
Molecular Pathomechanisms of Impaired Flow-Induced Constriction of Cerebral Arteries Following Traumatic Brain Injury: A Potential Impact on Cerebral Autoregulation

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Traumatic Brain Injury (TBI)
2.3. Isolation of Rat Middle Cerebral Arteries and Use of Flow-Chamber to Obtain Vasomotor Responses
2.4. Myogenic Tone and Flow-Induced Responses of Isolated Cerebral Arteries
2.5. Administration of Vasoactive Agents
2.6. Quantitative Real-Time qRT-PCR
2.7. Statistical Analysis
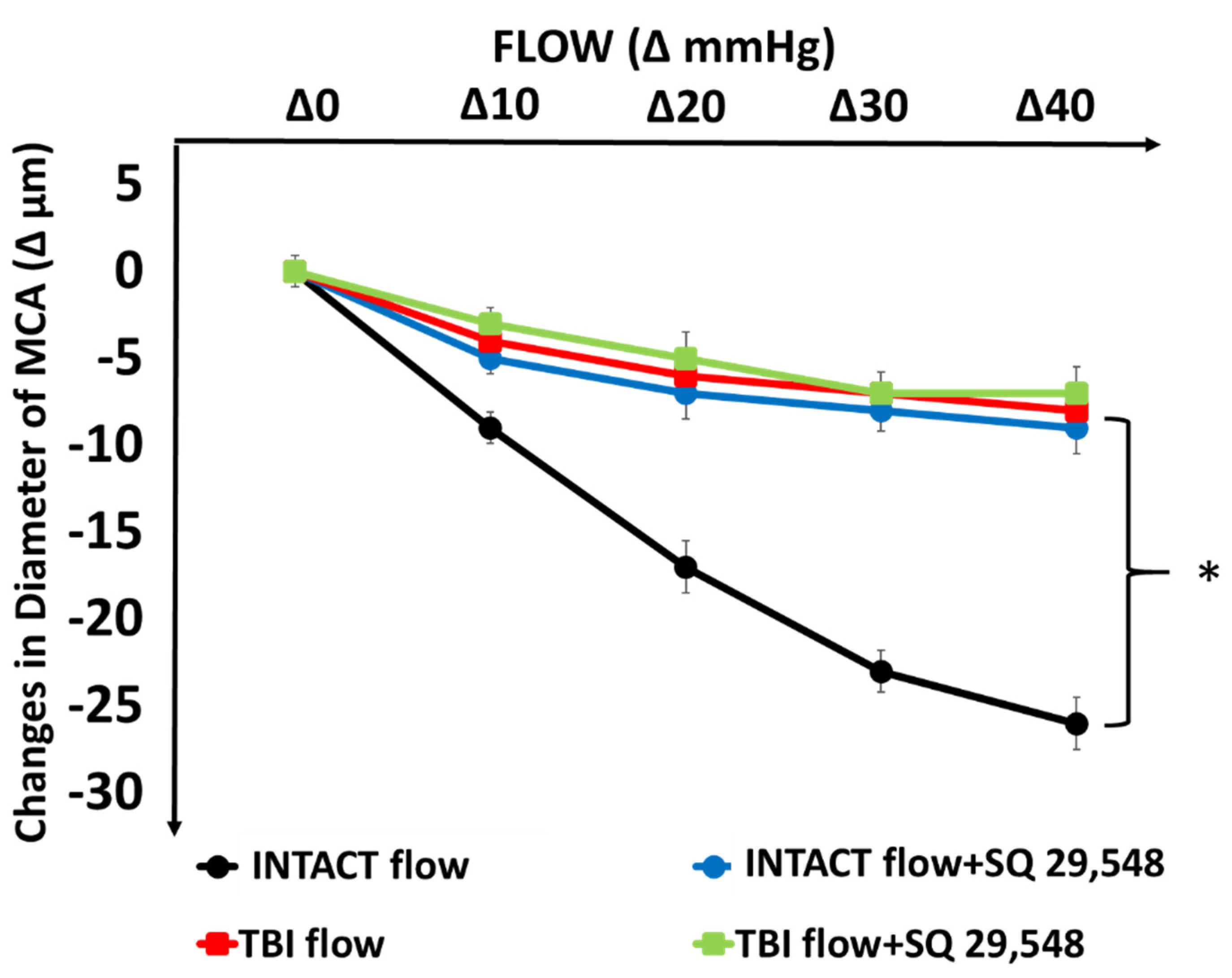
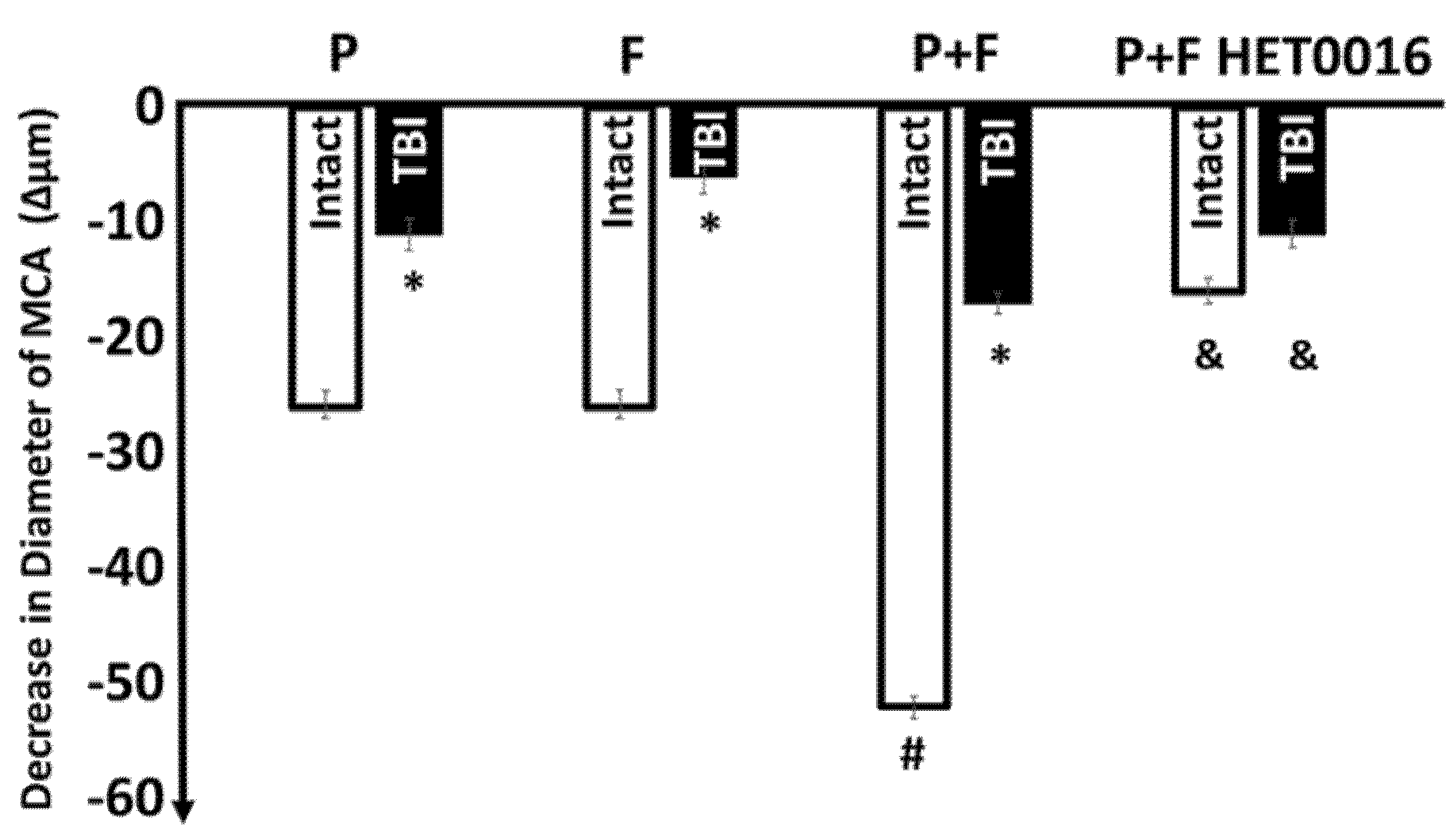
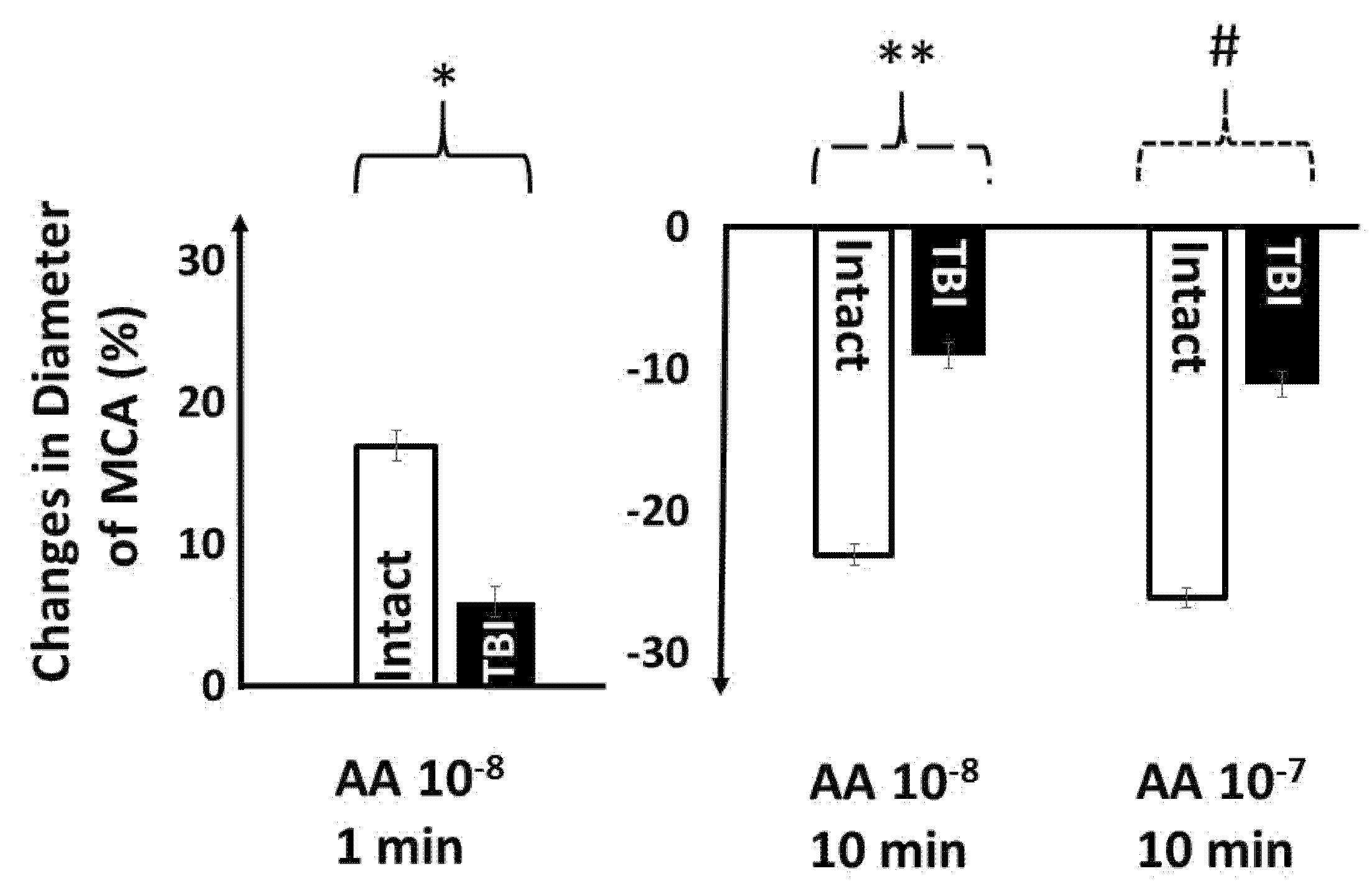
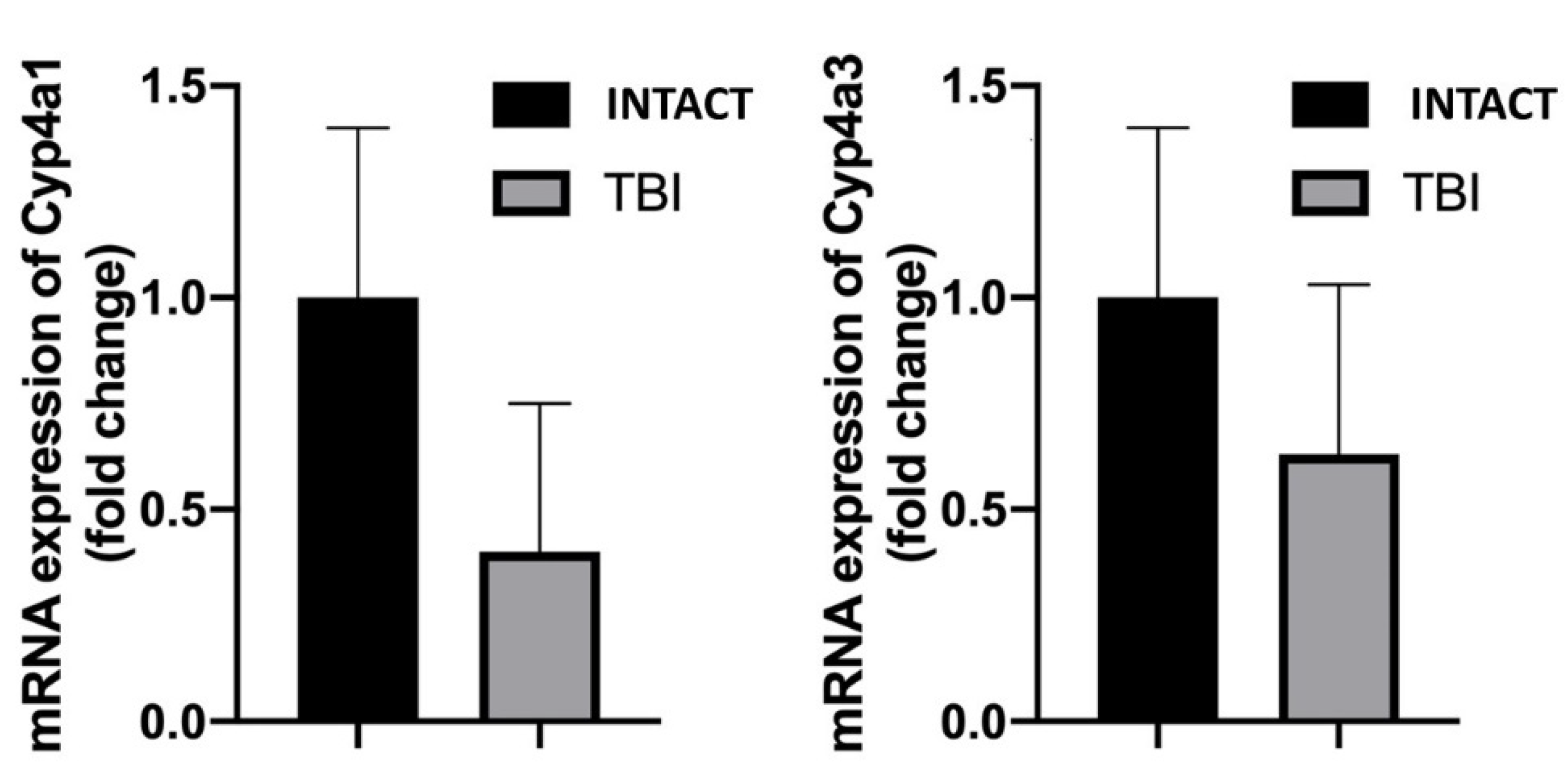
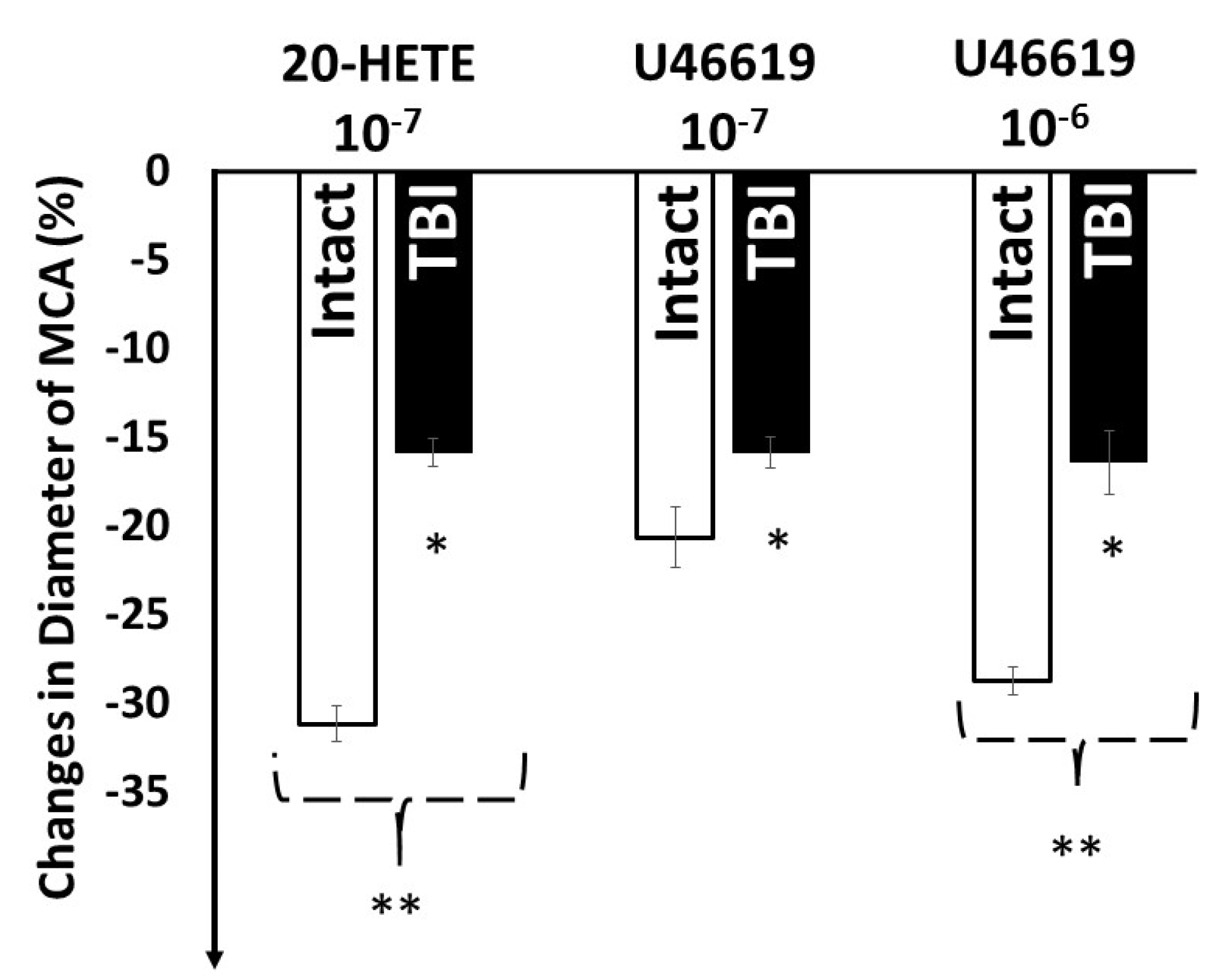
3. Results
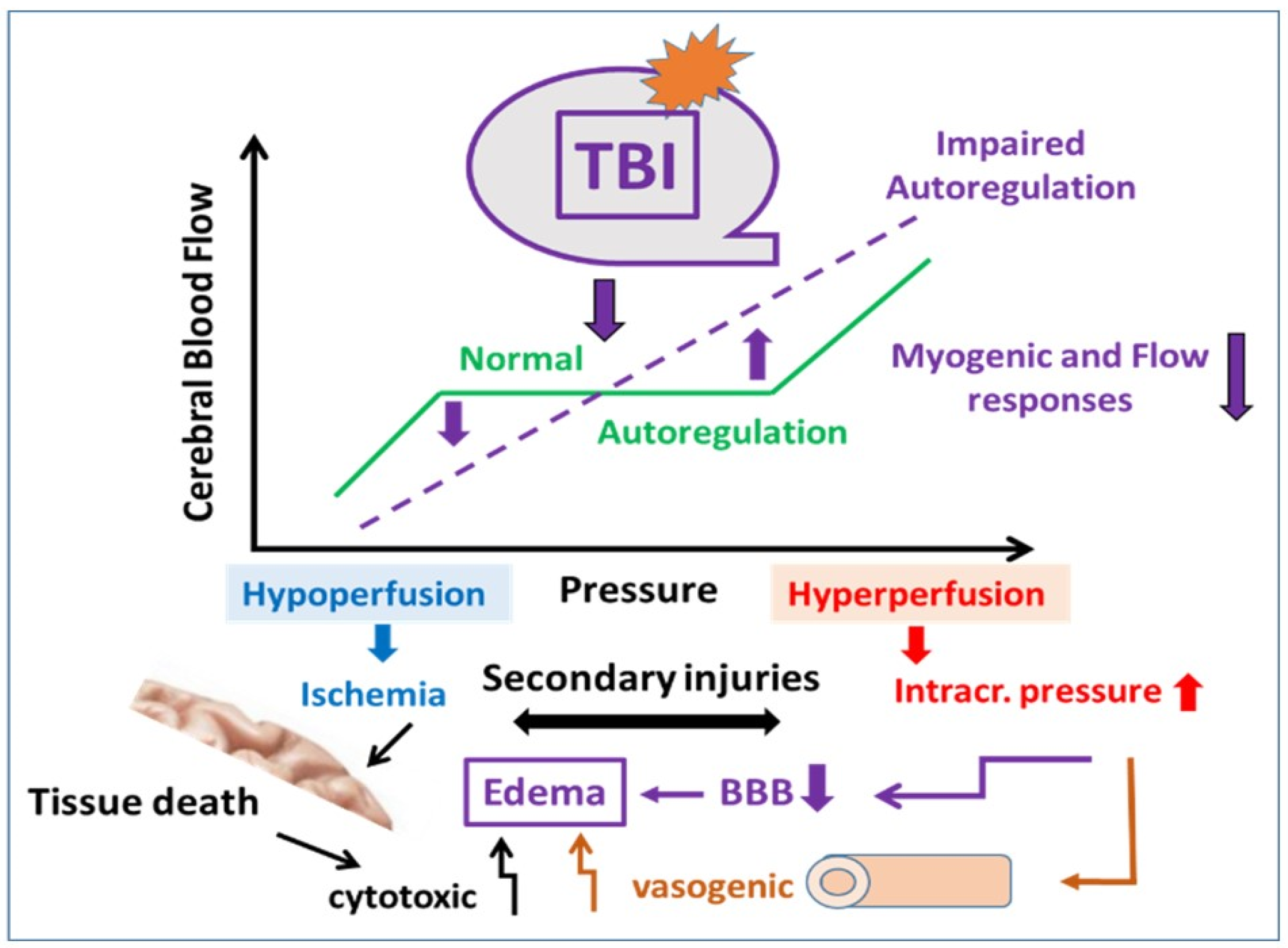
4. Discussion
4.1. Traumatic Brain Injury and Consequences
4.2. Traumatic Brain Injury Impairs Pressure and Flow Sensitive Vasomotor Mechanisms
4.3. Consequences of the TBI-Impaired Mechanosensitive Vasomotor Mechanisms on the Autoregulation of CBF
4.4. Limitation of the Present Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Menyhart, A.; Makra, P.; Szepes, B.E.; Toth, O.M.; Hertelendy, P.; Bari, F.; Farkas, E. High incidence of adverse cerebral blood flow responses to spreading depolarization in the aged ischemic rat brain. Neurobiol. Aging 2015, 36, 3269–3277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapi, D.; Scuri, R.; Colantuoni, A. Trigeminal Cardiac Reflex and Cerebral Blood Flow Regulation. Front. Neurosci. 2016, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Iadecola, C.; Yang, G.; Ebner, T.J.; Chen, G. Local and propagated vascular responses evoked by focal synaptic activity in cerebellar cortex. J. Neurophysiol. 1997, 78, 651–659. [Google Scholar] [CrossRef] [Green Version]
- Koller, A.; Toth, P. Contribution of Flow-Dependent Vasomotor Mechanisms to the Autoregulation of Cerebral Blood Flow. J. Vasc. Res. 2012, 49, 375–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dora, E.; Koller, A.; Kovach, A.G. Effect of topical adenosine deaminase treatment on the functional hyperemic and hypoxic responses of cerebrocortical microcirculation. J. Cereb. Blood Flow Metab. 1984, 4, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipolla, M.J.; Gokina, N.I.; Osol, G. Pressure-induced actin polymerization in vascular smooth muscle as a mechanism underlying myogenic behavior. FASEB J. 2002, 16, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Cipolla, M.J. The Cerebral Circulation; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2009. [Google Scholar]
- Chennupati, R.; Wirth, A.; Favre, J.; Li, R.; Bonnavion, R.; Jin, Y.J.; Wietelmann, A.; Schweda, F.; Wettschureck, N.; Henrion, D.; et al. Myogenic vasoconstriction requires G12/G13 and LARG to maintain local and systemic vascular resistance. eLife 2019, 8, e49374. [Google Scholar] [CrossRef]
- Mandalà, M.; Pedatella, A.L.; Morales Palomares, S.; Cipolla, M.J.; Osol, G. Maturation is associated with changes in rat cerebral artery structure, biomechanical properties and tone. Acta Physiol. 2012, 205, 363–371. [Google Scholar] [CrossRef]
- Osol, G.; Brekke, J.F.; McElroy-Yaggy, K.; Gokina, N.I. Myogenic tone, reactivity, and forced dilatation: A three-phase model of in vitro arterial myogenic behavior. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2260–H2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, P.; Szarka, N.; Farkas, E.; Ezer, E.; Czeiter, E.; Amrein, K.; Ungvari, Z.; Hartings, J.A.; Buki, A.; Koller, A. Traumatic brain injury-induced autoregulatory dysfunction and spreading depression-related neurovascular uncoupling: Pathomechanisms, perspectives, and therapeutic implications. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1118–H1131. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhin, D.; Lange, A.R.; Lowry, T.F.; Taheri, M.R.; Birks, E.K.; Hudetz, A.G.; Narayanan, J.; Falck, J.R.; Okamoto, H.; Roman, R.J.; et al. Production of 20-HETE and its role in autoregulation of cerebral blood flow. Circ. Res. 2000, 87, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Henrion, D. Pressure and flow-dependent tone in resistance arteries. Role of myogenic tone. Arch. Mal. Coeur Vaiss. 2005, 98, 913–921. [Google Scholar]
- Jukic, I.; Mihaljevic, Z.; Matic, A.; Mihalj, M.; Kozina, N.; Selthofer-Relatic, K.; Mihaljevic, D.; Koller, A.; Tartaro Bujak, I.; Drenjancevic, I. Angiotensin II type 1 receptor is involved in flow-induced vasomotor responses of isolated middle cerebral arteries: Role of oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1609–H1624. [Google Scholar] [CrossRef]
- Harder, D.R.; Narayanan, D.J. Gebremedhin Pressure-induced myogenic tone and role of 20-HETE in mediating autoregulation of cerebral blood flow. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H1557–H1565. [Google Scholar] [CrossRef]
- Nemeth, Z.; Hildebrandt, E.; Ryan, M.J.; Granger, J.P.; Drummond, H.A. Pressure-induced constriction of the middle cerebral artery is abolished in TrpC6 knockout mice. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H42–H50. [Google Scholar] [CrossRef] [PubMed]
- Czigler, A.; Toth, L.; Szarka, N.; Szilágyi, K.; Kellermayer, Z.; Harci, A.; Vecsernyes, M.; Ungvari, Z.; Szolics, A.; Koller, A.; et al. Prostaglandin E2, a postulated mediator of neurovascular coupling, at low concentrations dilates whereas at higher concentrations constricts human cerebral parenchymal arterioles. Prostaglandins Lipid Mediat. 2020, 146, 106389. [Google Scholar] [CrossRef]
- Garcia-Roldan, J.L.; Bevan, J.A. Flow-induced constriction and dilation of cerebral resistance arteries. Circ. Res. 1990, 66, 1445–1448. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.; Mandala, M.; Casson, P.; Osol, G. Endothelial dysfunction in a rat model of PCOS: Evidence of increased vasoconstrictor prostanoid activity. Endocrinology 2011, 152, 4927–4936. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, S.; Liu, R.; Travis, O.K.; Roman, R.J.; Fan, F. Cerebral Autoregulation in Hypertension and Ischemic Stroke: A Mini Review. J. Pharm. Sci. Exp. Pharmacol. 2017, 2017, 21–27. [Google Scholar]
- Ungvari, Z.; Pacher, P.; Kecskeméti, V.; Koller, A. Fluoxetine dilates isolated small cerebral arteries of rats and attenuates constrictions to serotonin, norepinephrine, and a voltage-dependent Ca(2+) channel opener. Stroke 1999, 30, 1949–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langlois, J.A.; Sattin, R.W. Traumatic brain injury in the United States: Research and programs of the Centers for Disease Control and Prevention (CDC). J. Head Trauma Rehabil. 2005, 20, 187–188. [Google Scholar] [CrossRef]
- Roozenbeek, B.; Maas, A.I.; Menon, D.K. Changing patterns in the epidemiology of traumatic brain injury. Nat. Rev. Neurol. 2013, 9, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Szarka, N.; Pabbidi, M.R.; Amrein, K.; Czeiter, E.; Berta, G.; Pohoczky, K.; Helyes, Z.; Ungvari, Z.; Koller, A.; Buki, A.; et al. Traumatic Brain Injury Impairs Myogenic Constriction of Cerebral Arteries: Role of Mitochondria-Derived H2O2 and TRPV4-Dependent Activation of BKca Channels. J. Neurotrauma 2018, 35, 930–939. [Google Scholar] [CrossRef]
- Toth, P.; Rozsa, B.; Springo, Z.; Doczi, T.; Koller, A. Isolated human and rat cerebral arteries constrict to increases in flow: Role of 20-HETE and TP receptors. J. Cereb. Blood Flow Metab. 2011, 31, 2096–2105. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.B.; Mechtler, L.; Ali, B.; Swedler, D.; Kelley-Baker, T. Cannabis and crash risk among older drivers. Accid. Anal. Prev. 2021, 152, 105987. [Google Scholar] [CrossRef] [PubMed]
- Coronado, V.G.; Xu, L.; Basavaraju, S.V.; McGuire, L.C.; Wald, M.M.; Faul, M.D.; Guzman, B.R.; Hemphill, J.D. Surveillance for traumatic brain injury-related deaths—United States, 1997–2007; Centers for Disease Control and Prevention (CDC). MMWR Surveill Summ. 2011, 60, 1–32. [Google Scholar]
- Schaller, B.; Graf, R. Different Compartments of Intracranial Pressure and Its Relationship to Cerebral Blood Flow. J. Trauma Inj. Infect. Crit. Care 2005, 59, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Toth, P.; Tucsek, Z.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Tarantini, S.; Deak, F.; Koller, A.; Sonntag, W.E.; Csiszar, A.; et al. Age-related autoregulatory dysfunction and cerebromicrovascular injury in mice with angiotensin II-induced hypertension. J. Cereb. Blood Flow Metab. 2013, 33, 1732–1742. [Google Scholar] [CrossRef]
- Toth, P.; Tucsek, Z.; Tarantini, S.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Koller, A.; Sonntag, W.E.; Csiszar, A.; Ungvari, Z. IGF-1 deficiency impairs cerebral myogenic autoregulation in hypertensive mice. J. Cereb. Blood Flow Metab. 2014, 34, 1887–1897. [Google Scholar] [CrossRef] [Green Version]
- Escalante, B.; Sessa, W.C.; Falck, J.R.; Yadagiri, P.; Schwartzman, M.L. Vasoactivity of 20-hydroxyeicosatetraenoic acid is dependent on metabolism by cyclooxygenase. J. Pharmacol. Exp. Ther. 1989, 248, 229–232. [Google Scholar]
- Schwartzman, M.L.; Falck, J.R.; Yadagiri, P.; Escalante, B. Metabolism of 20-hydroxyeicosatetraenoic acid by cyclooxygenase. Formation and identification of novel endothelium-dependent vasoconstrictor metabolites. J. Biol. Chem. 1989, 264, 11658–11662. [Google Scholar] [CrossRef]
- Harder, D.R.; Gebremedhin, D.; Narayanan, J.; Jefcoat, C.; Falck, J.R.; Campbell, W.B.; Roman, R. Formation and action of a P-450 4A metabolite of arachidonic acid in cat cerebral microvessels. Am. J. Physiol. 1994, 266, H2098–H2107. [Google Scholar] [CrossRef]
- Yu, M.; Cambj-Sapunar, L.; Kehl, F.; Maier, K.G.; Takeuchi, K.; Miyata, N.; Ishimoto, T.; Reddy, L.M.; Falck, J.R.; Gebremedhin, D.; et al. Effects of a 20-HETE antagonist and agonists on cerebral vascular tone. Eur. J. Pharmacol. 2004, 486, 297–306. [Google Scholar] [CrossRef]
- Marmarou, A.; Foda, M.A.; van den Brink, W.; Campbell, J.; Kita, H.; Demetriadou, K. A new model of diffuse brain injury in rats. PartI: Pathopphysiology and biomechanics. J. Neurosurg. 1994, 80, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szarka, N.; Toth, L.; Czigler, A.; Kellermayer, Z.; Ungvari, Z.; Amrein, K.; Czeiter, E.; Bali, Z.K.; Tadepalli, S.A.; Wahr, M.; et al. Single Mild Traumatic Brain Injury Induces Persistent Disruption of the Blood-Brain Barrier, Neuroinflammation and Cognitive Decline in Hypertensive Rats. Int. J. Mol. Sci. 2019, 20, 3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashida, M.; Fukunaga, A.; Hanaoka, K. An animal model for surgical anesthesia and analgesia: Characterization with isoflurane anesthesia and remifentanil analgesia. Anesth. Analg. 2003, 97, 1340–1346. [Google Scholar] [CrossRef]
- Golding, E.M.; Contant, C.F., Jr.; Robertson, C.S.; Bryan, R.M., Jr. Temporal effect of severe controlled cortical impact injury in the rat on the myogenic response of the middle cerebral artery. J. Neurotrauma 1998, 15, 973–984. [Google Scholar] [CrossRef]
- Racz, A.; Veresh, Z.; Lotz, G.; Bagi, Z.; Koller, A. Cyclooxygenase-2 derived thromboxane A(2) and reactive oxygen species mediate flow-induced constrictions of venules in hyperhomocysteinemia. Atherosclerosis 2010, 208, 43–49. [Google Scholar] [CrossRef]
- Szarka, N.; Amrein, K.; Horvath, P.; Ivic, I.; Czeiter, E.; Buki, A.; Koller, A.; Toth, P. Hypertension-Induced Enhanced Myogenic Constriction of Cerebral Arteries Is Preserved after Traumatic Brain Injury. J. Neurotrauma 2017, 34, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.P.; Fleming, J.T.; Falck, J.R.; Jacobs, E.R.; Gebremedhin, D.; Harder, D.R.; Roman, R.J. 20-HETE is an endogenous inhibitor of the large-conductance Ca2+-activated K+ channel in renal arterioles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1996, 270, R228–R237. [Google Scholar] [CrossRef]
- Gebremedhin, D.; Yamaura, K.; Harder, D.R. Role of 20-HETE in the hypoxia-induced activation of Ca2+-activated K+ channel currents in rat cerebral arterial muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H107–H120. [Google Scholar] [CrossRef]
- Toth, P.; Csiszar, A.; Tucsek, Z.; Sosnowska, D.; Gautam, T.; Koller, A.; Schwartzman, M.L.; Sonntag, W.E.; Ungvari, Z. Role of 20-HETE, TRPC channels, and BKCa in dysregulation of pressure-induced Ca2+ signaling and myogenic constriction of cerebral arteries in aged hypertensive mice. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1698–H1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkin, J.J.; Vink, R. Mechanisms of cerebral edema in traumatic brain injury: Therapeutic developments. Curr. Opin. Neurol. 2010, 23, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Marmarou, A. Traumatic brain edema: An overview. Acta Neurochir. Suppl. 1994, 60, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Poloyac, S.M.; Reynolds, R.B.; Yonas, H.; Kerr, M.E. Identification and quantification of the hydroxyeicosatetraenoic acids, 20-HETE and 12-HETE, in the cerebrospinal fluid after subarachnoid hemorrhage. J. Neurosci. Methods 2005, 144, 257–263. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szenasi, A.; Amrein, K.; Czeiter, E.; Szarka, N.; Toth, P.; Koller, A. Molecular Pathomechanisms of Impaired Flow-Induced Constriction of Cerebral Arteries Following Traumatic Brain Injury: A Potential Impact on Cerebral Autoregulation. Int. J. Mol. Sci. 2021, 22, 6624. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126624
Szenasi A, Amrein K, Czeiter E, Szarka N, Toth P, Koller A. Molecular Pathomechanisms of Impaired Flow-Induced Constriction of Cerebral Arteries Following Traumatic Brain Injury: A Potential Impact on Cerebral Autoregulation. International Journal of Molecular Sciences. 2021; 22(12):6624. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126624
Chicago/Turabian StyleSzenasi, Annamaria, Krisztina Amrein, Endre Czeiter, Nikolett Szarka, Peter Toth, and Akos Koller. 2021. "Molecular Pathomechanisms of Impaired Flow-Induced Constriction of Cerebral Arteries Following Traumatic Brain Injury: A Potential Impact on Cerebral Autoregulation" International Journal of Molecular Sciences 22, no. 12: 6624. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126624