
Oxytocin Signaling Acts as a Marker for Environmental Stressors in Zebrafish


Department of Life Science, National Taiwan University, Taipei 10617, Taiwan
*
Author to whom correspondence should be addressed.
†
H.-J. Chuang and C.-Y. Chang contributed equally to this study.
Int. J. Mol. Sci. 2021, 22(14), 7459; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147459
Submission received: 25 May 2021
/
Revised: 24 June 2021
/
Accepted: 9 July 2021
/
Published: 12 July 2021
(This article belongs to the Special Issue Nonapeptide Hormones)
Abstract
:The oxytocin system plays a role in stress responses and behavior modulation. However, the effects of oxytocin signaling on stress adaptation remain unclear. Here, we demonstrated the roles of oxytocin signaling as a biomarker under stress conditions in the peripheral tissues (the gills) and central nervous system (the brain). All the environmental stressors downregulated the expression of oxytocin receptors in the gills, and the alteration of the expression of oxytocin receptors was also found in the brain after the acidic (AC) and high-ammonia (HA) treatments. The number of oxytocin neurons was increased after double-deionized (DI) treatment. By transgenic line, Tg(oxtl:EGFP), we also investigated the projections of oxytocin neurons and found oxytocin axon innervations in various nuclei that might regulate the anxiety levels and aggressiveness of adult zebrafish under different environmental stresses. The oxytocin system integrates physiological responses and behavioral outcomes to ensure environmental adaptation in adult zebrafish. Our study provides insight into oxytocin signaling as a stress indicator upon environmental stressors.
1. Introduction
Animals adapt to the external environment throughout their life. Environmental stimuli can be classified as biotic or abiotic. The stimuli that disturb the internal dynamic equilibrium are defined as stressors [1,2,3], and they include pH, nitrogenous waste, salinity, temperature, anoxia, pollutants, and predation [4,5,6,7,8,9,10,11,12]. Stress responses are conserved in vertebrates and are divided into three phases [13,14]. To cope with stress, in animals, the neuroendocrine system is first activated to secrete hormones that alter physiological and metabolic pathways. Subsequently, systemic changes occur, which may reverse the negative impact of stressors.
The hypothalamus plays a primary role in stress responses; it integrates sensory inputs, controls hormone secretion, and regulates various functions, including the readjustment of biological activity and energy allocation [3,15,16]. Many hormones have been reported to be involved in stress responses [17,18,19]. Among them, cortisol, which is regulated by the hypothalamic–pituitary–adrenal (HPA) axis, is the most studied stress indicator, and it promotes “fight and flight” abilities in fish [2,20]. In zebrafish, the knockdown of glucocorticoid receptor (GR) by morpholino (MO) downregulated the expression of acid-secreting transporters and acid secretion, which decreased the ability of the zebrafish to cope with acidic environments [17]. Intraperitoneal injection with cortisol upregulated the expression and activity of Na-K-ATPase in the gills of milkfish under hypotonic stress [21]. However, the cortisol concentration in the blood is not always increased during stress. The cortisol level in serum was decreased in yellow perch treated with multiple stressors, including chemical pollutants and capture stress [22]. In sea bream, only 80 µg/L gold nanoparticles coated with PVP (AuNP-PVP) upregulated cortisol levels in the plasma, and 4 and 1600 µg/L AuNP-PVP did not change plasma cortisol levels [23]. Furthermore, some moderate stressors are insufficiently strong to trigger cortisol release. Compared with 25 mg/L aluminum oxide (Al2O3), 1 and 5 mg/L Al2O3 did not change cortisol levels in the blood of tilapia [24]. To date, several hypothalamic hormones have been demonstrated to participate in stress responses [25].
Oxytocin is a polypeptide neurotransmitter/hormone that mediates uterus construction and milk ejection [26,27,28]. Recent studies have shown that hypothalamic oxytocin is involved in stress responses. In rodents, forced swimming stress and shaker stress induced the parallel release of oxytocin in the hypothalamus and peripheral circulation [29,30]. Oxytocin-knockout mice displayed anxiety-like behaviors and an increased corticosterone level in the blood after psychogenic stressor on a platform shaker or in novel environments [31]. In patients with generalized social anxiety disorder, higher plasma oxytocin levels led to higher social anxiety and dissatisfaction [32]. Furthermore, recent studies have clarified the regulatory role of oxytocin on glucose homeostasis. Glucose levels were elevated in the blood of oxytocin-knockout mice fed a high fat diet [33]. Oxytocin attenuated the peak value of plasma glucose in men after intranasal administration [34]. The oxytocin receptor is expressed in the amygdala [35], and this receptor mediates an increase in oxytocin release in stimulated fear-attenuating responses, which may help individuals better respond to urgent situations [36]. Oxytocin is present in the hypothalamus and is widely present in various tissues [37,38]; furthermore, it plays roles in stress responses [39]. The oxytocin system (oxytocin and oxytocin receptors) may serve as an indicator for evaluating whether the individual is under stress conditions.
Because of aquatic habitats, fish possess high tolerance and sensitivity to harsh environments compared with terrestrial organisms [14,40], making fish popular experimental animals for studies of environmental acclimation [12,41,42]. For example, the blood glucose level was increased in Labeo capensis (mudfish) and Micropterus salmoides (largemouth bass) after exposure to high-ammonia stress [43,44]. Cortisol concentrations in the blood were increased under hypo- or hyper-osmotic stress in milkfish and tilapia [21,45]. However, the roles of oxytocin in stress responses and the underlying mechanisms in fish are unclear; more data are required to elucidate this research gap.
To address this research gap, we used zebrafish (Danio rerio) as the animal model to investigate the roles of the oxytocin system under different environmental challenges. Previous studies indicated that the oxytocin controlled the social behaviors in the adult zebrafish [46,47]. However, the role of oxytocin in stress response is still unknown. Zebrafish possesses similar physiological responses compared to mammals/humans [48]. A small size, rapid lifecycle, and ability to be easily genetically manipulated make zebrafish convenient for many experimental designs. Various molecular physiological approaches can be applied to study the functions of zebrafish genes and cells, and many behavioral paradigms can be used to evaluate the emotional states and social interactions of adult zebrafish [49,50]. The main goal of the present study was to evaluate the potential of oxytocin system as a stress indicator and investigate the role of oxytocin in stress responses in the zebrafish. In present study, we exposed adult zebrafish to various environmental stresses and measured the expression of oxytocin receptors in the gills and brain. We investigated the numbers of oxytocin neurons in the brain under different environmental stresses. We also traced the projections of oxytocin neurons in the brain and performed behavioral tests to evaluate behavioral alteration under different environmental challenges. Hence, that the oxytocin system is a potential biomarker of the stress response in aquatic teleosts is an expected result of this study.
2. Results
2.1. Oxyr and Oxyrl Were Expressed in the Brains and Gills of Adult Zebrafish
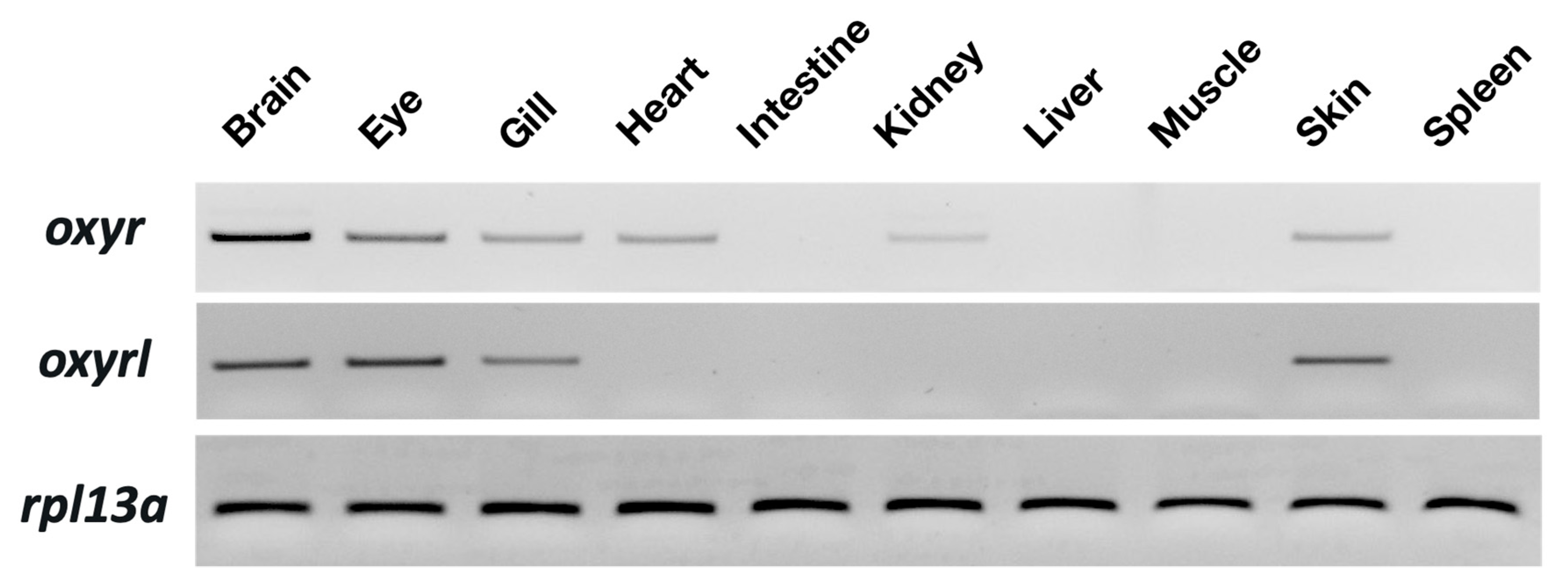
Tissue distribution of oxyr and oxyrl in the zebrafish was examined through semi-quantitative PCR with 40 amplification cycles. Rpl13a was used as an internal control to confirm the equal abundance of RNA in various tissues. Oxyr showed the highest expression in the brain and moderate expression in the eyes, gills, hearts, intestines, and skin of the adult zebrafish (Figure 1). In the kidneys, a weak signal of oxry was detected. For oxyrl, the expression in the eyes and brains was the highest, and moderate signals were found in the gills and skin of the adult zebrafish (Figure 1).
2.2. Environmental Stressors Downregulated Oxyr mRNA Expression in the Gills
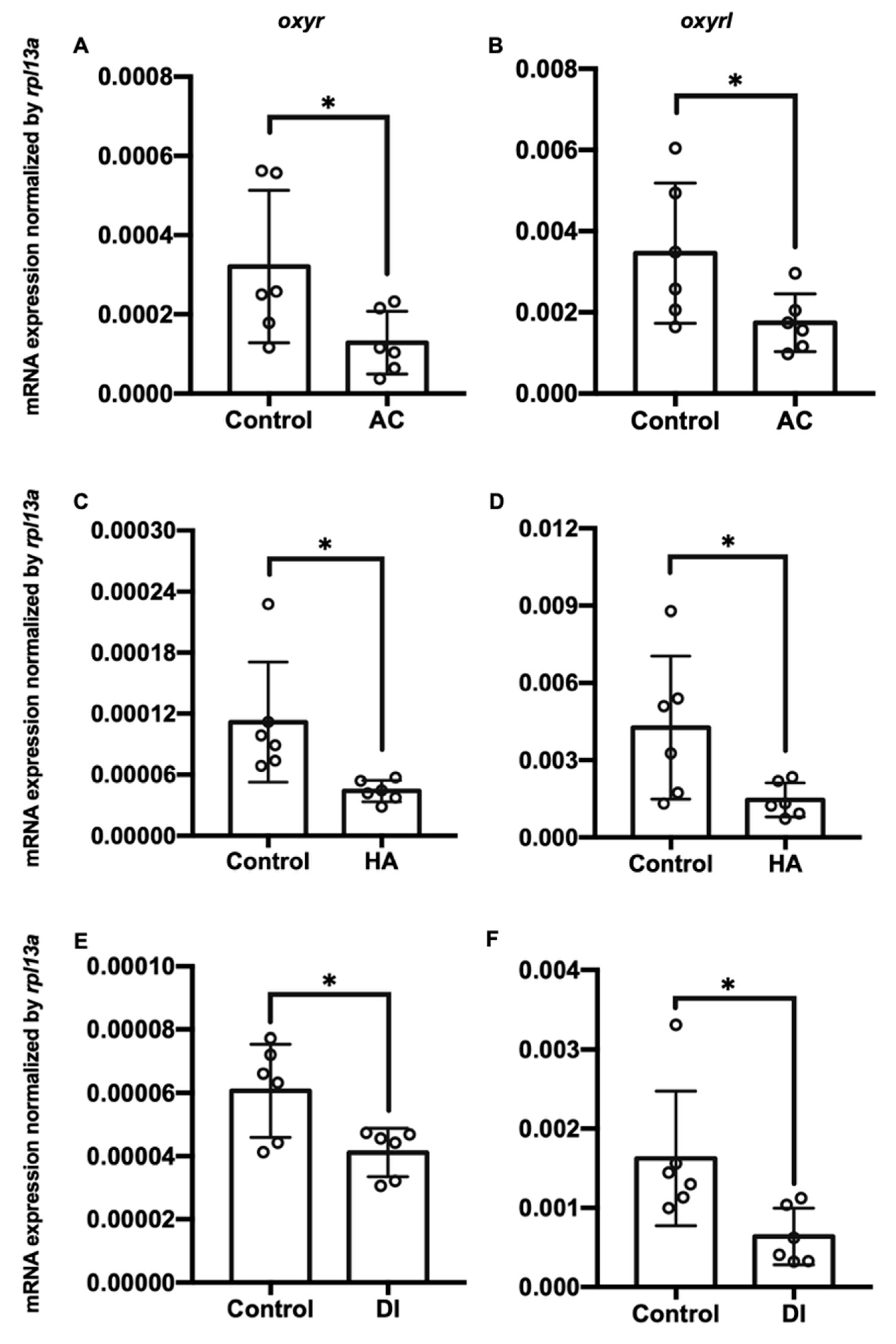
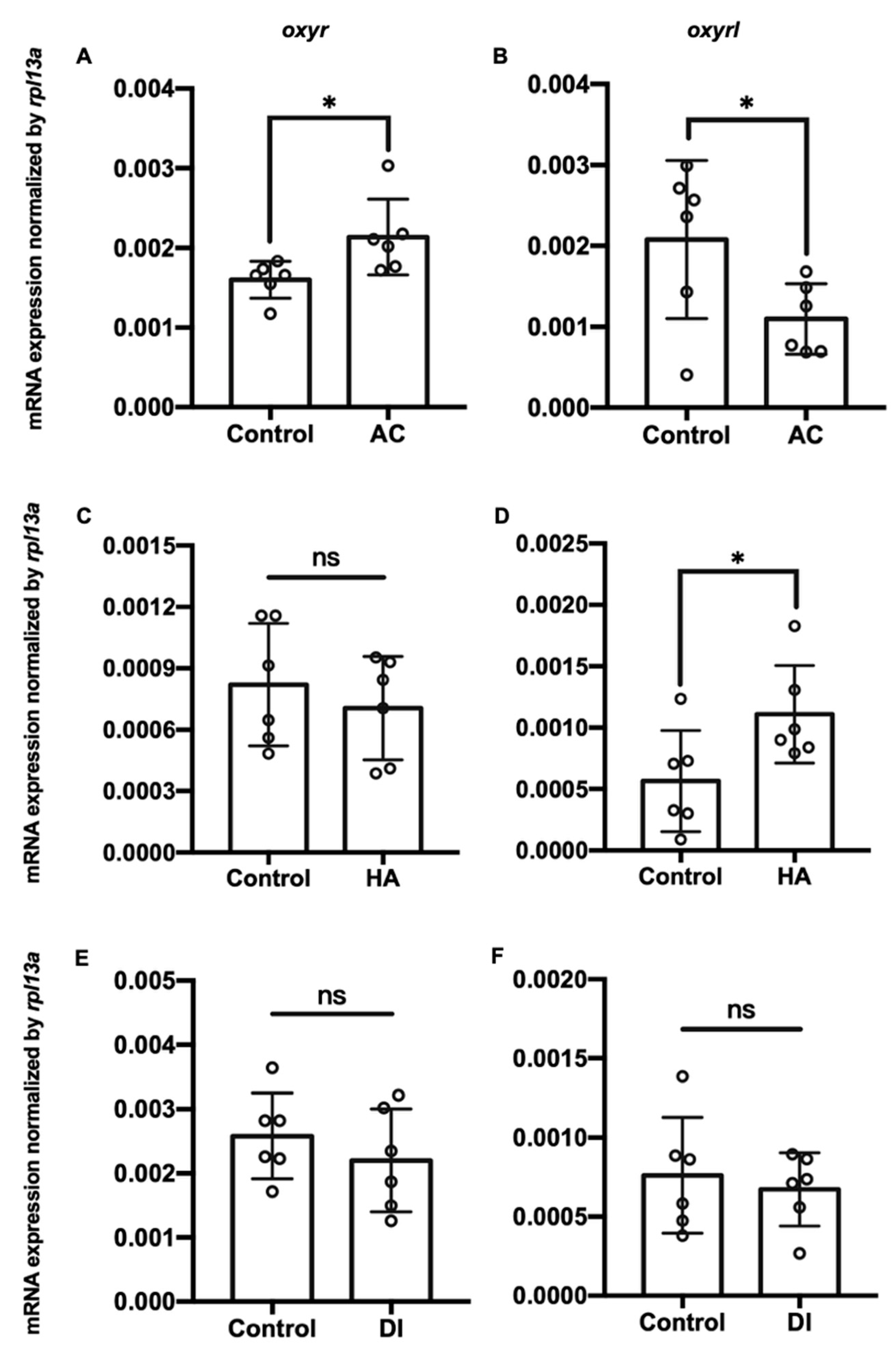
Three environmental challenges, namely acidic (AC), high-ammonia (HA), and double-deionized (DI) water, were applied to investigate the effects of environmental fluctuations on oxyr and oxyrl expression in the adult zebrafish. After the exposure of the zebrafish to these challenges, their gills and brains were dissected, and the total RNA of these two tissues was extracted for RT-qPCR analysis. AC, HA, and DI treatments significantly downregulated the mRNA expression of oxyr and oxyrl in the gills (Figure 2). After exposure to AC water for 7 days, the mRNA expression of oxyr and oxyrl in the gills declined by approximately 54% and 66% compared with that in the control fish gills, respectively (Figure 2A,B); HA treatment reduced the expression of oxyr and oxyrl in the gills by 61% and 69% for 7 days, respectively (Figure 2C,D). DI treatment caused 32% and 63% reductions in oxyr and oxyrl mRNA expression in the gills, respectively, compared with that in the control fish gills (Figure 2E,F). In the brain, oxyr expression increased by 38% after AC water exposure for 7 days, whereas oxyrl expression decreased by 58% after AC treatment (Figure 3A,B). After exposure to HA water, oxyr expression in the brain was similar to that in the control fish brains, and HA treatment significantly upregulated oxyrl expression by 83% (Figure 3C,D). DI treatment did not affect the expression of oxyr and oxyrl in the brain (Figure 3E,F).
2.3. DI Water Elevated the Number of Oxytocin Neurons in the Brain
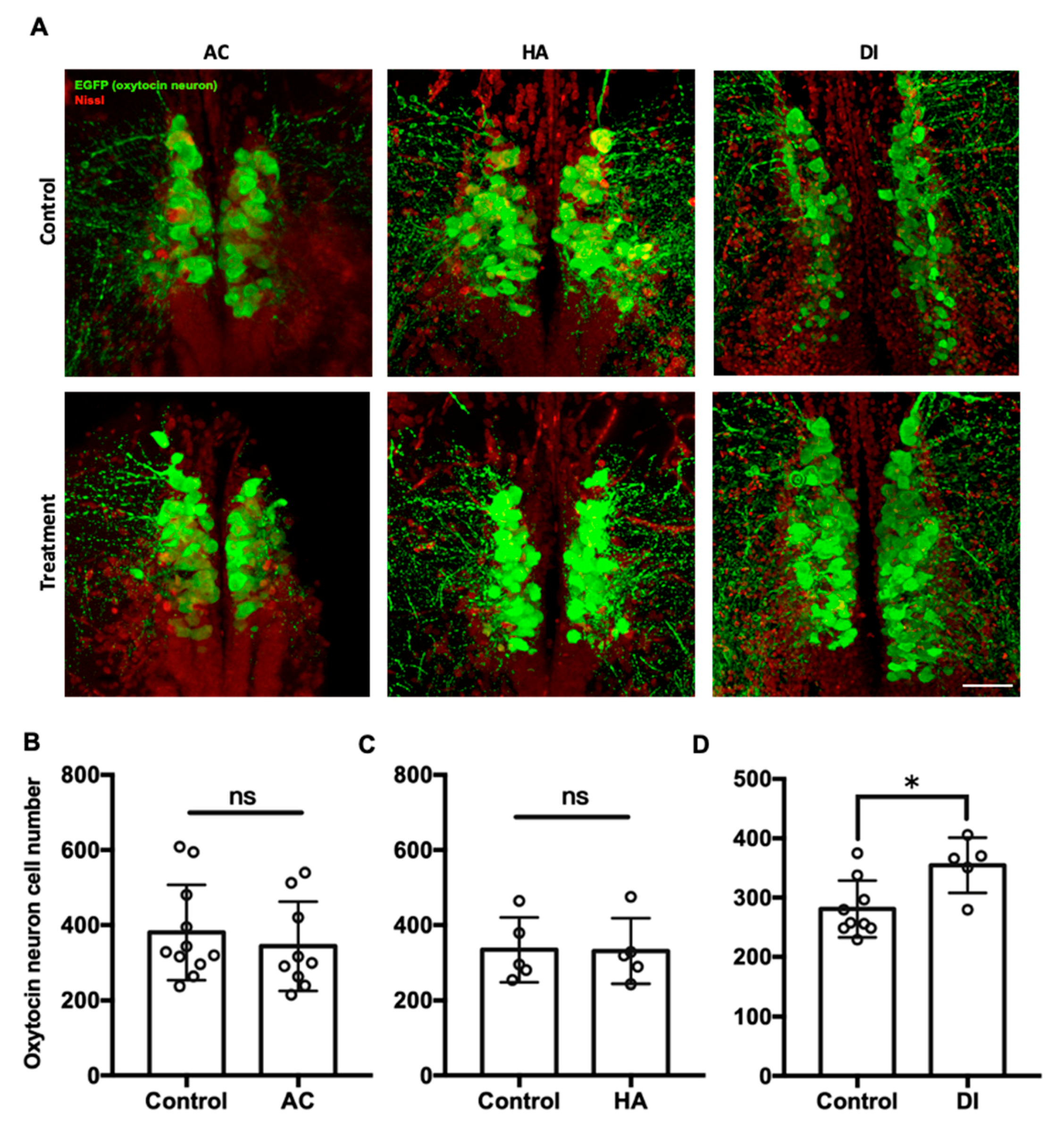
To investigate the effects of environmental stressors on the oxytocin system in the brain, the transgenic zebrafish Tg(oxtl:EGFP) were used, in which oxytocin neurons and their projections are labeled. In Figure 4, each circle represented the number of oxytocin neuron number in the brain of a single individual. After exposure to AC water for 7 days, the number of oxytocin neurons in the preoptic area of the brain was similar between the AC group (343.7 ± 118.6) and the control group (380.1 ± 127.2) (Figure 4B). Similarly, HA treatment did not affect the number of oxytocin neurons in the brain (HA group, 330.7 ± 87.3; control group, 334.2 ± 86.4) (Figure 4C). After DI treatment for 7 days, the number of oxytocin neurons in the DI group (476.6 ± 63.7) was significantly higher than that in the control group (378.3 ± 35.8) (Figure 4D).
2.4. Mapping Central Projections of Oxytocin Neurons in Adult Zebrafish
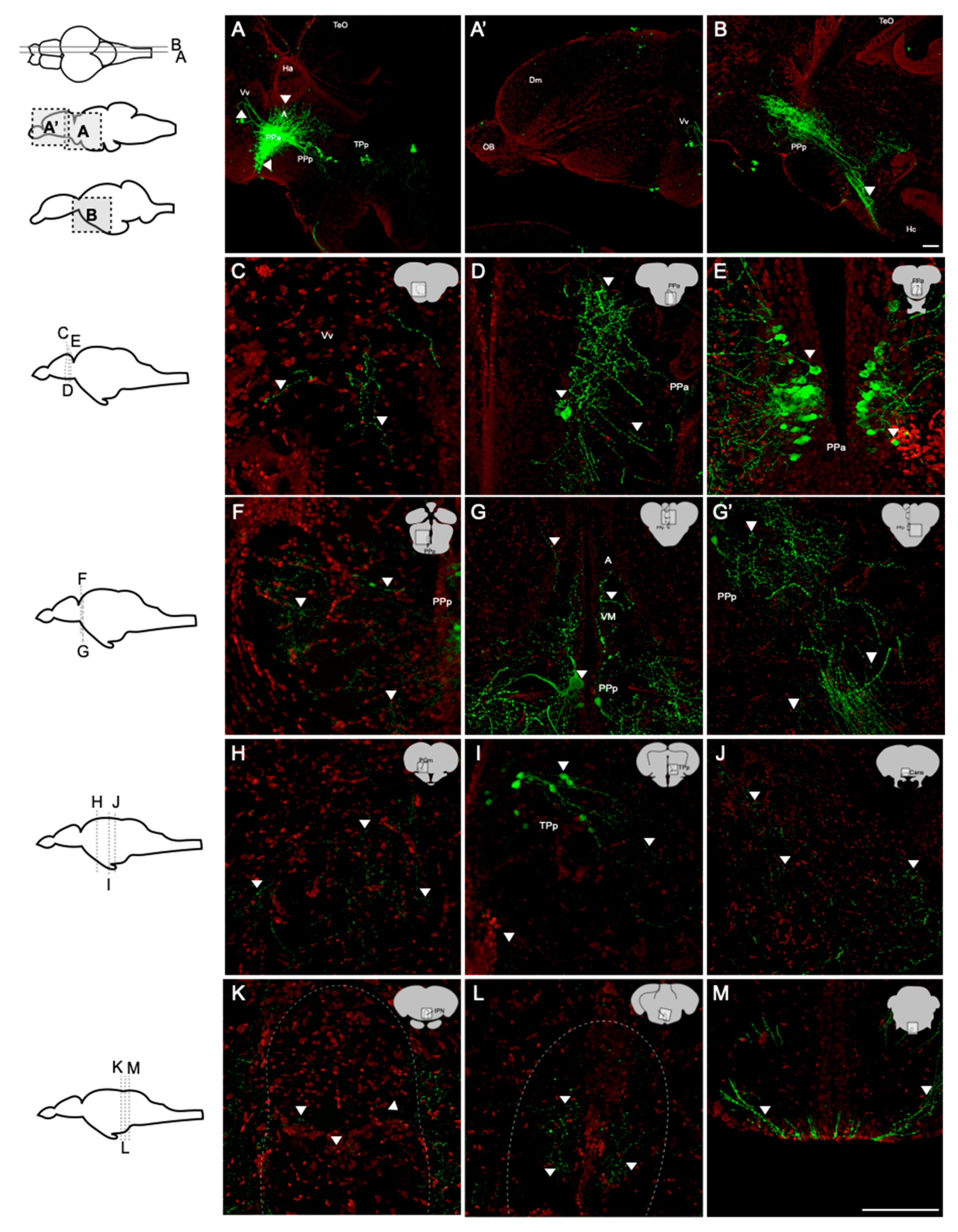
To investigate the projections of oxytocin neurons in the brain of the adult zebrafish, the whole-brain sections of a transgenic zebrafish line, Tg(oxtl:EGFP), was used to visualize the cell bodies of oxytocin neurons and their projections. Immunohistochemistry of GFP was performed to enhance green fluorescent signals. In the sagittal brain sections, the cell bodies of oxytocin neurons were mainly located in the parvocellular preoptic nucleus including the anterior part (PPa) and the posterior part (PPp) (Figure 5A,B). Few neurons were also found in the periventricular nucleus of the posterior tuberculum (TPp), and these neurons were smaller than those in the parvocellular preoptic nucleus (Figure 5A). Oxytocin neurons in the PPa and PPp sent a high density of axons toward the caudal zone of the periventricular hypothalamus (Hc) to the pituitary gland (Figure 5B). Oxytocin neurons also projected to the ventral nuclei of the ventral telencephalic area (Vv) (Figure 5A) but not to the olfactory bulb (Figure 5A). To observe the innervation of oxytocin neurons more clearly, we performed coronal sectioning using a vibratome. In the telencephalon, few fibers of oxytocin neurons were found in the Vv (Figure 5C), and many fibers were observed in the PPa (Figure 5D) as well as many oxytocin neuron cell bodies (Figure 5E). In the midbrain, few cell bodies were found in the PPp, and these neurons sent axons dorsally to the anterior and ventromedial thalamic nuclei (A and VM) (Figure 5F,G) and ventral-caudally to the pituitary (Figure 5G). Few cell bodies were also found in the TPp (Figure 5I). In the preglomerular nuclei (PGm), TPp, commissura ansulata (Cans), interpeduncular nucleus (IPN), and raphe, the signals of oxytocin axons were found (Figure 5H–L). In the hindbrain, oxytocinergic axons extended to the crossed tecto-bulbar tract (TTBc; Figure 5M). No cell body was found in the hindbrain.
2.5. Effects of Environmental Stressors on Swimming and Aggressive Behaviors
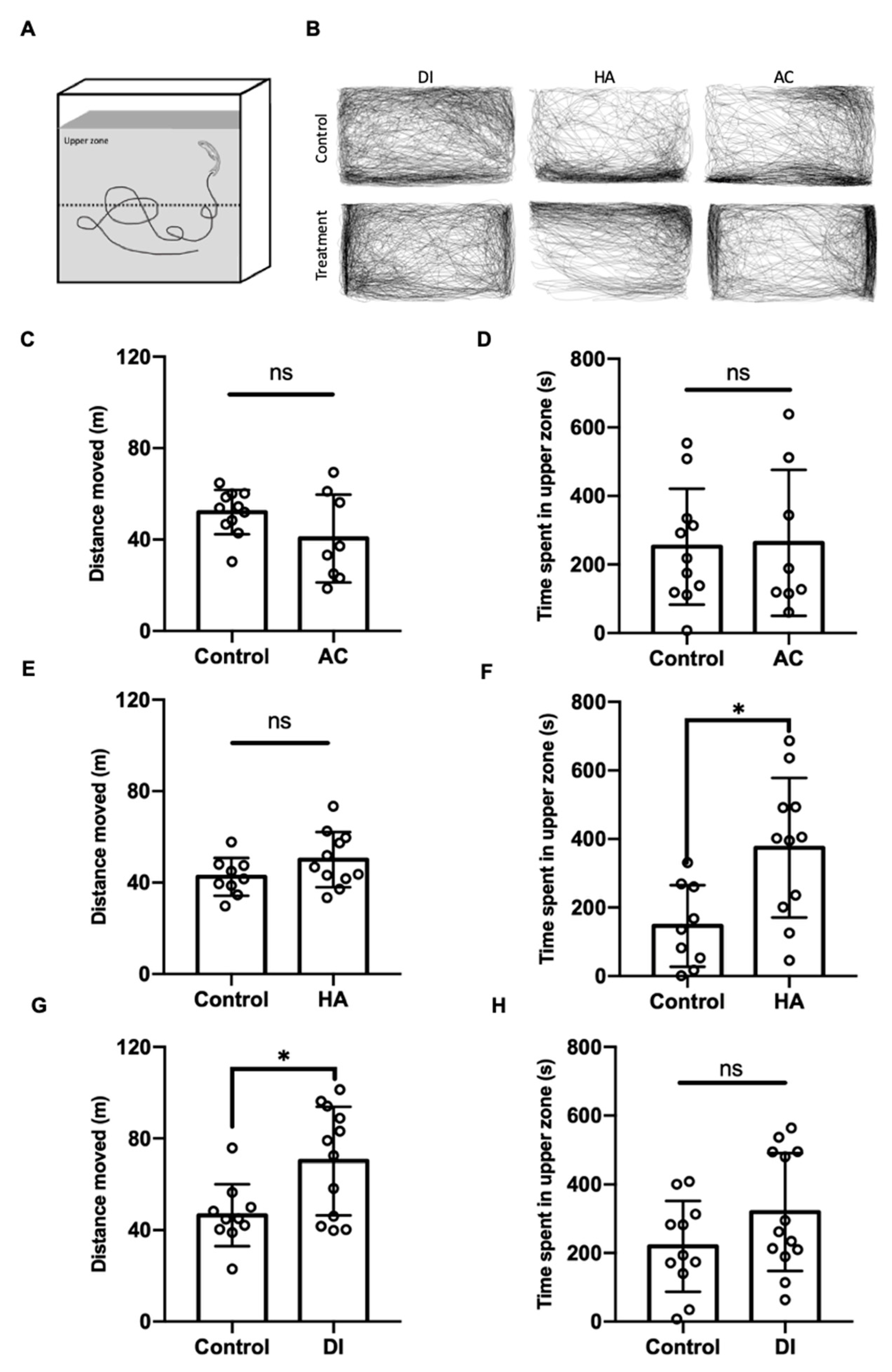
To evaluate the anxiety level of the adult zebrafish under various environmental stresses, the novel tank diving test was performed. After exposure to DI water for 24 h, the zebrafish’s total moving distance was significantly increased, but the time spent in the upper part of the tank was similar to that of the control fish (Figure 6G,H); by contrast, after exposure to HA water for 24 h, relative to the control fish, the fish spent more time in the upper part of tank but showed a similar moving distance (Figure 6E,F). AC treatment did not affect the anxiety level and locomotion activity of the fish (Figure 6C,D).
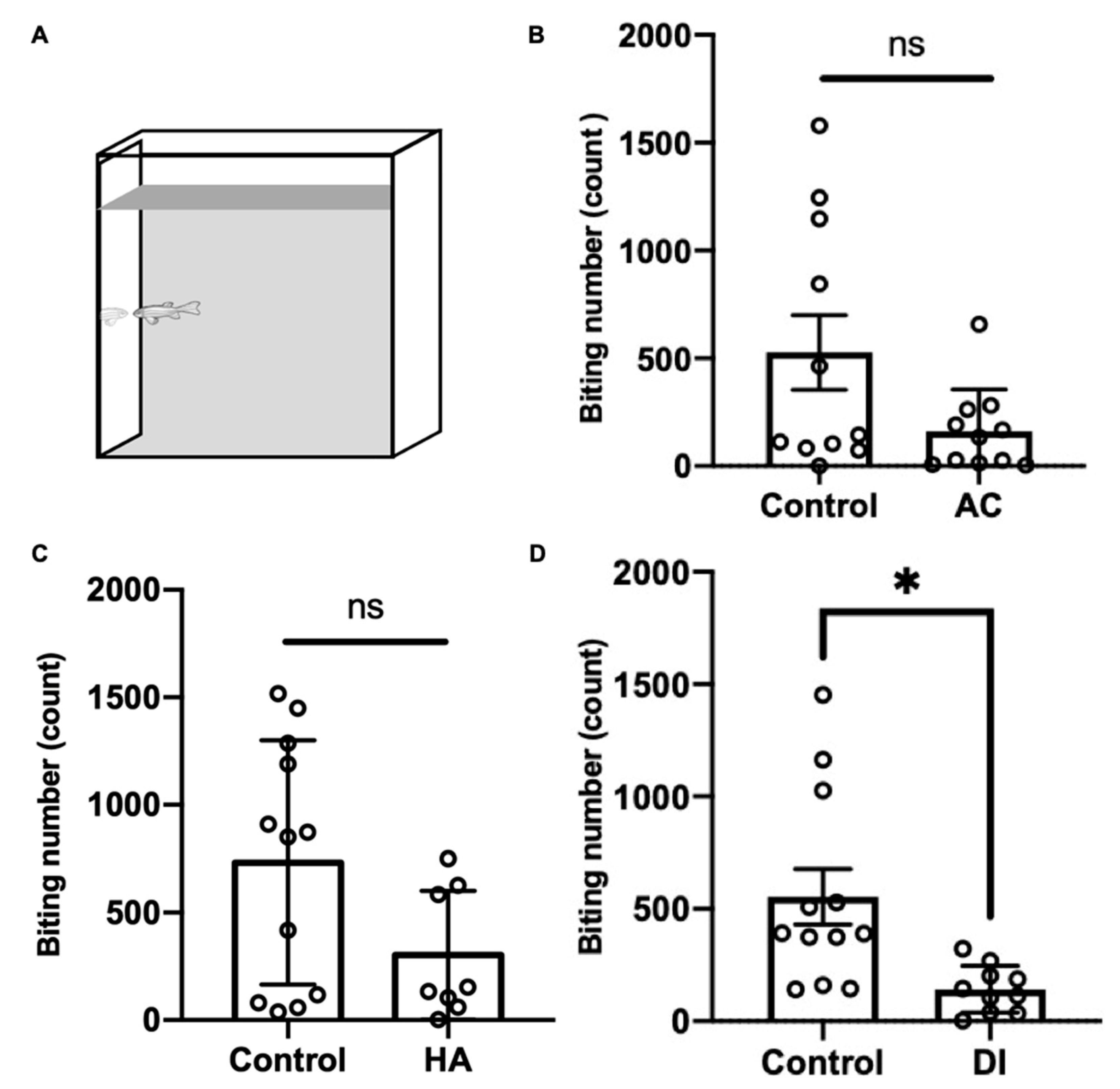
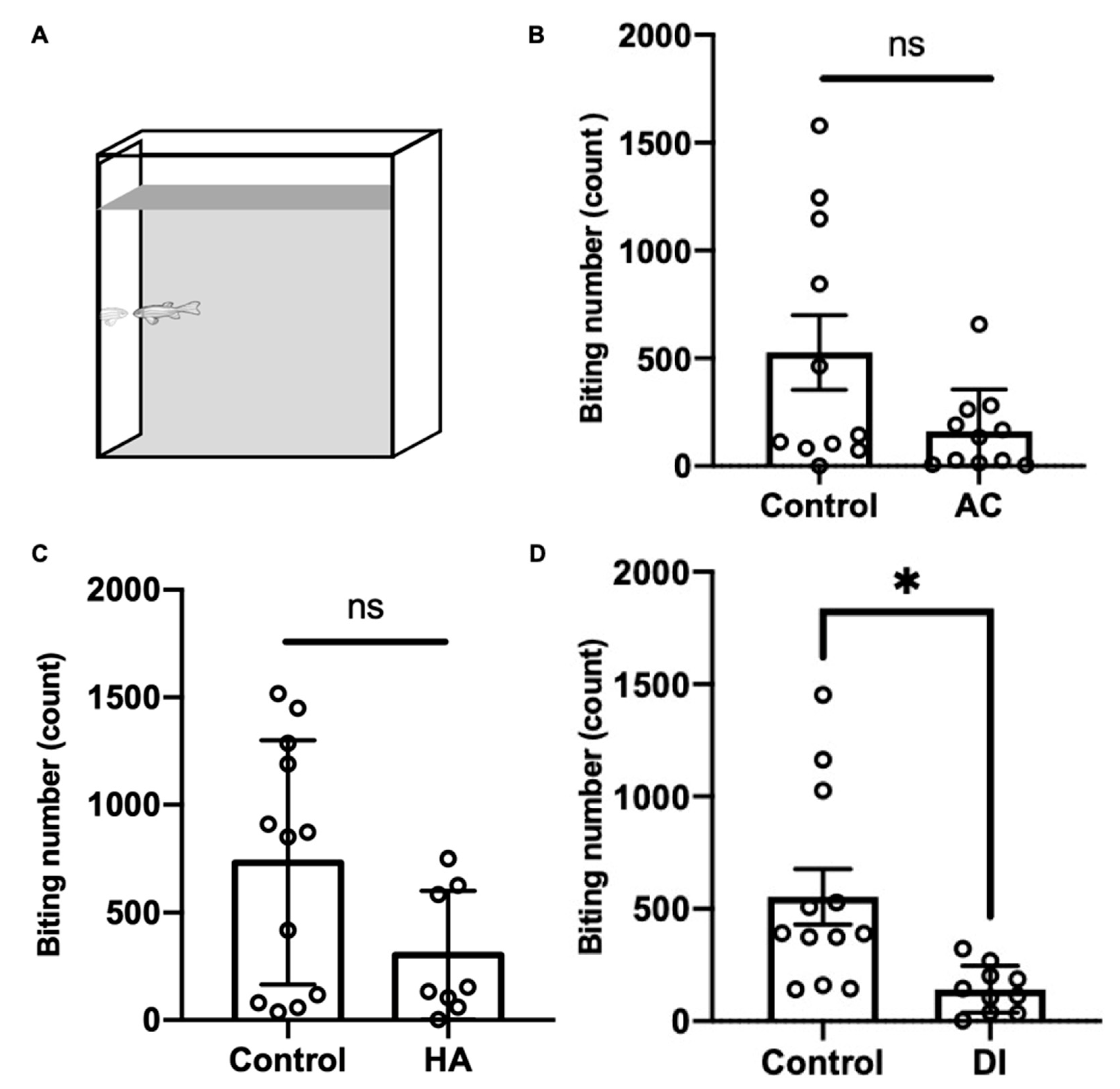
To evaluate the aggressiveness of the adult zebrafish under various environmental stresses, the mirror biting test was performed. After AC, HA, and DI water treatments for 24 h, the fish showed a lower biting frequency than did the control fish; however, the finding did not reach statistical significance in the AC and HA groups (Figure 7).
3. Discussion
The present study demonstrated that the oxytocin system participated in the acclimation of zebrafish to environmental fluctuations. The expression of oxytocin receptors in the peripheral tissue (the gills) and the central nervous system (the brain) was altered by environmental stresses; this finding indicates that the oxytocin system serves as a stress indicator. We also provided anatomical and behavioral evidences induced by stressors through the oxytocin system. This study evaluated the role of oxytocin signaling as a potential stress indicator, and our study results provided insights into the effects of oxytocin signaling on stress responses.
Oxytocin is a conserved neuropeptide with multiple functions, including adaptation to stress [39,51]. Oxytocin in the central nervous system reduced corticosterone in rats exposed to noise stress [52]. A higher plasma oxytocin level was discovered in older women under social stress [51]. Moreover, recent studies have indicated that the expression and functions of oxytocin receptors are important for adaptation to stress among animals. Forced swimming stress increased the expression of oxytocin receptors in the brains of rats [53], whereas isolation stress downregulated the mRNA expression of oxytocin receptors in mice [54]. In humans, the polymorphism of oxytocin receptor genes is related to empathy and stress reactivity [55]. The expression of oxytocin receptors was upregulated by hypomethylation in patients with social anxiety disorder [56]. These findings indicate that oxytocin signaling is involved in stress responses. In the present study, the expression of two oxytocin receptor genes (oxyr and oxyrl) in the gills was downregulated by AC, HA, and DI treatments (Figure 2). Studies have demonstrated that physiological biomarkers (stress markers) are changed (either stimulated or suppressed) by stressors [3,13]. In tilapia, pollution stress upregulated the levels of cortisol, blood urea nitrogen, and thyroxine (T4) and downregulated the levels of blood glucose, triglyceride, and triiodothyronine (T3) [24]. Multiple stresses (netting, chasing, air exposure, and confinement stresses) increased plasma glucose and lactate levels in common carp [57]. Nickel (Ni) exposure decreased the levels of total protein, lipids, glucose, and glycogen in the gills of carp [58]. Chasing or netting stress upregulated the levels of cortisol, glucose, and lactate in the gills of zebrafish and rainbow trout [59]. In addition to physiological parameters, stress-related molecules are applied to evaluate whether individuals are under stress conditions. Hyperthermal stress caused an increase in heat shock protein 70 (HSP70) and a decrease in GR in rainbow trout [60]. Under hypertonic stress, the expression of HSP70 and HSP90 was upregulated in the gills of milkfish [12]. Under cold stress, the expression of GRs and mineralocorticoid receptors was downregulated in carp [61]. All the aforementioned observations indicate that the expression of molecular biomarkers reflect the stress conditions of fish. According to the aforementioned findings, the alteration of oxyr and oxyrl expression in the gills provides evidence that oxytocin signaling serves as a biomarker for stress conditions.
In the present study, the highest expression of oxytocin receptors was found in the brain (Figure 1). AC water induced oxyr expression in the brain (Figure 3A), and oxyrl expression was downregulated by AC water but upregulated by HA treatment (Figure 3B,D). The different expression patterns of oxyr and oxyrl in the brain under AC treatment revealed the different mechanisms of these two receptors. After DI treatment, the expression of oxyr and oxyrl did not change (Figure 3E,F). To investigate the effects of various environmental stresses on the oxytocin system, we measured the numbers of oxytocin neurons after various stress challenges. A previous study found that the cell bodies of oxytocin neurons were located in the PPa in rainbow trout [62]. In the present study, we observed oxytocin neuron somas in the parvocellular preoptic nucleus (PPa and PPp) and periventricular nucleus of TPp (Figure 4 and Figure 5). The oxytocin neuron numbers increased after DI treatment but remained unchanged after AC and HA treatments (Figure 4). In mice, repeated defeat experiences and vicarious social stress increased the number of oxytocin neurons in the paraventricular nucleus of the hypothalamus (PVN) and bed nucleus of the stria terminalis, respectively [63,64]. Under chronic homotypic stress, the number of oxytocin neurons in the PVN also increased [65,66]. Given that the expression of oxytocin receptors in the brain changed according to various stressors and that the number of oxytocin neurons increased under hypotonic stress, the oxytocin system may be involved in acclimation to environmental fluctuations.
In addition to hormones, oxytocin serves as a neurotransmitter [67]. Oxytocin receptors are widely present in many brain regions of mammals, including the hypothalamus, prefrontal cortex (PFC), hippocampus, and amygdala [39,68]. The innervations of oxytocin neurons were observed in the cortex, pallidum, thalamic area, striatum, hypothalamic area, midbrain, hindbrain, and medulla of mice [69]. Through single-cell labeling and neurite tracing, Wircer and colleagues (2017) reported that zebrafish oxytocin neurons projected to the hindbrain and spinal cord 5 days post-fertilization (dpf) embryos [70]. Besides the projection of oxytocin neurons to hindbrain and spinal cord, Wee and her colleagues further demonstrated that this innervation of oxytocin neurons drove the nocifensive behavior of zebrafish via the premotor targets in the brainstem of 6–8 dpf embryos [71]. However, the projections of oxytocin neurons in the adult brain remained unclear. In the present study, for the first time, we observed oxytocin axons in the telencephalic area (Vv), anterior and ventromedial thalamic nuclei (A and VM), preglomerular nuclei (PGm), TPp, Cans, IPN, and raphe in the adult zebrafish brain (Figure 5). The projections and innervations of oxytocin neurons might regulate fish behaviors.
Oxytocin exerts anxiolytic and stress-alleviating effects in animals under stress [72]. Serotonergic neurons in the median raphe express oxytocin receptors and are activated by oxytocin, and such activation reduces anxiety-related behavior in mice [73]. Medial PFC (mPFC) interneurons expressing oxytocin receptors also exert anxiolytic effects by producing corticotropin-releasing hormone-binding protein that blocks the effects of corticotropin-releasing hormone [74]. The intranasal administration of oxytocin attenuated the activity of the amygdala in patients shown a fearful face [75]. The activation of presynaptic oxytocin receptors enhanced depolarization-evoked glutamate release in the hippocampus and reduced anxiety- and depressive-like behaviors [76]. Moreover, oxytocin neurons have been reported to project into the raphe, cortex areas, amygdala, and hippocampus [69], suggesting that the reduction in anxiety and fear responses is due to the oxytocin neuron innervation in these brain regions. Knobloch and colleagues (2012) demonstrated that hypothalamic oxytocin neurons projected to the central amygdala and that optogenetic stimulation of these neurons inhibited the output neurons of the central amygdala and decreased the freezing response after fear conditioning in rats [36]. In the present study, the axons of oxytocin neurons innervated the telencephalon and thalamus in adult zebrafish (Figure 5C,G, respectively). The telencephalon of fish is believed to be homologous to the mammalian limbic system, which controls anxiety, emotion, and motivation [77]. In zebrafish, the activation of cannabinoid receptor type-1 (CB1 receptor) in the telencephalon decreased anxiety from acute restraint stress [78]. Pharmacological activation of the GABAergic system in the telencephalon also reduced the anxiety levels of Japanese medaka [79]. The thalamus consists of several nuclei and is involved in various functions, including the regulation of anxiety and fear in humans [80]. Pharmacological suppression of c-fos expression in the reuniens (RE) and rhomboid (RH) nuclei in the thalamus increased avoidance/defensive behaviors in the elevated plus-maze in rats [81]. The abnormal asymmetry of the thalamic volume was demonstrated to be related to social anxiety disorder in children and adolescents under chronic family stress [82]. In the present study, HA treatment decreased the anxiety level in fish (Figure 6F). This result together with the finding that oxytocin neurons innervated the telencephalon and thalamus in the adult zebrafish brain provides evidence of the anxiolytic effects of oxytocin neuron innervation in specific brain regions under HA stress.
Through aggressive behavior, animals consolidate resources and status in society [83]. Oxytocin was reported to mediate aggression and affiliative behaviors in several species. Oxytocin-knockout mice showed reduced attacking time to their opponents but exhibited normal sensorimotor performance [84]. GABAergic neurons expressing oxytocin receptors in the lateral and capsular division of the central amygdala (CeL/C) inhibited the output of the medial part of the central amygdala (CeM) as well as decreased aggression toward pups and then enhanced the attacks toward intruders [85,86]. The intranasal administration of oxytocin in humans promoted an aggressive choice of fictitious partners in monetary games [87] and increased the inclination toward intimate partner violence in participants prone to physical aggression [88]. In zebrafish, the habenula–IPN circuit regulates aggression and social conflict [89,90]. The medial subregion of the dorsal habenula (dHbM)–intermediate/ventral IPN (i/vIPN) pathway and the lateral subregion of the dorsal habenula (dHbL)–dorsal/intermediate IPN (d/iIPN) pathway facilitated loser and winner behaviors through projections into the medial raphe and dorsal tegmental area, respectively, in male zebrafish [89]. Hunger potentiated the dHbL–d/iIPN pathway and increased the probability of winning a fight during dyadic fighting [90]. In addition, the raphe was reported to control the escalation of aggression. Pharmacological activation of presynaptic GABAB receptors inhibited 5-HT neurons in the dorsal raphe nucleus (DRN) and enhanced aggressive behaviors in mice [91]. Microinjection of L-glutamate into the DRN dose-dependently upregulated the number of attacking bites toward intruders, and the glutamate level in the DRN was increased during aggressive encounters [92]. Optogenetic stimulation of the median raphe decreased aggression in a phasic-like manner, whereas the stimulation of the dorsal raphe marginally but continuously diminished aggression and promoted social interactions in mice [93]. In the present study, the axons of oxytocin neurons were found to reach the IPN and raphe of the adult zebrafish brain (Figure 5K,L, respectively). We also found that environmental fluctuations downregulated the aggressiveness of the fish (Figure 7). Altogether, these findings suggest that the innervation of oxytocin neurons modified neural transmission and/or plasticity in the IPN and raphe and subsequently reduced aggressiveness in the adult zebrafish under AC, HA, and DI water treatments.
4. Materials and Methods
4.1. Experimental Animals
Mature zebrafish (D. rerio, AB strain and transgenic line) were reared in tanks filled with circulating filtered tap water at 28 °C ± 1 °C that was partially refreshed periodically. They were reared under a photoperiod cycle of 14-h light:10-h darkness (9:00–23:00) [94], and the pH value of water was monitored and maintained at 7.2 ± 2. The transgenic line (oxtl:EGFP) was obtained from Gil Levkowitz’s laboratory, and the location of oxytocin neurons was determined in the fish [95]. All experiments were conducted according to the principles and procedures approved by the Institutional Animal Care and Use Committee of National Taiwan University (IACUC approval no. NTU107-EL-00205 to M.Y. Chou).
4.2. Environmental Challenges to Adult Zebrafish
The zebrafish were gently transferred from their home tanks to different extreme environments at 28 °C ± 1 °C for 7 days. During the experiments, the fish were housed at a density of 1 fish/L in independent 10 L tanks with aeration and starved to avoid the effects of feeding and there were the independent control groups for different treatments. At the same time, both male and female zebrafish were existed in the same tank (sex ratio about 50%). In the tanks, 8 L water was changed daily to maintain environmental stability. In the present study, acidic (AC) water, high-ammonia (HA) water, and double-deionized (DI) water were used as extreme environments. DI water was collected from a water-purification system (A4-A8SL, Prema, Taipei, Taiwan). HA water (water containing 5 mM NH4+) was prepared by adding 5 mL of stock solution (1 M NH4Cl) and 300 mM 3-(N-morpholino) propanesulfonic acid buffer (MOPS; K35208029, Merck, Darmstadt, Germany; the pH value stabilizer) to 1 L fresh water (FW, filtered tap water). AC water was prepared through the addition of concentrated H2SO4 to FW (pH 7.2); the pH of this acidic water was adjusted to 4 ± 0.1. Measurement of pH was conducted by using a portable pH meter (pH 3310, WTW GmbH, Weilheim, Germany). For DI and AC treatments, FW was used as the control. For HA, FW with 300 mM MOPS was used as the control.
4.3. Total RNA Isolation and Complementary DNA Preparation
Tissues were frozen in liquid nitrogen and then homogenized in an appropriate amount of TRIzol reagent (Ambion, Woodward, TX, USA), using TissueLyser II (Qiagen, Valencia, CA, USA), and total RNA was purified by following the manufacturer’s protocol. To remove genomic DNA, total RNA was treated with DNase I (Promega, Madison, WI, USA), and the quality and quantity of total RNA were determined through agarose gel electrophoresis and NanoDrop ND-1000 (Thermo Scientific, Wilmington, DE, USA), respectively. For complementary DNA (cDNA) preparation, SuperScript IV reverse transcriptase (Invitrogen, Carlsbad, CA, USA) was used for the conversion of 5 μg of total RNA by following the manufacturer’s protocol, and the cDNA was stored at −20 °C for further experiments.
4.4. mRNA Expression of Oxytocin Receptors
Total RNA of the following tissues was isolated from the adult zebrafish: brain, eyes, gills, heart, intestine, kidney, liver, muscle, skin, and spleen. Tissue distributions of the following genes were determined (with the encoded proteins given in parentheses): oxyr (oxytocin receptor), oxyrl (oxytocin receptor–like), and rpl13a (ribosomal protein L13a), which served as the internal positive control. The sequences and Ensemble ID of primers are listed in Table 1. The genes were detected through semi-quantitative polymerase chain reaction (PCR) with Q5 high-fidelity DNA polymerase (M0491L, New England BioLabs, Ipswich, MA, USA) by following the manufacturer’s protocol. Different reaction cycles and amount of templates were tested to make sure the sampling condition was in a linear/quantifiable range (data not shown), and the amplified conditions were finally decided as 40 cycles. All the amplicons were sequenced to confirm the predicted products.
4.5. Real-Time Quantitative Polymerase Chain Reaction
Real-time quantitative PCR (RT-qPCR) was performed by using a LightCycler 480 real-time PCR system (Roche, Penzberg, Germany) in a final volume of 10 μL. The reaction mixture consisted of 5 μL 2× SYBR Green I Master mix (Roche) and 500 nM primer pairs. The reaction conditions were as follows: preincubation at 95 °C for 5 min and 45 cycles of amplification at 95 °C for 10 s, 60 °C for 10 s, and 72 °C for 10 s. The primer sets used in RT-qPCR are provided in Table 1. Moreover, rpl13a and β-actin were used as internal controls to obtain the confident quantification [96]. In the present study, normalization through the use of two reference genes showed similar patterns; therefore, only data normalized to rpl13a are shown. The products from each primer pair were subjected to melting-curve analysis and Sanger sequencing to verify their specificity. The efficiency of amplification was confirmed through serial dilution of cDNA for each primer set [97]. A non-template control (NTC) was conducted with sterile water to determine background DNA contamination. The relative expression of target genes normalized to internal control [98] was determined by using the following equation: 2−ΔΔC(t).
4.6. Brain Fixation and Sectioning
Transgenic zebrafish (oxtl:EGFP) were anesthetized on ice, and the heads of fish were removed immediately and fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS, 137 mM NaCl, 2.7 mM KCl, 1.8 mM KH2PO4, and 10 mM Na2HPO4; pH 7.4) at 4 °C, overnight. After fixation, the skull was removed carefully, and the whole brain was then mounted in 2% agarose prepared with PBS. Subsequently, 45 and 60 µm sections were obtained by using a Vibratory Microtome (Leica VT1200 S, Leica Microsystems, Wetzlar, Germany), and the sections were maintained in PBS and stored at 4 °C for analysis.
4.7. Immunohistochemistry
The sections were rinsed with PBS-Tr (PBS with 0.1% Triton X-100; T8787, Sigma-Aldrich, UK) three times and then incubated in 1% blocking reagent (11096176001, Roche) diluted with PBS for at least 2 h, at room temperature.
For mapping the projections of oxytocin neurons, 60 µm–thick sections were incubated with rabbit anti-GFP antibody (1: 2000; ab13970) at 4 °C, overnight, followed by incubation with CF 568 goat anti-rabbit secondary antibody (20102; Biotium, Fremont, CA, USA), at room temperature, for 2 h. After the section was rinsed with PBS-Tr, it was reacted with NeuroTrace 640/660 (N21483, Invitrogen) and then incubated with 1 µg/mL DAPI for 20 min for nucleus targeting.
For cell counting, 45 µm–thick sections were incubated with rabbit anti-GFP antibody (1: 2000; ab13970) at 4 °C, overnight, followed by incubation with CF 488 goat anti-rabbit secondary antibody (20012; Biotium), at room temperature, for 2 h. After the section was rinsed with PBS-Tr, it was reacted with NeuroTrace 530/615 (N21482, Invitrogen); then cell counting was conducted.
4.8. Cell Counting for Oxytocin Neurons
Whole-brain sections were applied to quantify the number of oxytocin neurons in a single individual. About 10 sections were collected from one brain. The free software Fiji was used to analyze the images of each section that was taken by a Zeiss Axio Imager Z2 Upright Microscope equipped with a Prime 95B sCMOS camera (TELEDYNE Photometrics, Tucson, AZ, USA). The final cell-counting data were corrected by the thickness of sections and the diameter of the nuclei through Abercrombie’s equation [99].
4.9. Novel Tank Diving Test
The novel tank diving test was performed to measure the locomotion activity and anxiety level of the fish [100], and the procedure in our previous study was followed to ensure a consistent behavioral paradigm [94]. After exposure to DI, HA, and AC water for 24 h, the adult zebrafish were transported individually from their home tank to a 3.5 L tank (26 cm × 8 cm× 17 cm; water volume: 2.5 L), through careful handling, to reduce stress. Fish behaviors were recorded by a digital camera (SONY HDR-SR11, Tokyo, Japan), which was mounted at the lateral site of the test tank, and the videos were analyzed by using EthoVision XT 14 (Noldus Information Technology, Wageningen, The Netherlands). During the behavioral record, the tank was in an independent room without any interference to avoid the effects from surrounding area. The center of gravity of the fish was defined as the fish position. The total swimming distance and the time spent at the top of the tank were analyzed for 15 min after transfer of the fish to evaluate the locomotion activity and anxiety level. The test was repeated thrice.
4.10. Mirror-Biting Test
The mirror-biting test was used to measure aggressiveness [100] according to a previously described method [94]. A 3.5 L tank (26 cm × 8 cm × 18 cm; water volume: 2.5 L) with a mirror attached to one of its sidewalls was used. Before the fish were placed in the test tank, an opaque partition was placed to prevent the fish from seeing the mirror. After exposure to AC, HA, and DI water for 24 h, the zebrafish were carefully transferred individually from their home tank to the test tank, through careful handling, to reduce stress. During the behavioral record, the tank was in an independent room, without any interference, to avoid the effects from surrounding area. After the fish were allowed 15 min of acclimation, the opaque partition was removed without excessive disturbance. Zebrafish behaviors were videotaped for 10 min by a digital camera (SONY HDR-SR11), which was mounted at the lateral site of the test tank, and an experienced observer analyzed the mirror-biting frequency.
4.11. Statistical Analysis
Data are expressed as the mean ± deviation (SD) for the parametric data and the mean ± standard error of the mean (SEM) for nonparametric data. Before statistical analysis was performed, ROUT method was used to identify the outlier. Then the Shapiro–Wilk normality test was applied to evaluate normality in all the datasets. The datasets showing normal distribution were analyzed by Student’s t-test. The dataset of the mirror-biting test with AC treatment was not normally distributed and was analyzed by Mann–Whitney test (Figure 7B). The raw data used for all statistical analyses can be found at https://tinyurl.com/oxytocinzebrafish, accessed on 1 June 2021. Statistical analysis was conducted by using GraphPad Prism 7.
5. Conclusions
In summary, our results demonstrated that the oxytocin system serves as a potential stress indicator that enables animals to adapt to environmental fluctuations. Oxytocin regulates physiological functions through the endocrine system and affects behavioral outcomes by innervating downstream nuclei in the brain. Under stress, behavioral change is a strategy through which animals adapt to extreme environments. Such behavioral changes may allocate energy metabolism, which improves animal survival with minimal adjustments to physiological and metabolic responses and ultimately increases the fitness of animals for environmental adaptation.
Author Contributions
Conceptualization, H.-J.C. and M.-Y.C.; methodology, H.-J.C., C.-Y.C., H.-P.H.; software, C.-Y.C., H.-P.H.; validation, H.-J.C., C.-Y.C. and M.-Y.C.; formal analysis, H.-J.C., C.-Y.C. and H.-P.H.; investigation, H.-J.C. and M.-Y.C.; data curation, H.-J.C. and M.-Y.C.; writing—original draft preparation, H.-J.C. and M.-Y.C.; writing—review and editing, H.-J.C., C.-Y.C. and M.-Y.C.; visualization, H.-J.C.; supervision, M.-Y.C.; project administration, M.-Y.C.; funding acquisition, M.-Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Taiwan’s Ministry of Science and Technology (MOST-109-2628-B-002-038, MOST-109-2321-B-002-042 and MOST-110-2628-B-002-050) and National Taiwan University (NTU) (NTU-CC-109L892701), which were awarded to Ming-Yi Chou. This article was subsidized for English editing by National Taiwan University under the Excellence Improvement Program for Doctoral Students (grant number 108-2926-I-002-002-MY4), sponsored by Ministry of Science and Technology, Taiwan.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Animal Care and Use Committee of National Taiwan University (IACUC approval no. NTU107-EL-00205 to M.Y. Chou).
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data used for all statistical analyses can be found at https://tinyurl.com/oxytocinzebrafish, accessed on 1 June 2021.
Acknowledgments
We thank the Technology Commons, College of Life Science, NTU, for the technical support in our molecular biological experiments. This article was subsidized by NTU. The funding source had no influence in the study design; collection, analysis, or interpretation of the data; or writing of the manuscript. This manuscript was edited by Wallace Academic Editing.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AC | acidic |
| HA | high ammonia |
| DI | double-ionized |
| HPA | hypothalamic–pituitary–adrenal |
| GR | glucocorticoid receptor |
| MO | morpholino |
| AuNP-PVP | gold nanoparticles coated with PVP |
| PPa | the anterior part of the parvocellular preoptic nucleus |
| PPp | the posterior part of the parvocellular preoptic nucleus |
| TPp | the periventricular nucleus of the posterior tuberculum |
| Hc | the caudal zone of the periventricular hypothalamus |
| Vv | the ventral nuclei of the ventral telencephalic area |
| VM | the ventromedial thalamic nuclei |
| A | the anterior thalamic nuclei |
| PGm | the preglomerular nuclei |
| TPp | the periventricular nucleus of the posterior tuberculum |
| Cans | the commissura ansulata |
| IPN | the interpeduncular nucleus |
| TTBc | the crossed tecto-bulbar tract |
| PVN | the paraventricular nucleus of the hypothalamus |
| PFC | the prefrontal cortex |
| Dpf | day-post-fertilization |
| mPFC | the medial prefrontal cortex |
| CB1 receptor | cannabinoid receptor type-1 |
| RE | the reuniens nuclei in the thalamus |
| RH | the rhomboid nuclei in the thalamus |
| CeL/C | the lateral and capsular division of the central amygdala |
| CeM | the central amygdala |
| dHbM | the medial subregion of the dorsal habenula |
| i/vIPN | the intermediate/ventral interpeduncular nucleus |
| dHbL | the lateral subregion of the dorsal habenula |
| d/iIPN | the dorsal/intermediate interpeduncular nucleus |
| DRN | the dorsal raphe nucleus |
References
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the stress response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef] [PubMed]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Fishelson, Z.; Hochman, I.; Greene, L.E.; Eisenberg, E. Contribution of heat shock proteins to cell protection from complement-mediated lysis. Int. Immunol. 2001, 13, 983–991. [Google Scholar] [CrossRef] [Green Version]
- Fromm, P.O. A review of some physiological and toxicological responses of freshwater fish to acid stress. Env. Biol. Fishes 1980, 5, 79–93. [Google Scholar] [CrossRef]
- Hattingh, J. The surface area of mudfish. J. Fish. Biol. 1976, 8, 19–22. [Google Scholar] [CrossRef]
- Iwama, G.K.; Afonso, L.O.; Todgham, A.; Ackerman, P.; Nakano, K. Are hsps suitable for indicating stressed states in fish? J. Exp. Biol. 2004, 207, 15–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmini, E.; Rani, M.U. Evaluation of oxidative stress biomarkers in hepatocytes of grey mullet inhabiting natural and polluted estuaries. Sci. Total Environ. 2009, 407, 4533–4541. [Google Scholar] [CrossRef] [PubMed]
- Taleb, M.; Brandon, C.S.; Lee, F.S.; Lomax, M.I.; Dillmann, W.H.; Cunningham, L.L. Hsp70 inhibits aminoglycoside-induced hair cell death and is necessary for the protective effect of heat shock. J. Assoc. Res. Otolaryngol. 2008, 9, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.H.; Chang, I.C.; Chen, C.H.; Lee, T.H.; Hwang, P.P. Phenotypic changes in mitochondrion-rich cells and responses of Na+/K+-ATPase in gills of tilapia exposed to deionized water. Zool. Sci. 2008, 25, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Tine, M.; Bonhomme, F.; McKenzie, D.J.; Durand, J.D. Differential expression of the heat shock protein Hsp70 in natural populations of the tilapia, Sarotherodon melanotheron, acclimatised to a range of environmental salinities. BMC Ecol. 2010, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Khotinul, U.; Chuang, H.J.; Chiu, L.; Yang, W.K.; Wang, Y.C.; Wu, W.Y.; Lee, T.H. Potential osmoprotective roles of branchial heat shock proteins towards Na+, K+ -ATPase in milkfish (Chanos chanos) exposed to hypotonic stress. Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 2020, 248, 110749. [Google Scholar]
- Harper, C.; Wolf, J.C. Morphologic effects of the stress response in fish. ILAR J. 2009, 50, 387–396. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Lowell, B.B. The hypothalamus. Curr. Biol. 2014, 24, R1111–R1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedemeyer, G.; Barton, B.B.; McLeay, D.J. Stress and acclimation. In Methods for Fish Biology; Schreck, C.B., Moyle, P.B., Eds.; American Fisheries Society: Bethesda, MD, USA, 1990; pp. 451–489. [Google Scholar]
- Lin, C.H.; Shih, T.H.; Liu, S.T.; Hsu, H.H.; Hwang, P.P. Cortisol regulates acid secretion of H+-ATPase-rich ionocytes in zebrafish (Danio rerio) embryos. Front. Physiol. 2015, 6, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jezova, D.; Skultetyova, I.; Tokarev, D.I.; Bakos, P.; Vigas, M. Vasopressin and oxytocin in stress. Ann. N. Y. Acad. Sci. 1995, 771, 192–203. [Google Scholar] [CrossRef]
- Weninger, S.C.; Dunn, A.; Muglia, L.J.; Dikkes, P.; Miczek, K.A.; Swiergiel, A.H.; Berridge, C.W.; Majzoub, J.A. Stress-induced behaviors require the corticotropin-releasing hormone (CRH) receptor, but not CRH. Proc. Natl. Acad. Sci. USA 1999, 96, 8283–8288. [Google Scholar] [CrossRef] [Green Version]
- Cannon, W.B. Organization for physiological homeostasis. Physiol. Rev. 1929, 9, 399–431. [Google Scholar] [CrossRef]
- Hu, Y.C.; Chu, K.F.; Hwang, L.Y.; Lee, T.H. Cortisol regulation of Na+, K+ -ATPase β1 subunit transcription via the pre-receptor 11β-hydroxysteroid dehydrogenase 1-like (11β-Hsd1L) in gills of hypothermal freshwater milkfish, Chanos chanos. J. Steroid Biochem. Mol. Biol. 2019, 192, 105381. [Google Scholar] [CrossRef]
- Hontela, A.; Rasmussen, J.B.; Audet, C.; Chevalier, G. Impaired cortisol stress response in fish from environments polluted by PAHs, PCBs, and Mercury. Arch. Environ. Contam. Toxicol. 1992, 22, 278–283. [Google Scholar] [CrossRef]
- Teles, M.; Soares, A.M.V.M.; Tort, L.; Guimarães, L.; Oliveira, M. Linking cortisol response with gene expression in fish exposed to gold nanoparticles. Sci. Total Environ. 2017, 584–585, 1004–1011. [Google Scholar] [CrossRef]
- Canli, E.G.; Dogan, A.; Canli, M. Serum biomarker levels alter following nanoparticle (Al2O3, CuO, TiO2) exposures in freshwater fish (Oreochromis niloticus). Environ. Toxicol. Pharmacol. 2018, 62, 181–187. [Google Scholar] [CrossRef]
- Ranabir, S.; Reetu, K. Stress and hormones. Indian J. Endocrinol. Metab. 2011, 1, 18–22. [Google Scholar] [CrossRef]
- Dale, H.H. On some physiological actions of ergot. J. Physiol. 1906, 34, 163–206. [Google Scholar] [CrossRef] [PubMed]
- Gimpl, G.; Fahrenholz, F. The oxytocin receptor system: Structure, function, and regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef] [Green Version]
- Sharpey-Schafer, E.A.; Mackenzie, K. The action of animal extracts on milk secretion. Proc. R. Soc. Lond. B Biol. Sci. 1911, 84, 16–22. [Google Scholar]
- Nishioka, T.; Anselmo-Franci, J.A.; Li, P.; Callahan, M.F.; Morris, M. Stress increases oxytocin release within the hypothalamic paraventricular nucleus. Brain Res. 1998, 781, 57–61. [Google Scholar] [CrossRef]
- Torner, L.; Plotsky, P.M.; Neumann, I.D.; Jong, T.R. Forced swimming-induced oxytocin release into blood and brain: Effects of adrenalectomy and corticosterone treatment. Psychoneuroendocrinology 2017, 77, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Amico, J.A.; Mantella, R.C.; Vollmer, R.R.; Li, X. Anxiety and stress responses in female oxytocin deficient mice. J. Neuroendocrinol. 2004, 16, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Hoge, E.A.; Pollack, M.H.; Kaufman, R.E.; Zak, P.J.; Simon, N.M. Oxytocin levels in social anxiety disorder. Cns Neurosci. Ther. 2008, 14, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Wei, F.Y.; Matsunaga, T.; Matsunaga, N.; Kaitsuka, T.; Tomizawa, K. Oxytocin protects against stress-induced cell death in murine pancreatic β-cells. Sci. Rep. 2016, 6, 25185. [Google Scholar] [CrossRef] [PubMed]
- Klement, J.; Ott, V.; Rapp, K.; Brede, S.; Piccinini, F.; Cobelli, C.; Lehnert, H.; Hallschmid, M. Oxytocin improves β-cells responsivity and glucose tolerance in healthy men. Diabetes 2017, 66, 264–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, D.D.; Champagne, F.C.; Meaney, M.J. Variations in maternal behaviour are associated with differences in oxytocin receptor levels in the rat. J. Neuroendocrinol. 2000, 12, 1145–1148. [Google Scholar] [CrossRef]
- Knobloch, H.S.; Charlet, A.; Hoffmann, L.C.; Eliava, M.; Khrulev, S.; Certin, A.H.; Osten, P.; Schwarz, M.K.; Seeburg, P.H.; Stoop, R.; et al. Evoked axonal oxytocin release in the central amygdala attenuates fear response. Neuron 2012, 73, 553–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, M.Y.; Hung, J.C.; Wu, L.C.; Hwang, S.P.L.; Hwang, P.P. Isotocin controls ion regulation through regulating ionocytes progenitor differentiation and proliferation. Cell. Mol. Life Sci. 2011, 68, 2797–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warfvinge, K.; Krause, D.; Edvinsson, L. The distribution of oxytocin and the oxytocin receptor in rat brain: Relation to regions active in migraine. J. Headache Pain. 2020, 21, 10. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, H.; Latt, H.M.; Koga, Y.; Nishiki, T.; Matsui, H. Oxytocin and stress: Neural mechanisms, stress-related disorders and therapeutic approaches. Neuroscience 2019, 417, 1–10. [Google Scholar] [CrossRef]
- Iwama, G.K.; Vijayan, M.M.; Forsyth, R.B.; Ackerman, P.A. Heat shock proteins and physiological stress in fish. Am. Zool. 1999, 39, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.H.; Lee, T.H. Early response of protein quality control in gills is associated with survival of hypertonic shock in Mozambique tilapia. PLoS ONE 2013, 8, e63112. [Google Scholar] [CrossRef] [Green Version]
- Utne-Palm, A.C.; Smith, A. Fish as laboratory animals. In The Welfare of Fish; Kristiansen, T.S., Fernö, A., Pavlids, M.A., van de Vis, H., Eds.; Springer: Cham, Switzerland, 2020; pp. 375–400. [Google Scholar]
- Hatting, J. The influence of carbon dioxide on the blood sugar concentration in the fresh-water fish, Labeo capensis (smith). Comp. Biol. Physiol. A Comp. Physiol. 1976, 53, 235–236. [Google Scholar] [CrossRef]
- Zhao, L.; Cui, C.; Liu, Q.; Sun, J.; He, K.; Adam, A.A.; Luo, J.; Li, Z.; Wang, Y.; Yang, S. Combined exposure to hypoxia and ammonia aggravated biological effects on glucose metabolism, oxidative stress, inflammation and apoptosis in largemouth bass (Micropterus salmoides). Aquat. Toxicol. 2020, 224, 105514. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.P.; Wu, S.M. Role of cortisol in hypoosmoregulation in larvae of the tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 1993, 92, 318–324. [Google Scholar] [CrossRef]
- Landin, J.; Hovey, D.; Xu, B.; Lagman, D.; Zettergren, A.; Larhammar, D.; Kettunen, P.; Westberg, L. Oxytocin receptors regulate social preference in zebrafish. Sci. Rep. 2020, 10, 5435. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Nunes, A.R.; Teles, M.; Anbalagan, S.; Blechman, J.; Levkowitz, G.; Oliveira, R.F. Genetic variation in the social environment affects behavioral phenotypes of oxytocin receoptor mutants in zebrafish. eLife 2020, 9, e56973. [Google Scholar] [CrossRef]
- Outtandy, P.; Russell, C.; Kleta, R.; Bockenhauer, D. Zebrafish as a model for kidney function and disease. Pediatr. Nephrol. 2019, 34, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miklósi, A.; Andrew, R.J. The zebrafish as a model for behavioral studies. Zebrafish 2006, 3, 227–234. [Google Scholar] [CrossRef]
- Zhang, K.; Liang, J.; Brun, N.R.; Zhao, Y.; Werdich, A.A. Rapid zebrafish behavioral profiling assay accelerates the identification of environmental neurodevelopmental toxicants. Environ. Sci. Technol. 2021, 55, 1919–1929. [Google Scholar] [CrossRef]
- Taylor, S.E.; Gonzaga, G.C.; Klein, L.C.; Hu, P.; Greendale, G.A.; Seeman, T.E. Relation of oxytocin to psychological stress responses and hypothalamic-pituitary-adrenocortical axis activity in older women. Psychosom. Med. 2006, 68, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Windle, R.J.; Shank, N.; Lightman, S.L.; Ingram, C.D. Central oxytocin administration reduces stress-induced corticosterone release and anxiety. Endocrinol. 1997, 138, 2829–2834. [Google Scholar] [CrossRef]
- Liberzon, I.; Young, E.A. Effects of stress and glucocorticoids on CNS oxytocin receptor binding. Psychoneuroendocrinology 1997, 22, 411–422. [Google Scholar] [CrossRef]
- Han, R.T.; Kim, Y.B.; Park, E.H.; Kim, J.Y.; Ryu, C.; Kimm, H.Y.; Lee, J.; Pahk, K.; Shanyu, C.; Kim, H.; et al. Long-term isolation elicits depression and anxiety-related behaviors by reducing oxytocin-induced GABAergic transmission in central amygdala. Front. Mol. Neurosci. 2018, 11, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, S.M.; Saslow, L.R.; Garcia, N.; John, O.P.; Keltner, D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 21437–21441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, C.; Dannlowski, U.; Bräuer, D.; Steven, S.; Laeger, I.; Wittmann, H.; Kugel, H.; Dobel, C.; Hurlemann, R.; Reif, A.; et al. Oxytocin receptor gene methylation: Converging multilevel evidence for a role in social anxiety. Neuropsychopharmacology 2015, 40, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Aerts, J.; Metz, J.R.; Ampe, B.; Decostere, A.; Filk, G.; Saeger, S.D. Scales tell a story on the stress history of fish. PLoS ONE 2015, 10, e0123411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shagirtha, K.; Priya; Arthi; Arun; Prabu, M. Histopathological and biochemical changes in grass carp gill and muscle tissues due to nickel exposure may be related to the induction of oxidative stress. Eur. J. Biomed. Pharm. 2018, 5, 439–447. [Google Scholar]
- Gesto, M.; Hernández, J.; López-Patiño, M.A.; Soengas, J.L.; Míguez, J.M. Is gill cortisol concentration a good acute stress indicator in fish? A study in rainbow trout and zebrafish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 188, 65–69. [Google Scholar] [CrossRef]
- Basu, N.; Kennedy, C.J.; Iwama, G.K. The effects of stress on the association between hsp70 and the glucocorticoid receptor in rainbow trout. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 134, 655–663. [Google Scholar] [CrossRef]
- Stolte, E.H.; Mazon, A.F.; Leon-Koosterziel, K.M.; Jesiak, M.; Bury, N.R.; Sturm, A.; Savelkoul, H.F.J.; Verburg van Kemenade, B.M.L.; Flik, G. Corticosteroid receptors involved in stress regulation in common carp, Cyprinus carpio. J. Endocrinol. 2008, 198, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Saito, D.; Komatsuda, M.; Urano, A. Functional organization of preoptic vasotocin and isotocin neurons in the brain of rainbow trout: Central and neurohypophysial projections of single neurons. Neuroscience 2004, 124, 973–984. [Google Scholar] [CrossRef]
- Duque-Wilckens, N.; Torres, L.Y.; Yokoyama, S.; Minie, V.A.; Tran, A.M.; Petkova, S.P.; Hao, R.; Ramos-Maciel, S.; Rios, R.A.; Jackson, K.; et al. Extrahypothalamic oxytocin neurons drive stress-induced social vigilance and avoidance. Proc. Natl. Acad. Sci. USA 2020, 117, 26406–26413. [Google Scholar] [CrossRef]
- Steinman, M.Q.; Duque-Wilckens, N.; Greenberg, G.D.; Hao, R.; Campi, K.L.; Laredo, S.A.; Laman-Maharg, A.; Manning, C.E.; Doig, I.E.; Lopez, E.M.; et al. Sex-specific effects of stress on oxytocin neurons correspond with responses to intranasal oxytocin. Biol. Psychiatry 2016, 80, 406–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Coleman, F.H.; Doheny, K.K.; Travagli, R.A. Stress adaptation upregulates oxytocin within hypothalamo-vagal neurocircuits. Neuroscience 2018, 390, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Babygirijia, R.; Bulbul, M.; Cerjak, D.; Ludwig, K.; Takahashi, T. Hypothalamic oxytocin mediates adaptation mechanism against chronic stress in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G946–G953. [Google Scholar] [CrossRef] [Green Version]
- Insel, T.R.; Young, L.; Wang, Z. Central oxytocin and reproductive behaviours. Rev. Reprod. 1997, 2, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Jurek, B.; Neumann, I.D. The oxytocin receptor: From intracellular signaling to behavior. Physiol. Rev. 2018, 98, 1805–1908. [Google Scholar] [CrossRef]
- Liao, P.Y.; Chiu, Y.M.; Yu, J.H.; Chen, S.K. Mapping central projection of oxytocin neurons in unmated mice using Cre and alkaline phosphatase reporter. Front. Neuroanat. 2020, 14, 559402. [Google Scholar] [CrossRef]
- Wircer, E.; Blechman, J.; Borodovsky, N.; Tsoory, M.; Nunes, A.R.; Oliveira, R.F.; Levkowitz, G. Homeodomain protein Otp affects developmental neuropeptide switching in oxytocin neurons associated with a long-term effect on social behavior. eLife 2017, 6, e22170. [Google Scholar] [CrossRef]
- Wee, C.L.; Nikitchenko, M.; Wang, W.C.; Luks-Morgan, S.J.; Song, E.; Gagnon, J.A.; Randlett, O.; Bianco, I.H.; Lacoste, A.M.B.; Glushenkova, E.; et al. Zebrafish oxytocin neurons drive nocifensive behavior via brainstem premotor targets. Nat. Neurosci. 2019, 22, 1477–1492. [Google Scholar] [CrossRef]
- Olff, M.; Frijling, J.L.; Kubzansky, L.D.; Bradley, B.; Ellenbogen, M.A.; Cardoso, C.; Bartz, J.A.; Yee, J.R.; van Zuiden, M. The role of oxytocin in social bonding, stress regulation and mental health: An update on the moderating effects of context and interindividual differences. Psychoneuroendocrinology 2013, 38, 1883–1894. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Takayanagi, Y.; Inoue, K.; Kimura, T.; Young, L.J.; Onaka, T.; Nishimori, K. Evidence that oxytocin exerts anxiolytic effects via oxytocin receptor expressed in serotonergic neurons in mice. J. Neurosci. 2009, 29, 2259–2271. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Nakajima, M.; Ibañez-Tallon, I.; Heintz, N. A cortical circuit for sexually dimorphic oxytocin-dependent anxiety behaviors. Cell 2016, 167, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Labuschagne, I.; Phan, K.L.; Wood, A.; Angstadt, M.; Chua, P.; Heinrichs, M.; Stout, J.C.; Nathan, P.J. Oxytocin attenuates amygdala reactivity to fear in generalized social disorder. Neuropsychopharmacology 2010, 35, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Mairesse, J.; Gatta, E.; Reynaert, M.L.; Marrocco, J.; Morley-Fletcher, S.; Soichot, M.; Deruyter, L.; Camp, G.V.; Bouwalerh, H.; Fagioli, F.; et al. Activation of presynaptic oxytocin receptors enhances glutamaterelease in the ventral hippocampus of prenatally restraint stressed rats. Psychoneuroendocrinology 2015, 62, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Von Trotha, J.W.; Vernier, P.; Bally-Cuif, L. Emotions and motivated behavior converge on an amygdala-like structure in the zebrafish. Eur. J. Neurosci. 2014, 40, 3302–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas Luz, W.; Santos-Silva, M.; Cardoso, P.B.; Assad, N.; da Silva Moraes, E.R.; Grisólia, A.B.A.; Braga, D.V.; Leão, L.K.R.; de Moraes, S.A.S.; Passos, A.C.; et al. Putative Activation of the CB1 Cannabinoid Receptors Prevents Anxiety-Like Behavior, Oxidative Stress, and GABA Decrease in the Brain of Zebrafish Submitted to Acute Restraint Stress. Front. Behav. Neurosci. 2021, 14, 598812. [Google Scholar] [CrossRef] [PubMed]
- Tsubokawa, T.; Saito, K.; Kawano, H.; Kawamura, K.; Shinozuka, K.; Watanabe, S. Pharmacological effects on mirror approaching behavior and neurochemical aspects of the telencephalon in the fish, medaka (Oryzias latipes). Soc. Neurosci. 2009, 4, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Asami, T.; Yoshida, H.; Takaishi, M.; Nakamura, R.; Yoshimi, A.; Whitford, T.J.; Hirayasu, Y. Thalamic shape and volume abnormalities in female patients with panic disorder. PLoS ONE 2018, 13, e0208152. [Google Scholar] [CrossRef]
- Linley, S.B.; Athanason, A.C.; Rojas, A.K.P.; Vertes, R.P. Role of the reuniens and rhomboid thalamic nuclei in anxiety-like avoidance behavior in the rat. Hippocampus 2021, 31, 756–769. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Lebowitz, E.R.; Zhang, F.; Hu, Y.; Liu, Z.; Yang, H.; Wu, J.; Wang, Y.; Silverman, W.K.; et al. Abnormal asymmetry of thalamic volume moderates stress from parents and anxiety symptoms in children and adolescents with social anxiety disorder. Neuropharmacology 2020, 180, 108301. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.K.S.; Chow, T.S.C. The effects of environmental resource and security on aggressive behavior. Aggress. Behav. 2017, 43, 304–314. [Google Scholar] [CrossRef]
- DeVries, A.C.; Young, W.S.; Nelson, R.J. Reduced aggressive behaviour in mice with targeted disruption of the oxytocin gene. J. Neuroendocrinol. 1997, 9, 363–368. [Google Scholar] [CrossRef]
- Campbell, A. Attachment, aggression and affiliation: The role of oxytocin in female social behavior. Biol. Psychol. 2008, 77, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debiec, J. Peptides of love and fear: Vasopressin and oxytocin modulate the integration of information in the amygdala. Bioassays 2005, 27, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Ne’eman, R.; Perach-barzilay, N.; Fischer-Shofty, M.; Atias, A.; Shamay-Tsoory, S.G. Intranasal administration of oxytocin increases human aggressive behavior. Horm. Behav. 2016, 80, 125–131. [Google Scholar] [CrossRef] [PubMed]
- DeWall, C.N.; Gillath, O.; Pressman, S.D.; Black, L.L.; Bartz, J.A.; Moskovitz, J.; Stetler, D.A. When the love hormone leads to violence: Oxytocin increases intimate partner violence inclinations among high trait aggressive people. Soc. Psychol. Personal. Sci. 2014, 5, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.Y.; Amo, R.; Kinoshita, M.; Cherng, B.W.; Shimazaki, H.; Agetsuma, M.; Shiraki, T.; Aoki, T.; Takahoko, M.; Yamazaki, M.; et al. Social conflict resolution regulated by two dorsal habenular subregions in zebrafish. Science 2016, 352, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Nakajo, H.; Chou, M.Y.; Kinoshita, M.; Appelbaum, L.; Shimazaki, H.; Tsuboi, T.; Okamoto, H. Hunger potentiates the habenular winner pathway for social conflict by orexin-promoted biased alternative splicing of the AMPA receptor gene. Cell Rep. 2020, 31, 107790. [Google Scholar] [CrossRef]
- Takahashi, A.; Shimamoto, A.; Boyson, C.O.; DeBold, J.F.; Miczek, K.A. GABA(B) receptor modulation of serotonin neurons in the dorsal raphé nucleus and escalation of aggression in mice. J. Neurosci. 2010, 30, 11771–11780. [Google Scholar] [CrossRef]
- Takahashi, A.; Lee, R.X.; Iwasato, T.; Itohara, S.; Arima, H.; Bettler, B.; Miczek, K.A.; Koide, T. Glutamate input in the dorsal raphe nucleus as a determinant of escalated aggression in male mice. J. Neurosci. 2015, 35, 6452–6463. [Google Scholar] [CrossRef] [Green Version]
- Balázsfi, D.; Zelena, D.; Demeter, K.; Miskolczi, C.; Varga, Z.K.; Nagyváradi, A.; Nyíri, G.; Cserép, C.; Baranyi, M.; Sperlágh, B.; et al. Differential roles of the two raphe nuclei in amiable social behavior and aggression-An optogenetic study. Front. Behav. Neurosci. 2018, 12, 163. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.W.; Horng, J.L.; Tong, S.K.; Cherng, B.W.; Liao, B.K.; Lin, L.Y.; Chou, M.Y. Exposure to silver impairs learning and social behaviors in adult zebrafish. J. Hazard. Mater. 2021, 403, 124031. [Google Scholar] [CrossRef]
- Gutnick, A.; Blechmen, J.; Kaslin, J.; Herwig, L.; Belting, H.G.; Affolter, M.; Bonkowsky, J.L.; Levkowitz, G. The hypothalamic neuropeptide oxytocin is required for formation of the neurovascular interface of the pituitary. Dev. Cell 2011, 21, 642–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Li, C.; Zeng, Q.; Agrawal, I.; Zhu, X.; Gong, Z. Genome-wide identification of suitable zebrafish Danio rerio reference genes for normalization of gene expression data by RT-qPCR. J. Fish. Biol. 2016, 88, 2095–2110. [Google Scholar] [CrossRef]
- Walker, S.J.; Worst, T.J.; Vrana, K.E. Semiquantitative real-time PCR for analysis of mRNA levels. In Drugs of Abuse; Methods in Molecular Medicine; Wang, J.Q., Ed.; Humana Press: Totowa, NJ, USA, 2013; pp. 220–221. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Abercrombei, M. Estimation of nuclear population from microtome sections. Anat. Rec. 1946, 94, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlai, R.; Lahav, M.; Guo, S.; Rosenthal, A. Drinks like a fish: Zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects. Pharmacol. Biochem. Behav. 2000, 67, 773–782. [Google Scholar] [CrossRef]
Figure 1.
Distribution of oxyr and oxyrl in different tissues of adult zebrafish. The mRNA expression of target genes was analyzed through semi-quantitative PCR; rpl13a was used as an internal control to confirm the cDNA quality of different tissues.
Figure 1.
Distribution of oxyr and oxyrl in different tissues of adult zebrafish. The mRNA expression of target genes was analyzed through semi-quantitative PCR; rpl13a was used as an internal control to confirm the cDNA quality of different tissues.
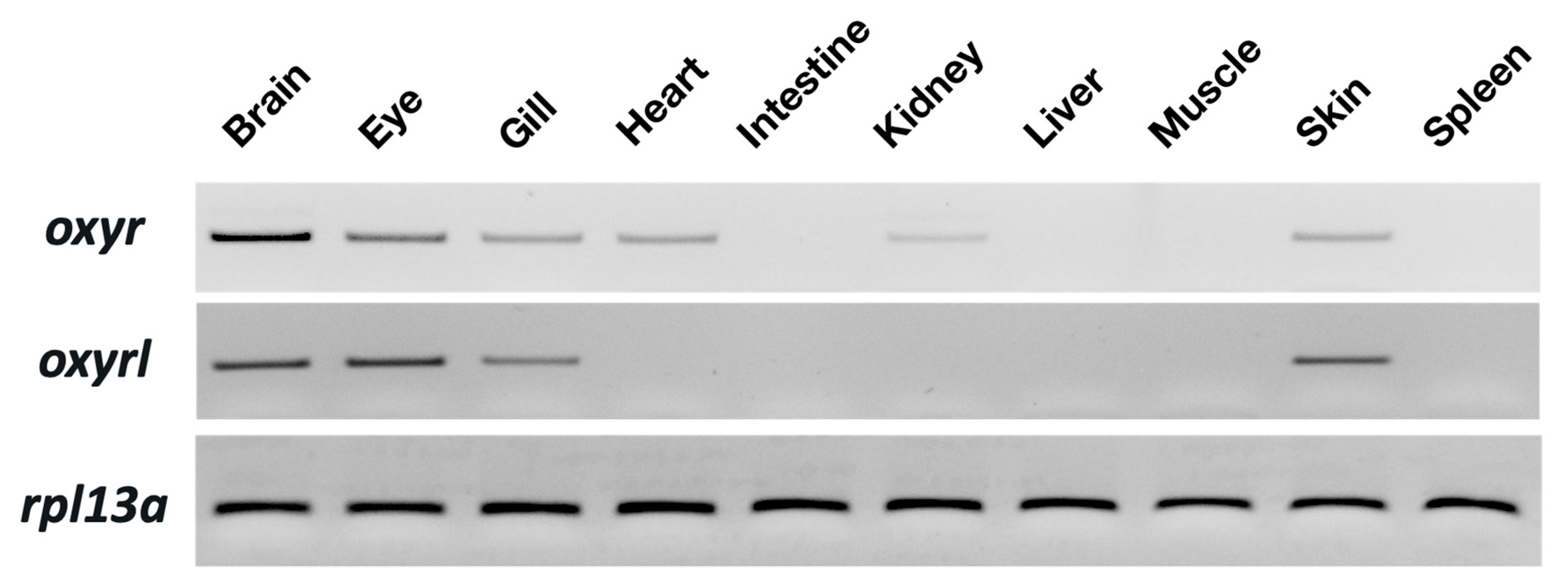
Figure 2.
Charts showing mRNA expression of oxyr and oxyrl in the gills of adult zebrafish acclimated to (A,B) acidic (AC), (C,D) high-ammonia (HA), or (E,F) double-deionized (DI) water for 7 days. Rpl13a was used as an internal control to normalize relative expression. Each circle represents the data of one single fish. The asterisks (*) indicate significant differences between the control and treatment groups. Values are means ± SD (n = 6, p < 0.05 (Student’s t-test)).
Figure 2.
Charts showing mRNA expression of oxyr and oxyrl in the gills of adult zebrafish acclimated to (A,B) acidic (AC), (C,D) high-ammonia (HA), or (E,F) double-deionized (DI) water for 7 days. Rpl13a was used as an internal control to normalize relative expression. Each circle represents the data of one single fish. The asterisks (*) indicate significant differences between the control and treatment groups. Values are means ± SD (n = 6, p < 0.05 (Student’s t-test)).

Figure 3.
Charts showing mRNA expression of oxyr and oxyrl in the brains of adult zebrafish acclimated to (A,B) AC; (C,D) HA, or (E,F) DI water for 7 days. Rpl13a was used as an internal control to normalize relative expression. Each circle represents the data of one single fish. The asterisks (*) indicate significant differences between the control and treatment groups; ns indicates that no significant difference was found between the control and treatment groups. Values are means ± SD (n = 6, p < 0.05, (Student’s t-test)).
Figure 3.
Charts showing mRNA expression of oxyr and oxyrl in the brains of adult zebrafish acclimated to (A,B) AC; (C,D) HA, or (E,F) DI water for 7 days. Rpl13a was used as an internal control to normalize relative expression. Each circle represents the data of one single fish. The asterisks (*) indicate significant differences between the control and treatment groups; ns indicates that no significant difference was found between the control and treatment groups. Values are means ± SD (n = 6, p < 0.05, (Student’s t-test)).

Figure 4.
Confocal laser scanning micrographs of the sections of the brain of transgenic zebrafish (oxtl:EGFP) acclimated to AC, HA, or DI water stained with anti-GFP (green; oxytocin) and Nissl (red) (A). Scale bar = 50 µm. The total numbers of oxytocin neurons in the brain of transgenic zebrafish treated with AC (B), HA (C), and DI (D) water were counted through the section sets from whole brain. Each circle represents the data from one single fish. In AC treatment, n = 11 for control group; n = 9 for the treatment group. In HA treatment, n = 5 for both control and treatment group. In DI treatment, n = 9 for control group; n = 5 for the treatment group. The asterisks (*) indicate significant differences between the control and treatment groups; ns indicates that no significant difference was found between the control and treatment groups. Values are mean ± SD (p < 0.05 (Student’s t-test)).
Figure 4.
Confocal laser scanning micrographs of the sections of the brain of transgenic zebrafish (oxtl:EGFP) acclimated to AC, HA, or DI water stained with anti-GFP (green; oxytocin) and Nissl (red) (A). Scale bar = 50 µm. The total numbers of oxytocin neurons in the brain of transgenic zebrafish treated with AC (B), HA (C), and DI (D) water were counted through the section sets from whole brain. Each circle represents the data from one single fish. In AC treatment, n = 11 for control group; n = 9 for the treatment group. In HA treatment, n = 5 for both control and treatment group. In DI treatment, n = 9 for control group; n = 5 for the treatment group. The asterisks (*) indicate significant differences between the control and treatment groups; ns indicates that no significant difference was found between the control and treatment groups. Values are mean ± SD (p < 0.05 (Student’s t-test)).
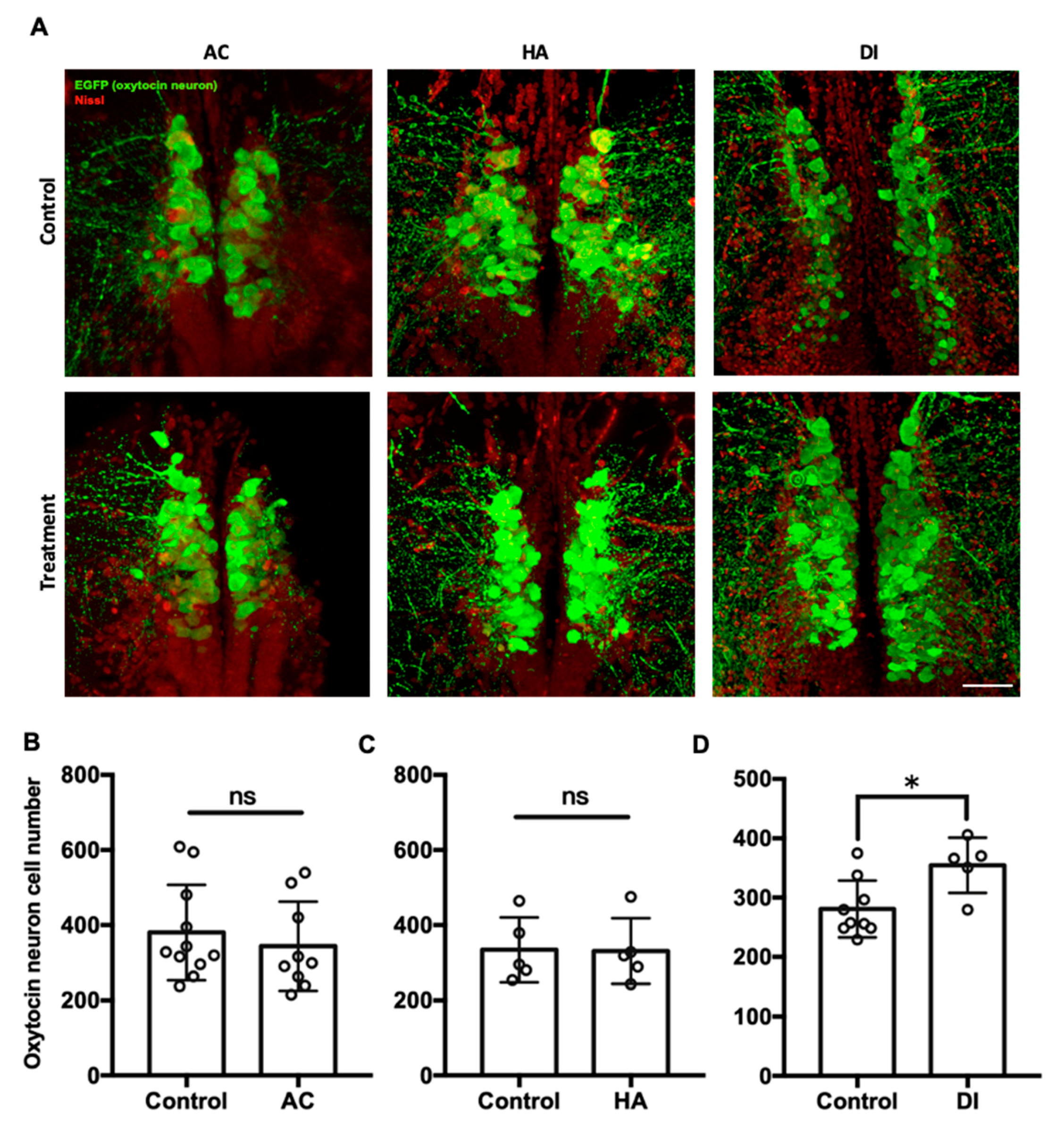
Figure 5.
Distribution and projection of oxytocin neurons in the adult zebrafish brain (n = 5). Whole-brain sections were stained with anti-GFP (green; oxytocin) and Nissl (red). (A,B) Sagittal sections showed that the fibers of oxytocin neurons were widely present in the brain. Somas were found in (D,E) PPa, (F,G) PPp, and (I) TPp. From the rostral end to the caudal end, the regions containing the fibers were found in (C) Vv, (D,E) PPa, (F) PPp, (G) A, VM, (H) PGm, (I) TPp, (J) Cans, (K) IPN, (L) raphe, and (M) TTBc. Scale bar = 100 µm.
Figure 5.
Distribution and projection of oxytocin neurons in the adult zebrafish brain (n = 5). Whole-brain sections were stained with anti-GFP (green; oxytocin) and Nissl (red). (A,B) Sagittal sections showed that the fibers of oxytocin neurons were widely present in the brain. Somas were found in (D,E) PPa, (F,G) PPp, and (I) TPp. From the rostral end to the caudal end, the regions containing the fibers were found in (C) Vv, (D,E) PPa, (F) PPp, (G) A, VM, (H) PGm, (I) TPp, (J) Cans, (K) IPN, (L) raphe, and (M) TTBc. Scale bar = 100 µm.

Figure 6.
Effects of AC, HA, and DI treatments in a novel tank diving test. (A) Schematic of the novel tank diving test. (B) Trajectories of fish moving after exposure to AC, HA, or DI water. (C,E,G) Total moving distance of fish was calculated after exposure to different stressors. (D,F,H) The period fish stayed in upper zone was analyzed after exposure to different stressors. In AC treatment, n = 11 for the treatment group, n = 8 for the treatment group. In HA treatment, n = 9 for control group; n = 11 for the treatment group. In DI treatment, n = 11 for control group; n = 13 for the treatment group. The asterisks (*) indicate significant differences between the control and treatment groups.; ns indicates that no significant difference was found between the control and treatment groups. Values are means ± SD (p < 0.05 (Student’s t-test)).
Figure 6.
Effects of AC, HA, and DI treatments in a novel tank diving test. (A) Schematic of the novel tank diving test. (B) Trajectories of fish moving after exposure to AC, HA, or DI water. (C,E,G) Total moving distance of fish was calculated after exposure to different stressors. (D,F,H) The period fish stayed in upper zone was analyzed after exposure to different stressors. In AC treatment, n = 11 for the treatment group, n = 8 for the treatment group. In HA treatment, n = 9 for control group; n = 11 for the treatment group. In DI treatment, n = 11 for control group; n = 13 for the treatment group. The asterisks (*) indicate significant differences between the control and treatment groups.; ns indicates that no significant difference was found between the control and treatment groups. Values are means ± SD (p < 0.05 (Student’s t-test)).

Figure 7.
Effects of AC, HA, and DI treatments in the mirror biting test. (A) Schematic of the mirror biting test. Adult zebrafish were exposed (B) AC, (C) HA, or (D) DI, and the mirror-biting frequency was determined. Each circle represents the data of one single fish. In AC treatment, n = 11 for control group; n = 10 for the treatment group. In HA treatment, n = 12 for control group; n = 8 for the treatment group. In DI treatment, n = 11 for control group; n = 10 for the treatment group. All the treatments reduced mirror-biting frequency relative to control fish. The asterisks (*) indicate significant differences between the control and treatment groups.; ns indicates that no significant difference was found between the control and treatment groups. Values are means ± SD in (A) and (C) (p < 0.05 (Student’s t-test)). Values are means ± SEM in (B) (p < 0.05 (Mann–Whitney)).
Figure 7.
Effects of AC, HA, and DI treatments in the mirror biting test. (A) Schematic of the mirror biting test. Adult zebrafish were exposed (B) AC, (C) HA, or (D) DI, and the mirror-biting frequency was determined. Each circle represents the data of one single fish. In AC treatment, n = 11 for control group; n = 10 for the treatment group. In HA treatment, n = 12 for control group; n = 8 for the treatment group. In DI treatment, n = 11 for control group; n = 10 for the treatment group. All the treatments reduced mirror-biting frequency relative to control fish. The asterisks (*) indicate significant differences between the control and treatment groups.; ns indicates that no significant difference was found between the control and treatment groups. Values are means ± SD in (A) and (C) (p < 0.05 (Student’s t-test)). Values are means ± SEM in (B) (p < 0.05 (Mann–Whitney)).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The information of primer sets used in the present study.
| Genes | Primer Sequence (5′–3′) | Amplicon Size | EF | Ensemble ID |
|---|---|---|---|---|
| oxyr | F: TTCAGCATCCCGCAGGTTTA | 123 bp | 95.4% | ENSDART00000176856.2 |
| R: GCACTGGTCCCTCTTCGTCTT | ||||
| oxyrl | F: ACGCCCTTCTTCTTCGTTCAG | 147 bp | 98.4% | ENSDART00000064853.4 |
| R: TATTTCTCCAGTGCCTCTTACAGC | ||||
| rpl13a | F: CCTCGGTCGTCTTTCCGCTATTG | 247 bp | 95.2% | ENSDART00000180298.1 |
| R: CAGCCTGACCCCTCTTGGTTTTG |
F, forward primer; R, reverse primer; EF, efficiency of primer pair in semi-quantitative PCR and RT-qPCR.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chuang, H.-J.; Chang, C.-Y.; Ho, H.-P.; Chou, M.-Y. Oxytocin Signaling Acts as a Marker for Environmental Stressors in Zebrafish. Int. J. Mol. Sci. 2021, 22, 7459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147459
AMA Style
Chuang H-J, Chang C-Y, Ho H-P, Chou M-Y. Oxytocin Signaling Acts as a Marker for Environmental Stressors in Zebrafish. International Journal of Molecular Sciences. 2021; 22(14):7459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147459
Chicago/Turabian StyleChuang, Hsin-Ju, Chun-Yung Chang, Huai-Ping Ho, and Ming-Yi Chou. 2021. "Oxytocin Signaling Acts as a Marker for Environmental Stressors in Zebrafish" International Journal of Molecular Sciences 22, no. 14: 7459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147459
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.