
Symptoms of Prenatal Depression Associated with Shorter Telomeres in Female Placenta

 and
and
Abstract
:1. Introduction
2. Methods
2.1. Cohort
2.2. Materials
2.2.1. Maternal Demographics and Birth Outcomes
2.2.2. Maternal Depression and Anxiety Symptoms
2.2.3. Placental Biopsies
2.2.4. STELA
2.3. Statistical Analysis
2.3.1. Research Question
2.3.2. Hypothesis
2.3.3. Statistical Steps
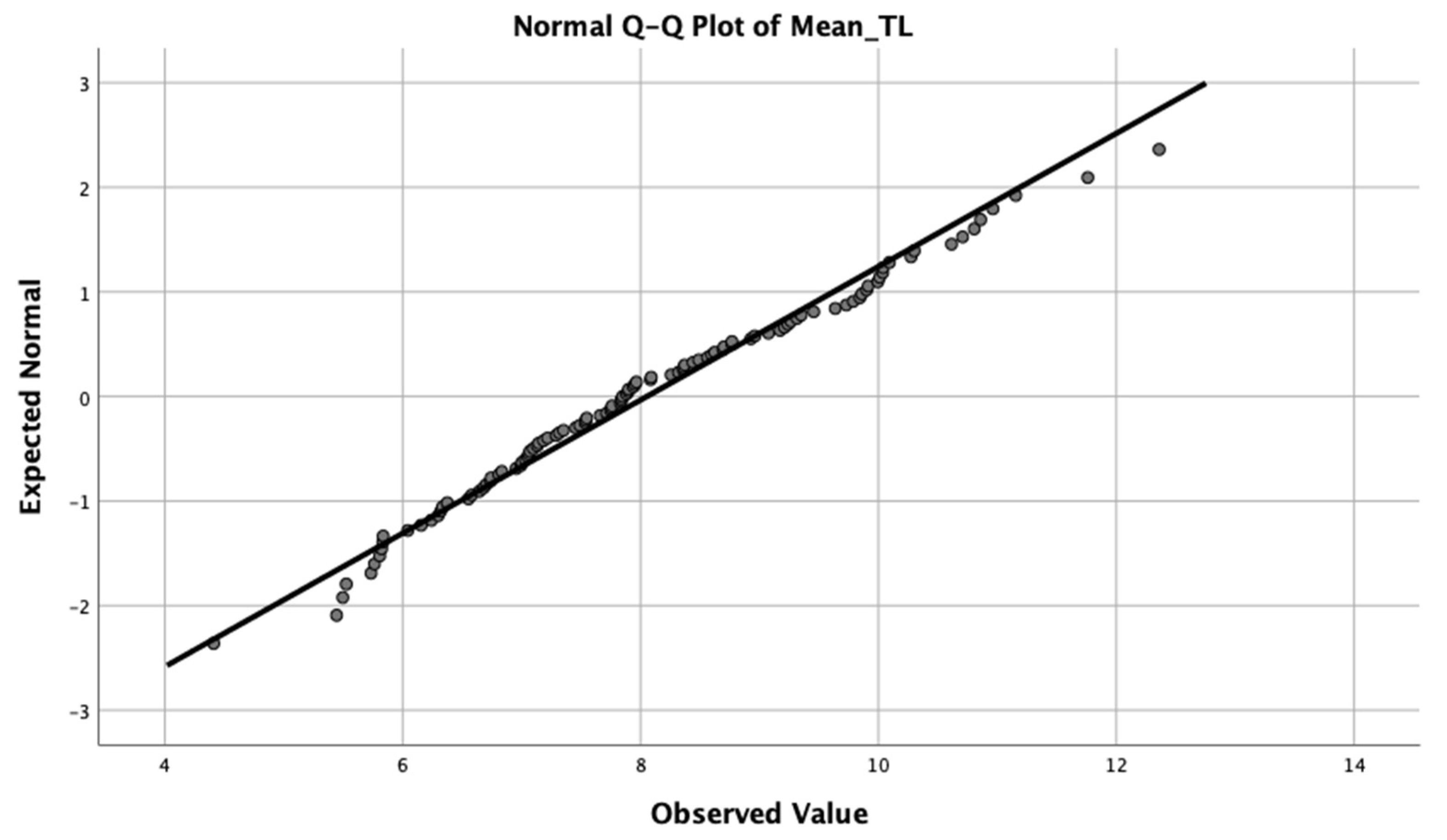
3. Results
3.1. Association between Telomere Length and Potential Confounders
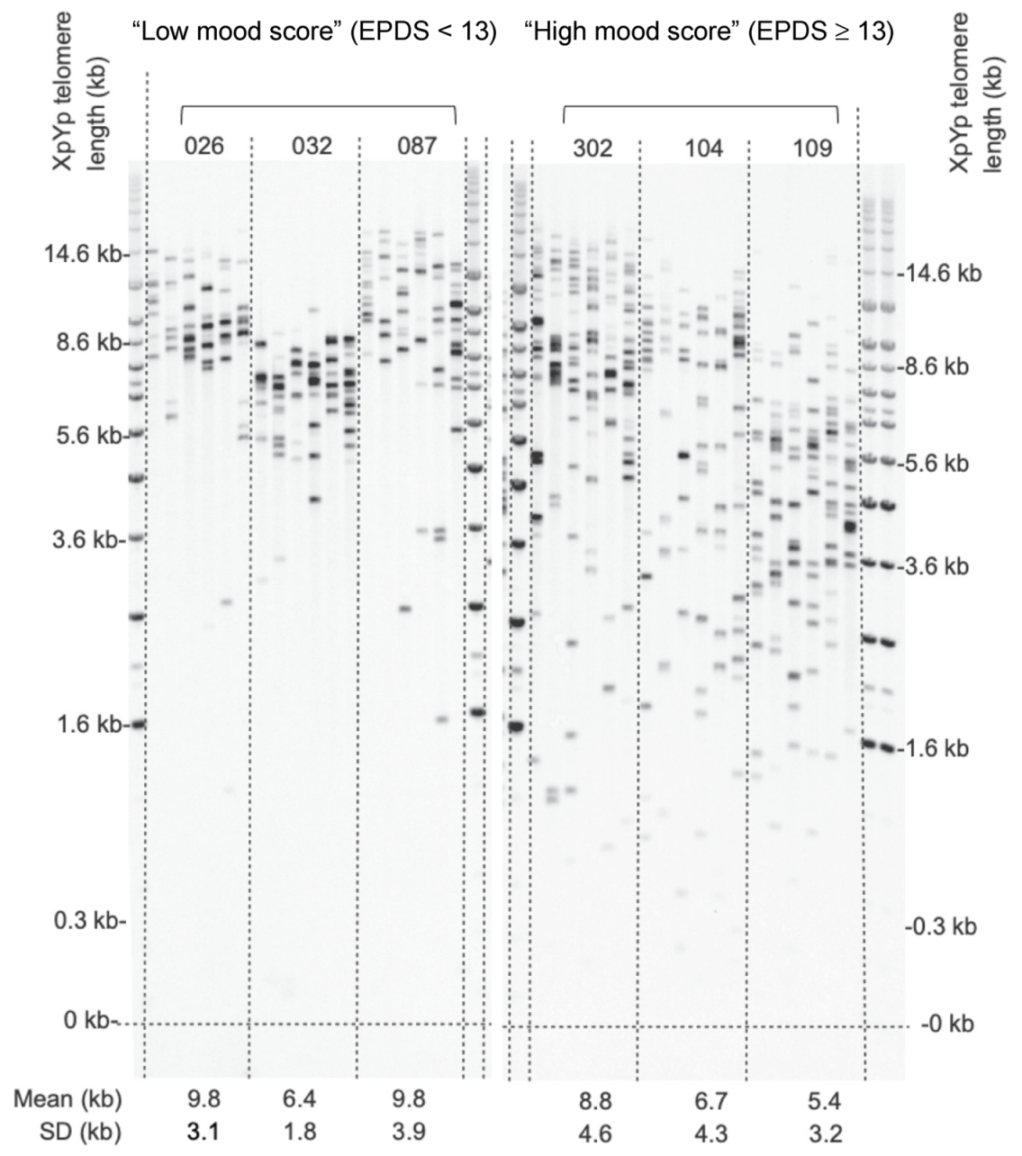
3.2. Analysis of High and Low Mood Score Groups
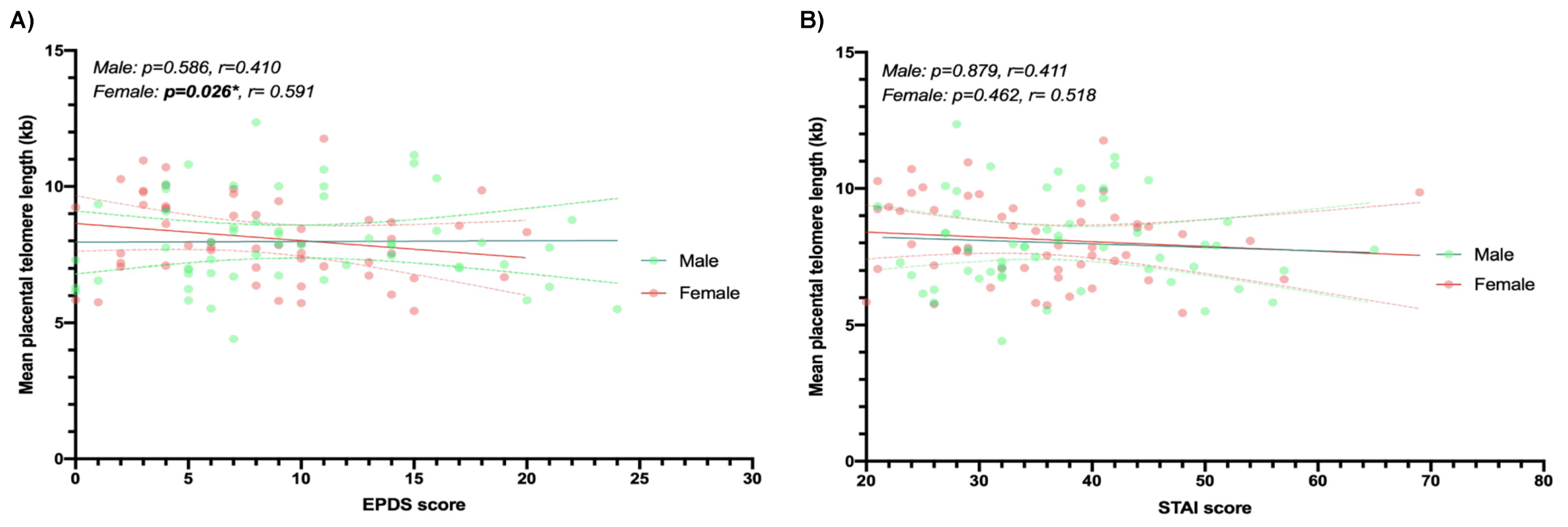
3.3. Multiple Linear Regression
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| STELA | Single telomere length analysis |
| GiW | Grown in Wales |
| ELCS | Elective caesarean section |
| WIMD | Welsh Index of Multiple Deprivation |
| EPDS | Edinburgh Postnatal Depression Scale |
| STAI | State Trait Anxiety Index |
| BMI | Body mass index |
References
- Van de Velde, S.; Bracke, P.; Levecque, K. Gender differences in depression in 23 European countries. Cross-national variation in the gender gap in depression. Soc. Sci. Med. 2010, 71, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.E. Clinical practice. Depression during pregnancy. N. Engl. J. Med. 2011, 365, 1605–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, S.R.; Brennan, P.A. Impact of antenatal and postpartum maternal mental illness: How are the children? Clin Obs. Gynecol 2009, 52, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, M.T.; Monk, C. Impact of maternal stress, depression and anxiety on fetal neurobehavioral development. Clin. Obs. Gynecol. 2009, 52, 425–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, L.; Ryan, E.; Trevillion, K.; Anderson, F.; Bick, D.; Bye, A.; Pickles, A. Accuracy of the Whooley questions and the Edinburgh Postnatal Depression Scale in identifying depression and other mental disorders in early pregnancy. Br. J. Psychiatry 2018, 212, 50–56. [Google Scholar] [CrossRef]
- Janssen, A.B.; Savory, K.A.; Garay, S.M.; Sumption, L.; Watkins, W.; Garcia-Martin, I.; Savory, N.A.; Ridgway, A.; Isles, A.R.; Penketh, R.; et al. Persistence of anxiety symptoms after elective caesarean delivery. Bjpsych Open 2018, 4, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.M.; Carnegie, R.E.; Cree, C.; Rollings, C.; Rena-Jones, L.; Evans, J.; Stein, A.; Tilling, K.; Lewcock, M.; Lawlor, D.A. Prevalence of Prenatal Depression Symptoms Among 2 Generations of Pregnant Mothers: The Avon Longitudinal Study of Parents and Children. Jama Netw. Open 2018, 1, e180725. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.; Parsonage, M.; Knapp, M.; Iemmi, V.; Adelaja, B. Costs of perinatal mental health problems. J. Affect. Disord. 2014, 192, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Biaggi, A.; Conroy, S.; Pawlby, S.; Pariante, C.M. Identifying the women at risk of antenatal anxiety and depression: A systematic review. J. Affect. Disord. 2016, 191, 62–77. [Google Scholar] [CrossRef] [Green Version]
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar] [CrossRef] [Green Version]
- De Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [Green Version]
- Takubo, K.; Izumiyama-Shimomura, N.; Honma, N.; Sawabe, M.; Arai, T.; Kato, M.; Oshimura, M.; Nakamura, K. Telomere lengths are characteristic in each human individual. Exp. Gerontol. 2002, 37, 523–531. [Google Scholar] [CrossRef]
- Baird, D.M.; Britt-Compton, B.; Rowson, J.; Amso, N.N.; Gregory, L.; Kipling, D. Telomere instability in the male germline. Hum. Mol. Genet. 2006, 15, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Baird, D.M.; Rowson, J.; Wynford-Thomas, D.; Kipling, D. Extensive allelic variation and ultrashort telomeres in senescent human cells. Nat. Genet. 2003, 33, 203–207. [Google Scholar] [CrossRef]
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Collins, K.; Mitchell, J.R. Telomerase in the human organism. Oncogene 2002, 21, 564–579. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J. Senescent cells, tumor suppression, and organismal aging: Good citizens, bad neighbors. Cell 2005, 120, 513–522. [Google Scholar] [CrossRef]
- Jylhava, J.; Pedersen, N.L.; Hagg, S. Biological Age Predictors. EBioMedicine 2017, 21, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, K.; Bardeguez, A.; Gardner, J.P.; Rodriguez, P.; Ganesh, V.; Kimura, M.; Skurnick, J.; Awad, G.; Aviv, A. Telomere length in the newborn. Pediatr. Res. 2002, 52, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Gielen, M.; Hageman, G.; Pachen, D.; Derom, C.; Vlietinck, R.; Zeegers, M.P. Placental telomere length decreases with gestational age and is influenced by parity: A study of third trimester live-born twins. Placenta 2014, 35, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.L.; Liu, Y.; Robinson, W.P. Placental telomere length decline with gestational age differs by sex and TERT, DNMT1, and DNMT3A DNA methylation. Placenta 2016, 48, 26–33. [Google Scholar] [CrossRef]
- Whiteman, V.E.; Goswami, A.; Salihu, H.M. Telomere length and fetal programming: A review of recent scientific advances. Am. J. Reprod. Immunol. 2017, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, N.; Boehner, M.; Groenen, F.; Kalb, R. Telomere length of patients with major depression is shortened but independent from therapy and severity of the disease. Depress Anxiety 2010, 27, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Simon, N.M.; Smoller, J.W.; McNamara, K.L.; Maser, R.S.; Zalta, A.K.; Pollack, M.H.; Nierenberg, A.A.; Fava, M.; Wong, K.K. Telomere shortening and mood disorders: Preliminary support for a chronic stress model of accelerated aging. Biol. Psychiatry 2006, 60, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Bosquet Enlow, M.; Bollati, V.; Sideridis, G.; Flom, J.D.; Hoxha, M.; Hacker, M.R.; Wright, R.J. Sex differences in effects of maternal risk and protective factors in childhood and pregnancy on newborn telomere length. Psychoneuroendocrinology 2018, 95, 74–85. [Google Scholar] [CrossRef]
- John, R.; Hemberger, M. A placenta for life. Reprod Biomed Online 2012, 25, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Izutsu, T.; Kudo, T.; Sato, T.; Nishiya, I.; Ohyashiki, K.; Mori, M.; Nakagawara, K. Telomerase activity in human chorionic villi and placenta determined by TRAP and in situ TRAP assay. Placenta 1998, 19, 613–618. [Google Scholar] [CrossRef]
- Biron-Shental, T.; Kidron, D.; Sukenik-Halevy, R.; Goldberg-Bittman, L.; Sharony, R.; Fejgin, M.D.; Amiel, A. TERC telomerase subunit gene copy number in placentas from pregnancies complicated with intrauterine growth restriction. Early Hum. Dev. 2011, 87, 73–75. [Google Scholar] [CrossRef]
- Welsh Index of Multiple Deprivation. Available online: https://gov.wales/welsh-index-multiple-deprivation (accessed on 3 January 2021).
- Cox, J.L.; Holden, J.M.; Sagovsky, R. Detection of postnatal depression. Development of the 10-item Edinburgh Postnatal Depression Scale. Br. J. Psychiatry J. Ment. Sci. 1987, 150, 782–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, K.A.; McMahon, C.; Austin, M.P. Maternal anxiety during the transition to parenthood: A prospective study. J. Affect. Disord. 2008, 108, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Meades, R.; Ayers, S. Anxiety measures validated in perinatal populations: A systematic review. J. Affect. Disord. 2011, 133, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Martin, I.; Janssen, A.B.; Jones, R.E.; Grimstead, J.W.; Penketh, R.J.A.; Baird, D.M.; John, R.M. Telomere length heterogeneity in placenta revealed with high-resolution telomere length analysis. Placenta 2017, 59, 61–68. [Google Scholar] [CrossRef]
- Hogan, B.; Beddington, R.; Constantini., F.; Lacy, E. Manipulating the Mouse Embryo: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1994. [Google Scholar]
- Garcia-Martin, I.; Penketh, R.J.A.; Janssen, A.B.; Jones, R.E.; Grimstead, J.; Baird, D.M.; John, R.M. Metformin and insulin treatment prevent placental telomere attrition in boys exposed to maternal diabetes. PLoS ONE 2018, 13, e0208533. [Google Scholar] [CrossRef]
- Sumption, L.A.; Garay, S.M.; John, R.M. Low serum placental lactogen at term is associated with postnatal symptoms of depression and anxiety in women delivering female infants. Psychoneuroendocrinology 2020, 116, 104655. [Google Scholar] [CrossRef] [PubMed]
- Dingsdale, H.; Nan, X.; Garay, S.M.; Mueller, A.; Sumption, L.A.; Chacon-Fernandez, P.; Martinez-Garay, I.; Ghevaert, C.; Barde, Y.A.; John, R.M. The placenta protects the fetal circulation from anxiety-driven elevations in maternal serum levels of brain-derived neurotrophic factor. Transl. Psychiatry 2021, 11, 62. [Google Scholar] [CrossRef]
- Hjort, L.; Vryer, R.; Grunnet, L.G.; Burgner, D.; Olsen, S.F.; Saffery, R.; Vaag, A. Telomere length is reduced in 9- to 16-year-old girls exposed to gestational diabetes in utero. Diabetologia 2018, 61, 870–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davy, P.; Nagata, M.; Bullard, P.; Fogelson, N.S.; Allsopp, R. Fetal growth restriction is associated with accelerated telomere shortening and increased expression of cell senescence markers in the placenta. Placenta 2009, 30, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biron-Shental, T.; Sukenik Halevy, R.; Goldberg-Bittman, L.; Kidron, D.; Fejgin, M.D.; Amiel, A. Telomeres are shorter in placental trophoblasts of pregnancies complicated with intrauterine growth restriction (IUGR). Early Hum. Dev. 2010, 86, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Toutain, J.; Prochazkova-Carlotti, M.; Cappellen, D.; Jarne, A.; Chevret, E.; Ferrer, J.; Idrissi, Y.; Pelluard, F.; Carles, D.; Maugey-Laulon, B.; et al. Reduced placental telomere length during pregnancies complicated by intrauterine growth restriction. PLoS ONE 2013, 8, e54013. [Google Scholar] [CrossRef] [Green Version]
- Paules, C.; Dantas, A.P.; Miranda, J.; Crovetto, F.; Eixarch, E.; Rodriguez-Sureda, V.; Dominguez, C.; Casu, G.; Rovira, C.; Nadal, A.; et al. Premature placental aging in term small-for-gestational-age and growth-restricted fetuses. Ultrasound Obs. Gynecol. 2019, 53, 615–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Goldberg-Bittman, L.; Kidron, D.; Fejgin, M.D.; Amiel, A. Short telomeres may play a role in placental dysfunction in preeclampsia and intrauterine growth restriction. Am. J. Obs. Gynecol. 2010, 202, 381.e1–381.e7. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.W.; Esteves, K.C.; Gray, S.A.O.; Clarke, T.N.; Callerame, K.; Theall, K.P.; Drury, S.S. The transgenerational transmission of maternal adverse childhood experiences (ACEs): Insights from placental aging and infant autonomic nervous system reactivity. Psychoneuroendocrinology 2019, 106, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Matthey, S.; Henshaw, C.; Elliott, S.; Barnett, B. Variability in use of cut-off scores and formats on the Edinburgh Postnatal Depression Scale: Implications for clinical and research practice. Arch. Women’s Ment. Health 2006, 9, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Murray, L.; Carothers, A.D. The validation of the Edinburgh Post-natal Depression Scale on a community sample. Br. J. Psychiatry J. Ment. Sci. 1990, 157, 288–290. [Google Scholar] [CrossRef]
- Entringer, S.; Epel, E.S.; Lin, J.; Buss, C.; Shahbaba, B.; Blackburn, E.H.; Simhan, H.N.; Wadhwa, P.D. Maternal psychosocial stress during pregnancy is associated with newborn leukocyte telomere length. Am. J. Obs. Gynecol. 2013, 208, 134.e1–134.e7. [Google Scholar] [CrossRef] [Green Version]
- Marchetto, N.M.; Glynn, R.A.; Ferry, M.L.; Ostojic, M.; Wolff, S.M.; Yao, R.; Haussmann, M.F. Prenatal stress and newborn telomere length. Am. J. Obs. Gynecol. 2016, 215, 94. [Google Scholar] [CrossRef] [Green Version]
- Send, T.S.; Gilles, M.; Codd, V.; Wolf, I.; Bardtke, S.; Streit, F.; Strohmaier, J.; Frank, J.; Schendel, D.; Sutterlin, M.W.; et al. Telomere Length in Newborns is Related to Maternal Stress During Pregnancy. Neuropsychopharmacology 2017, 42, 2407–2413. [Google Scholar] [CrossRef] [PubMed]
- Entringer, S.; Epel, E.S.; Kumsta, R.; Lin, J.; Hellhammer, D.H.; Blackburn, E.H.; Wust, S.; Wadhwa, P.D. Stress exposure in intrauterine life is associated with shorter telomere length in young adulthood. Proc. Natl. Acad. Sci. USA 2011, 108, E513–E518. [Google Scholar] [CrossRef] [Green Version]
- Rosa, M.J.; Hsu, H.L.; Just, A.C.; Brennan, K.J.; Bloomquist, T.; Kloog, I.; Pantic, I.; Mercado Garcia, A.; Wilson, A.; Coull, B.A.; et al. Association between prenatal particulate air pollution exposure and telomere length in cord blood: Effect modification by fetal sex. Env. Res. 2019, 172, 495–501. [Google Scholar] [CrossRef]
- Bosquet Enlow, M.; Petty, C.R.; Hacker, M.R.; Burris, H.H. Maternal psychosocial functioning, obstetric health history, and newborn telomere length. Psychoneuroendocrinology 2021, 123, 105043. [Google Scholar] [CrossRef] [PubMed]
- Martens, D.S.; Janssen, B.G.; Bijnens, E.M.; Clemente, D.B.P.; Vineis, P.; Plusquin, M.; Nawrot, T.S. Association of Parental Socioeconomic Status and Newborn Telomere Length. Jama. Netw. Open 2020, 3, e204057. [Google Scholar] [CrossRef] [PubMed]
- Roger, L.; Jones, R.E.; Heppel, N.H.; Williams, G.T.; Sampson, J.R.; Baird, D.M. Extensive telomere erosion in the initiation of colorectal adenomas and its Assoc. with chromosomal instability. J. Natl. Cancer Inst. 2013, 105, 1202–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Potential Confounder | Association with Mean Telomere Length |
|---|---|
| Maternal age | r = 0.09 CI = −0.030, 0.078 p = 0.37 |
| Gestation age | r = 0.15 CI = −0.100, 0.832 p = 0.12 |
| Parity | r = 0.09 CI = −0.173, 0.479 p = 0.35 |
| Smoking 14/109 (12.9%) | r = −0.00 CI = −0.765, 1.023 p = 0.776 |
| Alcohol 8/109 (7.3%) | r = 0.11 CI = −0.421, 1.85 p = 0.214 |
| BMI | r = −0.03 CI = −0.056, 0.093 p = 0.792 |
| WIMD | r = 0.22 CI = 0.000, 0.001 p = 0.027 |
| Characteristics | Low Mood Score Group: EPDS < 13 Group (n = 79) | High Mood Score Group: EPDS ≥ 13 Group (n = 30) | p-Value |
|---|---|---|---|
| Caucasian ethnicity | 73 (92%) | 29 (96%) | 0.52 |
| Parity: | 0.82 | ||
| Primiparous | 14 (17.7%) | 7(23.3%) | |
| Multiparous | 65 (82.2%) | 23 (76.7%) | |
| Maternal age | 32 (5.51)/19–44 | 30 (5.68)/20–39 | 0.31 |
| ELCS | 79 (100%) | 30 (100%) | N/A |
| Birth weight (g) | 3491 (620)/2260–5080 | 3488 (499)/2460–5110 | 0.98 |
| Gestational age (weeks) | 39 (0.61)/38–41 | 38 (0.68)/37–41 | 0.12 |
| Placental weight (g) | 663 (129)/376–941 | 671 (150)/455–1060 | 0.76 |
| Fetal sex: | 0.42 | ||
| Female | 38 (48%) | 17 (56%) | |
| Male | 41 (52%) | 13 (44%) | |
| Smoking during pregnancy | 9 (11.4%) | 5 (16.7%) | 0.46 |
| Alcohol during pregnancy | 5 (6.3%) | 3 (10%) | 0.51 |
| Mood Scores | All | Male | Female | ||||||
|---|---|---|---|---|---|---|---|---|---|
| B | 95%CI | p | B | 95%CI | p | B | 95%CI | p | |
| Unadjusted linear regressions | |||||||||
| EPDS | −0.025 | −0.079, 0.028 | 0.346 | 0.002 | −0.073, 0.078 | 0.949 | −0.063 | −0.143, 0.017 | 0.121 |
| STAI | −0.016 | −0.046, 0.015 | 0.305 | −0.013 | −0.060, 0.034 | 0.582 | −0.017 | −0.059, 0.024 | 0.399 |
| Adjusted linear regressions | |||||||||
| EPDS | −0.004 | −0.061, 0.052 | 0.884 | 0.022 | −0.059, 0.103 | 0.586 | −0.098 | −0.184, −0.012 | 0.026 |
| STAI | −0.002 | −0.033, 0.030 | 0.914 | −0.004 | −0.054, 0.046 | 0.879 | −0.016 | −0.058, 0.027 | 0.462 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Martin, I.; Penketh, R.J.A.; Garay, S.M.; Jones, R.E.; Grimstead, J.W.; Baird, D.M.; John, R.M. Symptoms of Prenatal Depression Associated with Shorter Telomeres in Female Placenta. Int. J. Mol. Sci. 2021, 22, 7458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147458
Garcia-Martin I, Penketh RJA, Garay SM, Jones RE, Grimstead JW, Baird DM, John RM. Symptoms of Prenatal Depression Associated with Shorter Telomeres in Female Placenta. International Journal of Molecular Sciences. 2021; 22(14):7458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147458
Chicago/Turabian StyleGarcia-Martin, Isabel, Richard J. A. Penketh, Samantha M. Garay, Rhiannon E. Jones, Julia W. Grimstead, Duncan M. Baird, and Rosalind M. John. 2021. "Symptoms of Prenatal Depression Associated with Shorter Telomeres in Female Placenta" International Journal of Molecular Sciences 22, no. 14: 7458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147458