
High-Throughput Screen Detects Calcium Signaling Dysfunction in Hutchinson-Gilford Progeria Syndrome


Abstract
:1. Introduction
2. Results
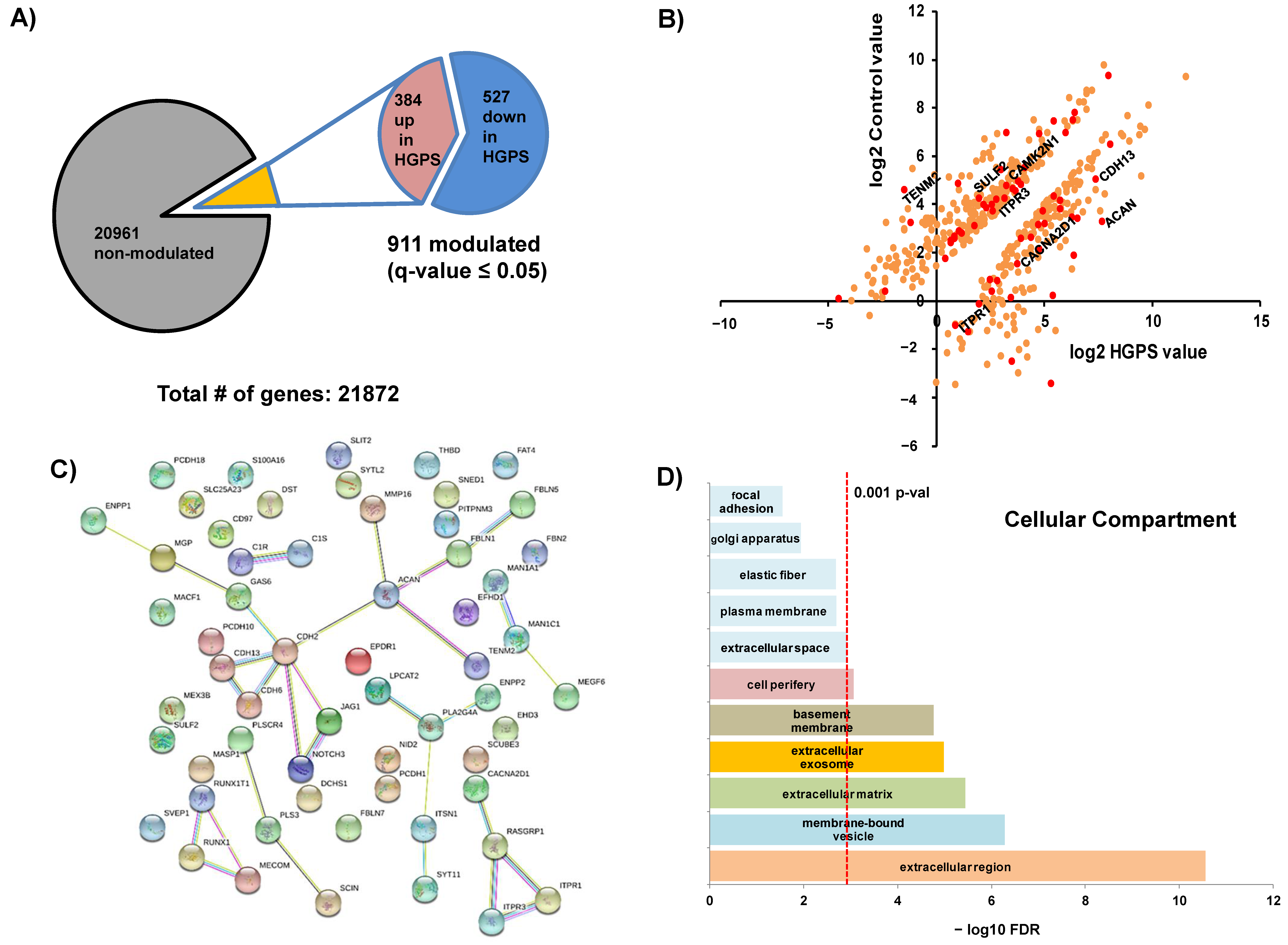
2.1. RNAseq Analysis of HGPS and Healthy Control Cell Lines
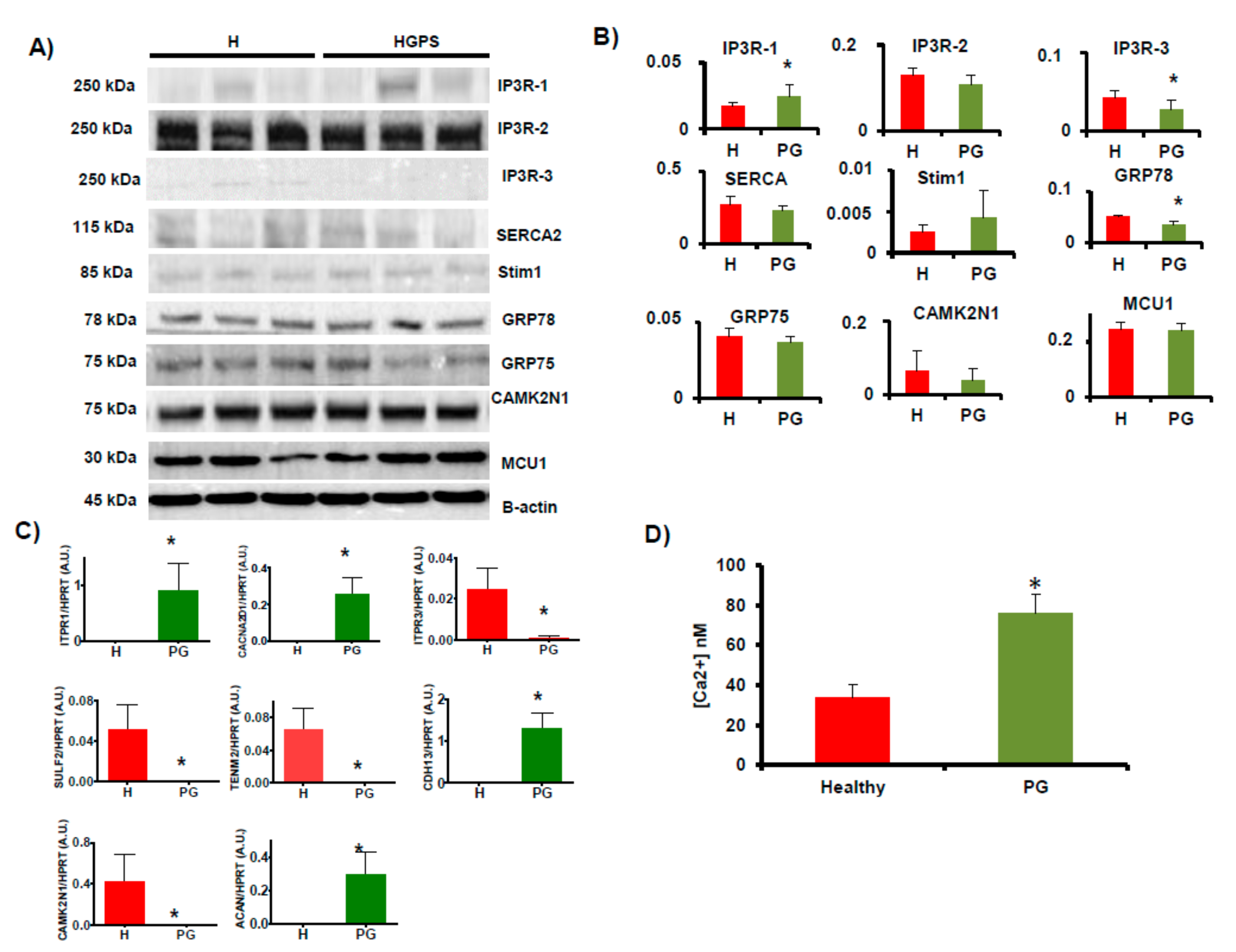
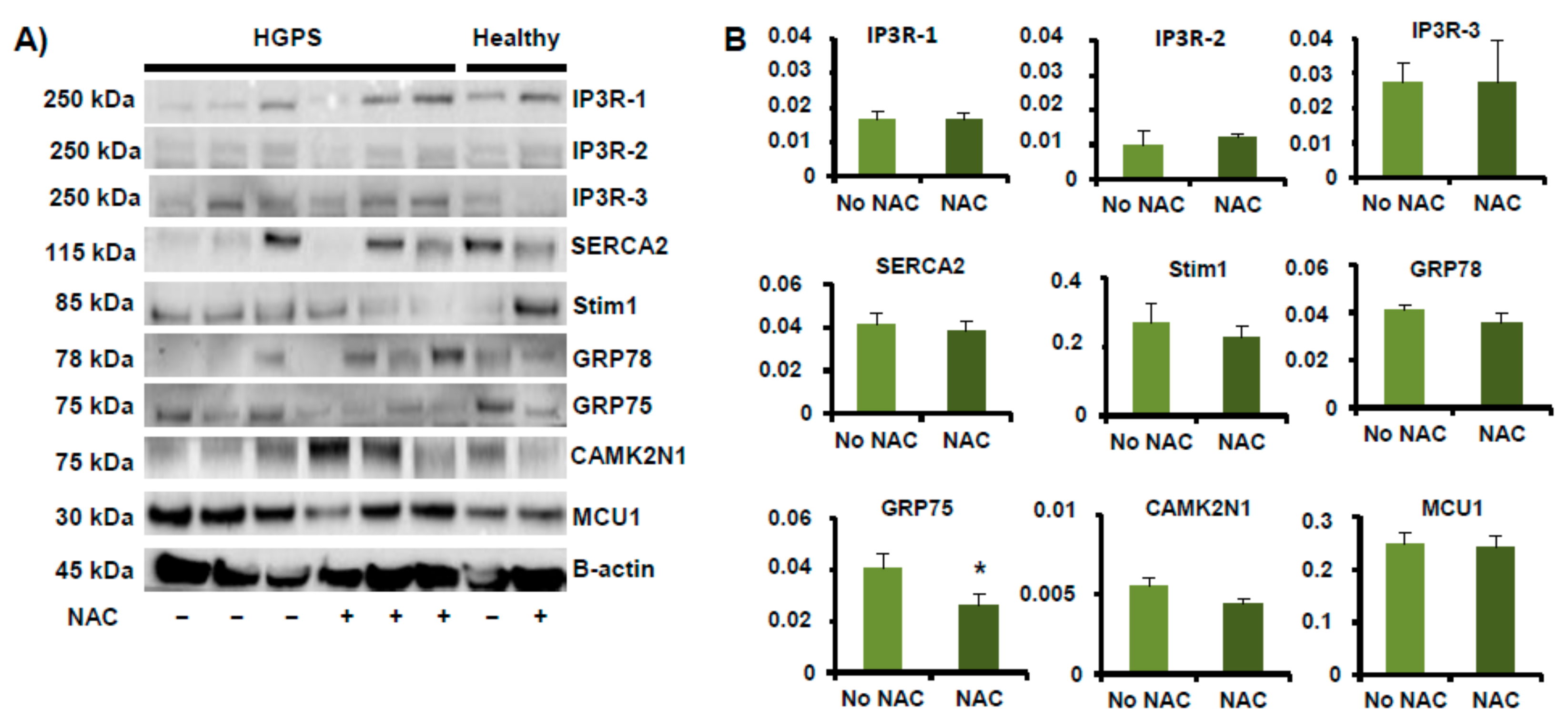
2.2. Orthogonal Validation of Ca2+ Flux-Related Proteins and Transcripts
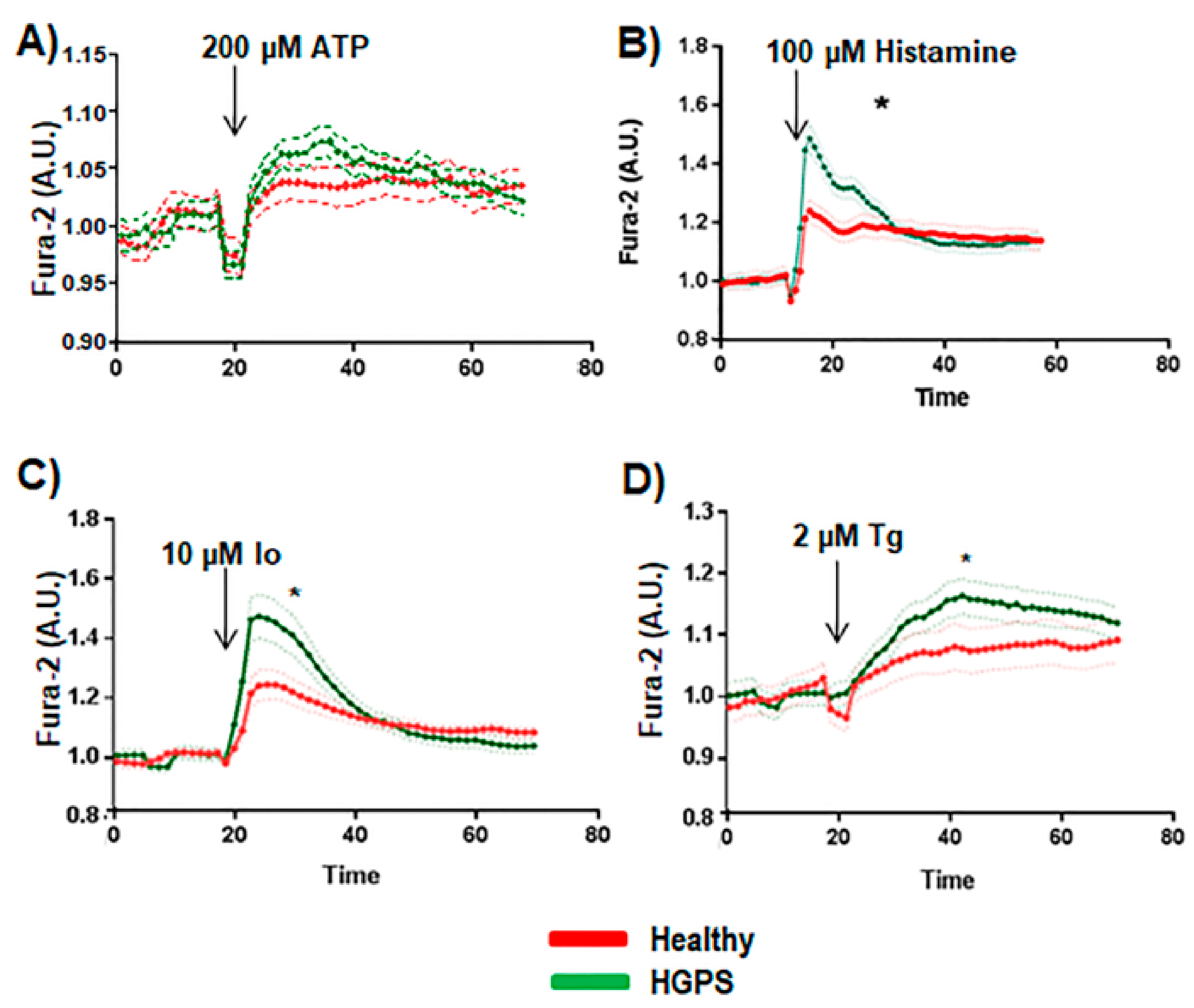
2.3. Calcium Dynamics Measurement
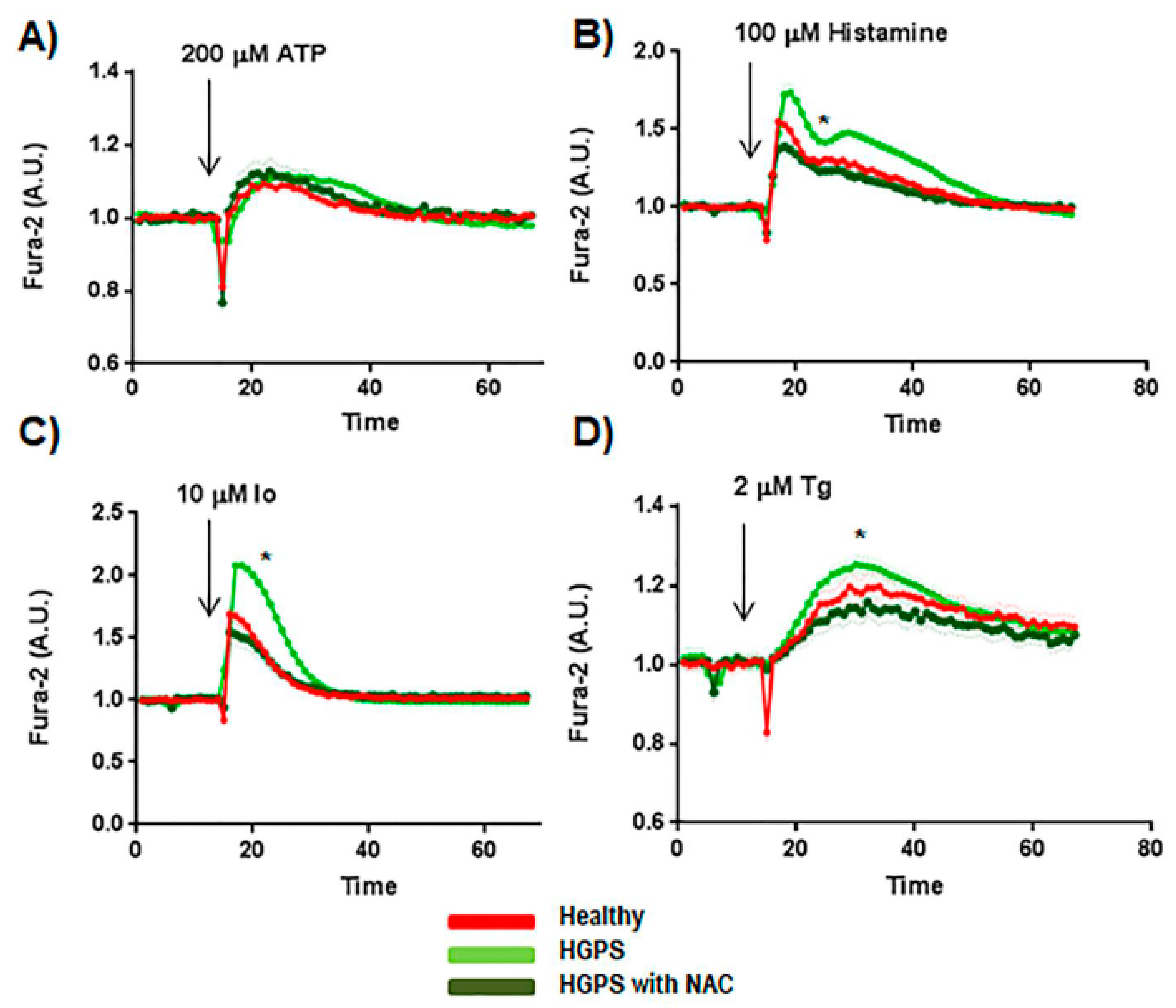
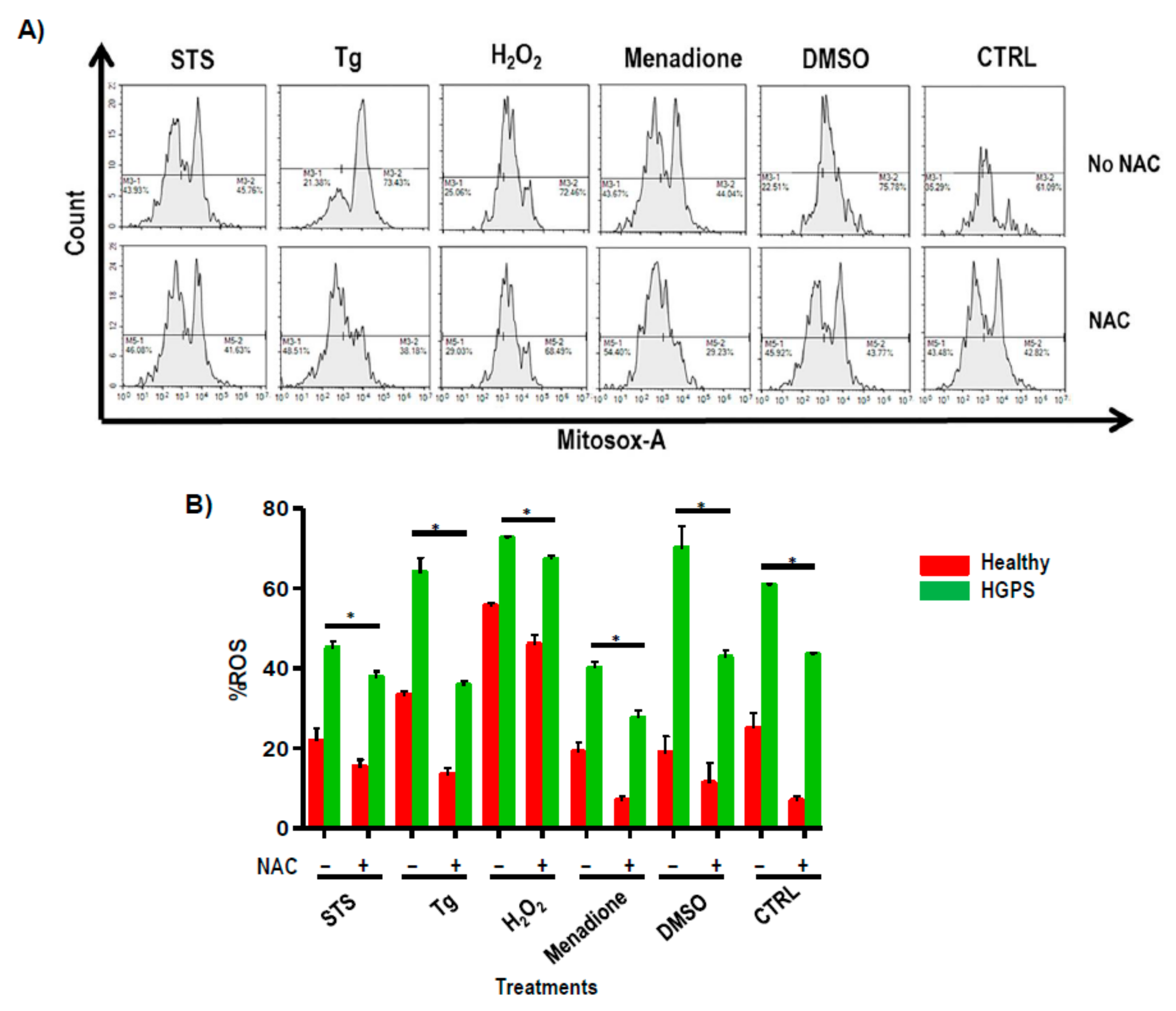
2.4. Effect of ROS Scavengers in the Ca2+ Flux
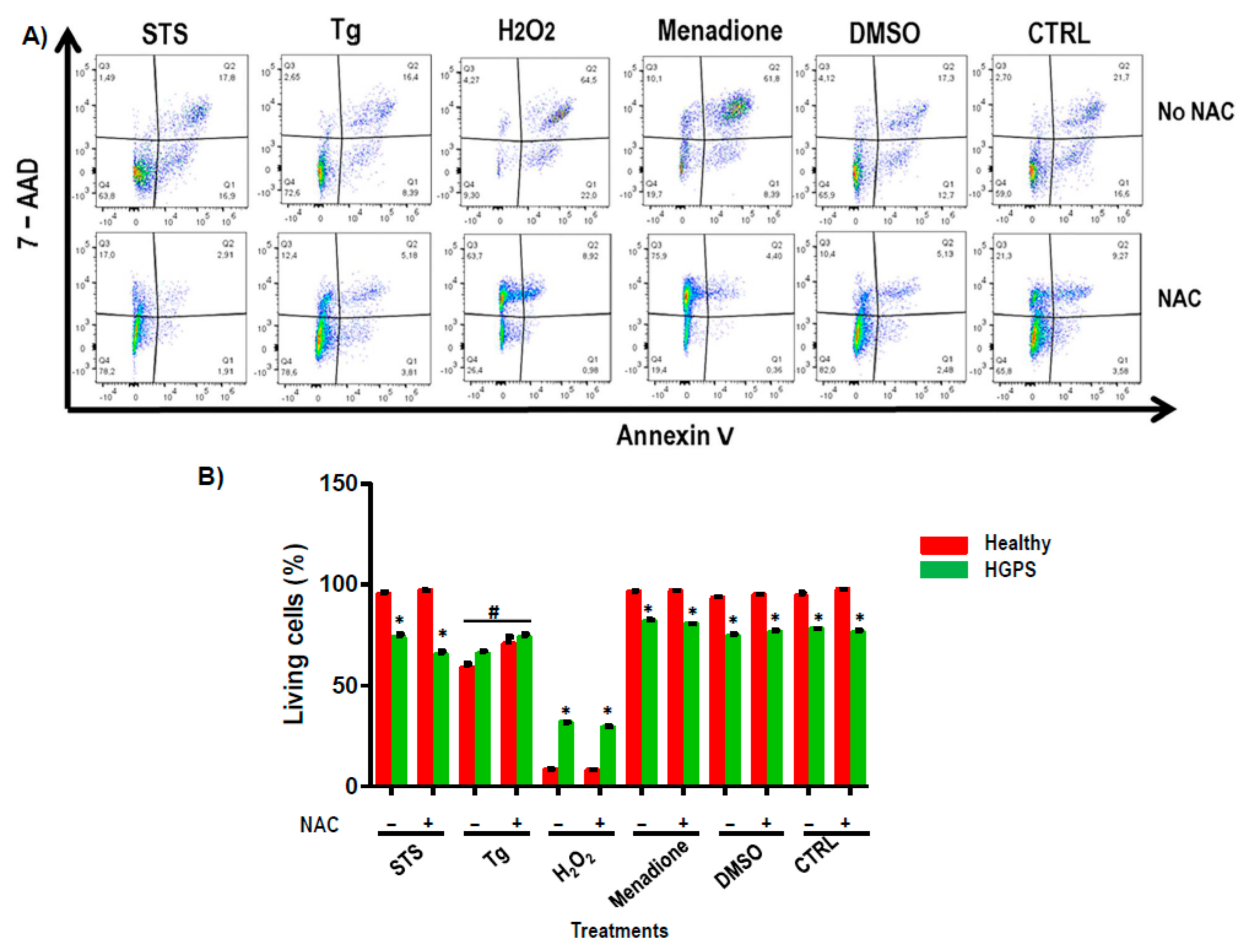
2.5. Apoptosis Study
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA-Seq Protocol
4.3. Bioinformatic Analysis
4.4. qRT-PCR
4.5. Western Blot Analysis
4.6. Basal Calcium Measurement
- Kd = dissociation constant of Fura-2 for Ca2+ at room temperature (241 nM)
- R = basal ratio
- Rmin = minimum ratio (R-value after the addition of EGTA)
- Rmax = maximum ratio (R-value after the addition of digitonin)
- Sf2 = F380 max
- Sb2 = FF380 min
4.7. Cytosolic Calcium Handling
4.8. Flow Cytometry
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | RNAsec Data: Up in | Cellular Component |
|---|---|---|---|
| ITPR1 | Inositol 1,4,5-trisphosphate receptor type 1 | HGPS | Endoplasmic reticulum membrane |
| CACNA2D1 | Voltage-dependent calcium channel subunit alpha-2/delta-1 | HGPS | Sarcoplasmic reticulum, extracelular exosome, plasma membrane |
| ITPR3 | Inositol 1,4,5-trisphosphate receptor type 3 | Healthy | Endoplasmic reticulum membrane |
| CAMK2N1 | Calcium/calmodulin-dependent protein kinase II inhibitor 1 | Healthy | Endoplasmic reticulum membrane |
| SULF2 | Extracellular sulfatase Sulf-2 | Healthy | Golgi apparatus, endoplasmic reticulum |
| TENM2 | Teneurin-2 | Healthy | Nucleus, Golgi apparatus, endoplasmic reticulum, plasma membrane |
| CDH13 | Cadherin-13 | HGPS | Plasma membrane |
| ACAN | Aggrecan core protein | HGPS | Extracellular matrix |
| Gene Name | Forward 5′ | Reverse 5′ | mRNA ID |
|---|---|---|---|
| ITPR1 | CCTGTTCAAGTTGTGTGCGA | CCTCCCACCAACTGAGATGT | NG_016144.1 |
| CACNA2D1 | AGTGGGAGGAGTGAGAGGAT | TCTACAATCCCAAACGGCCT | NG_009358.2 |
| ITPR3 | GTCTCCTGCTCCCTGTCTTT | AGAAATAAAGGTCGGGGCCA | NG_027729.1 |
| CAMK2N1 | TTGCAGTCTGTTGTGTGAGC | CCAAGCATTCCTGTACGTGG | NM_018584.6 |
| SULF2 | AAGGATGGTGGGGACTTCAG | CGGACTTCCCTCAGGTTCTT | AB033073.2 |
| TENM2 | CCTTTCAACAGCAGCTTGGT | CACCTCCATTTGCGATCAGG | AB032953.2 |
| CDH13 | GAATGAGAAAGCAGGCGAGG | TTGCCCTCCATGAGCTGTTA | NG_052819.1 |
| ACAN | TCCCCGTAGAAGAGGAGACA | AAGACAGGGGTATGCAGCTT | NG_012794.1 |
| HPRT | ATGACCAGTCAACAGGGGACA | GCTTGCGACCTTGACCATCT | NG_012329.2 |
| Gene Name | Antibody | Source | Commercial Brand |
|---|---|---|---|
| ITPR3 | Inositol 1,4,5-trisphosphate receptor type 3 | Rabbit | ACC-010 Alomone Labs. |
| VDAC | Voltage-dependent anion-selective channel protein 1 | Rabbit | AVC-001 Alomone Labs. |
| ITPR1 | Inositol 1,4,5-trisphosphate receptor type 1 | Rabbit | ACC-019 Alomone Labs. |
| GRP78 | Endoplasmic reticulum chaperone BiP | Rabbit | 3183 Cell Signaling |
| MCU | Calcium uniporter protein, mitochondrial | Rabbit | ACC-328 Alomone Labs. |
| SERCA2 | Sarcoplasmic/endoplasmic reticulum calcium ATPase 2 | Rabbit | ACP-012 Alomone Labs. |
| STIM1 | Stromal interaction molecule 1 | Rabbit | ACC-063-AO. Alomone Labs. |
| GRP75 | Stress-70 protein, mitochondrial | Rabbit | 3177. Cell Signaling |
| Tubulin | Tubulin alpha-1B chain | Rabbit | 2128S. Cell Signaling |
| B-actin | Actin-related protein 3B | Rabbit | 4970S. Cell Signaling |
| TOM20 | Mitochondrial import receptor subunit TOM20 homolog | Mouse | 612278 BD |
| CALM1 | Calmodulin-1 | Mouse | Ab-38841. Abcam |
| Anti-rabbit IgG-HRP | Goat | Sc-2004. Sta. Cruz | |
| Anti-mouse IgG-HRP | Goat | Sc-2005. Sta. Cruz |
References
- Guenantin, A.C.; Briand, N.; Bidault, G.; Afonso, P.; Bereziat, V.; Vatier, C.; Lascols, O.; Caron-Debarle, M.; Capeau, J.; Vigouroux, C. Nuclear envelope-related lipodystrophies. Semin. Cell Dev. Biol. 2014, 29, 148–157. [Google Scholar] [CrossRef]
- Worman, H.J. Prelamin A prenylation and the treatment of progeria. J. Lipid Res. 2010, 51, 223–225. [Google Scholar] [CrossRef] [Green Version]
- Hennekam, R.C. Hutchinson-Gilford progeria syndrome: Review of the phenotype. Am. J. Med. Genet. A 2006, 140, 2603–2624. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, M.; Brown, W.T.; Gordon, L.B.; Glynn, M.W.; Singer, J.; Scott, L.; Erdos, M.R.; Robbins, C.M.; Moses, T.Y.; Berglund, P.; et al. Recurrent de novo point mutations in lamin A cause Hutchinson-Gilford progeria syndrome. Nature 2003, 423, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.; Comai, L.; Lamin, A. Farnesylation and aging. Exp. Cell Res. 2012, 318, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, V.; Roedl, D.; Gabriel, D.; Gordon, L.B.; Herlyn, M.; Schneider, R.; Ring, J.; Djabali, K. Naïve adult stem cells from patients with Hutchinson-Gilford progeria syndrome express low levels of progerin in vivo. Biol. Open 2012, 1, 516–526. [Google Scholar] [CrossRef] [Green Version]
- Viteri, G.; Chung, Y.W.; Stadtman, E.R. Effect of progerin on the accumulation of oxidized proteins in fibroblasts from Hutchinson Gilford progeria patients. Mech. Ageing Dev. 2010, 131, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Mateos, J.; De la Fuente, A.; Lesende-Rodriguez, I.; Fernández-Pernas, P.; Arufe, M.C.; Blanco, F.J. Lamin A deregulation in human mesenchymal stem cells promotes an impairment in their chondrogenic potential and imbalance in their response to oxidative stress. Stem Cell Res. 2013, 11, 1137–1148. [Google Scholar] [CrossRef] [Green Version]
- Mateos, J.; Landeira-Abia, A.; Fafián-Labora, J.A.; Fernández-Pernas, P.; Lesende-Rodríguez, I.; Fernández-Puente, P.; Fernández-Moreno, M.; Delmiro, A.; Martín, M.A.; Blanco, F.J.; et al. iTRAQ-based analysis of progerin expression reveals mitochondrial dysfunction, reactive oxygen species accumulation and altered proteostasis. Stem Cell Res. Ther. 2015, 6, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollimuntha, S.; Selvaraj, S.; Singh, B.B. Emerging roles of canonical TRP channels in neuronal function. Adv. Exp. Med. Biol. 2011, 704, 573–593. [Google Scholar] [PubMed] [Green Version]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.; Gutiérrez, A.; Núñez, M.; Cocca, C.; Martín, G.; Cricco, G.; Medina, V.; Rivera, E.; Bergoc, R. Mitochondrial apoptotic pathways. Biocell 2005, 29, 149–161. [Google Scholar] [CrossRef]
- Vance, J.E. MAM (mitochondria-associated membranes) in mammalian cells: Lipids and beyond. Biochim. Biophys. Acta 2014, 1841, 595–609. [Google Scholar] [CrossRef]
- Shimizu, S. Organelle zones in mitochondria. J. Biochem. 2019, 165, 101–107. [Google Scholar] [CrossRef]
- Chen, W.M.; Chiang, J.C.; Lin, Y.C.; Lin, Y.N.; Chuang, P.Y.; Chang, Y.C.; Chen, C.C.; Wu, K.Y.; Hsieh, J.C.; Chen, S.K.; et al. Lysophosphatidic acid receptor LPA3 prevents oxidative stress and cellular senescence in Hutchinson-Gilford progeria syndrome. Aging Cell 2020, 19, e13064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateos, J.; Fafian-Labora, J.; Morente-López, M.; Lesende-Rodriguez, I.; Monserrat, L.; Ódena, M.A.; Oliveira, E.D.; De Toro, J.; Arufe, M.C. Next-Generation Sequencing and Quantitative Proteomics of Hutchinson-Gilford progeria syndrome-derived cells point to a role of nucleotide metabolism in premature aging. PLoS ONE 2018, 13, e0205878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bárcena, C.; López-Otín, C.; Kroemer, G. Methionine restriction for improving progeria: Another autophagy-inducing anti-aging strategy? Autophagy 2019, 15, 558–559. [Google Scholar] [CrossRef] [Green Version]
- Kasri, N.N.; Kocks, S.L.; Verbert, L.; Hébert, S.S.; Callewaert, G.; Parys, J.B.; Missiaen, L.; De Smedt, H. Up-regulation of inositol 1,4,5-trisphosphate receptor type 1 is responsible for a decreased endoplasmic-reticulum Ca2+ content in presenilin double knock-out cells. Cell Calcium 2006, 40, 41–51. [Google Scholar] [CrossRef]
- Lo, C.Y.; Tjong, Y.W.; Ho, J.C.Y.; Siu, C.W.; Cheung, S.Y.; Tang, N.L.; Yu, S.; Tse, H.F.; Yao, X. An upregulation in the expression of vanilloid transient potential channels 2 enhances hypotonicity-induced cytosolic Ca2⁺ rise in human induced pluripotent stem cell model of Hutchinson-Gillford Progeria. PLoS ONE 2014, 9, e87273. [Google Scholar] [CrossRef] [Green Version]
- Prasad, M.; Pawlak, K.J.; Burak, W.E.; Perry, E.E.; Marshall, B.; Whittal, R.M.; Bose, H.S. Mitochondrial metabolic regulation by GRP78. Sci. Adv. 2017, 3, e1602038. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.; Mitchell, C.; Park, M.A.; Yacoub, A.; Rahmani, M.; Häussinger, D.; Reinehr, R.; Voelkel-Johnson, C.; Fisher, P.B.; Grant, S.; et al. 17-allylamino-17-demethoxygeldanamycin and MEK1/2 inhibitors kill GI tumor cells via Ca2+-dependent suppression of GRP78/BiP and induction of ceramide and reactive oxygen species. Mol. Cancer Ther. 2010, 9, 1378–1395. [Google Scholar] [CrossRef] [Green Version]
- Paillard, M.; Csordás, G.; Huang, K.T.; Várnai, P.; Joseph, S.K.; Hajnóczky, G. MICU1 Interacts with the D-Ring of the MCU Pore to Control Its Ca. Mol. Cell 2018, 72, 778–785.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, B.; Lee, A.S. The critical roles of endoplasmic reticulum chaperones and unfolded protein response in tumorigenesis and anticancer therapies. Oncogene 2013, 32, 805–818. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Li, Y.; Zhang, D.; Zhang, A.; Guo, W.; Zhu, S. HMGB1-induced asthmatic airway inflammation through GRP75-mediated enhancement of ER-mitochondrial Ca. J. Cell Biochem. 2018, 119, 4205–4215. [Google Scholar] [CrossRef]
- Baumgartner, H.K.; Gerasimenko, J.V.; Thorne, C.; Ferdek, P.; Pozzan, T.; Tepikin, A.V.; Petersen, O.H.; Sutton, R.; Watson, A.J.; Gerasimenko, O.V. Calcium elevation in mitochondria is the main Ca2+ requirement for mitochondrial permeability transition pore (mPTP) opening. J. Biol. Chem. 2009, 284, 20796–20803. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.Y.; Cho, H.J.; Yu, S.N.; Kim, S.H.; Yu, H.S.; Park, Y.M.; Mirkheshti, N.; Kim, S.Y.; Song, C.S.; Chatterjee, B.; et al. Interplay of reactive oxygen species, intracellular Ca2+ and mitochondrial homeostasis in the apoptosis of prostate cancer cells by deoxypodophyllotoxin. J. Cell Biochem. 2013, 114, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.A.; Muter, J.; Pamela Ritchie, P.; Lattanzi, G.; Hutchison, C.J. The accumulation of un-repairable DNA damage in laminopathy progeria fibroblasts is caused by ROS generation and is prevented by treatment with N-acetyl cysteine. Hum. Mol. Genet. 2011, 20, 3997–4004. [Google Scholar] [CrossRef] [Green Version]
- Paillard, M.; Tubbs, E.; Thiebaut, P.A.; Gomez, L.; Fauconnier, J.; Crola Da Silva, C.; Teixeira, G.; Mewton, N.; Belaidi, E.; Durand, A.; et al. Depressing mitochondria-reticulum interactions protects cardiomyocytes from lethal hypoxia-reoxygenation injury. Circulation 2013, 128, 1555–1565. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chang, X.; Zhang, Y.; Wu, C.; Li, R.; Tang, L.; Zhou, Z. Fluorochloridone induces primary cultured Sertoli cells apoptosis: Involvement of ROS and intracellular calcium ions-mediated ERK1/2 activation. Toxicology 2018, 47, 228–237. [Google Scholar] [CrossRef]
- Mohsin, A.A.; Thompson, J.; Hu, Y.; Hollander, J.; Lesnefsky, E.J.; Chen, Q. Endoplasmic reticulum stress-induced complex I defect: Central role of calcium overload. Arch. Biochem. Biophys. 2020, 683, 108299. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Torres, J.; Acin-Perez, R.; Cabezas-Sánchez, P.; Osorio, F.G.; Gonzalez-Gomez, C.; Megias, D.; Cámara, C.; López-Otín, C.; Enríquez, J.A.; Luque-García, J.L.; et al. Identification of mitochondrial dysfunction in Hutchinson-Gilford progeria syndrome through use of stable isotope labeling with amino acids in cell culture. J. Proteom. 2013, 91, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Paschen, W.; Mengesdorf, T.; Althausen, S.; Hotop, S. Peroxidative stress selectively down-regulates the neuronal stress response activated under conditions of endoplasmic reticulum dysfunction. J. Neurochem. 2001, 76, 1916–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younce, C.W.; Burmeister, M.A.; Ayala, J.E. Exendin-4 attenuates high glucose-induced cardiomyocyte apoptosis via inhibition of endoplasmic reticulum stress and activation of SERCA2a. Am. J. Physiol. Cell Physiol. 2013, 304, C508–C518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scrima, R.; Piccoli, C.; Moradpour, D.; Capitanio, N. Targeting Endoplasmic Reticulum and/or Mitochondrial Ca. Front. Chem. 2018, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 408. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fafián-Labora, J.A.; Morente-López, M.; de Toro, F.J.; Arufe, M.C. High-Throughput Screen Detects Calcium Signaling Dysfunction in Hutchinson-Gilford Progeria Syndrome. Int. J. Mol. Sci. 2021, 22, 7327. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147327
Fafián-Labora JA, Morente-López M, de Toro FJ, Arufe MC. High-Throughput Screen Detects Calcium Signaling Dysfunction in Hutchinson-Gilford Progeria Syndrome. International Journal of Molecular Sciences. 2021; 22(14):7327. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147327
Chicago/Turabian StyleFafián-Labora, Juan A., Miriam Morente-López, Fco. Javier de Toro, and María C. Arufe. 2021. "High-Throughput Screen Detects Calcium Signaling Dysfunction in Hutchinson-Gilford Progeria Syndrome" International Journal of Molecular Sciences 22, no. 14: 7327. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147327