google Scholar] [CrossRef]
- Monda, E.; Palmiero, G.; Rubino, M.; Verrillo, F.; Amodio, F.; Di Fraia, F.; Pacileo, R.; Fimiani, F.; Esposito, A.; Cirillo, A.; et al. Molecular Basis of Inflammation in the Pathogenesis of Cardiomyopathies. Int. J. Mol. Sci. 2020, 21, 6462. [Google Scholar] [CrossRef]
- Murphy, S.P.; Kakkar, R.; McCarthy, C.P.; Januzzi, J.L., Jr. Inflammation in Heart Failure: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 75, 1324–1340. [Google Scholar] [CrossRef]
- Levine, B.; Kalman, J.; Mayer, L.; Fillit, H.M.; Packer, M. Elevated Circulating Levels of Tumor Necrosis Factor in Severe Chronic Heart Failure. N. Engl. J. Med. 1990, 323, 236–241. [Google Scholar] [CrossRef]
- Podesser, B.K.; Kreibich, M.; Dzilic, E.; Santer, D.; Forster, L.; Trojanek, S.; Abraham, D.; Krssak, M.; Klein, K.U.; Tretter, E.V.; et al. Tenascin-C promotes chronic pressure overload-induced cardiac dysfunction, hypertrophy and myocardial fibrosis. J. Hypertens 2018, 36, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, I.F.; Acar, E.; Costantino, S.; Szabo, P.L.; Hamza, O.; Tretter, E.V.; Klein, K.U.; Trojanek, S.; Abraham, D.; Paneni, F.; et al. Epigenetic modulation of tenascin C in the heart: Implications on myocardial ischemia, hypertrophy and metabolism. J. Hypertens 2019, 37, 1861–1870. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K.; Hiroe, M.; Nishikawa, T.; Ishiyama, S.; Shimojo, T.; Ohta, Y.; Sakakura, T.; Yoshida, T. Tenascin-C modulates adhesion of cardiomyocytes to extracellular matrix during tissue remodeling after myocardial infarction. Lab. Investig. 2001, 81, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Imanaka-Yoshida, K.; Hiroe, M.; Yasutomi, Y.; Toyozaki, T.; Tsuchiya, T.; Noda, N.; Maki, T.; Nishikawa, T.; Sakakura, T.; Yoshida, T. Tenascin-C is a useful marker for disease activity in myocarditis. J. Pathol. 2002, 197, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Imanaka-Yoshida, K.; Hiramitsu, S.; Kato, S.; Ohtsuki, M.; Uemura, A.; Kato, Y.; Nishikawa, T.; Toyozaki, T.; Hishida, H.; et al. Diagnostic utility of tenascin-C for evaluation of the activity of human acute myocarditis. J. Pathol. 2005, 205, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Suzuki, M.; Onishi, K.; Takakura, N.; Inada, H.; Yoshida, T.; Hiroe, M.; Imanaka-Yoshida, K. Eplerenone attenuates myocardial fibrosis in the angiotensin II-induced hypertensive mouse: Involvement of tenascin-C induced by aldosterone-mediated inflammation. J. Cardiovasc. Pharmacol. 2007, 49, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Shimojo, N.; Terasaki, F.; Otsuka, K.; Hosotani, N.; Kohda, Y.; Tanaka, T.; Nishioka, T.; Yoshida, T.; Hiroe, M.; et al. Atrial natriuretic peptide exerts protective action against angiotensin II-induced cardiac remodeling by attenuating inflammation via endothelin-1/endothelin receptor A cascade. Heart Vessel. 2013, 28, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Imanaka-Yoshida, K.; Hiroe, M.; Yoshida, T. Interaction between cell and extracellular matrix in heart disease: Multiple roles of tenascin-C in tissue remodeling. Histol. Histopathol. 2004, 19, 517–525. [Google Scholar] [PubMed]
- Giblin, S.P.; Midwood, K.S. Tenascin-C: Form versus function. Cell Adh. Migr. 2015, 9, 48–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [Green Version]
- Okada, T.; Suzuki, H. The Role of Tenascin-C in Tissue Injury and Repair After Stroke. Front. Immunol. 2020, 11, 607587. [Google Scholar] [CrossRef]
- Nakajima, Y.; Imanaka-Yoshida, K. New insights into the developmental mechanisms of coronary vessels and epicardium. Int. Rev. Cell Mol. Biol. 2013, 303, 263–317. [Google Scholar] [PubMed]
- Cao, Y.X.; Duca, S.; Cao, J.L. Epicardium in Heart Development. Cold Spring Harb. Perspect. Biol. 2020, 12, a037143. [Google Scholar] [CrossRef]
- Vrancken Peeters, M.P.; Gittenberger-de Groot, A.C.; Mentink, M.M.; Hungerford, J.E.; Little, C.D.; Poelmann, R.E. Differences in development of coronary arteries and veins. Cardiovasc. Res. 1997, 36, 101–110. [Google Scholar] [CrossRef]
- Vrancken Peeters, M.P.; Gittenberger-de Groot, A.C.; Mentink, M.M.; Hungerford, J.E.; Little, C.D.; Poelmann, R.E. The development of the coronary vessels and their differentiation into arteries and veins in the embryonic quail heart. Dev. Dyn. 1997, 208, 338–348. [Google Scholar] [CrossRef]
- Kattan, J.; Dettman, R.W.; Bristow, J. Formation and remodeling of the coronary vascular bed in the embryonic avian heart. Dev. Dyn. 2004, 230, 34–43. [Google Scholar] [CrossRef]
- Ando, K.; Takahashi, M.; Yamagishi, T.; Miyagawa-Tomita, S.; Imanaka-Yoshida, K.; Yoshida, T.; Nakajima, Y. Tenascin C may regulate the recruitment of smooth muscle cells during coronary artery development. Differentiation 2011, 81, 299–306. [Google Scholar] [CrossRef]
- Ishigaki, T.; Imanaka-Yoshida, K.; Shimojo, N.; Matsushima, S.; Taki, W.; Yoshida, T. Tenascin-C enhances crosstalk signaling of integrin alphavbeta3/PDGFR-beta complex by SRC recruitment promoting PDGF-induced proliferation and migration in smooth muscle cells. J. Cell Physiol. 2011, 226, 2617–2624. [Google Scholar] [CrossRef] [PubMed]
- Imanaka-Yoshida, K.; Yoshida, T.; Miyagawa-Tomita, S. Tenascin-C in development and disease of blood vessels. Anat. Rec. 2014, 297, 1747–1757. [Google Scholar] [CrossRef]
- Forsberg, E.; Hirsch, E.; Fröhlich, L.; Meyer, M.; Ekblom, P.; Aszodi, A.; Werner, S.; Fässler, R. Skin wounds and severed nerves heal normally in mice lacking tenascin-C. Proc. Natl. Acad. Sci. USA 1996, 93, 6594–6599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saga, Y.; Yagi, T.; Ikawa, Y.; Sakakura, T.; Aizawa, S. Mice develop normally without tenascin. Genes Dev. 1992, 6, 1821–1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, K.; Imanaka-Yoshida, K.; Yoshida, T.; Sugimura, Y. Role of stromal tenascin-C in mouse prostatic development and epithelial cell differentiation. Dev. Biol. 2008, 324, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Roth-Kleiner, M.; Hirsch, E.; Schittny, J.C. Fetal lungs of tenascin-C-deficient mice grow well, but branch poorly in organ culture. Am. J. Respir. Cell Mol. Biol. 2004, 30, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mund, S.I.; Schittny, J.C. Tenascin-C deficiency impairs alveolarization and microvascular maturation during postnatal lung development. J. Appl. Physiol. 2020, 128, 1287–1298. [Google Scholar] [CrossRef]
- Morellini, F.; Schachner, M. Enhanced novelty-induced activity, reduced anxiety, delayed resynchronization to daylight reversal and weaker muscle strength in tenascin-C-deficient mice. Eur. J. Neurosci. 2006, 23, 1255–1268. [Google Scholar] [CrossRef] [PubMed]
- De Chevigny, A.; Lemasson, M.; Saghatelyan, A.; Sibbe, M.; Schachner, M.; Lledo, P.M. Delayed onset of odor detection in neonatal mice lacking tenascin-C. Mol. Cell. Neurosci. 2006, 32, 174–186. [Google Scholar] [CrossRef]
- Gurevicius, K.; Kuang, F.; Stoenica, L.; Irintchev, A.; Gureviciene, I.; Dityatev, A.; Schachner, M.; Tanila, H. Genetic ablation of tenascin-C expression leads to abnormal hippocampal CA1 structure and electrical activity in vivo. Hippocampus 2009, 19, 1232–1246. [Google Scholar] [CrossRef]
- Gremlich, S.; Roth-Kleiner, M.; Equey, L.; Fytianos, K.; Schittny, J.C.; Cremona, T.P. Tenascin-C inactivation impacts lung structure and function beyond lung development. Sci. Rep. 2020, 10, 5118. [Google Scholar] [CrossRef] [Green Version]
- Santer, D.; Nagel, F.; Gonçalves, I.F.; Kaun, C.; Wojta, J.; Fagyas, M.; Krššák, M.; Balogh, Á.; Papp, Z.; Tóth, A.; et al. Tenascin-C aggravates ventricular dilatation and angiotensin-converting enzyme activity after myocardial infarction in mice. ESC Heart Fail. 2020, 7, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Perera-Gonzalez, M.; Kiss, A.; Kaiser, P.; Holzweber, M.; Nagel, F.; Watzinger, S.; Acar, E.; Szabo, P.L.; Gonçalves, I.F.; Weber, L.; et al. The Role of Tenascin C in Cardiac Reverse Remodeling Following Banding-Debanding of the Ascending Aorta. Int. J. Mol. Sci. 2021, 22, 2023. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, L.; Li, F.Q.; Yukht, A.; Qin, M.H.; Ruther, H.; Yang, M.J.; Chaux, A.; Shah, P.K.; Sharifi, B.G. Bone Marrow-Derived Tenascin-C Attenuates Cardiac Hypertrophy by Controlling Inflammation. J. Am. Coll. Cardiol. 2017, 70, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Onishi, K.; Shimojo, N.; Nagano, Y.; Matsusaka, H.; Ikeuchi, M.; Ide, T.; Tsutsui, H.; Hiroe, M.; Yoshida, T.; et al. Tenascin-C may aggravate left ventricular remodeling and function after myocardial infarction in mice. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1072–H1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machino-Ohtsuka, T.; Tajiri, K.; Kimura, T.; Sakai, S.; Sato, A.; Yoshida, T.; Hiroe, M.; Yasutomi, Y.; Aonuma, K.; Imanaka-Yoshida, K. Tenascin-C aggravates autoimmune myocarditis via dendritic cell activation and Th17 cell differentiation. J. Am. Heart Assoc. 2014, 3, e001052. [Google Scholar] [CrossRef] [Green Version]
- Shimojo, N.; Hashizume, R.; Kanayama, K.; Hara, M.; Suzuki, Y.; Nishioka, T.; Hiroe, M.; Yoshida, T.; Imanaka-Yoshida, K. Tenascin-C May Accelerate Cardiac Fibrosis by Activating Macrophages via the Integrin alphaVbeta3/Nuclear Factor-kappaB/Interleukin-6 Axis. Hypertension 2015, 66, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Tajiri, K.; Sato, A.; Sakai, S.; Wang, Z.; Yoshida, T.; Uede, T.; Hiroe, M.; Aonuma, K.; Ieda, M.; et al. Tenascin-C accelerates adverse ventricular remodelling after myocardial infarction by modulating macrophage polarization. Cardiovasc. Res. 2019, 115, 614–624. [Google Scholar] [CrossRef]
- Nagel, F.; Santer, D.; Stojkovic, S.; Kaun, C.; Schaefer, A.K.; Krššák, M.; Abraham, D.; Bencsik, P.; Ferdinandy, P.; Kenyeres, E.; et al. The impact of age on cardiac function and extracellular matrix component expression in adverse post-infarction remodeling in mice. Exp. Gerontol. 2019, 119, 193–202. [Google Scholar] [CrossRef]
- Sato, A.; Aonuma, K.; Imanaka-Yoshida, K.; Yoshida, T.; Isobe, M.; Kawase, D.; Kinoshita, N.; Yazaki, Y.; Hiroe, M. Serum tenascin-C might be a novel predictor of left ventricular remodeling and prognosis after acute myocardial infarction. J. Am. Coll. Cardiol. 2006, 47, 2319–2325. [Google Scholar] [CrossRef] [Green Version]
- Willems, I.E.; Arends, J.W.; Daemen, M.J. Tenascin and fibronectin expression in healing human myocardial scars. J. Pathol. 1996, 179, 321–325. [Google Scholar] [CrossRef]
- Tamaoki, M.; Imanaka-Yoshida, K.; Yokoyama, K.; Nishioka, T.; Inada, H.; Hiroe, M.; Sakakura, T.; Yoshida, T. Tenascin-C regulates recruitment of myofibroblasts during tissue repair after myocardial injury. Am. J. Pathol. 2005, 167, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Murphy-Ullrich, J.E.; Sage, E.H. Revisiting the matricellular concept. Matrix Biol. 2014, 37, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Danowski, B.A.; Imanaka-Yoshida, K.; Sanger, J.M.; Sanger, J.W. Costameres are sites of force transmission to the substratum in adult rat cardiomyocytes. J. Cell Biol. 1992, 118, 1411–1420. [Google Scholar] [CrossRef] [Green Version]
- Imanaka-Yoshida, K.; Danowski, B.A.; Sanger, J.M.; Sanger, J.W. Living adult rat cardiomyocytes in culture: Evidence for dissociation of costameric distribution of vinculin from costameric distributions of attachments. Cell Motil. Cytoskelet. 1996, 33, 263–275. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K.; Enomoto-Iwamoto, M.; Yoshida, T.; Sakakura, T. Vinculin, Talin, Integrin alpha6beta1 and laminin can serve as components of attachment complex mediating contraction force transmission from cardiomyocytes to extracellular matrix. Cell Motil. Cytoskelet. 1999, 42, 1–11. [Google Scholar] [CrossRef]
- Mazhari, R.; Omens, J.H.; Covell, J.W.; McCulloch, A.D. Structural basis of regional dysfunction in acutely ischemic myocardium. Cardiovasc. Res. 2000, 47, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Oberhauser, A.F.; Marszalek, P.E.; Erickson, H.P.; Fernandez, J.M. The molecular elasticity of the extracellular matrix protein tenascin. Nature 1998, 393, 181–185. [Google Scholar] [CrossRef]
- Marín, J.L.; Muñiz, J.; Huerta, M.; Trujillo, X. Folding-unfolding of FN-III domains in tenascin: An elastically coupled two-state system. J. Biomech. 2003, 36, 1733–1737. [Google Scholar] [CrossRef]
- Kimura, T.; Shiraishi, K.; Furusho, A.; Ito, S.; Hirakata, S.; Nishida, N.; Yoshimura, K.; Imanaka-Yoshida, K.; Yoshida, T.; Ikeda, Y.; et al. Tenascin C protects aorta from acute dissection in mice. Sci. Rep. 2014, 4, 4051. [Google Scholar] [CrossRef]
- De Haan, J.J.; Smeets, M.B.; Pasterkamp, G.; Arslan, F. Danger signals in the initiation of the inflammatory response after myocardial infarction. Mediat. Inflamm. 2013, 2013, 206039. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.A. Inflammatory and fibrotic responses of cardiac fibroblasts to myocardial damage associated molecular patterns (DAMPs). J. Mol. Cell Cardiol. 2016, 94, 189–200. [Google Scholar] [CrossRef]
- Marzeda, A.M.; Midwood, K.S. Internal Affairs: Tenascin-C as a Clinically Relevant, Endogenous Driver of Innate Immunity. J. Histochem. Cytochem. 2018, 66, 289–304. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G. The immune system and cardiac repair. Pharmacol. Res. 2008, 58, 88–111. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, S.D.; Frangogiannis, N.G. The Biological Basis for Cardiac Repair After Myocardial Infarction: From Inflammation to Fibrosis. Circ. Res. 2016, 119, 91–112. [Google Scholar] [CrossRef]
- Midwood, K.; Sacre, S.; Piccinini, A.M.; Inglis, J.; Trebaul, A.; Chan, E.; Drexler, S.; Sofat, N.; Kashiwagi, M.; Orend, G.; et al. Tenascin-C is an endogenous activator of Toll-like receptor 4 that is essential for maintaining inflammation in arthritic joint disease. Nat. Med. 2009, 15, 774–780. [Google Scholar] [CrossRef]
- Zuliani-Alvarez, L.; Marzeda, A.M.; Deligne, C.; Schwenzer, A.; McCann, F.E.; Marsden, B.D.; Piccinini, A.M.; Midwood, K.S. Mapping tenascin-C interaction with toll-like receptor 4 reveals a new subset of endogenous inflammatory triggers. Nat. Commun. 2017, 8, 1595. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, A.; Spary, E.J.; Manfield, I.W.; Ruhmann, M.; Zuliani-Alvarez, L.; Gamboa-Esteves, F.O.; Porter, K.E.; Drinkhill, M.J.; Midwood, K.S.; Turner, N.A. Tenascin C upregulates interleukin-6 expression in human cardiac myofibroblasts via toll-like receptor 4. World J. Cardiol. 2016, 8, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Piccinini, A.M.; Zuliani-Alvarez, L.; Lim, J.M.P.; Midwood, K.S. Distinct microenvironmental cues stimulate divergent TLR4-mediated signaling pathways in macrophages. Sci. Signal. 2016, 9, ra86. [Google Scholar] [CrossRef] [Green Version]
- Benbow, J.H.; Thompson, K.J.; Cope, H.L.; Brandon-Warner, E.; Culberson, C.R.; Bossi, K.L.; Li, T.; Russo, M.W.; Gersin, K.S.; McKillop, I.H.; et al. Diet-Induced Obesity Enhances Progression of Hepatocellular Carcinoma through Tenascin-C/Toll-Like Receptor 4 Signaling. Am. J. Pathol. 2016, 186, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Kuriyama, N.; Duarte, S.; Hamada, T.; Busuttil, R.W.; Coito, A.J. Tenascin-C: A novel mediator of hepatic ischemia and reperfusion injury. Hepatology 2011, 54, 2125–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nieuwenhoven, F.A.; Turner, N.A. The role of cardiac fibroblasts in the transition from inflammation to fibrosis following myocardial infarction. Vascul. Pharmacol. 2013, 58, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honold, L.; Nahrendorf, M. Resident and Monocyte-Derived Macrophages in Cardiovascular Disease. Circ. Res. 2018, 122, 113–127. [Google Scholar] [CrossRef]
- Chen, B.; Frangogiannis, N.G. The Role of Macrophages in Nonischemic Heart Failure. JACC Basic Transl. Sci. 2018, 3, 245–248. [Google Scholar] [CrossRef]
- De Couto, G.; Liu, W.; Tseliou, E.; Sun, B.; Makkar, N.; Kanazawa, H.; Arditi, M.; Marbán, E. Macrophages mediate cardioprotective cellular postconditioning in acute myocardial infarction. J. Clin. Investig. 2015, 125, 3147–3162. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, S.A.; Dunne, A.; Monaghan, M.G. The Role of Macrophages in the Infarcted Myocardium: Orchestrators of ECM Remodeling. Front. Cardiovasc. Med. 2019, 6, 101. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Mouton, A.J.; Lindsey, M.L. Cardiac macrophage biology in the steady-state heart, the aging heart, and following myocardial infarction. Transl. Res. 2018, 191, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzai, T. Inflammatory Mechanisms of Cardiovascular Remodeling. Circ. J. 2018, 82, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.Y.; Lee, K.W.; Shin, D.; An, S.; Cho, K.H.; Kim, S.H. A positive feedback loop bi-stably activates fibroblasts. Nat. Commun. 2018, 9, 3016. [Google Scholar] [CrossRef] [PubMed]
- Hesse, J.; Leberling, S.; Boden, E.; Friebe, D.; Schmidt, T.; Ding, Z.P.; Dieterich, P.; Deussen, A.; Roderigo, C.; Rose, C.R.; et al. CD73-derived adenosine and tenascin-C control cytokine production by epicardium-derived cells formed after myocardial infarction. FASEB J. 2017, 31, 3040–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, S.; Midwood, K.S.; Yin, H.; Varga, J. Toll-Like Receptor-4 Signaling Drives Persistent Fibroblast Activation and Prevents Fibrosis Resolution in Scleroderma. Adv. Wound Care 2017, 6, 356–369. [Google Scholar] [CrossRef] [Green Version]
- Frangogiannis, N.G. The extracellular matrix in myocardial injury, repair, and remodeling. J. Clin. Investig. 2017, 127, 1600–1612. [Google Scholar] [CrossRef] [Green Version]
- Midwood, K.S.; Hussenet, T.; Langlois, B.; Orend, G. Advances in tenascin-C biology. Cell Mol. Life Sci. 2011, 68, 3175–3199. [Google Scholar] [CrossRef] [Green Version]
- Katoh, D.; Kozuka, Y.; Noro, A.; Ogawa, T.; Imanaka-Yoshida, K.; Yoshida, T. Tenascin-C Induces Phenotypic Changes in Fibroblasts to Myofibroblasts with High Contractility through the Integrin αvβ1/Transforming Growth Factor β/SMAD Signaling Axis in Human Breast Cancer. Am. J. Pathol. 2020, 190, 2123–2135. [Google Scholar] [CrossRef]
- Sato, A.; Hiroe, M.; Akiyama, D.; Hikita, H.; Nozato, T.; Hoshi, T.; Kimura, T.; Wang, Z.; Sakai, S.; Imanaka-Yoshida, K.; et al. Prognostic value of serum tenascin-C levels on long-term outcome after acute myocardial infarction. J. Card. Fail. 2012, 18, 480–486. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Pathophysiology of Myocardial Infarction. Compr. Physiol. 2015, 5, 1841–1875. [Google Scholar] [PubMed]
- Lindsey, M.L.; Iyer, R.P.; Jung, M.; DeLeon-Pennell, K.Y.; Ma, Y. Matrix metalloproteinases as input and output signals for post-myocardial infarction remodeling. J. Mol. Cell Cardiol. 2016, 91, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhao, Q.; Kong, W. Extracellular matrix remodeling and cardiac fibrosis. Matrix Biol. 2018, 68–69, 490–506. [Google Scholar] [CrossRef]
- Kong, P.; Christia, P.; Frangogiannis, N.G. The pathogenesis of cardiac fibrosis. Cell Mol. Life Sci. 2014, 71, 549–574. [Google Scholar] [CrossRef] [Green Version]
- Regan, T.J.; Lyons, M.M.; Ahmed, S.S.; Levinson, G.E.; Oldewurtel, H.A.; Ahmad, M.R.; Haider, B. Evidence for cardiomyopathy in familial diabetes mellitus. J. Clin. Investig. 1977, 60, 884–899. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.Y.; López, B.; Coelho-Filho, O.R.; Lakdawala, N.K.; Cirino, A.L.; Jarolim, P.; Kwong, R.; González, A.; Colan, S.D.; Seidman, J.G.; et al. Myocardial fibrosis as an early manifestation of hypertrophic cardiomyopathy. N. Engl. J. Med. 2010, 363, 552–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krejci, J.; Mlejnek, D.; Sochorova, D.; Nemec, P. Inflammatory Cardiomyopathy: A Current View on the Pathophysiology, Diagnosis, and Treatment. Biomed. Res. Int. 2016, 2016, 4087632. [Google Scholar] [CrossRef] [Green Version]
- Assomull, R.G.; Prasad, S.K.; Lyne, J.; Smith, G.; Burman, E.D.; Khan, M.; Sheppard, M.N.; Poole-Wilson, P.A.; Pennell, D.J. Cardiovascular magnetic resonance, fibrosis, and prognosis in dilated cardiomyopathy. J. Am. Coll. Cardiol. 2006, 48, 1977–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenning, G.; Zeisberg, E.M.; Kalluri, R. The origin of fibroblasts and mechanism of cardiac fibrosis. J. Cell Physiol. 2010, 225, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Manabe, I. Chronic Inflammation Links Cardiovascular, Metabolic and Renal Diseases. Circ. J. 2011, 75, 2739–2748. [Google Scholar] [CrossRef] [Green Version]
- Oishi, Y.; Manabe, I. Macrophages in age-related chronic inflammatory diseases. NPJ Aging Mech. Dis. 2016, 2, 16018. [Google Scholar] [CrossRef] [Green Version]
- Van Linthout, S.; Elsanhoury, A.; Klein, O.; Sosnowski, M.; Miteva, K.; Lassner, D.; Abou-El-Enein, M.; Pieske, B.; Kuhl, U.; Tschope, C. Telbivudine in chronic lymphocytic myocarditis and human parvovirus B19 transcriptional activity. ESC Heart Fail. 2018, 5, 818–829. [Google Scholar] [CrossRef]
- Hsieh, C.S.; Macatonia, S.E.; O’Garra, A.; Murphy, K.M. T cell genetic background determines default T helper phenotype development in vitro. J. Exp. Med. 1995, 181, 713–721. [Google Scholar] [CrossRef]
- Abbadi, D.; Laroumanie, F.; Bizou, M.; Pozzo, J.; Daviaud, D.; Delage, C.; Calise, D.; Gaits-Iacovoni, F.; Dutaur, M.; Tortosa, F.; et al. Local production of tenascin-C acts as a trigger formonocyte/macrophage recruitment that provokes cardiac dysfunction. Cardiovasc. Res. 2018, 114, 123–137. [Google Scholar] [CrossRef] [PubMed]
- JCS Joint Working Group and treatment of myocarditis (JCS 2009): Digest version. Circ. J. 2011, 75, 734–743. [CrossRef] [Green Version]
- Imanaka-Yoshida, K. Inflammation in myocardial disease: From myocarditis to dilated cardiomyopathy. Pathol. Int. 2020, 70, 1–11. [Google Scholar] [CrossRef]
- Kawada, J.I.; Takeuchi, S.; Imai, H.; Okumura, T.; Horiba, K.; Suzuki, T.; Torii, Y.; Yasuda, K.; Imanaka-Yoshida, K.; Ito, Y. Immune cell infiltration landscapes in pediatric acute myocarditis analyzed by CIBERSORT. J. Cardiol. 2021, 77, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Ammirati, E.; Veronese, G.; Bottiroli, M.; Wang, D.W.; Cipriani, M.; Garascia, A.; Pedrotti, P.; Adler, E.D.; Frigerio, M. Update on acute myocarditis. Trends Cardiovasc. Med. 2020, S1050-1738, 30077–30079. [Google Scholar] [CrossRef]
- Suzuki, H.; Nishikawa, H.; Kawakita, F. Matricellular proteins as possible biomarkers for early brain injury after aneurysmal subarachnoid hemorrhage. Neural Regen Res. 2018, 13, 1175–1178. [Google Scholar] [CrossRef]
- Tajiri, K.; Aonuma, K.; Sekine, I. Immune checkpoint inhibitor-related myocarditis. Jpn J. Clin. Oncol. 2018, 48, 7–12. [Google Scholar] [CrossRef]
- Trachtenberg, B.H.; Hare, J.M. Inflammatory Cardiomyopathic Syndromes. Circ. Res. 2017, 121, 803–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschope, C.; Ammirati, E.; Bozkurt, B.; Caforio, A.L.P.; Cooper, L.T.; Felix, S.B.; Hare, J.M.; Heidecker, B.; Heymans, S.; Hubner, N.; et al. Myocarditis and inflammatory cardiomyopathy: Current evidence and future directions. Nat. Rev. Cardiol. 2021, 18, 169–193. [Google Scholar] [CrossRef]
- Basso, C.; Leone, O.; Rizzo, S.; De Gaspari, M.; van der Wal, A.C.; Aubry, M.-C.; Bois, M.C.; Lin, P.T.; Maleszewski, J.J.; Stone, J.R. Pathological features of COVID-19-associated myocardial injury: A multicentre cardiovascular pathology study. Eur. Heart J. 2020, 41, 3827–3835. [Google Scholar] [CrossRef] [PubMed]
- Halushka, M.K.; Vander Heide, R.S. Myocarditis is rare in COVID-19 autopsies: Cardiovascular findings across 277 postmortem examinations. Cardiovasc. Pathol. 2021, 50, 107300. [Google Scholar] [CrossRef] [PubMed]
- Kawai, C. From myocarditis to cardiomyopathy: Mechanisms of inflammation and cell death: Learning from the past for the future. Circulation 1999, 99, 1091–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodruff, J.F.; Woodruff, J.J. Involvement of T lymphocytes in the pathogenesis of coxsackie virus B3 heart disease. J. Immunol. 1974, 113, 1726–1734. [Google Scholar]
- Lasrado, N.; Reddy, J. An overview of the immune mechanisms of viral myocarditis. Rev. Med. Virol. 2020, 30, 1–14. [Google Scholar] [CrossRef]
- Gangaplara, A.; Massilamany, C.; Brown, D.M.; Delhon, G.; Pattnaik, A.K.; Chapman, N.; Rose, N.; Steffen, D.; Reddy, J. Coxsackievirus B3 infection leads to the generation of cardiac myosin heavy chain-alpha-reactive CD4 T cells in A/J mice. Clin. Immunol. 2012, 144, 237–249. [Google Scholar] [CrossRef] [Green Version]
- Basavalingappa, R.H.; Arumugam, R.; Lasrado, N.; Yalaka, B.; Massilamany, C.; Gangaplara, A.; Riethoven, J.J.; Xiang, S.H.; Steffen, D.; Reddy, J. Viral myocarditis involves the generation of autoreactive T cells with multiple antigen specificities that localize in lymphoid and non-lymphoid organs in the mouse model of CVB3 infection. Mol. Immunol. 2020, 124, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Havari, E.; Pinto, S.; Gottumukkala, R.V.; Cornivelli, L.; Raddassi, K.; Matsui, T.; Rosenzweig, A.; Bronson, R.T.; Smith, R.; et al. Impaired thymic tolerance to alpha-myosin directs autoimmunity to the heart in mice and humans. J. Clin. Investig. 2011, 121, 1561–1573. [Google Scholar] [CrossRef]
- Eriksson, U.; Ricci, R.; Hunziker, L.; Kurrer, M.O.; Oudit, G.Y.; Watts, T.H.; Sonderegger, I.; Bachmaier, K.; Kopf, M.; Penninger, J.M. Dendritic cell-induced autoimmune heart failure requires cooperation between adaptive and innate immunity. Nat. Med. 2003, 9, 1484–1490. [Google Scholar] [CrossRef]
- Tajiri, K.; Imanaka-Yoshida, K.; Matsubara, A.; Tsujimura, Y.; Hiroe, M.; Naka, T.; Shimojo, N.; Sakai, S.; Aonuma, K.; Yasutomi, Y. Suppressor of cytokine signaling 1 DNA administration inhibits inflammatory and pathogenic responses in autoimmune myocarditis. J. Immunol. 2012, 189, 2043–2053. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.T.; Schwenzer, A.; Marsh, E.K.; Edwards, M.R.; Sabroe, I.; Midwood, K.S.; Parker, L.C. Airway Epithelial Cells Generate Pro-inflammatory Tenascin-C and Small Extracellular Vesicles in Response to TLR3 Stimuli and Rhinovirus Infection. Front. Immunol. 2019, 10, 1987. [Google Scholar] [CrossRef] [Green Version]
- Fouda, G.G.; Jaeger, F.H.; Amos, J.D.; Ho, C.; Kunz, E.L.; Anasti, K.; Stamper, L.W.; Liebl, B.E.; Barbas, K.H.; Ohashi, T.; et al. Tenascin-C is an innate broad-spectrum, HIV-1–neutralizing protein in breast milk. Proc. Natl. Acad. Sci. USA 2013, 110, 18220. [Google Scholar] [CrossRef] [Green Version]
- Mansour, R.G.; Stamper, L.; Jaeger, F.; McGuire, E.; Fouda, G.; Amos, J.; Barbas, K.; Ohashi, T.; Alam, S.M.; Erickson, H.; et al. The Presence and Anti-HIV-1 Function of Tenascin C in Breast Milk and Genital Fluids. PLoS ONE 2016, 11, e0155261. [Google Scholar] [CrossRef]
- Mangan, R.J.; Stamper, L.; Ohashi, T.; Eudailey, J.A.; Go, E.P.; Jaeger, F.H.; Itell, H.L.; Watts, B.E.; Fouda, G.G.; Erickson, H.P.; et al. Determinants of Tenascin-C and HIV-1 envelope binding and neutralization. Mucosal. Immunol. 2019, 12, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Khatun, M.; Steele, R.; Isbell, T.S.; Ray, R.; Ray, R.B. Exosomes from COVID-19 Patients Carry Tenascin-C and Fibrinogen-β in Triggering Inflammatory Signals in Cells of Distant Organ. Int. J. Mol. Sci. 2021, 22, 3184. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A.L.; Faal, H.; Moss, D.; Addengast, L.; Fanta, L.; Eyster, K.; Huber, V.C.; Chaussee, M.S. The Streptococcus pyogenes fibronectin/tenascin-binding protein PrtF.2 contributes to virulence in an influenza superinfection. Sci. Rep. 2018, 8, 12126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leipner, C.; Grün, K.; Müller, A.; Buchdunger, E.; Borsi, L.; Kosmehl, H.; Berndt, A.; Janik, T.; Uecker, A.; Kiehntopf, M.; et al. Imatinib mesylate attenuates fibrosis in coxsackievirus b3-induced chronic myocarditis. Cardiovasc. Res. 2008, 79, 118–126. [Google Scholar] [CrossRef]
- Sato, T.; Nakamori, S.; Watanabe, S.; Nishikawa, K.; Inoue, T.; Imanaka-Yoshida, K.; Ishida, M.; Sakuma, H.; Ito, M.; Dohi, K. Monitoring of the Evolution of Immune Checkpoint Inhibitor Myocarditis With Cardiovascular Magnetic Resonance. Circ. Cardiovasc. Imaging 2020, 13, e010633. [Google Scholar] [CrossRef]
- Tajiri, K.; Yonebayashi, S.; Li, S.; Ieda, M. Immunomodulatory Role of Tenascin-C in Myocarditis and Inflammatory Cardiomyopathy. Front. Immunol. 2021, 12, 624703. [Google Scholar] [CrossRef]
- Cihakova, D.; Rose, N.R. Pathogenesis of Myocarditis and Dilated Cardiomyopathy. In Advances in Immunology; Academic Press: Cambridge, MA, USA, 2008; Volume 99, pp. 95–114. [Google Scholar]
- Sonderegger, I.; Röhn, T.A.; Kurrer, M.O.; Iezzi, G.; Zou, Y.; Kastelein, R.A.; Bachmann, M.F.; Kopf, M. Neutralization of IL-17 by active vaccination inhibits IL-23-dependent autoimmune myocarditis. Eur. J. Immunol. 2006, 36, 2849–2856. [Google Scholar] [CrossRef]
- Chiquet-Ehrismann, R.; Orend, G.; Chiquet, M.; Tucker, R.P.; Midwood, K.S. Tenascins in stem cell niches. Matrix Biol. 2014, 37, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.; Erickson, H.P.; Springer, T.A. Tenascin supports lymphocyte rolling. J. Cell Biol. 1997, 137, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Ishizu, A.; Okuno, Y.; Omatsu, Y.; Okabe, K.; Morimoto, J.; Uede, T.; Nagasawa, T.; Suda, T.; Kubota, Y. Extracellular matrix protein tenascin-C is required in the bone marrow microenvironment primed for hematopoietic regeneration. Blood 2012, 119, 5429–5437. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.L.; Heazlewood, S.Y.; Williams, B.; Reitsma, A.J.; Grassinger, J.; Borg, J.; Heazlewood, C.K.; Chidgey, A.P.; Nilsson, S.K. The role of Tenascin C in the lymphoid progenitor cell niche. Exp. Hematol. 2013, 41, 1050–1061. [Google Scholar] [CrossRef]
- Nakahara, H.; Gabazza, E.C.; Fujimoto, H.; Nishii, Y.; D’Alessandro-Gabazza, C.N.; Bruno, N.E.; Takagi, T.; Hayashi, T.; Maruyama, J.; Maruyama, K.; et al. Deficiency of tenascin C attenuates allergen-induced bronchial asthma in the mouse. Eur. J. Immunol. 2006, 36, 3334–3345. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, M.; Kurotaki, D.; Morimoto, J.; Asano, T.; Matsui, Y.; Nakayama, Y.; Saito, Y.; Ito, K.; Kimura, C.; Iwasaki, N.; et al. Alpha9 integrin and its ligands constitute critical joint microenvironments for development of autoimmune arthritis. J. Immunol. 2009, 182, 8015–8025. [Google Scholar] [CrossRef] [Green Version]
- Kanayama, M.; Morimoto, J.; Matsui, Y.; Ikesue, M.; Danzaki, K.; Kurotaki, D.; Ito, K.; Yoshida, T.; Uede, T. alpha9beta1 integrin-mediated signaling serves as an intrinsic regulator of pathogenic Th17 cell generation. J. Immunol. 2011, 187, 5851–5864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhmann, M.; Piccinini, A.M.; Kong, P.L.; Midwood, K.S. Endogenous activation of adaptive immunity: Tenascin-C drives interleukin-17 synthesis in murine arthritic joint disease. Arthritis Rheum 2012, 64, 2179–2190. [Google Scholar] [CrossRef]
- Matsumoto, K.I.; Aoki, H. The Roles of Tenascins in Cardiovascular, Inflammatory, and Heritable Connective Tissue Diseases. Front. Immunol. 2020, 11, 609752. [Google Scholar] [CrossRef]
- Tajiri, K.; Imanaka-Yoshida, K.; Tsujimura, Y.; Matsuo, K.; Hiroe, M.; Aonuma, K.; Ieda, M.; Yasutomi, Y. A New Mouse Model of Chronic Myocarditis Induced by Recombinant Bacille Calmette-Guèrin Expressing a T-Cell Epitope of Cardiac Myosin Heavy Chain-α. Int. J. Mol. Sci. 2021, 22, 794. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, M.; Dohi, K.; Imanaka-Yoshida, K.; Omori, T.; Moriwaki, K.; Nakamori, S.; Yamada, N.; Ito, M. Fulminant Myocarditis With Prolonged Active Lymphocytic Infiltration After Hemodynamic Recovery. Int. Heart J. 2017, 58, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Weintraub, R.G.; Semsarian, C.; Macdonald, P. Dilated cardiomyopathy. Lancet 2017, 390, 400–414. [Google Scholar] [CrossRef]
- McNally, E.M.; Mestroni, L. Dilated Cardiomyopathy: Genetic Determinants and Mechanisms. Circ. Res. 2017, 121, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Kindermann, I.; Kindermann, M.; Kandolf, R.; Klingel, K.; Bültmann, B.; Müller, T.; Lindinger, A.; Böhm, M. Predictors of Outcome in Patients With Suspected Myocarditis. Circulation 2008, 118, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Sugano, Y.; Yokokawa, T.; Nagai, T.; Matsuyama, T.-a.; Ohta-Ogo, K.; Ikeda, Y.; Ishibashi-Ueda, H.; Nakatani, T.; Ohte, N.; et al. Clinical impact of the presence of macrophages in endomyocardial biopsies of patients with dilated cardiomyopathy. Eur. J. Heart Fail. 2017, 19, 490–498. [Google Scholar] [CrossRef]
- Yokokawa, T.; Sugano, Y.; Nakayama, T.; Nagai, T.; Matsuyama, T.A.; Ohta-Ogo, K.; Ikeda, Y.; Ishibashi-Ueda, H.; Nakatani, T.; Yasuda, S.; et al. Significance of myocardial tenascin-C expression in left ventricular remodelling and long-term outcome in patients with dilated cardiomyopathy. Eur. J. Heart Fail. 2016, 18, 375–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukada, B.; Terasaki, F.; Shimomura, H.; Otsuka, K.; Otsuka, K.; Katashima, T.; Fujita, S.; Imanaka-Yoshida, K.; Yoshida, T.; Hiroe, M.; et al. High prevalence of chronic myocarditis in dilated cardiomyopathy referred for left ventriculoplasty: Expression of tenascin C as a possible marker for inflammation. Hum. Pathol. 2009, 40, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Odaka, K.; Uehara, T.; Imanaka-Yoshida, K.; Kato, Y.; Oyama, H.; Tadokoro, H.; Akizawa, H.; Tanada, S.; Hiroe, M.; et al. Toward in vivo imaging of heart disease using a radiolabeled single-chain Fv fragment targeting tenascin-C. Anal. Chem. 2011, 83, 9123–9130. [Google Scholar] [CrossRef] [PubMed]
- Dhaouadi, S.; Ben Abderrazek, R.; Loustau, T.; Abou-Faycal, C.; Ksouri, A.; Erne, W.; Murdamoothoo, D.; Mörgelin, M.; Kungl, A.; Jung, A.; et al. Novel Human Tenascin-C Function-Blocking Camel Single Domain Nanobodies. Front. Immunol. 2021, 12, 635166. [Google Scholar] [CrossRef]
- Sato, M.; Toyozaki, T.; Odaka, K.; Uehara, T.; Arano, Y.; Hasegawa, H.; Yoshida, K.; Imanaka-Yoshida, K.; Yoshida, T.; Hiroe, M.; et al. Detection of experimental autoimmune myocarditis in rats by 111In monoclonal antibody specific for tenascin-C. Circulation 2002, 106, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Taki, J.; Inaki, A.; Wakabayashi, H.; Imanaka-Yoshida, K.; Ogawa, K.; Hiroe, M.; Shiba, K.; Yoshida, T.; Kinuya, S. Dynamic expression of tenascin-C after myocardial ischemia and reperfusion: Assessment by 125I-anti-tenascin-C antibody imaging. J. Nucl. Med. 2010, 51, 1116–1122. [Google Scholar] [CrossRef] [Green Version]
- Taki, J.; Inaki, A.; Wakabayashi, H.; Matsunari, I.; Imanaka-Yoshida, K.; Ogawa, K.; Hiroe, M.; Shiba, K.; Yoshida, T.; Kinuya, S. Effect of postconditioning on dynamic expression of tenascin-C and left ventricular remodeling after myocardial ischemia and reperfusion. EJNMMI Res. 2015, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Ageyama, N.; Kurosawa, H.; Fujimoto, O.; Uehara, T.; Hiroe, M.; Arano, Y.; Yoshida, T.; Yasutomi, Y.; Imanaka-Yoshida, K. Successful Inflammation Imaging of Non-Human Primate Hearts Using an Antibody Specific for Tenascin-C. Int. Heart J. 2019, 60, 151–158. [Google Scholar] [CrossRef]
- Terasaki, F.; Okamoto, H.; Onishi, K.; Sato, A.; Shimomura, H.; Tsukada, B.; Imanaka-Yoshida, K.; Hiroe, M.; Yoshida, T.; Kitaura, Y.; et al. Higher serum tenascin-C levels reflect the severity of heart failure, left ventricular dysfunction and remodeling in patients with dilated cardiomyopathy. Circ. J. 2007, 71, 327–330. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, N.; Onishi, K.; Sato, A.; Terasaki, F.; Tsukada, B.; Nozato, T.; Yamada, T.; Imanaka-Yoshida, K.; Yoshida, T.; Ito, M.; et al. Incremental prognostic values of serum tenascin-C levels with blood B-type natriuretic peptide testing at discharge in patients with dilated cardiomyopathy and decompensated heart failure. J. Card Fail. 2009, 15, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Kotby, A.A.; Abdel Aziz, M.M.; El Guindy, W.M.; Moneer, A.N. Can serum tenascin-C be used as a marker of inflammation in patients with dilated cardiomyopathy? Int. J. Pediatr. 2013, 2013, 608563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, M.; Matusiak-Bruckner, M.; Richter, P.; Grun, K.; Ziffels, B.; Neri, D.; Maschek, H.; Schulz, U.; Pfeil, A.; Jung, C.; et al. De novo expression of fetal ED-A(+) fibronectin and B (+) tenascin-C splicing variants in human cardiac allografts: Potential impact for targeted therapy of rejection. J. Mol. Histol. 2014, 45, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Gellen, B.; Thorin-Trescases, N.; Thorin, E.; Gand, E.; Sosner, P.; Brishoual, S.; Rigalleau, V.; Montaigne, D.; Javaugue, V.; Pucheu, Y.; et al. Serum tenascin-C is independently associated with increased major adverse cardiovascular events and death in individuals with type 2 diabetes: A French prospective cohort. Diabetologia 2020, 63, 915–923. [Google Scholar] [CrossRef]
- Kanagala, P.; Arnold, J.R.; Khan, J.N.; Singh, A.; Gulsin, G.S.; Chan, D.C.S.; Cheng, A.S.H.; Yang, J.; Li, Z.; Gupta, P.; et al. Plasma Tenascin-C: A prognostic biomarker in heart failure with preserved ejection fraction. Biomarkers 2020, 25, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Kass, D.A. Cellular and molecular pathobiology of heart failure with preserved ejection fraction. Nat. Rev. Cardiol. 2020. in print. [Google Scholar]
- Nattel, S. Molecular and Cellular Mechanisms of Atrial Fibrosis in Atrial Fibrillation. JACC Clin. Electrophysiol. 2017, 3, 425–435. [Google Scholar] [CrossRef]
- Ozmen, C.; Deniz, A.; Deveci, O.S.; Cagliyan, C.E.; Celik, A.I.; Yildiz, İ.; Yildiz, P.; Demir, M.; Kanadasi, M. Association among tenascin-C and NT-proBNP levels and arrhythmia prevalence in heart failure. Clin. Investig. Med. 2017, 40, E219–E227. [Google Scholar] [CrossRef] [Green Version]
- Shiomi, Y.; Yokokawa, M.; Uzui, H.; Hisazaki, K.; Morishita, T.; Ishida, K.; Fukuoka, Y.; Hasegawa, K.; Ikeda, H.; Tama, N.; et al. Serum tenascin-C levels in atrium predict atrial structural remodeling processes in patients with atrial fibrillation. J. Interv. Card Electrophysiol. 2019, 59, 401–406. [Google Scholar] [CrossRef]
- Golledge, J.; Clancy, P.; Maguire, J.; Lincz, L.; Koblar, S. The role of tenascin C in cardiovascular disease. Cardiovasc. Res. 2011, 92, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Franz, M.; Jung, C.; Lauten, A.; Figulla, H.R.; Berndt, A. Tenascin-C in cardiovascular remodeling: Potential impact for diagnosis, prognosis estimation and targeted therapy. Cell Adh. Migr. 2015, 9, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erer, H.B.; Guvenc, T.S.; Kemik, A.S.; Yilmaz, H.; Kul, S.; Altay, S.; Oz, D.; Zeren, G.; Ekmekci, A.; Zencirci, A.E.; et al. Assessment of tenascin-C levels in ventricular noncompaction/hypertrabeculation patients: A cross-sectional study. Echocardiography 2014, 31, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, H.; Kubo, T.; Baba, Y.; Yamasaki, N.; Matsumura, Y.; Furuno, T.; Doi, Y.L. Serum tenascin-C levels as a prognostic biomarker of heart failure events in patients with hypertrophic cardiomyopathy. J. Cardiol. 2012, 59, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokouchi, Y.; Oharaseki, T.; Enomoto, Y.; Sato, W.; Imanaka-Yoshida, K.; Takahashi, K. Expression of tenascin C in cardiovascular lesions of Kawasaki disease. Cardiovasc. Pathol. 2018, 38, 25–30. [Google Scholar] [CrossRef]
- Karatas, Z.; Baysal, T.; Alp, H.; Toker, A. Serum tenascin-C: A novel biomarker for diagnosis and predicting prognosis of rheumatic carditis? J. Trop Pediatr. 2013, 59, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Karatas, Z.; Baysal, T.; Sap, F.; Altin, H.; Cicekler, H. The role of tenascin-C and oxidative stress in rheumatic and congenital heart valve diseases: An observational study. Anadolu Kardiyol. Derg. 2013, 13, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Sarli, B.; Topsakal, R.; Kaya, E.G.; Akpek, M.; Lam, Y.Y.; Kaya, M.G. Tenascin-C as predictor of left ventricular remodeling and mortality in patients with dilated cardiomyopathy. J. Investig. Med. 2013, 61, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Yoshikane, Y.; Okuma, Y.; Miyamoto, T.; Hashimoto, J.; Fukazawa, R.; Kato, T.; Takeda, A.; Suda, K.; Hiroe, M.; Imanaka-Yoshida, K. Serum tenascin-C predicts resistance to steroid combination therapy in high-risk Kawasaki disease: A multicenter prospective cohort study. Pediatric Rhematol. 2020, in press. [Google Scholar]
- Sun, R.R.; Lu, L.; Liu, M.; Cao, Y.; Li, X.C.; Liu, H.; Wang, J.; Zhang, P.Y. Biomarkers and heart disease. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2927–2935. [Google Scholar]
- Kii, I. Practical Application of Periostin as a Biomarker for Pathological Conditions. Adv. Exp. Med. Biol. 2019, 1132, 195–204. [Google Scholar] [PubMed]
- Abdelaziz Mohamed, I.; Gadeau, A.P.; Hasan, A.; Abdulrahman, N.; Mraiche, F. Osteopontin: A Promising Therapeutic Target in Cardiac Fibrosis. Cells 2019, 8, 1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Chow, S.L.; Maisel, A.S.; Anand, I.; Bozkurt, B.; de Boer, R.A.; Felker, G.M.; Fonarow, G.C.; Greenberg, B.; Januzzi, J.L., Jr.; Kiernan, M.S.; et al. Role of Biomarkers for the Prevention, Assessment, and Management of Heart Failure: A Scientific Statement From the American Heart Association. Circulation 2017, 135, e1054–e1091. [Google Scholar] [CrossRef]
- Kii, I.; Nishiyama, T.; Li, M.; Matsumoto, K.; Saito, M.; Amizuka, N.; Kudo, A. Incorporation of tenascin-C into the extracellular matrix by periostin underlies an extracellular meshwork architecture. J. Biol. Chem. 2010, 285, 2028–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probstmeier, R.; Montag, D.; Schachner, M. Galectin-3, a beta-galactoside-binding animal lectin, binds to neural recognition molecules. J. Neurochem. 1995, 64, 2465–2472. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Fujimoto, M.; Shiba, M.; Kawakita, F.; Liu, L.; Ichikawa, N.; Kanamaru, K.; Imanaka-Yoshida, K.; Yoshida, T. The Role of Matricellular Proteins in Brain Edema after Subarachnoid Hemorrhage. Acta Neurochir. Suppl. 2016, 121, 151–156. [Google Scholar] [PubMed]
Figure 1.
Schematic illustration showing a hypothetical mechanism by which tenascin-C enhances proinflammatory responses of macrophages.
Figure 1.
Schematic illustration showing a hypothetical mechanism by which tenascin-C enhances proinflammatory responses of macrophages.

Figure 2.
Sequential changes in the expression of TNC in a mouse myocarditis model of coxsackievirus B3 infection. The expression of TNC is observed in the early stage, on day 5 of inoculation. The expression of TNC persists during active inflammation and disappears in the healed phase when mature collagen fibers are formed. Bar, 50 µm.
Figure 2.
Sequential changes in the expression of TNC in a mouse myocarditis model of coxsackievirus B3 infection. The expression of TNC is observed in the early stage, on day 5 of inoculation. The expression of TNC persists during active inflammation and disappears in the healed phase when mature collagen fibers are formed. Bar, 50 µm.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Imanaka-Yoshida, K. Tenascin-C in Heart Diseases—The Role of Inflammation. Int. J. Mol. Sci. 2021, 22, 5828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115828
AMA Style
Imanaka-Yoshida K. Tenascin-C in Heart Diseases—The Role of Inflammation. International Journal of Molecular Sciences. 2021; 22(11):5828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115828
Chicago/Turabian StyleImanaka-Yoshida, Kyoko. 2021. "Tenascin-C in Heart Diseases—The Role of Inflammation" International Journal of Molecular Sciences 22, no. 11: 5828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115828
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.