
Neural Stem Cells for Early Ischemic Stroke

1
Department of Pharmacology, Tulane University School of Medicine, 1430 Tulane Ave, New Orleans, LA 70112, USA
2
Department of Physiology, Tulane University School of Medicine, 1430 Tulane Ave, New Orleans, LA 70112, USA
3
Tulane Brain Institute, Tulane University, 1430 Tulane Ave, New Orleans, LA 70112, USA
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(14), 7703; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147703
Submission received: 23 June 2021
/
Revised: 14 July 2021
/
Accepted: 15 July 2021
/
Published: 19 July 2021
(This article belongs to the Special Issue Molecular Mechanisms of Cerebrovascular Diseases)
{kind=link}
{kind=link}
Abstract
:Clinical treatments for ischemic stroke are limited. Neural stem cell (NSC) transplantation can be a promising therapy. Clinically, ischemia and subsequent reperfusion lead to extensive neurovascular injury that involves inflammation, disruption of the blood-brain barrier, and brain cell death. NSCs exhibit multiple potentially therapeutic actions against neurovascular injury. Currently, tissue plasminogen activator (tPA) is the only FDA-approved clot-dissolving agent. While tPA’s thrombolytic role within the vasculature is beneficial, tPA’s non-thrombolytic deleterious effects aggravates neurovascular injury, restricting the treatment time window (time-sensitive) and tPA eligibility. Thus, new strategies are needed to mitigate tPA’s detrimental effects and quickly mediate vascular repair after stroke. Up to date, clinical trials focus on the impact of stem cell therapy on neuro-restoration by delivering cells during the chronic stroke stage. Also, NSCs secrete factors that stimulate endogenous repair mechanisms for early-stage ischemic stroke. This review will present an integrated view of the preclinical perspectives of NSC transplantation as a promising treatment for neurovascular injury, with an emphasis on early-stage ischemic stroke. Further, this will highlight the impact of early sub-acute NSC delivery on improving short-term and long-term stroke outcomes.
1. Introduction
Stroke remains a leading cause of death and long-term disability in the United States, and ischemic stroke accounts for 87% of all strokes [1]. Aging is one of the primary risk factors for neurovascular diseases, and overall, two-thirds of strokes afflict patients over 65 years of age [2].
Clinically, ischemia-reperfusion (IR) leads to extensive neurovascular injury and neurological dysfunction [3,4]. Although there is a high incidence of ischemic stroke, treatment options are limited to mechanical endovascular treatment (thrombectomy) [5,6] and intravenous thrombolysis by tissue plasminogen activator (tPA) [7]. Moreover, major limitations of tPA treatment include a narrow therapeutic window within 4.5 h after stroke onset [8], and a greater potential for hemorrhagic transformation [9].
Strategies extending tPA’s and thrombectomy’s narrow time windows have been explored [10,11]. While thrombectomy has shown to be beneficial up to 24 h in a highly selected sub-group of patients by perfusion imaging [12,13,14,15], the outcome of tPA treatment between 4.5 h and 9 h post-stroke onset in selected patients with salvageable tissue is inconclusive [11]. However, delaying reperfusion might cause greater disruption of the blood-brain barrier (BBB), which is a promising target for reducing stroke injury. Preclinical studies show that IR triggers 2 episodes of BBB disruption: the first is reversible, but the second, which occurs following 24 to 72 h of ischemic stroke, is irreversible and contributes significantly to cell death [16]. Therefore, new strategies are needed to extend tPA’s therapeutic window, minimize deleterious effects, and improve stroke outcome.
Neural stem cells (NSCs) possess pleiotropic effects that are beneficial for early stroke pathophysiology, which is highly complex. Engrafted NSCs can differentiate into functional neurons in the brain [17], and also exhibit bystander (chaperone) effects that include delivery of neurotrophic factors, which could mitigate the toxic microenvironment and protect endangered host cells [18,19,20,21,22]. Preclinical studies demonstrate that engrafted NSCs improve stroke outcome through multiple mechanisms, such as protection of the BBB, decreased cerebral vascular inflammation, increased neurogenesis and angiogenesis, and enhanced neurological function [23,24]. Furthermore, therapeutic outcomes of NSCs can be different based on time and route of NSC administration. While current clinical trials focus on the outcome of stem cells on neuro-restoration by injecting cells during the chronic stroke stage [25], this review will focus on the therapeutic mechanisms and potential of NSC transplantation in the early (subacute) phase of ischemic stroke, thus improving long-term outcome.
2. Biology of Neural Stem Cells
2.1. Endogenous Neural Stem Cells
NSCs have functional properties of self-renewal and multipotency to generate neurons, astrocytes, and oligodendrocytes in the central nervous system (CNS). In mammals, NSCs are present in neurogenic “niches” [26] that include the subgranular zone (SGZ) in the dentate gyrus of the hippocampus [27] and the subventricular zone (SVZ) of the lateral ventricles [28,29,30]. While these areas are especially enriched in NSCs, experimental evidence suggests that neurogenesis can also occur in other brain regions such as the amygdala and the hypothalamus [31,32,33]. In the SVZ, along the walls of the lateral ventricles, progenitor cells proliferate, differentiate and migrate along the rostral migratory system [26]. With aging, there is a significant decrease in SVZ neurogenesis accompanied by changes in the niche, including diminished thickness [34], changes in the choroid plexus secretome [35], and increased microglial activation [36]. Furthermore, 3D image analysis reveals more significant vascular changes and NSC deficits in males, suggesting sex differences [37]. Intriguingly, several differences between the rodent and human SVZ have been reported [38]. NSCs are predominantly quiescent in healthy adults and neurogenesis is tightly regulated by the neurogenic niche [39,40]. Pathological insults such as ischemia stimulate neurogenesis in the brain [41,42,43]. However, injury-induced neurogenesis does not provide sufficient cells necessary for repairing extensive tissue damage after cerebral infarction [42,44]. Therefore, NSC transplantation could be a promising therapy to repair ischemia-induced neurovascular damage [19,45,46,47,48,49,50,51].
2.2. NSC Derivation
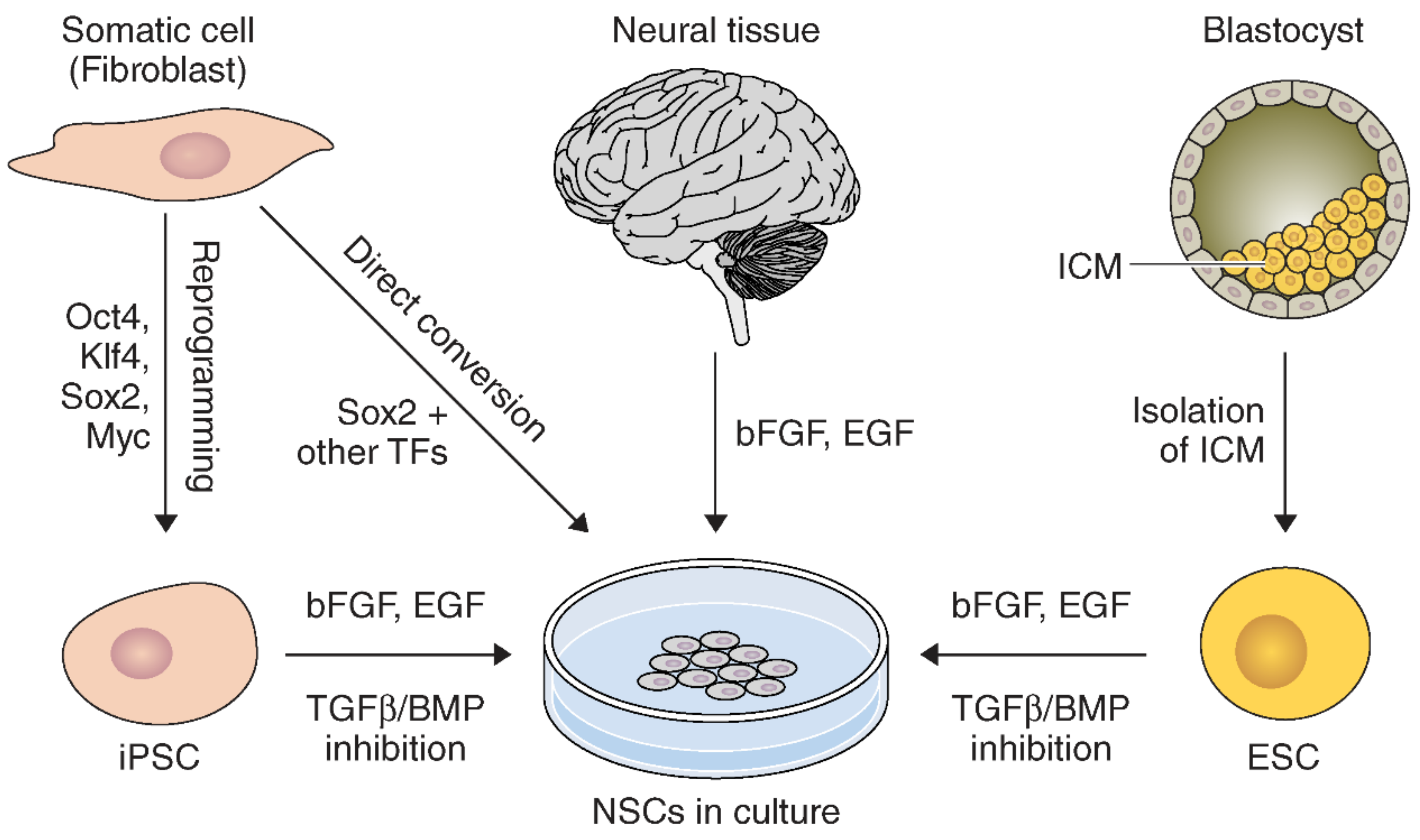
Neural stem cells can be derived by many approaches from various sources (Figure 1). NSCs can be directly harvested from neuroectoderm in fetal tissue, or the SVZ and SGZ in adults [52]. These primary NSCs can be expanded and maintained in culture using basic fibroblast growth factor (bFGF) and epidermal growth factor (EGF) [52].
NSCs can also be derived from other sources. For example, NSCs can be generated from embryonic stem cells (ESCs) [53,54,55]. However, a great deal of manipulation and intermediate steps are required for ESCs to fully differentiate into NSCs [53,55]. Neuroinduction of ESCs in culture can be achieved by inhibiting TGFβ/BMP signaling pathways during expansion with bFGF and EGF [56].
Similarly, NSCs can be generated from induced pluripotent stem cells (iPSCs) [56,57]. Many types of somatic cells can be readily obtained to dedifferentiate into iPSCs. These include fibroblasts, keratinocytes [58], blood [59], and hair follicles [60]. Notably, a similar methodology using dual-inhibiting SMAD signaling for NSC derivation from ESCs can generate iPSCs into NSCs [56]. However, generating iPSCs requires extra reprogramming to dedifferentiate somatic cells [61]. Microarray analysis studies confirmed that iPSC-NSCs and ESC-NSCs have very similar but not identical genetic expression profiles [62,63]. Use of iPSCs is advantageous due to fewer ethical concerns and pathological immune responses, since they can be generated by reprogramming from the patient’s own tissue [57]. iPSC-NSCs have been reported to be therapeutic in preclinical neurological disease models, which raises hope as a clinically promising source for cell therapy.
Direct conversion methods for reprograming somatic cells into induced NSCs (iNSCs) have been developed [64,65]. For instance, mouse fibroblasts were successfully transformed into NSCs by the introduction of pluripotency factors Oct4, Sox2, Klf4, and c-Myc [64]. Similarly, NSCs can be generated by constitutively inducing Sox2, Klf4, and c-Myc while stringently limiting Oct4 activity to the initial phase of reprogramming [65]. Generation of iNSCs has been reported with different combinations of NSC-specific transcription factors as well [66,67]. Further, iNSCs can also be generated by a single transcription factor, Sox2 or ZFP521 from mouse and human fibroblasts [68,69]. Pharmacological reprogramming can be used to generate iNSCs by signaling-directed transcriptional activation [70]. A direct iNSC generation methodology provides a new strategy for the generation of NSCs through direct cell transdifferentiation and avoids the lengthy intermediate step to generate iPSCs. Thus, direct conversion of somatic cells to NSCs can be a more efficient strategy [65]. Further, iNSCs from the patient’s own cells lower the risk of rejection of iNSCs following transplantation [71]. Therefore, research advancements in direct differentiation of NSCs can be promising for future therapeutic applications against stroke.
2.3. Labeling and Tracking Exogenous NSCs
Various methods for labeling and tracing NSCs have been established. Direct labeling is a widely used method by introducing a marker into stem cells or onto the cell surface before transplantation. Prelabeling NSCs with the thymidine analog, bromodeoxyuridine (BrdU), which incorporates into the nuclei during the S phase, is a reliable method. Newly-divided BrdU+ NSCs can then be detected with an antibody [52]. This method is useful to quantify the number of divisions of engrafted NSCs following transplantation [52] and is preferentially used for short-term tracking studies because BrdU becomes diluted over time [52]. To trace engrafted NSCs using clinically translational magnetic resonance imaging (MRI), NSCs can be labeled with super-paramagnetic iron oxide particles (SPIO) [72]. Stem cells can also be effectively labeled with fluorescent probes (e.g., orange cell tracker CMTMR [73] or CM-Dil [74]) for optical imaging and also radiotracers for radionuclide imaging [75]. However, direct radiotracer labeling can lead to potential radiation damage to the cells.
NSCs can be labeled via viral transduction to express readily identifiable markers such as GFP and lacZ. However, this method raises concerns about transgene inactivation and altering the properties of stem cells [52]. When human NSCs (hNSCs) are engrafted into rodents (e.g., species mismatch), hNSCs can be identified by human-specific antibodies. Engrafted stem cells can also be verified by donor-to-recipient sex-mismatch [52].
2.4. Stem Cell Migration
NSCs can migrate to areas of injury and neurodegeneration in the CNS. Migration and engagement of NSCs with a pathologic niche is the first step in cell-mediated restoration of homeostasis to the injured region [18,76,77,78,79]. NSC pathotropism, which is the natural tendency of homing to the site of injury, is partly achieved by chemokine receptors on NSCs that respond to proinflammatory cytokines secreted from the pathologic region [80,81,82,83,84,85,86]. For example, chemokine stromal cell-derived factor-1α (SDF-1α/CXCL12) interacts with CXCR4 receptors on NSCs as a major step in directing NSCs to injured brain regions [74,76,83,87]. The role of SDF-1α in stem cell homing is also implicated in cardiovascular and renal disease [88,89,90]. Further, in vitro transwell chemotaxis experiments demonstrated that NSCs preferentially migrate toward a higher SDF-1α concentration [76,91] and also synthetic SDF-1α [76]. SDF-1α is increased in stroke-affected brain tissue, and NSCs migrated toward the infarct area [92]. Targeting the SDF-1α/CRCX4 signaling pathway could be a highly efficient strategy for increasing the migration capacity and number of NSCs that cover the stroke-afflicted site.
Stroke injury generates an irreversible “necrotic core” and also a salvageable “penumbra” that surrounds the core. The penumbral region is metabolically active and structurally intact for longer than the infarct core. Intracranial transplantation of NSCs into the penumbral parenchyma has been reported to be beneficial [52,78,79,93]. Also, migration of NSCs from the intravascular space to the pathologic niche has been shown after tail vein [94] or intra-arterial [95] injection. However, intravenous delivery of stem cells can be trapped in filtering organs including the lung and liver [96,97,98,99]. Thus, intra-arterial injection may provide a more direct route to the lesion and exhibit better stroke outcome [46,100]. Also, intranasally delivered stem cells have been reported to extensively migrate to areas afflicted by experimental hypoxia-ischemia and ischemic stroke [101,102].
3. Pathophysiology of Early Ischemic Stroke
3.1. Blood-Brain Barrier and Ischemia-Reperfusion Injury
Integrity of the BBB is very critical as it forms a physical barrier created by tight junction proteins between endothelial cells that strictly regulate transcellular trafficking [103,104,105,106]. The BBB is created by spatiotemporal coordination between neurons, astrocytes, vascular cells (e.g., endothelial cells and pericytes), and the extracellular matrix (ECM). This neurovascular unit (NVU) is crucial for maintaining CNS homeostasis [107,108,109]. The BBB also controls leukocyte trafficking into the CNS for immune surveillance and response to infections [110], or removal of tissue damage [111].
BBB dysfunction is a prominent pathophysiological feature of acute ischemic stroke. Acute ischemia results in rapid decrease of cerebral blood flow and causes metabolic dysregulation due to oxygen and glucose deprivation. Consequent decrease of energy supply leads to lactic acidosis, alterations in ion transport, and excess extracellular accumulation of glutamate [112], which results in endothelial swelling and BBB damage [113].
Restoration of cerebral blood flow supplying oxygen and nutrients is crucial to attenuating ischemic stroke damage. However, reperfusion following ischemia further challenges the BBB, which causes a biphasic opening of the BBB. Experimental studies report that the initial phase of the BBB opening is reversible and occurs within several hours after reperfusion [114], but the second opening is irreversible and occurs 24–72 h after reperfusion [115]. While the exact mechanisms underlying increased BBB permeability in the early IR stages remain under extensive investigation, accumulating evidence shows that IR promotes greater disruption of the BBB through reactive oxygen species (ROS) damage to cellular molecules, upregulation of inflammatory factors and matrix metalloproteinases (MMPs), and alterations of tight junction proteins (TJPs) [116,117,118]. The initial BBB opening is linked to subtle alterations of tight junction complexes [119,120], followed by a second opening involving enzymatic cleavage of TJPs [114,121,122].
3.2. MMPs
MMPs are a family of zinc-binding proteolytic enzymes that are capable of degrading components of the ECM [123]. Although MMPs play critical roles in angiogenesis, tissue remodeling, and wound healing [124,125], there is a growing body of research highlighting strong associations between unrestrained MMP activity, neuroinflammation, and progression of neurodegenerative disease [126,127,128,129,130,131]. MMPs play a dual role following ischemic stroke [132], enhancing neurovascular injury during the acute phase but playing a beneficial role during the chronic recovery state. Neuroinflammation-associated MMPs degrade perivascular basement membranes and TJPs of the BBB, which contribute to increased BBB leakiness and the passage of toxic substances into ischemic tissue [133]. Consequences of damage to tight junctions in the BBB include upregulation of proinflammatory cytokines and infiltration of peripheral immune cells into the CNS.
3.2.1. MMP-2 and MMP-9
Already, the involvement of MMP-2 and MMP-9 has been extensively studied in stroke. Following stroke, the levels of MMP-2 and MMP-9 are elevated in ischemic tissue [134,135] and they contribute to BBB disruption [136,137]. For example, prior studies show upregulation of MMP-2 in the acute stages of IR in rodents and nonhuman primates [138,139]. Also, during the first 24 h post-IR, there is a significant increase in the level of MMP-9, which is associated with more extensive damage to TJPs, especially zona occludens-1 (ZO-1), a protein that connects the actin cytoskeleton of microvascular endothelial cells (ECs) to occludin [137,140]. Further, previous studies have reported that MMP-9 activity is positively correlated with BBB breakdown after stroke [141] and linked to tPA-induced hemorrhage in stroke patients [142] and animal models [143,144]. TJPs, such as claudin-5, occludin, and ZO-1 are crucial for BBB integrity [120,145]. It has been shown that at least one of the mechanisms by which MMP-9 participates in BBB damage is through the degradation of specific TJPs [146]. Preclinical investigations using the well-established experimental stroke model, middle cerebral artery occlusion-reperfusion (MCAO/R), report that BBB leakage begins to occur 24 h post-MCAO/R, regardless of claudin-5 or occludin disruption [147]. However, MCAO/R does result in the destruction of ZO-1 [145]. Given that MMP-9 is known to degrade ZO-1 [137,148], MMP-9 inhibition may ameliorate proteolytic degradation of ZO-1 and preserve BBB integrity.
3.2.2. MMP-3 (Stromelysin-1)
MMP-3, a 51-kDa protein [149], is one of the major inducible MMPs that can activate latent pro-MMP-9 [150,151,152]. MMP-3 has broad substrate specificity and can degrade various ECM proteins including fibronectin, denatured collagens (gelatin), laminin, and proteoglycans [153], and is critical to brain tissue remodeling and wound healing. Injury-induced MMP-3 is upregulated within several hours after stroke, suggesting association with the initial opening of the BBB [154]. MMP-3 deficient mice exhibited reduced degradation of TJPs of the BBB and less neutrophil infiltration caused by intracerebral lipopolysaccharide (LPS)-induced BBB opening [128] or spinal cord injury [130] in young adult mice. Further, MMP-3 reportedly exacerbates tPA-induced intracerebral hemorrhage (ICH) post-stroke in thrombotic MCAO mouse brains [154]. While knockout of MMP-3 in stroke mice reduced the tPA-enhanced risk of ICH, delayed tPA administration (4 h post-MCAO) in the thrombotic ischemic stroke model was found to further augment MMP-3 expression selectively in ECs in the ischemic hemisphere, which suggests the involvement of MMP-3 in disruption of the BBB and ICH. While MMP-9 expression is also significantly upregulated in wild-type mouse ischemic brains, ICH was less prominent in MMP-9 KO mice. However, the location and levels of MMP-9 were not altered following tPA treatment 4 h post-MCAO, suggesting that MMP-3 may be a greater contributor to tPA-induced ICH than MMP-9 [154]. Combined laser microdissection and protein array studies showed significant upregulation of MMP-3 in the human ischemic stroke brain, along with MMP-9 [155].
In vitro, MMP-3 is upregulated either by tPA treatment or ischemic conditions in cultured murine brain ECs. However, this effect is attenuated by inhibition of either lipoprotein receptor-related protein (LRP), a scavenger receptor that can bind tPA [156], or nuclear factor-kappa B (NF-κB) activation, suggesting an underlying mechanism of MMP-3 induction via the LRP/NF-κB pathway [157]. While the role of MMP-3 in hemorrhagic transformation in rodents has been reported [154,158,159], the critical role of MMP-3 in BBB integrity and function remains understudied in cerebral IR injury post-stroke.
3.3. Inflammatory and Immune Responses after Stroke
Inflammation is a prominent feature of early stroke pathophysiology [160,161]. Stroke leads to increased BBB permeability that enables infiltration of neutrophils, macrophages, and T lymphocytes into the CNS [162]. Although inflammation is crucial for brain repair, an unchecked inflammatory response is detrimental in early ischemic stroke [163].
Microglia and macrophages play critical roles in modulating CNS repair [164]. Cerebral ischemic injury activates resident microglia that constantly survey their surroundings in the brain. Microglia exist in two functional phenotypic states and there is a dynamic interplay between their functional status responding to extracellular signals. For example, ischemic stroke promotes M1 microglial polarization, which leads to secretion of detrimental factors such as TNF-α, IL-1β, and ROS [165]. Alternatively, activation of M2 microglial polarization promotes the resolution of neuroinflammation by releasing neurotrophic factors and anti-inflammatory cytokines such as IL-10 and TGF-β [165,166].
Less than 24 h after stroke, peripheral immune cells start to infiltrate the brain through the compromised BBB [167]. Macrophages, the major inflammatory cell infiltrate during acute stroke [168], are highly plastic and can display functionally different phenotypes [169]. M1 macrophages increase CNS damage by secreting proinflammatory molecules including TNF-α, IL-8, and IL-12 [161,170]. Infiltrating M1 macrophages recruit neutrophils into the CNS through secretion of IL-8, which promote further inflammation and tissue damage by releasing NO, MMPs, and cathepsins [161]. Secreted factors by both macrophages and neutrophils constitute the initial inflammatory cascade after stroke. In contrast, inflammation-resolving M2 macrophages release anti-inflammatory cytokines such as TFG-β and IL-10 [161,171]. Further, M2 macrophages are also beneficial for stroke outcome by removing ischemic debris [170].
Neuroinflammation after stroke is eventually resolved and the M1-to-M2 shift occurs when proinflammatory mediators become further reduced by anti-inflammatory mediators [161]. The involvement of the STAT family is reported in regulating functional status of immune cells [172,173]. While STAT6 signaling drives M2 macrophage polarization [173,174], STAT1 activation promotes M1 macrophages [175]. IL-4, a STAT6 activator, drives microglia and macrophages toward a beneficial M2 phenotype and facilitates stroke recovery [176].
3.4. Ischemic Tissue Loss and Neurological Dysfunction
Stroke results in extensive cell death and tissue infarction in the affected region. The brain consumes high levels of oxygen and glucose and relies on oxidative phosphorylation for its energy source. Ischemia limits oxygen and glucose supplies thus, leading to significant loss of endogenous energy stores and disruption of ionic balance or neurotransmitter reuptake [182]. In particular, a metabolic insult after stroke leads to extracellular accumulation of glutamate, which activates NMDA and AMPA receptors, leading to high calcium influx and consequent cell death [183]. For example, Ca2+ overload drives mitochondrial function toward releasing cytochrome C, which activates cell death signaling pathways [184,185].
Edema is also major contributor of cell death and tissue infarction after ischemic stroke. Hyperactivation of glutamate receptors leads to increased Na+ and water influx, which causes hypotonic-induced cell swelling [183]. Furthermore, BBB damage enables the entry of foreign molecules, fluid, and immune cells into brain interstitial spaces, which promotes vasoactive edema and exacerbates tissue injury [186]. Although vasoactive edema is eventually resolved, clearance of debris and angiogenesis are often too slow to attenuate acute edema-linked brain injury after stroke.
4. Endogenous Repair Mechanisms
4.1. Angiogenesis
Spontaneous stroke recovery occurs [189,190] despite the fact the adult brain has limited endogenous repair capabilities [191]. Angiogenesis, the formation of new capillary blood vessels from preexisting vessels, occurs as a result of cerebral ischemia and can contribute to CNS plasticity and post-stroke recovery [192,193,194,195,196]. Endothelial cell proliferation occurs in the subacute stage of stroke and can continue for several weeks due to increased expression of angiogenesis genes and growth factors [197,198,199]. While angiogenesis can improve the repair process following stroke, the initial angiogenic vascular remodeling process is accompanied with a compromised endothelial barrier function. For example, vascular endothelial growth factor (VEGF) is the major initiator for stimulating angiogenesis and is highly upregulated as early as 1 h following cerebral ischemia. However, VEGF also promotes vascular permeability and barrier dysfunction [200]. Therefore, VEGF delivery aiming for increasing angiogenesis in early phase stroke can be both beneficial and detrimental. For instance, targeted local delivery in the subacute stage is better tolerated [201] but intraperitoneal systemic injection of VEGF in the early phase of stroke exacerbated BBB leakage, vasogenic edema and further increased infarct volume. Moreover, studies focusing on VEGF-induced BBB leakage demonstrated that blocking VEGF receptors after ischemia mitigates BBB permeability, brain edema, and reduces cerebral infarct volume [201]. Furthermore, regarding increased BBB permeability for initial stages of angiogenesis, high levels of MMPs such as MMP-2 and MMP-9 [202] has been associated with ECM remodeling during angiogenesis.
An angiogenic response is stimulated in the potentially salvageable penumbra for post-stroke recovery [192]. A higher vessel density in the penumbra is linked to improved survival after ischemic stroke [192,199,203], and further penumbra damage following stroke can have a catastrophic neurological outcome [204]. Angiogenic growth factors play an important role for cell survival in the penumbra [205]. In addition to enhancing local blood supply after stroke, angiogenesis also promotes the removal of necrotic brain tissue [206,207]. However, angiogenesis can be negatively influenced by several factors, including aging [208]. Given that stroke usually affects the aging population and clinical trials using stem cells show beneficial outcome in stroke patients [47], translational studies investigating the effects of neural stem cells on vascular remodeling during stroke in aged animals is greatly warranted.
4.2. Endogenous Neurogenesis
Endogenous neurogenesis is activated following stroke [42,44,49]. NSCs proliferate, generate new neurons and secrete trophic factors [41,42,50,189]. Further, neurogenesis and angiogenesis are coupled in the neurovascular unit in that endothelial cells release trophic factors that regulate both responses [209,210], and VEGF as a possible mediator for coupling angiogenesis and neurogenesis after stroke [210]. Microvessel characteristics also change after stroke [211], and neuroblasts migrate to the vascular remodeling site [212]. Additionally, NSCs enhance angiogenesis through trophic factors and can improve cerebral capillary blood flow [210,213]. These studies highlight stroke-induced coupling between neurogenesis and brain vascular remodeling. However, enhanced endogenous neurogenesis is not sufficient to yield substantial impact on repairing neuronal damage [42,214,215]. Therefore, transplantation of exogenous NSCs may be an effective strategy to reduce stroke brain damage.
Astrocytes also play a critical role in neurogenesis and angiogenesis by secreting neurotrophic and vascular growth factors [216,217]. Astrocytes can also mediate neurotransmission and help maintain neurovascular coupling [218]. During ischemia, astrocytes show neuroprotective effects including the removal of excess glutamate and enhancing revascularization [219,220,221]. However, in the chronic stroke phase, astrocytes can diminish neurological recovery by releasing growth inhibitory factors and forming glial scars, which lead to poor neuronal connectivity [222]. Thus, early stroke intervention with stem cells may reduce chronic stage stroke complications.
5. Transplantation of Pleiotropic Neural Stem Cells for Ischemic Stroke
There are prominent changes in the brain during initial ischemic insult. The timing of NSC transplantation is critical to the repair and regenerative therapeutic actions for ameliorating the complex, multifactorial pathophysiology of ischemic stroke. Engrafted NSCs release therapeutic trophic factors for brain tissue remodeling and facilitating neuronal plasticity [79,223,224]. NSC-secreted brain-derived neurotrophic factor (BDNF) [77,79] is a main neurotrophin that promotes neuroprotection, neurogenesis, and enhances neurological outcome following stroke [225,226,227,228]. Also, neurotrophic factors (NTFs) assist in maintaining neuronal health, ECM remodeling, and cell proliferation. NTFs also protect neuronal tissue from extensive damage [229]. VEGF is an important neurotrophic factor that facilitates angiogenesis and is involved in neural tissue repair [205,230]. Further, VEGF regulates mitogenesis and survival for vascular endothelial cells [231,232] and provides neuroprotective effects against ischemic injury [205,233,234]. When VEGF is pharmacologically inhibited using SU1498 and Flt-1Fc, NSC-mediated protection is dramatically thwarted [235]. Additionally, other neurotrophic factors such as ciliary neurotrophic factor, glial cell-line derived neurotrophic factor, and neural growth factor contribute to neuroprotection and repair [236,237,238,239,240,241]. In addition to secreting NTFs, engrafted NSCs generate electrophysiologically active functional neurons that make appropriate synaptic connections with host neurons [242].
5.1. Transplantation of NSCs for Early Stroke Intervention
Mostly, pharmacological-based therapy has been implemented to reduce acute/sub-acute stroke injury. For instance, minocycline, a commonly used semi-synthetic tetracycline antibiotic, has demonstrated potential as a safe, efficacious neuroprotectant for clinical treatment of stroke [243,244], partly attributed to reducing inflammation, cell death, and BBB damage [245,246,247,248,249,250]. However, NSC administration at the subacute stage may be used as part of a combinatorial strategy to protect neurons by reducing brain injury and cell death post-stroke. Preclinical studies report that NSC transplantation 24 h post-stroke leads to reduced inflammation and BBB damage, and better functional stroke outcome in mice [51,77,78,79]. The beneficial actions of NSC transplantation during early stroke are highlighted below.
5.1.1. BBB Support and MMPs
The BBB has great functional significance for protecting against the entry of neurotoxic agents and inflammatory mediators. Thus, preserving BBB integrity and function is a promising strategy for ischemic stroke. Early NSC delivery ameliorates BBB injury post-stroke. Intracranial hNSC administration in mice 24 h post-stroke leads to robust NSC migration to the injured region, decreased infarct size, and lessened BBB disruption [79]. This rapid therapeutic response of NSCs clearly suggests anti-inflammatory mechanisms of action. Consistent with these findings, hNSC-engrafted mice showed decreased microglia activation and reduced expression of IL-6, IL-1β, and MIP-1α [79]. Also, intracranial administration of hiPSC-NSCs in mice 24 h following stroke leads to decreased leakage of IgG into the brain parenchyma [78].
During ischemia reperfusion, MMPs are significantly upregulated in affected tissue and degrade the ECM. Higher MMP-9 activity is correlated with BBB disruption during stroke [148,251] and dysfunction of BBB tight junctions [78]. However, MMP-9 activity after stroke was decreased in NSC-transplanted young adult [78,79] and aged mice [77]. For instance, Western blot analysis shows that transplanted hNSCs attenuated MMP-9 at 48 h post-stroke, suggesting that hNSCs can reduce BBB disruption [79]. This finding was validated by gel zymography that measures MMP-9 enzyme activity. While MMP-9 activity is upregulated at 48 h post-stroke, engrafted hNSCs significantly reduced MMP-9 activity. In contrast, MMP-2 was low 48 h post-MCAO/R, implying that MMP-2 may not be a significant contributor at this time point. MMP-9 induction is associated with tPA-induced hemorrhage in stroke patients [142] and animal models [143,144]. MMP-9 is further elevated by delayed tPA treatment and associated with exacerbated BBB breakdown during stroke [141,148,251]. Aged mouse stroke brains show that hNSC administration reduces MMP-9 following delayed tPA treatment [77], suggesting BBB protection.
Tight junction proteins are essential in modulating BBB integrity [120,145]. However, cerebral ischemia leads to destruction of the tight junction protein, ZO-1 [145]. MMP-9 is known to degrade ZO-1 [137,148] thus hNSC-mediated MMP-9 downregulation may reduce ZO-1 degradation and consequently preserve BBB integrity. Indeed, hNSC-transplantation in mice showed higher ZO-1 [77], suggesting hNSCs reduced degradation of ZO-1. Since delayed tPA is known to augment the severity of neural cell death and BBB damage, adjuvant therapy with hNSCs after tPA administration may be an effective strategy to not only retain thrombolysis for acute ischemic stroke but also reduce BBB disruption in the early stage of stroke.
5.1.2. Inflammation
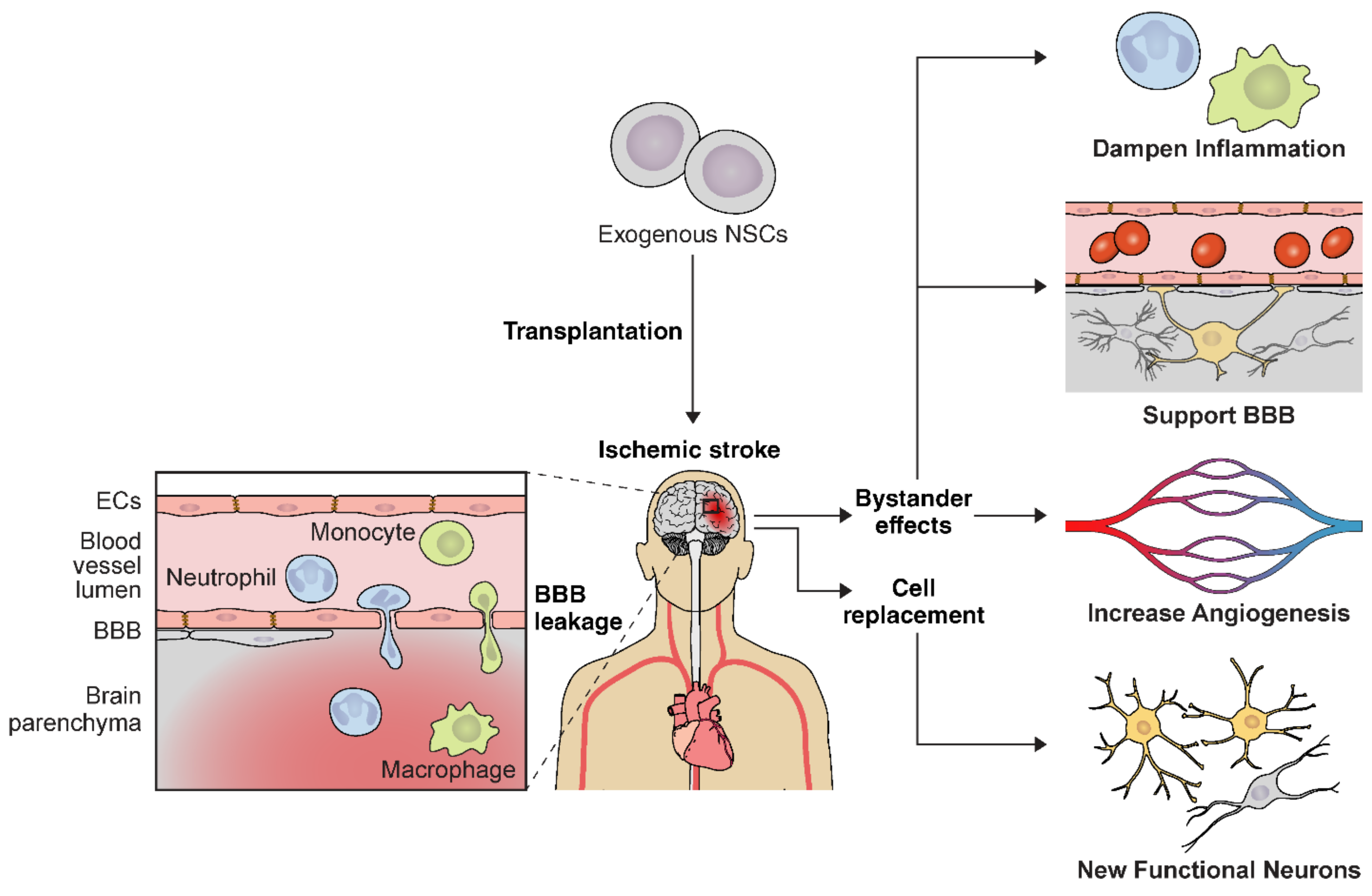
Ischemic stroke activates astrocytes and microglia and leads to inflammatory cell infiltration in the affected tissue due to increased BBB permeability. NSC delivery in the early stroke phase attenuates this complex inflammatory signaling cascade [223,252,253] (Figure 2). NSCs ameliorate ischemic damage by lowering proinflammatory mediators including TNF-α, IL-1β, IL-6, MCP-1, and Iba-1 [223,253]. For instance, hiPSC-NSC transplantation is associated with significantly decreased Iba-1 positive cells in the stroke mouse model [254]. Transplanted mice also had decreased numbers of GFAP-positive astrocytes [254]. Similarly, NSC-engrafted mice displayed lower CD45+ and Iba-1+/major histocompatibility II immune cells after ischemia [253]. Collectively, these studies demonstrate an important role of NSCs in BBB integrity and functional recovery from cerebral ischemia through influencing the extracellular microenvironment and reducing neuroinflammation. Thus, implementing stem cell therapies for immunomodulation and targeting the proinflammatory signaling cascades in the brain may be an effective therapeutic strategy for early ischemic stroke.
5.2. Long-Term Outcome of NSC Transplantation at the Early Stroke Phase
NSC transplantation in the early acute/subacute phases of stroke aims for neuroprotection from the deleterious extracellular surroundings after ischemia. Further, NSC delivery during the subacute phase will also stimulate neurorestorative mechanisms to benefit long-term outcome.
5.2.1. Angiogenesis
NSC transplantation promotes stroke recovery by stimulating angiogenesis, which is coupled to neurogenesis [209]. Early hNSC delivery post-stroke enhanced angiogenesis in rats [255]. When assessed at 14d after NSC delivery, BrdU+/vWF+ proliferating endothelial cells were increased in the afflicted area of NSC-engrafted rats, demonstrating increased angiogenesis [255]. Also, transplantation of NSCs overexpressing Cu/Zn-superoxide dismutase (SOD1) into peri-infarct mouse cortex 2d post-stroke enhanced angiogenesis, which is potentially mediated by upregulation of VEGF [256].
5.2.2. Cell Replacement
Early delivery of NSCs may facilitate stroke recovery through generation of new neurons or enhancing the neurogenic response. For instance, engrafted hNSCs into the rat striatum 2d post-stroke differentiated into neurons at 6 weeks and enhanced endogenous neurogenesis [257]. Similarly, other studies reported direct neural replacement after early NSC delivery during stroke. Engrafted hNSCs gave rise to neurons and astrocytes in the rat brain when they were implanted one day post-stroke [258,259]. When assessed at 30d post-transplant, the majority of engrafted iPSC-NSCs at 24 h post-stroke remained as undifferentiated NSCs, and only a modest amount of hiPSC-NSCs were positive for the neuronal marker (TuJ-1) [78]. Based on these findings, direct neural replacement may not be a primary reason for the beneficial effects of early NSC administration post-stroke.
5.2.3. Improved Neurological Outcome
Improved neurological behavior is considered to be the gold standard for evaluating long-term stroke outcome. Modified Neurologic Severity Score (mNSS) tests revealed improved neurological function two weeks post-transplant when hNSCs were engrafted at 24 h in rat stroke brains [255]. Similarly, intraventricular transplantation of rat NSCs overexpressing HIF-1α [260] or intravenous injection of mouse NSCs overexpressing bFGF [261] into rat brains 24 h after MCAO promoted behavioral recovery as assessed over 4 weeks by NSS scoring. Also, neurological function rapidly improved by 2d post-stroke when mice were engrafted with hNSCs 24 h post-MCAO [79]. These hNSC-engrafted mice displayed improved sensory motor function when assessed by the forepaw adhesive removal test [262]. In addition, engrafted mice show improved balance and motor coordination as evaluated by the beam walk and rotarod tests. Improved neurological outcome persisted when assessed for a month [79], suggesting that early stroke intervention with NSCs can lead to improved long-term neurological outcomes in ischemic stroke patients. Other studies also reported that intracranial iPSC-NSC delivery 24 h post-stroke leads to improved long-term neurological function [78]. Moreover, NSCs engrafted into the hippocampus expedited rapid migration of NSCs to injury sites and enhanced stroke recovery [78,79]. Another study using the murine NSC cell line, MHP36, showed greater functional recovery after intracranial injection 2d post-stroke in mice [263]. hNSC-engrafted animals showed better behavioral function when monitored for 28d post-stroke [263]. Furthermore, transplantation of hNSCs overexpressing Bcl2l1 into the infarct site 2d after permanent MCAO showed improved neurological function in mice [264]. Also, intracranial administration of SOD1-overexpressing NSCs 2d post-stroke improved behavioral performance as assessed over 28d by mNSS scoring and the rotarod test [256]. Based on these findings, early intervention of stroke with NSC administration clearly shows significant improvement for long-term neurological outcome and behavioral function.
6. Conclusions
To address the complex pathophysiology of neurovascular diseases, therapeutic strategies include pharmacologic, genetic, and cell-based tissue engineering. NSCs can be used alone or in combination with other interventions that can work synergistically. Specifically, NSCs possess anti-inflammatory actions that attenuate delayed tPA-associated adverse effects of early stroke. Further, diabetes increases the risk for stroke and worsens overall stroke outcome [265,266]. Diabetic patients are not eligible for tPA treatment due to greater risk of BBB permeability and hemorrhagic transformation caused by complications of ischemic stroke [265]. Diabetic db/db stroke mice show a further increase in the MMP-9 mRNA level and activity and inflammation [267]. Therefore, new therapeutic strategies to treat stroke patients with comorbidities are needed. Stem cell-based therapy for human trials is currently focusing on enhancing stroke rehabilitation by engrafting stem cells during the stroke recovery phase [47,268,269]. Phase 1 and 2 clinical trials delivering human NSCs during the chronic stroke phase were reported to show no safety concerns and improve motor recovery [268,270]. However, stem cell delivery during the sub-acute stroke phase may benefit more patients by ameliorating early stroke injury and consequently improving later stroke outcome. Therefore, deeper insight into early-phase stroke injury and identifying optimal stem cell strategies are needed for successful translation.
Author Contributions
J.-P.L. and M.H.H. drafted, edited and approved the final version of the manuscript. Both authors have read and agreed to the published version of the manuscript.
Funding
Jean-Pyo Lee is supported by the National Institute of Neurological Disorders and Stroke (Grant R01NS110370).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
Conflicts of Interest
The authors have no conflict of interest to disclose.
Abbreviations
| Abbreviation | Description |
| AMPA | α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| BBB | Blood-brain barrier |
| BDNF | Brain-derived neurotrophic factor |
| CNS | Central nervous system |
| CXCR4 | C-X-C chemokine type 4 receptor |
| ECASS-4 | European Cooperative Acute Stroke Study-4 |
| ECM | Extracellular matrix |
| EGF | Endothelial growth factor |
| ESC | Embryonic stem cell |
| FGF | Fibroblast growth factor |
| GFP | Green fluorescent protein |
| ICH | Intracerebral hemorrhage |
| IL-1β | Interleukin-1β |
| iPSC | Induced pluripotent stem cell |
| IR | Ischemic reperfusion |
| LRP | Lipoprotein receptor-related protein |
| MCAO | Middle cerebral artery occlusion |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MIP-1α | Macrophage inflammatory proteins |
| MMP | Matrix metalloproteinase |
| NF-κB | Nuclear factor kappa B |
| NMDA | N-Methyl-D-aspartic acid or N-methyl-D-aspartate |
| NSC | Neural stem cell |
| NTF | Neurotrophic factor |
| NVU | Neurovascular unit |
| ROS | Reactive oxygen species |
| SDF-1α | Stromal cell derived factor 1-α |
| STAT | Signal transducer and activator of transcription |
| SVZ | Subventricular zone |
| TIMP-2 | Tissue inhibitor of metalloproteinases 2 |
| TJPs | Tight junction proteins |
| TNF-α | Tumor necrosis factor |
| tPA | Tissue plasminogen activator |
| VEGF | Vascular endothelial growth factor |
| vWF | von Willebrand factor |
| ZO-1 | Zonula occludens-1 (ZO-1) |
References
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; de Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circulation 2017, 135, e146–e603. [Google Scholar] [CrossRef] [PubMed]
- Feigin, V.L.; Forouzanfar, M.H.; Krishnamurthi, R.; Mensah, G.A.; Connor, M.; Bennett, D.A.; Moran, A.E.; Sacco, R.L.; Anderson, L.; Truelsen, T.; et al. Global and regional burden of stroke during 1990–2010: Findings from the Global Burden of Disease Study 2010. Lancet 2014, 383, 245–254. [Google Scholar] [CrossRef]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The Science of Stroke: Mechanisms in Search of Treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef] [Green Version]
- Thom, T.; Haase, N.; Rosamond, W.; Howard, V.J.; Rumsfeld, J.; Manolio, T.; Zheng, Z.J.; Flegal, K.; O’Donnell, C.; Kittner, S.; et al. Heart disease and stroke statistics--2006 update: A report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation 2006, 113, e85–e151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sporns, P.B.; Minnerup, J.; Warneke, N.; Dziewas, R.; Hanning, U.; Berkemeyer, S.; Zoubi, T.; Heindel, W.; Schwindt, W.; Niederstadt, T. Impact of the Implementation of Thrombectomy with Stent Retrievers on the Frequency of Hemicraniectomy in Patients with Acute Ischemic Stroke. Clin. Neuroradiol. 2017, 27, 193–197. [Google Scholar] [CrossRef]
- Wahlgren, N.; Moreira, T.; Michel, P.; Steiner, T.; Jansen, O.; Cognard, C.; Mattle, H.P.; van Zwam, W.; Holmin, S.; Tatlisumak, T.; et al. Mechanical thrombectomy in acute ischemic stroke: Consensus statement by ESO-Karolinska Stroke Update 2014/2015, supported by ESO, ESMINT, ESNR and EAN. Int. J. Stroke 2016, 11, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, T.; Deguchi, K.; Nagotani, S.; Abe, K. Vascular protection and restorative therapy in ischemic stroke. Cell Transplant 2011, 20, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.M. Recombinant tissue plasminogen activator for the treatment of acute ischemic stroke. Proc. Baylor Univ. Med. Center 2011, 24, 257–259. [Google Scholar] [CrossRef]
- Kilic, E.; Bahr, M.; Hermann, D.M. Effects of recombinant tissue plasminogen activator after intraluminal thread occlusion in mice: Role of hemodynamic alterations. Stroke 2001, 32, 2641–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri, H.; Bluhmki, E.; Bendszus, M.; Eschenfelder, C.C.; Donnan, G.A.; Leys, D.; Molina, C.; Ringleb, P.A.; Schellinger, P.D.; Schwab, S.; et al. European Cooperative Acute Stroke Study-4: Extending the time for thrombolysis in emergency neurological deficits ECASS-4: ExTEND. Int. J. Stroke 2016, 11, 260–267. [Google Scholar] [CrossRef]
- Ringleb, P.; Bendszus, M.; Bluhmki, E.; Donnan, G.; Eschenfelder, C.; Fatar, M.; Kessler, C.; Molina, C.; Leys, D.; Muddegowda, G.; et al. Extending the time window for intravenous thrombolysis in acute ischemic stroke using magnetic resonance imaging-based patient selection. Int. J. Stroke 2019, 14, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, R.G.; Jadhav, A.P.; Haussen, D.C.; Bonafe, A.; Budzik, R.F.; Bhuva, P.; Yavagal, D.R.; Ribo, M.; Cognard, C.; Hanel, R.A.; et al. Thrombectomy 6 to 24 Hours after Stroke with a Mismatch between Deficit and Infarct. N. Engl. J. Med. 2018, 378, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Albers, G.W.; Marks, M.P.; Lansberg, M.G. Thrombectomy for Stroke with Selection by Perfusion Imaging. N. Engl. J. Med. 2018, 378, 1849–1850. [Google Scholar] [CrossRef]
- Albers, G.W.; Marks, M.P.; Kemp, S.; Christensen, S.; Tsai, J.P.; Ortega-Gutierrez, S.; McTaggart, R.A.; Torbey, M.T.; Kim-Tenser, M.; Leslie-Mazwi, T.; et al. Thrombectomy for Stroke at 6 to 16 Hours with Selection by Perfusion Imaging. N. Engl. J. Med. 2018, 378, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Powers, W.J.; Rabinstein, A.A.; Ackerson, T.; Adeoye, O.M.; Bambakidis, N.C.; Becker, K.; Biller, J.; Brown, M.; Demaerschalk, B.M.; Hoh, B.; et al. Guidelines for the Early Management of Patients With Acute Ischemic Stroke: 2019 Update to the 2018 Guidelines for the Early Management of Acute Ischemic Stroke: A Guideline for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke 2019, 50, e344–e418. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Ting, P.; Martinez, H.; Klatzo, I. The biphasic opening of the blood-brain barrier to proteins following temporary middle cerebral artery occlusion. Acta Neuropathol. 1985, 68, 122–129. [Google Scholar] [CrossRef]
- Yandava, B.D.; Billinghurst, L.L.; Snyder, E.Y. “Global” cell replacement is feasible via neural stem cell transplantation: Evidence from the dysmyelinated shiverer mouse brain. Proc. Natl. Acad. Sci. USA 1999, 96, 7029–7034. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.P.; Jeyakumar, M.; Gonzalez, R.; Takahashi, H.; Lee, P.J.; Baek, R.C.; Clark, D.; Rose, H.; Fu, G.; Clarke, J.; et al. Stem cells act through multiple mechanisms to benefit mice with neurodegenerative metabolic disease. Nat. Med. 2007, 13, 439–447. [Google Scholar] [CrossRef]
- Park, K.I.; Teng, Y.D.; Snyder, E.Y. The injured brain interacts reciprocally with neural stem cells supported by scaffolds to reconstitute lost tissue. Nat. Biotechnol. 2002, 20, 1111–1117. [Google Scholar] [CrossRef]
- Jeyakumar, M.; Lee, J.P.; Sibson, N.R.; Lowe, J.P.; Stuckey, D.J.; Tester, K.; Fu, G.; Newlin, R.; Smith, D.A.; Snyder, E.Y.; et al. Neural Stem Cell Transplantation Benefits a Monogenic Neurometabolic Disorder During the Symptomatic Phase of Disease. Stem Cells 2009, 27, 2362–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ourednik, J.; Ourednik, V.; Lynch, W.P.; Schachner, M.; Snyder, E.Y. Neural stem cells display an inherent mechanism for rescuing dysfunctional neurons. Nat. Biotechnol. 2002, 20, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Redmond, D.E., Jr.; Bjugstad, K.B.; Teng, Y.D.; Ourednik, V.; Ourednik, J.; Wakeman, D.R.; Parsons, X.H.; Gonzalez, R.; Blanchard, B.C.; Kim, S.U.; et al. Behavioral improvement in a primate Parkinson’s model is associated with multiple homeostatic effects of human neural stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 12175–12180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinden, J.D.; Vishnubhatla, I.; Muir, K.W. Prospects for stem cell-derived therapy in stroke. Prog. Brain Res. 2012, 201, 119–167. [Google Scholar] [CrossRef] [PubMed]
- Miljan, E.A.; Sinden, J.D. Stem cell treatment of ischemic brain injury. Curr. Opin. Mol. Ther. 2009, 11, 394–403. [Google Scholar] [PubMed]
- He, J.Q.; Sussman, E.S.; Steinberg, G.K. Revisiting Stem Cell-Based Clinical Trials for Ischemic Stroke. Front. Ag. Neurosci. 2020, 12, 575990. [Google Scholar] [CrossRef]
- Elder, G.A.; De Gasperi, R.; Sosa, M.A.G. Research update: Neurogenesis in adult brain and neuropsychiatric disorders. Mount. Sinai J. Med. 2006, 73, 931–940. [Google Scholar]
- Djavadian, R.L. Serotonin and neurogenesis in the hippocampal dentate gyrus of adult mammals. Acta Neurobiol. Exp. (Wars) 2004, 64, 189–200. [Google Scholar]
- Alvarez-Buylla, A.; Garcia-Verdugo, J.M. Neurogenesis in adult subventricular zone. J. Neurosci. 2002, 22, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, A.; Lois, C. Neuronal stem cells in the brain of adult vertebrates. Stem. Cells 1995, 13, 263–272. [Google Scholar] [CrossRef]
- Lledo, P.M.; Alonso, M.; Grubb, M.S. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 2006, 7, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Iacovitti, L. Classic and novel stem cell niches in brain homeostasis and repair. Brain Res. 2015, 1628, 327–342. [Google Scholar] [CrossRef] [Green Version]
- Sorrells, S.F.; Paredes, M.F.; Velmeshev, D.; Herranz-Perez, V.; Sandoval, K.; Mayer, S.; Chang, E.F.; Insausti, R.; Kriegstein, A.R.; Rubenstein, J.L.; et al. Immature excitatory neurons develop during adolescence in the human amygdala. Nat. Commun. 2019, 10, 2748. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.F. Hypothalamic neurogenesis in the adult brain. Front. Neuroendocrinol. 2013, 34, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Daniels, S.B.; Lennington, J.B.; Notti, R.Q.; Conover, J.C. The aging neurogenic subventricular zone. Aging Cell 2006, 5, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Silva-Vargas, V.; Maldonado-Soto, A.R.; Mizrak, D.; Codega, P.; Doetsch, F. Age-Dependent Niche Signals from the Choroid Plexus Regulate Adult Neural Stem Cells. Cell Stem Cell 2016, 19, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Solano Fonseca, R.; Mahesula, S.; Apple, D.M.; Raghunathan, R.; Dugan, A.; Cardona, A.; O’Connor, J.; Kokovay, E. Neurogenic Niche Microglia Undergo Positional Remodeling and Progressive Activation Contributing to Age-Associated Reductions in Neurogenesis. Stem. Cells Dev. 2016, 25, 542–555. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, Y.; Wait, E.; Mankowski, W.; Bjornsson, C.S.; Cohen, A.R.; Zuloaga, K.L.; Temple, S. 3D image analysis of the complete ventricular-subventricular zone stem cell niche reveals significant vasculature changes and progenitor deficits in males versus females with aging. Stem. Cell Rep. 2021. [Google Scholar] [CrossRef]
- Quinones-Hinojosa, A.; Sanai, N.; Soriano-Navarro, M.; Gonzalez-Perez, O.; Mirzadeh, Z.; Gil-Perotin, S.; Romero-Rodriguez, R.; Berger, M.S.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Cellular composition and cytoarchitecture of the adult human subventricular zone: A niche of neural stem cells. J. Comp. Neurol. 2006, 494, 415–434. [Google Scholar] [CrossRef]
- Bond, A.M.; Ming, G.L.; Song, H. Adult Mammalian Neural Stem Cells and Neurogenesis: Five Decades Later. Cell Stem. Cell 2015, 17, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obernier, K.; Alvarez-Buylla, A. Neural stem cells: Origin, heterogeneity and regulation in the adult mammalian brain. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernie, S.G.; Parent, J.M. Forebrain neurogenesis after focal Ischemic and traumatic brain injury. Neurobiol. Dis. 2010, 37, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Kokaia, Z.; Lindvall, O. Neurogenesis after ischaemic brain insults. Curr. Opin. Neurobiol. 2003, 13, 127–132. [Google Scholar] [CrossRef]
- Liu, Y.P.; Lang, B.T.; Baskaya, M.K.; Dempsey, R.J.; Vemuganti, R. The potential of neural stem cells to repair stroke-induced brain damage. Acta Neuropathol. 2009, 117, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Lindvall, O.; Kokaia, Z. Neurogenesis following Stroke Affecting the Adult Brain. Cold Spring Harb Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres, R.H.; Choi, R.; Steinberg, G.K.; Guzman, R. Potential of adult neural stem cells in stroke therapy. Regen. Med. 2008, 3, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Guzman, R.; De Los Angeles, A.; Cheshier, S.; Choi, R.; Hoang, S.; Liauw, J.; Schaar, B.; Steinberg, G. Intracarotid injection of fluorescence activated cell-sorted CD49d-positive neural stem cells improves targeted cell delivery and behavior after stroke in a mouse stroke model. Stroke 2008, 39, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, G.K.; Kondziolka, D.; Wechsler, L.R.; Lunsford, L.D.; Coburn, M.L.; Billigen, J.B.; Kim, A.S.; Johnson, J.N.; Bates, D.; King, B.; et al. Clinical Outcomes of Transplanted Modified Bone Marrow-Derived Mesenchymal Stem Cells in Stroke: A Phase 1/2a Study. Stroke 2016, 47, 1817–1824. [Google Scholar] [CrossRef] [Green Version]
- Burns, T.C.; Steinberg, G.K. Stem cells and stroke: Opportunities, challenges and strategies. Exp. Opin Biol. Ther. 2011, 11, 447–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.L.; Chopp, M.; Roberts, C.; Liu, X.; Wei, M.; Nejad-Davarani, S.P.; Wang, X.; Zhang, Z.G. Stroke increases neural stem cells and angiogenesis in the neurogenic niche of the adult mouse. PLoS ONE 2014, 9, e113972. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.L.; Zhang, Z.G.; Wang, L.; Wang, Y.; Gousev, A.; Zhang, L.; Ho, K.L.; Morshead, C.; Chopp, M. Activated neural stem cells contribute to stroke-induced neurogenesis and neuroblast migration toward the infarct boundary in adult rats. J. Cerebral Blood Flow Metabol. 2004, 24, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Boese, A.C.; Le, Q.E.; Pham, D.; Hamblin, M.H.; Lee, J.P. Neural stem cell therapy for subacute and chronic ischemic stroke. Stem. Cell Res. Ther. 2018, 9, 154. [Google Scholar] [CrossRef]
- Lee, J.P.; McKercher, S.; Muller, F.J.; Snyder, E.Y. Neural stem cell transplantation in mouse brain. Curr. Protoc. Neurosci. 2008. [Google Scholar] [CrossRef] [PubMed]
- Reubinoff, B.E.; Itsykson, P.; Turetsky, T.; Pera, M.F.; Reinhartz, E.; Itzik, A.; Ben-Hur, T. Neural progenitors from human embryonic stem cells. Nat. Biotechnol. 2001, 19, 1134–1140. [Google Scholar] [CrossRef]
- Daadi, M.M.; Maag, A.L.; Steinberg, G.K. Adherent self-renewable human embryonic stem cell-derived neural stem cell line: Functional engraftment in experimental stroke model. PLoS ONE 2008, 3, e1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.C.; Wernig, M.; Duncan, I.D.; Brustle, O.; Thomson, J.A. In vitro differentiation of transplantable neural precursors from human embryonic stem cells. Nat. Biotechnol. 2001, 19, 1129–1133. [Google Scholar] [CrossRef]
- Chambers, S.M.; Fasano, C.A.; Papapetrou, E.P.; Tomishima, M.; Sadelain, M.; Studer, L. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat. Biotechnol. 2009, 27, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Liu, J. Induced pluripotent stem cell-derived neural stem cells: New hope for stroke? Stem. Cell Res Ther. 2013, 4, 115. [Google Scholar] [CrossRef] [Green Version]
- Aasen, T.; Raya, A.; Barrero, M.J.; Garreta, E.; Consiglio, A.; Gonzalez, F.; Vassena, R.; Bilic, J.; Pekarik, V.; Tiscornia, G.; et al. Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes. Nat. Biotechnol. 2008, 26, 1276–1284. [Google Scholar] [CrossRef]
- Staerk, J.; Dawlaty, M.M.; Gao, Q.; Maetzel, D.; Hanna, J.; Sommer, C.A.; Mostoslavsky, G.; Jaenisch, R. Reprogramming of human peripheral blood cells to induced pluripotent stem cells. Cell Stem. Cell 2010, 7, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, I.; Kesner, N.S.; Karry, R.; Robicsek, O.; Aberdam, E.; Muller, F.J.; Aberdam, D.; Ben-Shachar, D. Induced pluripotent stem cells from hair follicles as a cellular model for neurodevelopmental disorders. Stem. Cell Res. 2012, 8, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Park, I.H.; Lerou, P.H.; Zhao, R.; Huo, H.; Daley, G.Q. Generation of human-induced pluripotent stem cells. Nat. Protocols 2008, 3, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.M.; Cooper, J.B. Lab-specific gene expression signatures in pluripotent stem cells. Cell Stem. Cell 2010, 7, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, M.H.; Mason, M.J.; Xie, W.; Volinia, S.; Singer, M.; Peterson, C.; Ambartsumyan, G.; Aimiuwu, O.; Richter, L.; Zhang, J.; et al. Induced Pluripotent Stem Cells and Embryonic Stem Cells Are Distinguished by Gene Expression Signatures. Cell Stem. Cell 2009, 5, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Efe, J.A.; Zhu, S.; Talantova, M.; Yuan, X.; Wang, S.; Lipton, S.A.; Zhang, K.; Ding, S. Direct reprogramming of mouse fibroblasts to neural progenitors. Proc. Natl. Acad. Sci. USA 2011, 108, 7838–7843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thier, M.; Worsdorfer, P.; Lakes, Y.B.; Gorris, R.; Herms, S.; Opitz, T.; Seiferling, D.; Quandel, T.; Hoffmann, P.; Nothen, M.M.; et al. Direct conversion of fibroblasts into stably expandable neural stem cells. Cell Stem. Cell 2012, 10, 473–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.W.; Tapia, N.; Hermann, A.; Hemmer, K.; Hoing, S.; Arauzo-Bravo, M.J.; Zaehres, H.; Wu, G.; Frank, S.; Moritz, S.; et al. Direct reprogramming of fibroblasts into neural stem cells by defined factors. Cell Stem. Cell 2012, 10, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lujan, E.; Chanda, S.; Ahlenius, H.; Sudhof, T.C.; Wernig, M. Direct conversion of mouse fibroblasts to self-renewing, tripotent neural precursor cells. Proc. Natl. Acad. Sci. USA 2012, 109, 2527–2532. [Google Scholar] [CrossRef] [Green Version]
- Ring, K.L.; Tong, L.M.; Balestra, M.E.; Javier, R.; Andrews-Zwilling, Y.; Li, G.; Walker, D.; Zhang, W.R.; Kreitzer, A.C.; Huang, Y. Direct reprogramming of mouse and human fibroblasts into multipotent neural stem cells with a single factor. Cell Stem. Cell 2012, 11, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Shahbazi, E.; Moradi, S.; Nemati, S.; Satarian, L.; Basiri, M.; Gourabi, H.; Zare Mehrjardi, N.; Gunther, P.; Lampert, A.; Handler, K.; et al. Conversion of Human Fibroblasts to Stably Self-Renewing Neural Stem Cells with a Single Zinc-Finger Transcription Factor. Stem. Cell Rep. 2016, 6, 539–551. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Lin, Y.H.; Sun, Y.J.; Zhu, S.; Zheng, J.; Liu, K.; Cao, N.; Li, K.; Huang, Y.; Ding, S. Pharmacological Reprogramming of Fibroblasts into Neural Stem Cells by Signaling-Directed Transcriptional Activation. Cell Stem Cell 2016, 18, 653–667. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.R.; Duan, W.M.; Reyes, M.; Keene, C.D.; Verfaillie, C.M.; Low, W.C. Human bone marrow stem cells exhibit neural phenotypes and ameliorate neurological deficits after grafting into the ischemic brain of rats. Exp. Neurol. 2002, 174, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Politi, L.S.; Bacigaluppi, M.; Brambilla, E.; Cadioli, M.; Falini, A.; Comi, G.; Scotti, G.; Martino, G.; Pluchino, S. Magnetic-resonance-based tracking and quantification of intravenously injected neural stem cell accumulation in the brains of mice with experimental multiple sclerosis. Stem. Cells 2007, 25, 2583–2592. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Methot, D.; Poppa, V.; Fujio, Y.; Walsh, K.; Murry, C.E. Cardiomyocyte grafting for cardiac repair: Graft cell death and anti-death strategies. J. Mol. Cell Cardiol. 2001, 33, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Imitola, J.; Raddassi, K.; Park, K.I.; Mueller, F.J.; Nieto, M.; Teng, Y.D.; Frenkel, D.; Li, J.; Sidman, R.L.; Walsh, C.A.; et al. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 18117–18122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Acton, P.D.; Ferrari, V.A. Imaging stem cells implanted in infarcted myocardium. J. Am. Coll Cardiol. 2006, 48, 2094–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.P.; Zhang, R.; Yan, M.; Duggineni, S.; Wakeman, D.R.; Niles, W.L.; Feng, Y.; Chen, J.; Hamblin, M.H.; Han, E.B.; et al. Chemical mutagenesis of a GPCR ligand: Detoxifying “inflammo-attraction” to direct therapeutic stem cell migration. Proc. Natl. Acad. Sci. USA 2020, 117, 31177–31188. [Google Scholar] [CrossRef]
- Boese, A.C.; Eckert, A.; Hamblin, M.H.; Lee, J.P. Human neural stem cells improve early stage stroke outcome in delayed tissue plasminogen activator-treated aged stroke brains. Exp. Neurol. 2020, 329, 113275. [Google Scholar] [CrossRef]
- Eckert, A.; Huang, L.; Gonzalez, R.; Kim, H.S.; Hamblin, M.H.; Lee, J.P. Bystander Effect Fuels Human Induced Pluripotent Stem Cell-Derived Neural Stem Cells to Quickly Attenuate Early Stage Neurological Deficits After Stroke. Stem. Cells Transl. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wong, S.; Snyder, E.Y.; Hamblin, M.H.; Lee, J.P. Human neural stem cells rapidly ameliorate symptomatic inflammation in early-stage ischemic-reperfusion cerebral injury. Stem. Cell Res. Ther. 2014, 5, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imitola, J.; Comabella, M.; Chandraker, A.K.; Dangond, F.; Sayegh, M.H.; Snyder, E.Y.; Khoury, S.J. Neural stem/progenitor cells express costimulatory molecules that are differentially regulated by inflammatory and apoptotic stimuli. Am. J. Pathol. 2004, 164, 1615–1625. [Google Scholar] [CrossRef] [Green Version]
- Belmadani, A.; Tran, P.B.; Ren, D.; Miller, R.J. Chemokines regulate the migration of neural progenitors to sites of neuroinflammation. J. Neurosci. 2006, 26, 3182–3191. [Google Scholar] [CrossRef]
- Li, M.; Hale, J.S.; Rich, J.N.; Ransohoff, R.M.; Lathia, J.D. Chemokine CXCL12 in neurodegenerative diseases: An SOS signal for stem cell-based repair. Trends Neurosci. 2012, 35, 619–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbajal, K.S.; Schaumburg, C.; Strieter, R.; Kane, J.; Lane, T.E. Migration of engrafted neural stem cells is mediated by CXCL12 signaling through CXCR4 in a viral model of multiple sclerosis. Proc. Natl. Acad Sci. USA 2010, 107, 11068–11073. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.B.; Ren, D.; Veldhouse, T.J.; Miller, R.J. Chemokine receptors are expressed widely by embryonic and adult neural progenitor cells. J. Neurosci. Res. 2004, 76, 20–34. [Google Scholar] [CrossRef]
- Tran, P.B.; Banisadr, G.; Ren, D.; Chenn, A.; Miller, R.J. Chemokine receptor expression by neural progenitor cells in neurogenic regions of mouse brain. J. Comp. Neurol. 2007, 500, 1007–1033. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.B.; Miller, R.J. Chemokine receptors: Signposts to brain development and disease. Nat. Rev. Neurosci. 2003, 4, 444–455. [Google Scholar] [CrossRef]
- Itoh, T.; Satou, T.; Ishida, H.; Nishida, S.; Tsubaki, M.; Hashimoto, S.; Ito, H. The relationship between SDF-1alpha/CXCR4 and neural stem cells appearing in damaged area after traumatic brain injury in rats. Neurol. Res. 2009, 31, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Schober, A.; Karshovska, E.; Zernecke, A.; Weber, C. SDF-1alpha-mediated tissue repair by stem cells: A promising tool in cardiovascular medicine? Trends Cardiovasc. Med. 2006, 16, 103–108. [Google Scholar] [CrossRef]
- Togel, F.; Isaac, J.; Hu, Z.; Weiss, K.; Westenfelder, C. Renal SDF-1 signals mobilization and homing of CXCR4-positive cells to the kidney after ischemic injury. Kidney Int. 2005, 67, 1772–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasyutina, E.; Stebler, J.; Brand-Saberi, B.; Schulz, S.; Raz, E.; Birchmeier, C. CXCR4 and Gab1 cooperate to control the development of migrating muscle progenitor cells. Genes Devel. 2005, 19, 2187–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, L.; Wang, J.; Wang, W.; Yang, Z.; Hu, Z.; Hu, M.; Ding, P. The effect of stromal cell-derived factor 1 in the migration of neural stem cells. Cell Biochem. Biophys. 2014, 70, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Robin, A.M.; Zhang, Z.G.; Wang, L.; Zhang, R.L.; Katakowski, M.; Zhang, L.; Wang, Y.; Zhang, C.; Chopp, M. Stromal cell-derived factor 1alpha mediates neural progenitor cell motility after focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2006, 26, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.J.; Stroemer, R.P.; Gorenkova, N.; Nakajima, M.; Crum, W.R.; Tang, E.; Stevanato, L.; Sinden, J.D.; Modo, M. Implantation Site and Lesion Topology Determine Efficacy of a Human Neural Stem Cell Line in a Rat Model of Chronic Stroke. Stem Cells 2012, 30, 785–796. [Google Scholar] [CrossRef]
- Song, M.; Kim, Y.; Kim, Y.; Ryu, S.; Song, I.; Kim, S.U.; Yoon, B.W. MRI tracking of intravenously transplanted human neural stem cells in rat focal ischemia model. Neurosci. Res. 2009, 64, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.Y.; Pendharkar, A.V.; Wang, N.; Choi, R.; Andres, R.H.; Gaeta, X.; Zhang, J.; Moseley, M.E.; Guzman, R. Intra-arterial injection of neural stem cells using a microneedle technique does not cause microembolic strokes. J. Cereb. Blood Flow Metabolism 2011, 31, 1263–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.T.; Chu, K.; Jung, K.H.; Kim, S.J.; Kim, D.H.; Kang, K.M.; Hong, N.H.; Kim, J.H.; Ban, J.J.; Park, H.K.; et al. Anti-inflammatory mechanism of intravascular neural stem cell transplantation in haemorrhagic stroke. Brain 2008, 131, 616–629. [Google Scholar] [CrossRef] [Green Version]
- Fischer, U.M.; Harting, M.T.; Jimenez, F.; Monzon-Posadas, W.O.; Xue, H.; Savitz, S.I.; Laine, G.A.; Cox, C.S., Jr. Pulmonary passage is a major obstacle for intravenous stem cell delivery: The pulmonary first-pass effect. Stem. Cells Dev. 2009, 18, 683–692. [Google Scholar] [CrossRef]
- Hauger, O.; Frost, E.E.; van Heeswijk, R.; Deminiere, C.; Xue, R.; Delmas, Y.; Combe, C.; Moonen, C.T.; Grenier, N.; Bulte, J.W. MR evaluation of the glomerular homing of magnetically labeled mesenchymal stem cells in a rat model of nephropathy. Radiology 2006, 238, 200–210. [Google Scholar] [CrossRef]
- Kraitchman, D.L.; Tatsumi, M.; Gilson, W.D.; Ishimori, T.; Kedziorek, D.; Walczak, P.; Segars, W.P.; Chen, H.H.; Fritzges, D.; Izbudak, I.; et al. Dynamic imaging of allogeneic mesenchymal stem cells trafficking to myocardial infarction. Circulation 2005, 112, 1451–1461. [Google Scholar] [CrossRef] [Green Version]
- Bliss, T.; Guzman, R.; Daadi, M.; Steinberg, G.K. Cell transplantation therapy for stroke. Stroke 2007, 38, 817–826. [Google Scholar] [CrossRef]
- Van Velthoven, C.T.; Kavelaars, A.; van Bel, F.; Heijnen, C.J. Mesenchymal stem cell treatment after neonatal hypoxic-ischemic brain injury improves behavioral outcome and induces neuronal and oligodendrocyte regeneration. Brain Behav. Immun. 2010, 24, 387–393. [Google Scholar] [CrossRef]
- Wei, N.; Yu, S.P.; Gu, X.; Taylor, T.M.; Song, D.; Liu, X.F.; Wei, L. Delayed intranasal delivery of hypoxic-preconditioned bone marrow mesenchymal stem cells enhanced cell homing and therapeutic benefits after ischemic stroke in mice. Cell Transplant 2013, 22, 977–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begley, D.J.; Brightman, M.W. Structural and functional aspects of the blood-brain barrier. Prog. Drug Res. Fort. Arzneimittelf. Prog. Recher. Pharm. 2003, 61, 39–78. [Google Scholar]
- Abbott, N.J. Astrocyte-endothelial interactions and blood-brain barrier permeability. J Anat. 2002, 200, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Saunders, N.R.; Dreifuss, J.J.; Dziegielewska, K.M.; Johansson, P.A.; Habgood, M.D.; Mollgard, K.; Bauer, H.C. The rights and wrongs of blood-brain barrier permeability studies: A walk through 100 years of history. Front. Neurosci. 2014, 8, 404. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Prat, A. The blood-brain barrier. Cold Spring Harb Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, maintenance and disruption of the blood-brain barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Schoknecht, K.; David, Y.; Heinemann, U. The blood-brain barrier-gatekeeper to neuronal homeostasis: Clinical implications in the setting of stroke. Semin. Cell Devel. Biol. 2015, 38, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Engelhardt, B. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat Rev. Immunol. 2012, 12, 623–635. [Google Scholar] [CrossRef]
- Shechter, R.; Schwartz, M. Harnessing monocyte-derived macrophages to control central nervous system pathologies: No longer ‘if’ but ‘how’. J. Pathol. 2013, 229, 332–346. [Google Scholar] [CrossRef]
- White, B.C.; Sullivan, J.M.; DeGracia, D.J.; ’O’Neil, B.J.; Neumar, R.W.; Grossman, L.I.; Rafols, J.A.; Krause, G.S. Brain ischemia and reperfusion: Molecular mechanisms of neuronal injury. J. Neurol. Sci. 2000, 179, 1–33. [Google Scholar] [CrossRef]
- Kulik, T.; Kusano, Y.; Aronhime, S.; Sandler, A.L.; Winn, H.R. Regulation of cerebral vasculature in normal and ischemic brain. Neuropharmacology 2008, 55, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Zhang, L.; Pu, H.; Mao, L.; Hu, X.; Jiang, X.; Xu, N.; Stetler, R.A.; Zhang, F.; Liu, X.; et al. Rapid endothelial cytoskeletal reorganization enables early blood-brain barrier disruption and long-term ischaemic reperfusion brain injury. Nat. Commun. 2016, 7, 10523. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Rosenberg, G.A. Blood-brain barrier breakdown in acute and chronic cerebrovascular disease. Stroke 2011, 42, 3323–3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.Y.; Betz, A.L. Reperfusion-induced injury to the blood-brain barrier after middle cerebral artery occlusion in rats. Stroke 1994, 25, 1658–1664. [Google Scholar] [CrossRef] [Green Version]
- Yemisci, M.; Gursoy-Ozdemir, Y.; Vural, A.; Can, A.; Topalkara, K.; Dalkara, T. Pericyte contraction induced by oxidative-nitrative stress impairs capillary reflow despite successful opening of an occluded cerebral artery. Nat. Med. 2009, 15, 1031–1037. [Google Scholar] [CrossRef]
- Daneman, R. The blood-brain barrier in health and disease. Ann. Neurol. 2012, 72, 648–672. [Google Scholar] [CrossRef]
- Pop, V.; Badaut, J. A neurovascular perspective for long-term changes after brain trauma. Transl. Stroke Res. 2011, 2, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Jiao, H.; Wang, Z.; Liu, Y.; Wang, P.; Xue, Y. Specific role of tight junction proteins claudin-5, occludin, and ZO-1 of the blood-brain barrier in a focal cerebral ischemic insult. J. Mol. Neurosci. 2011, 44, 130–139. [Google Scholar] [CrossRef]
- Knowland, D.; Arac, A.; Sekiguchi, K.J.; Hsu, M.; Lutz, S.E.; Perrino, J.; Steinberg, G.K.; Barres, B.A.; Nimmerjahn, A.; Agalliu, D. Stepwise recruitment of transcellular and paracellular pathways underlies blood-brain barrier breakdown in stroke. Neuron 2014, 82, 603–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Zoppo, G.J. Inflammation and the neurovascular unit in the setting of focal cerebral ischemia. Neuroscience 2009, 158, 972–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternlicht, M.D.; Lochter, A.; Sympson, C.J.; Huey, B.; Rougier, J.P.; Gray, J.W.; Pinkel, D.; Bissell, M.J.; Werb, Z. The stromal proteinase MMP3/stromelysin-1 promotes mammary carcinogenesis. Cell 1999, 98, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Werb, Z. The many faces of metalloproteases: Cell growth, invasion, angiogenesis and metastasis. Trends Cell Biol. 2001, 11, S37–S43. [Google Scholar] [CrossRef]
- Mannello, F.; Tonti, G.A.; Bagnara, G.P.; Papa, S. Role and function of matrix metalloproteinases in the differentiation and biological characterization of mesenchymal stem cells. Stem. Cells 2006, 24, 475–481. [Google Scholar] [CrossRef]
- Rosenberg, G.A.; Estrada, E.Y.; Dencoff, J.E. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke 1998, 29, 2189–2195. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Hendren, J.; Qin, X.J.; Liu, K.J. Normobaric hyperoxia reduces the neurovascular complications associated with delayed tissue plasminogen activator treatment in a rat model of focal cerebral ischemia. Stroke 2009, 40, 2526–2531. [Google Scholar] [CrossRef] [Green Version]
- Gurney, K.J.; Estrada, E.Y.; Rosenberg, G.A. Blood-brain barrier disruption by stromelysin-1 facilitates neutrophil infiltration in neuroinflammation. Neurobiol. Dis. 2006, 23, 87–96. [Google Scholar] [CrossRef]
- Mun-Bryce, S.; Rosenberg, G.A. Gelatinase B modulates selective opening of the blood-brain barrier during inflammation. Am. J. Physiol. 1998, 274, R1203–R1211. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choi, H.Y.; Ahn, H.J.; Ju, B.G.; Yune, T.Y. Matrix metalloproteinase-3 promotes early blood-spinal cord barrier disruption and hemorrhage and impairs long-term neurological recovery after spinal cord injury. Am. J. Pathol. 2014, 184, 2985–3000. [Google Scholar] [CrossRef]
- Noble, L.J.; Donovan, F.; Igarashi, T.; Goussev, S.; Werb, Z. Matrix metalloproteinases limit functional recovery after spinal cord injury by modulation of early vascular events. J. Neurosci. 2002, 22, 7526–7535. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Yang, G.J.; Li, G.H. Molecular insights and therapeutic targets for blood-brain barrier disruption in ischemic stroke: Critical role of matrix metalloproteinases and tissue-type plasminogen activator. Neurobiol. Dis. 2010, 38, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Zheng, G.; Xu, M.; Li, Y.; Chen, X.; Zhu, W.; Tong, Y.; Chung, S.K.; Liu, K.J.; Shen, J. Caveolin-1 regulates nitric oxide-mediated matrix metalloproteinases activity and blood-brain barrier permeability in focal cerebral ischemia and reperfusion injury. J. Neurochem. 2012, 120, 147–156. [Google Scholar] [CrossRef]
- Del Zoppo, G.J. The neurovascular unit, matrix proteases, and innate inflammation. Ann. N. Y. Acad. Sci. 2010, 1207, 46–49. [Google Scholar] [CrossRef]
- Heo, J.H.; Lucero, J.; Abumiya, T.; Koziol, J.A.; Copeland, B.R.; del Zoppo, G.J. Matrix metalloproteinases increase very early during experimental focal cerebral ischemia. J. Cereb. Blood Flow Metab. 1999, 19, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Romanic, A.M.; White, R.F.; Arleth, A.J.; Ohlstein, E.H.; Barone, F.C. Matrix metalloproteinase expression increases after cerebral focal ischemia in rats: Inhibition of matrix metalloproteinase-9 reduces infarct size. Stroke 1998, 29, 1020–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A.T.; Bürgers, H.F.; Rabie, T.; Marti, H.H. Matrix metalloproteinase-9 mediates hypoxia-induced vascular leakage in the brain via tight junction rearrangement. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cerebral Blood Flow Metab. 2010, 30, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.I.; Hosomi, N.; Lucero, J.; Heo, J.H.; Abumiya, T.; Mazar, A.P.; del Zoppo, G.J. Activation systems for latent matrix metalloproteinase-2 are upregulated immediately after focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2003, 23, 1408–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Rosenberg, G.A. Matrix metalloproteinases as therapeutic targets for stroke. Brain Res. 2015, 1623, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Chen, S.; Zhang, C.; Meng, F.; Wu, W.; Hu, R.; Hadass, O.; Lehmidi, T.; Blair, G.J.; Lee, M.; et al. Inhibition of MMP-9 by a selective gelatinase inhibitor protects neurovasculature from embolic focal cerebral ischemia. Mol. Neurodegen. 2012, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Sun, Y.; Xu, J.; Liu, W. Caveolin-1 mediates tissue plasminogen activator-induced MMP-9 up-regulation in cultured brain microvascular endothelial cells. J. Neurochem. 2015, 132, 724–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellanos, M.; Leira, R.; Serena, J.; Pumar, J.M.; Lizasoain, I.; Castillo, J.; Davalos, A. Plasma metalloproteinase-9 concentration predicts hemorrhagic transformation in acute ischemic stroke. Stroke 2003, 34, 40–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lee, S.R.; Arai, K.; Lee, S.R.; Tsuji, K.; Rebeck, G.W.; Lo, E.H. Lipoprotein receptor-mediated induction of matrix metalloproteinase by tissue plasminogen activator. Nat. Med. 2003, 9, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Mishiro, K.; Ishiguro, M.; Suzuki, Y.; Tsuruma, K.; Shimazawa, M.; Hara, H. A broad-spectrum matrix metalloproteinase inhibitor prevents hemorrhagic complications induced by tissue plasminogen activator in mice. Neuroscience 2012, 205, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosell, A.; Lo, E.H. Multiphasic roles for matrix metalloproteinases after stroke. Curr. Opin. Pharmacol. 2008, 8, 82–89. [Google Scholar] [CrossRef]
- Kuntz, M.; Mysiorek, C.; Petrault, O.; Petrault, M.; Uzbekov, R.; Bordet, R.; Fenart, L.; Cecchelli, R.; Berezowski, V. Stroke-induced brain parenchymal injury drives blood-brain barrier early leakage kinetics: A combined in vivo/in vitro study. J Cereb. Blood Flow Metab 2014, 34, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Asahi, M.; Wang, X.Y.; Mori, T.; Sumii, T.; Jung, J.C.; Moskowitz, M.A.; Fini, M.E.; Lo, E.H. Effects of matrix metalloproteinase-9 gene knock-out on the proteolysis of blood-brain barrier and white matter components after cerebral ischemia. J. Neurosci. 2001, 21, 7724–7732. [Google Scholar]
- Chin, J.R.; Murphy, G.; Werb, Z. Stromelysin, a connective tissue-degrading metalloendopeptidase secreted by stimulated rabbit synovial fibroblasts in parallel with collagenase. Biosynthesis, isolation, characterization, and substrates. J. Biol. Chem. 1985, 260, 12367–12376. [Google Scholar] [CrossRef]
- Hahn-Dantona, E.; Ramos-DeSimone, N.; Sipley, J.; Nagase, H.; French, D.L.; Quigley, J.P. Activation of proMMP-9 by a plasmin/MMP-3 cascade in a tumor cell model. Regulation by tissue inhibitors of metalloproteinases. Ann. N. Y. Acad. Sci. 1999, 878, 372–387. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Yang, Y.; Rosenberg, G.A. Diverse roles of matrix metalloproteinases and tissue inhibitors of metalloproteinases in neuroinflammation and cerebral ischemia. Neuroscience 2009, 158, 983–994. [Google Scholar] [CrossRef] [Green Version]
- Ramos-DeSimone, N.; Hahn-Dantona, E.; Sipley, J.; Nagase, H.; French, D.L.; Quigley, J.P. Activation of matrix metalloproteinase-9 (MMP-9) via a converging plasmin/stromelysin-1 cascade enhances tumor cell invasion. J. Biol. Chem. 1999, 274, 13066–13076. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Bischoff, R. Physiology and pathophysiology of matrix metalloproteases. Amino Acids 2011, 41, 271–290. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Nagai, N.; Umemura, K.; Collen, D.; Lijnen, H.R. Stromelysin-1 (MMP-3) is critical for intracranial bleeding after t-PA treatment of stroke in mice. J. Thromb Haemost 2007, 5, 1732–1739. [Google Scholar] [CrossRef]
- Cuadrado, E.; Rosell, A.; Penalba, A.; Slevin, M.; Alvarez-Sabin, J.; Ortega-Aznar, A.; Montaner, J. Vascular MMP-9/TIMP-2 and neuronal MMP-10 up-regulation in human brain after stroke: A combined laser microdissection and protein array study. J. Proteome Res. 2009, 8, 3191–3197. [Google Scholar] [CrossRef]
- Herz, J.; Strickland, D.K. LRP: A multifunctional scavenger and signaling receptor. J. Clin. Invest. 2001, 108, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Nagai, N.; Yamakawa, K.; Kawakami, J.; Lijnen, H.R.; Umemura, K. Tissue-type plasminogen activator (t-PA) induces stromelysin-1 (MMP-3) in endothelial cells through activation of lipoprotein receptor-related protein. Blood 2009, 114, 3352–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sole, S.; Petegnief, V.; Gorina, R.; Chamorro, A.; Planas, A.M. Activation of matrix metalloproteinase-3 and agrin cleavage in cerebral ischemia/reperfusion. J. Neuropathol. Exp. Neurol. 2004, 63, 338–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafez, S.; Abdelsaid, M.; El-Shafey, S.; Johnson, M.H.; Fagan, S.C.; Ergul, A. Matrix Metalloprotease 3 Exacerbates Hemorrhagic Transformation and Worsens Functional Outcomes in Hyperglycemic Stroke. Stroke 2016, 47, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mena, H.; Cadavid, D.; Rushing, E.J. Human cerebral infarct: A proposed histopathologic classification based on 137 cases. Acta Neuropathol. 2004, 108, 524–530. [Google Scholar] [CrossRef]
- Iadecola, C.; Anrather, J. The immunology of stroke: From mechanisms to translation. Nat. Med. 2011, 17, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Aktas, O.; Ullrich, O.; Infante-Duarte, C.; Nitsch, R.; Zipp, F. Neuronal damage in brain inflammation. Arch Neurol. 2007, 64, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Gendron, A.; Teitelbaum, J.; Cossette, C.; Nuara, S.; Dumont, M.; Geadah, D.; du Souich, P.; Kouassi, E. Temporal effects of left versus right middle cerebral artery occlusion on spleen lymphocyte subsets and mitogenic response in Wistar rats. Brain Res. 2002, 955, 85–97. [Google Scholar] [CrossRef]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.; Sansing, L.H. Microglial responses after ischemic stroke and intracerebral hemorrhage. Clin. Dev. Immunol. 2013, 2013, 746068. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Z.; Liu, C.; Zhao, Y.; Chen, Y. Activated microglia provide a neuroprotective role by balancing glial cell-line derived neurotrophic factor and tumor necrosis factor-alpha secretion after subacute cerebral ischemia. Int. J. Mol. Med. 2013, 31, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Rezai-Zadeh, K.; Gate, D.; Town, T. CNS infiltration of peripheral immune cells: D-Day for neurodegenerative disease? J. Neuroimmun. Pharm. 2009, 4, 462–475. [Google Scholar] [CrossRef] [Green Version]
- Ritzel, R.M.; Patel, A.R.; Grenier, J.M.; Crapser, J.; Verma, R.; Jellison, E.R.; McCullough, L.D. Functional differences between microglia and monocytes after ischemic stroke. J. Neuroinflamm. 2015, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Cho, S. Microglia and Monocyte-Derived Macrophages in Stroke. Neurother J. Am. Soc. Exp. NeuroTherapeutics 2016, 13, 702–718. [Google Scholar] [CrossRef]
- Xiong, X.Y.; Liu, L.; Yang, Q.W. Functions and mechanisms of microglia/macrophages in neuroinflammation and neurogenesis after stroke. Prog. Neurobiol. 2016, 142, 23–44. [Google Scholar] [CrossRef] [PubMed]
- Vivien, D.; Ali, C. Transforming growth factor-beta signalling in brain disorders. Cytokine Growth Factor Rev. 2006, 17, 121–128. [Google Scholar] [CrossRef]
- Hu, X.; Leak, R.K.; Shi, Y.; Suenaga, J.; Gao, Y.; Zheng, P.; Chen, J. Microglial and macrophage polarization-new prospects for brain repair. Nat. Rev. Neurol. 2015, 11, 56–64. [Google Scholar] [CrossRef]
- Cai, W.; Dai, X.; Chen, J.; Zhao, J.; Xu, M.; Zhang, L.; Yang, B.; Zhang, W.; Rocha, M.; Nakao, T.; et al. STAT6/Arg1 promotes microglia/macrophage efferocytosis and inflammation resolution in stroke mice. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Brunn, A.; Mihelcic, M.; Carstov, M.; Hummel, L.; Geier, F.; Schmidt, A.; Saupe, L.; Utermohlen, O.; Deckert, M. IL-10, IL-4, and STAT6 promote an M2 milieu required for termination of P0(106-125)-induced murine experimental autoimmune neuritis. Am. J. Pathol. 2014, 184, 2627–2640. [Google Scholar] [CrossRef]
- Qin, H.; Holdbrooks, A.T.; Liu, Y.; Reynolds, S.L.; Yanagisawa, L.L.; Benveniste, E.N. SOCS3 deficiency promotes M1 macrophage polarization and inflammation. J. Immunol. 2012, 189, 3439–3448. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Liu, J.; Zhao, S.; Zhang, H.; Cai, W.; Cai, M.; Ji, X.; Leak, R.K.; Gao, Y.; Chen, J.; et al. Interleukin-4 Is Essential for Microglia/Macrophage M2 Polarization and Long-Term Recovery After Cerebral Ischemia. Stroke 2016, 47, 498–504. [Google Scholar] [CrossRef]
- Stevens, S.L.; Bao, J.; Hollis, J.; Lessov, N.S.; Clark, W.M.; Stenzel-Poore, M.P. The use of flow cytometry to evaluate temporal changes in inflammatory cells following focal cerebral ischemia in mice. Brain Res. 2002, 932, 110–119. [Google Scholar] [CrossRef]
- Yilmaz, G.; Granger, D.N. Leukocyte recruitment and ischemic brain injury. Neuromol. Med. 2010, 12, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Kunis, G.; Baruch, K.; Rosenzweig, N.; Kertser, A.; Miller, O.; Berkutzki, T.; Schwartz, M. IFN-gamma-dependent activation of the brain’s choroid plexus for CNS immune surveillance and repair. Brain 2013, 136, 3427–3440. [Google Scholar] [CrossRef] [Green Version]
- Niu, F.N.; Zhang, X.; Hu, X.M.; Chen, J.; Chang, L.L.; Li, J.W.; Liu, Z.; Cao, W.; Xu, Y. Targeted mutation of Fas ligand gene attenuates brain inflammation in experimental stroke. Brain Behav. Immun. 2012, 26, 61–71. [Google Scholar] [CrossRef]
- Schilling, M.; Besselmann, M.; Muller, M.; Strecker, J.K.; Ringelstein, E.B.; Kiefer, R. Predominant phagocytic activity of resident microglia over hematogenous macrophages following transient focal cerebral ischemia: An investigation using green fluorescent protein transgenic bone marrow chimeric mice. Exp. Neurol. 2005, 196, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Besancon, E.; Guo, S.; Lok, J.; Tymianski, M.; Lo, E.H. Beyond NMDA and AMPA glutamate receptors: Emerging mechanisms for ionic imbalance and cell death in stroke. Trends Pharmacol. Sci. 2008, 29, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Lipton, P. Ischemic cell death in brain neurons. Physiol. Rev. 1999, 79, 1431–1568. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yenari, M.A.; Cheng, D.; Sapolsky, R.M.; Steinberg, G.K. Bcl-2 overexpression protects against neuron loss within the ischemic margin following experimental stroke and inhibits cytochrome c translocation and caspase-3 activity. J. Neurochem. 2003, 85, 1026–1036. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Fahy, B.; Fiskum, G. Cytochrome c release from brain mitochondria is independent of the mitochondrial permeability transition. FEBS Lett. 1998, 439, 373–376. [Google Scholar] [CrossRef] [Green Version]
- Petrovic-Djergovic, D.; Goonewardena, S.N.; Pinsky, D.J. Inflammatory Disequilibrium in Stroke. Circ. Res. 2016, 119, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Domercq, M.; Etxebarria, E.; Perez-Samartin, A.; Matute, C. Excitotoxic oligodendrocyte death and axonal damage induced by glutamate transporter inhibition. Glia 2005, 52, 36–46. [Google Scholar] [CrossRef]
- Schiene, K.; Bruehl, C.; Zilles, K.; Qu, M.; Hagemann, G.; Kraemer, M.; Witte, O.W. Neuronal hyperexcitability and reduction of GABAA-receptor expression in the surround of cerebral photothrombosis. J. Cereb Blood Flow Metab. 1996, 16, 906–914. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.L.; Wang, X.M.; Xie, L.; Mao, X.O.; Zhu, W.; Wang, Y.; Shen, J.F.; Mao, Y.; Banwait, S.; Greenberg, D.A. Evidence for stroke-induced neurogenesis in the human brain. Proc. Nat. Acad. Sci. USA 2006, 103, 13198–13202. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.S.; Washington, P.M.; Kernie, S.G. Injury-Induced Neurogenesis: Mechanisms and Relevance. Neuroscientist 2016, 22, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Nakagomi, T.; Molnar, Z.; Nakano-Doi, A.; Taguchi, A.; Saino, O.; Kubo, S.; Clausen, M.; Yoshikawa, H.; Nakagomi, N.; Matsuyama, T. Ischemia-induced neural stem/progenitor cells in the pia mater following cortical infarction. Stem. Cells Dev. 2011, 20, 2037–2051. [Google Scholar] [CrossRef]
- Krupinski, J.; Kaluza, J.; Kumar, P.; Kumar, S.; Wang, J.M. Role of angiogenesis in patients with cerebral ischemic stroke. Stroke 1994, 25, 1794–1798. [Google Scholar] [CrossRef] [Green Version]
- Plate, K.H.; Beck, H.; Danner, S.; Allegrini, P.R.; Wiessner, C. Cell type specific upregulation of vascular endothelial growth factor in an MCA-occlusion model of cerebral infarct. J. Neuropathol. Exp. Neurol. 1999, 58, 654–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Erinjeri, J.P.; Rovainen, C.M.; Woolsey, T.A. Collateral growth and angiogenesis around cortical stroke. Stroke 2001, 32, 2179–2184. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.W.; Friedman, B.; Cheng, Q.; Lyden, P.D. Stroke-evoked angiogenesis results in a transient population of microvessels. J. Cereb. Blood Flow Metab. 2007, 27, 755–763. [Google Scholar] [CrossRef]
- Adamczak, J.M.; Schneider, G.; Nelles, M.; Que, I.; Suidgeest, E.; van der Weerd, L.; Lowik, C.; Hoehn, M. In vivo bioluminescence imaging of vascular remodeling after stroke. Front. Cell Neurosci. 2014, 8, 274. [Google Scholar] [CrossRef] [Green Version]
- Marti, H.J.; Bernaudin, M.; Bellail, A.; Schoch, H.; Euler, M.; Petit, E.; Risau, W. Hypoxia-induced vascular endothelial growth factor expression precedes neovascularization after cerebral ischemia. Am. J. Pathol. 2000, 156, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Noshita, N.; Sugawara, T.; Chan, P.H. Temporal profile of angiogenesis and expression of related genes in the brain after ischemia. J. Cereb. Blood Flow Metab. 2003, 23, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.S.; Zhang, Z.G.; Zhang, R.L.; Gregg, S.; Morris, D.C.; Wang, Y.; Chopp, M. Stroke induces gene profile changes associated with neurogenesis and angiogenesis in adult subventricular zone progenitor cells. J. Cereb. Blood Flow Metab. 2007, 27, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Hermann, D.M.; Zechariah, A. Implications of vascular endothelial growth factor for postischemic neurovascular remodeling. J. Cereb. Blood Flow Metab. 2009, 29, 1620–1643. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zechariah, A.; Qu, Y.; Hermann, D.M. Effects of vascular endothelial growth factor in ischemic stroke. J. Neurosci. Res. 2012, 90, 1873–1882. [Google Scholar] [CrossRef]
- Qiu, J.; Xu, J.; Zheng, Y.; Wei, Y.; Zhu, X.; Lo, E.H.; Moskowitz, M.A.; Sims, J.R. High-mobility group box 1 promotes metalloproteinase-9 upregulation through Toll-like receptor 4 after cerebral ischemia. Stroke 2010, 41, 2077–2082. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, Y.; Katakowski, M.; Chen, X.; Wang, L.; Lu, D.; Lu, M.; Gautam, S.C.; Chopp, M. Intravenous bone marrow stromal cell therapy reduces apoptosis and promotes endogenous cell proliferation after stroke in female rat. J. Neurosci. Res. 2003, 73, 778–786. [Google Scholar] [CrossRef]
- Krupinski, J.; Stroemer, P.; Slevin, M.; Marti, E.; Kumar, P.; Rubio, F. Three-dimensional structure and survival of newly formed blood vessels after focal cerebral ischemia. Neuroreport 2003, 14, 1171–1176. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, K.; Xie, L.; Childs, J.; Mao, X.O.; Logvinova, A.; Greenberg, D.A. VEGF-induced neuroprotection, neurogenesis, and angiogenesis after focal cerebral ischemia. J. Clin. Invest. 2003, 111, 1843–1851. [Google Scholar] [CrossRef]
- Arenillas, J.F.; Sobrino, T.; Castillo, J.; Davalos, A. The role of angiogenesis in damage and recovery from ischemic stroke. Curr. Treat Options Cardiovasc. Med. 2007, 9, 205–212. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Cui, H.R.; Yang, S.Z.; Sun, H.P.; Qiu, M.H.; Feng, X.Y.; Sun, F.Y. VEGF enhance cortical newborn neurons and their neurite development in adult rat brain after cerebral ischemia. Neurochem. Int. 2009, 55, 629–636. [Google Scholar] [CrossRef]
- Petcu, E.B.; Smith, R.A.; Miroiu, R.I.; Opris, M.M. Angiogenesis in old-aged subjects after ischemic stroke: A cautionary note for investigators. J. Angiogen. Res. 2010, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Goderie, S.K.; Jin, L.; Karanth, N.; Sun, Y.; Abramova, N.; Vincent, P.; Pumiglia, K.; Temple, S. Endothelial cells stimulate self-renewal and expand neurogenesis of neural stem cells. Science 2004, 304, 1338–1340. [Google Scholar] [CrossRef] [Green Version]
- Teng, H.; Zhang, Z.G.; Wang, L.; Zhang, R.L.; Zhang, L.; Morris, D.; Gregg, S.R.; Wu, Z.; Jiang, A.; Lu, M.; et al. Coupling of angiogenesis and neurogenesis in cultured endothelial cells and neural progenitor cells after stroke. J. Cereb. Blood Flow Metab. 2008, 28, 764–771. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chang, C.; Cheung, W.M.; Lin, M.H.; Chen, J.J.; Hsu, C.Y.; Chen, J.H.; Lin, T.N. Dynamic changes in vascular permeability, cerebral blood volume, vascular density, and size after transient focal cerebral ischemia in rats: Evaluation with contrast-enhanced magnetic resonance imaging. J. Cereb. Blood Flow Metab 2008, 28, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Thored, P.; Wood, J.; Arvidsson, A.; Cammenga, J.; Kokaia, Z.; Lindvall, O. Long-term neuroblast migration along blood vessels in an area with transient angiogenesis and increased vascularization after stroke. Stroke 2007, 38, 3032–3039. [Google Scholar] [CrossRef] [Green Version]
- Lacar, B.; Herman, P.; Platel, J.C.; Kubera, C.; Hyder, F.; Bordey, A. Neural progenitor cells regulate capillary blood flow in the postnatal subventricular zone. J. Neurosci. 2012, 32, 16435–16448. [Google Scholar] [CrossRef]
- Thored, P.; Arvidsson, A.; Cacci, E.; Ahlenius, H.; Kallur, T.; Darsalia, V.; Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Persistent production of neurons from adult brain stem cells during recovery after stroke. Stem. Cells 2006, 24, 739–747. [Google Scholar] [CrossRef]
- Arvidsson, A.; Collin, T.; Kirik, D.; Kokaia, Z.; Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 2002, 8, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Goss, J.R.; O’Malley, M.E.; Zou, L.; Styren, S.D.; Kochanek, P.M.; DeKosky, S.T. Astrocytes are the major source of nerve growth factor upregulation following traumatic brain injury in the rat. Exp. Neurol. 1998, 149, 301–309. [Google Scholar] [CrossRef]
- Weidemann, A.; Krohne, T.U.; Aguilar, E.; Kurihara, T.; Takeda, N.; Dorrell, M.I.; Simon, M.C.; Haase, V.H.; Friedlander, M.; Johnson, R.S. Astrocyte hypoxic response is essential for pathological but not developmental angiogenesis of the retina. Glia 2010, 58, 1177–1185. [Google Scholar] [CrossRef] [Green Version]
- Venkat, P.; Chopp, M.; Chen, J. New insights into coupling and uncoupling of cerebral blood flow and metabolism in the brain. Croat. Med. J. 2016, 57, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Wu, Z.B.; Zhuge, Q.; Zheng, W.; Shao, B.; Wang, B.; Sun, F.; Jin, K. Glial scar formation occurs in the human brain after ischemic stroke. Int. J. Med. Sci. 2014, 11, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Roitbak, T.; Sykova, E. Diffusion barriers evoked in the rat cortex by reactive astrogliosis. Glia 1999, 28, 40–48. [Google Scholar] [CrossRef]
- Becerra-Calixto, A.; Cardona-Gomez, G.P. The Role of Astrocytes in Neuroprotection after Brain Stroke: Potential in Cell Therapy. Front. Mol. Neurosci. 2017, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Abeysinghe, H.C.; Phillips, E.L.; Chin-Cheng, H.; Beart, P.M.; Roulston, C.L. Modulating Astrocyte Transition after Stroke to Promote Brain Rescue and Functional Recovery: Emerging Targets Include Rho Kinase. Int. J. Mol. Sci. 2016, 17, 288. [Google Scholar] [CrossRef] [Green Version]
- De Feo, D.; Merlini, A.; Laterza, C.; Martino, G. Neural stem cell transplantation in central nervous system disorders: From cell replacement to neuroprotection. Curr. Opin Neurol. 2012, 25, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Andres, R.H.; Horie, N.; Slikker, W.; Keren-Gill, H.; Zhan, K.; Sun, G.; Manley, N.C.; Pereira, M.P.; Sheikh, L.A.; McMillan, E.L.; et al. Human neural stem cells enhance structural plasticity and axonal transport in the ischaemic brain. Brain 2011, 134, 1777–1789. [Google Scholar] [CrossRef]
- Emanueli, C.; Schratzberger, P.; Kirchmair, R.; Madeddu, P. Paracrine control of vascularization and neurogenesis by neurotrophins. Br. J. Pharmacol. 2003, 140, 614–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Lim, I.J.; Lee, M.C.; Kim, S.U. Human Neural Stem Cells Genetically Modified To Overexpress Brain-Derived Neurotrophic Factor Promote Functional Recovery and Neuroprotection in a Mouse Stroke Model. J. Neurosci. Res. 2010, 88, 3282–3294. [Google Scholar] [CrossRef] [PubMed]
- Schabitz, W.R.; Steigleder, T.; Cooper-Kuhn, C.M.; Schwab, S.; Sommer, C.; Schneider, A.; Kuhn, H.G. Intravenous brain-derived neurotrophic factor enhances poststroke sensorimotor recovery and stimulates neurogenesis. Stroke 2007, 38, 2165–2172. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.Y.; Park, S.I.; Oh, J.H.; Kim, S.M.; Jeong, C.H.; Jun, J.A.; Lee, K.S.; Oh, W.; Lee, J.K.; Jeun, S.S. Brain-derived neurotrophic factor stimulates the neural differentiation of human umbilical cord blood-derived mesenchymal stem cells and survival of differentiated cells through MAPK/ERK and PI3K/Akt-dependent signaling pathways. J. Neurosci. Res. 2008, 86, 2168–2178. [Google Scholar] [CrossRef]
- Abe, K. Therapeutic potential of neurotrophic factors and neural stem cells against ischemic brain injury. J. Cereb. Blood Flow Metab. 2000, 20, 1393–1408. [Google Scholar] [CrossRef]
- Sondell, M.; Sundler, F.; Kanje, M. Vascular endothelial growth factor is a neurotrophic factor which stimulates axonal outgrowth through the flk-1 receptor. Eur. J. Neurosci. 2000, 12, 4243–4254. [Google Scholar] [CrossRef]
- Leung, D.W.; Cachianes, G.; Kuang, W.J.; Goeddel, D.V.; Ferrara, N. Vascular endothelial growth factor is a secreted angiogenic mitogen. Science 1989, 246, 1306–1309. [Google Scholar] [CrossRef]
- Gerber, H.P.; McMurtrey, A.; Kowalski, J.; Yan, M.; Keyt, B.A.; Dixit, V.; Ferrara, N. Vascular endothelial growth factor regulates endothelial cell survival through the phosphatidylinositol 3’-kinase/Akt signal transduction pathway. Requirement for Flk-1/KDR activation. J. Biol. Chem. 1998, 273, 30336–30343. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Abe, K.; Itoyama, Y. Reduction of ischemic damage by application of vascular endothelial growth factor in rat brain after transient ischemia. J. Cereb. Blood Flow Metab. 1998, 18, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Kaya, D.; Gursoy-Ozdemir, Y.; Yemisci, M.; Tuncer, N.; Aktan, S.; Dalkara, T. VEGF protects brain against focal ischemia without increasing blood--brain permeability when administered intracerebroventricularly. J. Cereb. Blood. Flow Metab. 2005, 25, 1111–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, K.M.; Li, L.; Cunningham, L.A. Murine Neural Stem/Progenitor Cells Protect Neurons against Ischemia by HIF-1 alpha-Regulated VEGF Signaling. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Ann. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Multiple sclerosis in 54 twinships: Concordance rate is independent of zygosity. French Research Group on Multiple Sclerosis. Ann. Neurol. 1992, 32, 724–727. [Google Scholar] [CrossRef]
- Kang, S.S.; Keasey, M.P.; Arnold, S.A.; Reid, R.; Geralds, J.; Hagg, T. Endogenous CNTF mediates stroke-induced adult CNS neurogenesis in mice. Neurobiol. Dis. 2013, 49, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V.; Howe, C.L.; Mobley, W.C. Nerve growth factor signaling, neuroprotection, and neural repair. Ann. Rev. Neurosci. 2001, 24, 1217–1281. [Google Scholar] [CrossRef]
- Cattaneo, E.; McKay, R. Proliferation and differentiation of neuronal stem cells regulated by nerve growth factor. Nature 1990, 347, 762–765. [Google Scholar] [CrossRef]
- Yuan, M.; Wen, S.J.; Yang, C.X.; Pang, Y.G.; Gao, X.Q.; Liu, X.Q.; Huang, L.; Yuan, Q.L. Transplantation of neural stem cells overexpressing glial cell line-derived neurotrophic factor enhances Akt and Erk1/2 signaling and neurogenesis in rats after stroke. Chinese Med. J. 2013, 126, 1302–1309. [Google Scholar]
- Oki, K.; Tatarishvili, J.; Wood, J.; Koch, P.; Wattananit, S.; Mine, Y.; Monni, E.; Tornero, D.; Ahlenius, H.; Ladewig, J.; et al. Human-induced pluripotent stem cells form functional neurons and improve recovery after grafting in stroke-damaged brain. Stem Cells 2012, 30, 1120–1133. [Google Scholar] [CrossRef]
- Lampl, Y.; Boaz, M.; Gilad, R.; Lorberboym, M.; Dabby, R.; Rapoport, A.; Anca-Hershkowitz, M.; Sadeh, M. Minocycline treatment in acute stroke: An open-label, evaluator-blinded study. Neurology 2007, 69, 1404–1410. [Google Scholar] [CrossRef]
- Yang, F.; Zhou, L.; Wang, D.; Wang, Z.; Huang, Q.Y. Minocycline ameliorates hypoxia-induced blood-brain barrier damage by inhibition of HIF-1alpha through SIRT-3/PHD-2 degradation pathway. Neuroscience 2015, 304, 250–259. [Google Scholar] [CrossRef]
- Garrido-Mesa, N.; Camuesco, D.; Arribas, B.; Comalada, M.; Bailon, E.; Cueto-Sola, M.; Utrilla, P.; Nieto, A.; Zarzuelo, A.; Rodriguez-Cabezas, M.E.; et al. The intestinal anti-inflammatory effect of minocycline in experimental colitis involves both its immunomodulatory and antimicrobial properties. Pharmacol. Res. 2011, 63, 308–319. [Google Scholar] [CrossRef]
- Metz, L.M.; Li, D.K.B.; Traboulsee, A.L.; Duquette, P.; Eliasziw, M.; Cerchiaro, G.; Greenfield, J.; Riddehough, A.; Yeung, M.; Kremenchutzky, M.; et al. Trial of Minocycline in a Clinically Isolated Syndrome of Multiple Sclerosis. N. Engl. J. Med. 2017, 376, 2122–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshima, S.; Fujimoto, S.; Petrov, A.; Nakagami, H.; Haider, N.; Zhou, J.; Tahara, N.; Osako, M.K.; Fujimoto, A.; Zhu, J.; et al. Effect of an antimicrobial agent on atherosclerotic plaques: Assessment of metalloproteinase activity by molecular imaging. J. Am. Coll Cardiol. 2010, 55, 1240–1249. [Google Scholar] [CrossRef] [Green Version]
- Machado, L.S.; Kozak, A.; Ergul, A.; Hess, D.C.; Borlongan, C.V.; Fagan, S.C. Delayed minocycline inhibits ischemia-activated matrix metalloproteinases 2 and 9 after experimental stroke. BMC Neurosci. 2006, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Salayandia, V.M.; Thompson, J.F.; Yang, L.Y.; Estrada, E.Y.; Yang, Y. Attenuation of acute stroke injury in rat brain by minocycline promotes blood-brain barrier remodeling and alternative microglia/macrophage activation during recovery. J. Neuroinflam. 2015, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Yrjanheikki, J.; Tikka, T.; Keinanen, R.; Goldsteins, G.; Chan, P.H.; Koistinaho, J. A tetracycline derivative, minocycline, reduces inflammation and protects against focal cerebral ischemia with a wide therapeutic window. Proc. Natl. Acad. Sci. USA 1999, 96, 13496–13500. [Google Scholar] [CrossRef] [Green Version]
- Gasche, Y.; Fujimura, M.; Morita-Fujimura, Y.; Copin, J.C.; Kawase, M.; Massengale, J.; Chan, P.H. Early appearance of activated matrix metalloproteinase-9 after focal cerebral ischemia in mice: A possible role in blood-brain barrier dysfunction. J. Cereb. Blood Flow Meta. 1999, 19, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.K.; Gavins, F.N. The potential of stem cell therapy for stroke: Is PISCES the sign? FASEB J. 2012, 26, 2239–2252. [Google Scholar] [CrossRef] [Green Version]
- Bacigaluppi, M.; Pluchino, S.; Peruzzotti-Jametti, L.; Kilic, E.; Kilic, U.; Salani, G.; Brambilla, E.; West, M.J.; Comi, G.; Martino, G.; et al. Delayed post-ischaemic neuroprotection following systemic neural stem cell transplantation involves multiple mechanisms. Brain 2009, 132, 2239–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, D.J.; Lee, N.; Park, I.H.; Choi, C.; Jeon, I.; Kwon, J.; Oh, S.H.; Shin, D.A.; Do, J.T.; Lee, D.R.; et al. Therapeutic potential of human induced pluripotent stem cells in experimental stroke. Cell Transplant 2013, 22, 1427–1440. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Lee, S.H.; Kim, S.U.; Yoon, B.W. Human neural stem cells promote proliferation of endogenous neural stem cells and enhance angiogenesis in ischemic rat brain. Neural Regen. Res. 2016, 11, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Sakata, H.; Niizuma, K.; Wakai, T.; Narasimhan, P.; Maier, C.M.; Chan, P.H. Neural Stem Cells Genetically Modified to Overexpress Cu/Zn-Superoxide Dismutase Enhance Amelioration of Ischemic Stroke in Mice. Stroke 2012, 43, 2423. [Google Scholar] [CrossRef]
- Mine, Y.; Tatarishvili, J.; Oki, K.; Monni, E.; Kokaia, Z.; Lindvall, O. Grafted human neural stem cells enhance several steps of endogenous neurogenesis and improve behavioral recovery after middle cerebral artery occlusion in rats. Neurobiol. Dis. 2013, 52, 191–203. [Google Scholar] [CrossRef]
- Chu, K.; Kim, M.; Park, K.I.; Jeong, S.W.; Park, H.K.; Jung, K.H.; Lee, S.T.; Kang, L.; Lee, K.; Park, D.K.; et al. Human neural stem cells improve sensorimotor deficits in the adult rat brain with experimental focal ischemia. Brain Res. 2004, 1016, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.; Kim, M.; Jeong, S.W.; Kim, S.U.; Yoon, B.W. Human neural stem cells can migrate, differentiate, and integrate after intravenous transplantation in adult rats with transient forebrain ischemia. Neurosci. Lett. 2003, 343, 129–133. [Google Scholar] [CrossRef]
- Wu, W.F.; Chen, X.; Hu, C.L.; Li, J.F.; Yu, Z.; Cai, W.Q. Transplantation of neural stem cells expressing hypoxia-inducible factor-1 alpha (HIF-1 alpha) improves behavioral recovery in a rat stroke model. J. Clin. Neurosci. 2010, 17, 92–95. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhu, J.J.; Hu, Y.B.; Xiang, G.H.; Deng, L.C.; Wu, F.Z.; Wei, X.J.; Wang, Y.H.; Sun, L.Y.; Lou, X.Q.; et al. Transplantation of bFGF-expressing neural stem cells promotes cell migration and functional recovery in rat brain after transient ischemic stroke. Oncotarget 2017, 8, 102067–102077. [Google Scholar] [CrossRef] [PubMed]
- Bouet, V.; Boulouard, M.; Toutain, J.; Divoux, D.; Bernaudin, M.; Schumann-Bard, P.; Freret, T. The adhesive removal test: A sensitive method to assess sensorimotor deficits in mice. Nature Protocols 2009, 4, 1560–1564. [Google Scholar] [CrossRef] [PubMed]
- Patkar, S.; Tate, R.; Modo, M.; Plevin, R.; Carswell, H.V. Conditionally immortalised neural stem cells promote functional recovery and brain plasticity after transient focal cerebral ischaemia in mice. Stem Cell Res 2012, 8, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korshunova, I.; Rhein, S.; Garcia-Gonzalez, D.; Stolting, I.; Pfisterer, U.; Barta, A.; Dmytriyeva, O.; Kirkeby, A.; Schwaninger, M.; Khodosevich, K. Genetic modification increases the survival and the neuroregenerative properties of transplanted neural stem cells. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Ergul, A.; Kelly-Cobbs, A.; Abdalla, M.; Fagan, S.C. Cerebrovascular complications of diabetes: Focus on stroke. Endocrine Metab.Immune Disord. Drug Targets 2012, 12, 148–158. [Google Scholar] [CrossRef]
- Ergul, A.; Li, W.; Elgebaly, M.M.; Bruno, A.; Fagan, S.C. Hyperglycemia, diabetes and stroke: Focus on the cerebrovasculature. Vasc. Pharmacol. 2009, 51, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Tureyen, K.; Bowen, K.; Liang, J.; Dempsey, R.J.; Vemuganti, R. Exacerbated brain damage, edema and inflammation in type-2 diabetic mice subjected to focal ischemia. J. Neurochem. 2011, 116, 499–507. [Google Scholar] [CrossRef]
- Kalladka, D.; Sinden, J.; Pollock, K.; Haig, C.; McLean, J.; Smith, W.; McConnachie, A.; Santosh, C.; Bath, P.M.; Dunn, L.; et al. Human neural stem cells in patients with chronic ischaemic stroke (PISCES): A phase 1, first-in-man study. Lancet 2016, 388, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Muir, K.W. Clinical trial design for stem cell therapies in stroke: What have we learned? Neurochem. Int. 2016. [Google Scholar] [CrossRef] [Green Version]
- Muir, K.W.; Bulters, D.; Willmot, M.; Sprigg, N.; Dixit, A.; Ward, N.; Tyrrell, P.; Majid, A.; Dunn, L.; Bath, P.; et al. Intracerebral implantation of human neural stem cells and motor recovery after stroke: Multicentre prospective single-arm study (PISCES-2). J. Neurol Neurosurg. Psychiatry 2020, 91, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic diagram of NSC derivation through diverse methods and sources. NSCs can be directly extracted from neural tissue and expanded in vitro. Also, NSCs can be reprogrammed from iPSCs or ESCs by specific combinations of differentiation factors. Further, NSCs can be generated by direct conversion of somatic cells omitting the iPSC derivation step. ICM, inner cell mass.
Figure 1.
Schematic diagram of NSC derivation through diverse methods and sources. NSCs can be directly extracted from neural tissue and expanded in vitro. Also, NSCs can be reprogrammed from iPSCs or ESCs by specific combinations of differentiation factors. Further, NSCs can be generated by direct conversion of somatic cells omitting the iPSC derivation step. ICM, inner cell mass.

Figure 2.
Schematic diagram of NSC delivery illustrating multiple therapeutic mechanisms in ischemic stroke. NSCs can differentiate into functional neurons in the stroke brain and possess pleotropic bystander effects. These bystander effects include attenuation of blood-brain barrier (BBB) disruption, increased angiogenesis, and modulation of immune responses after ischemic stroke. ECs, endothelial cells.
Figure 2.
Schematic diagram of NSC delivery illustrating multiple therapeutic mechanisms in ischemic stroke. NSCs can differentiate into functional neurons in the stroke brain and possess pleotropic bystander effects. These bystander effects include attenuation of blood-brain barrier (BBB) disruption, increased angiogenesis, and modulation of immune responses after ischemic stroke. ECs, endothelial cells.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hamblin, M.H.; Lee, J.-P. Neural Stem Cells for Early Ischemic Stroke. Int. J. Mol. Sci. 2021, 22, 7703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147703
AMA Style
Hamblin MH, Lee J-P. Neural Stem Cells for Early Ischemic Stroke. International Journal of Molecular Sciences. 2021; 22(14):7703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147703
Chicago/Turabian StyleHamblin, Milton H., and Jean-Pyo Lee. 2021. "Neural Stem Cells for Early Ischemic Stroke" International Journal of Molecular Sciences 22, no. 14: 7703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22147703
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.