
Complex Analysis of Antioxidant Activity, Abscisic Acid Level, and Accumulation of Osmotica in Apple and Cherry In Vitro Cultures under Osmotic Stress

 , ,
, ,
Abstract
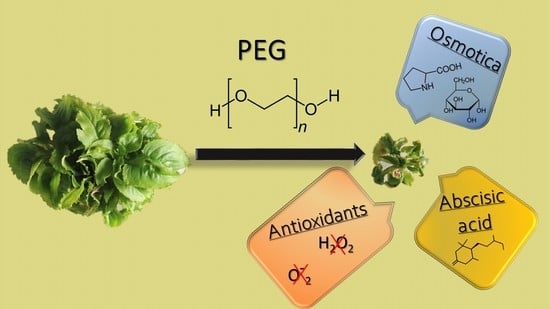
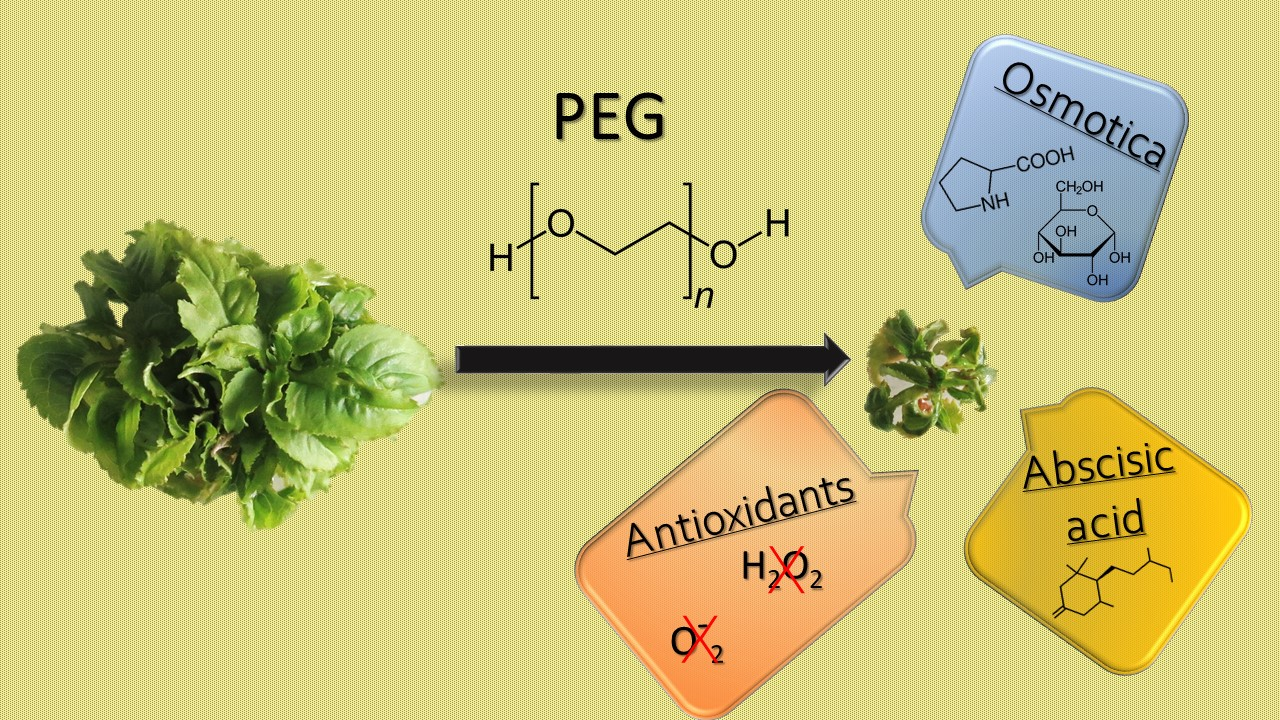
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Determination of Relative Water Content
2.3. Determination of Osmotic Regulating Compounds
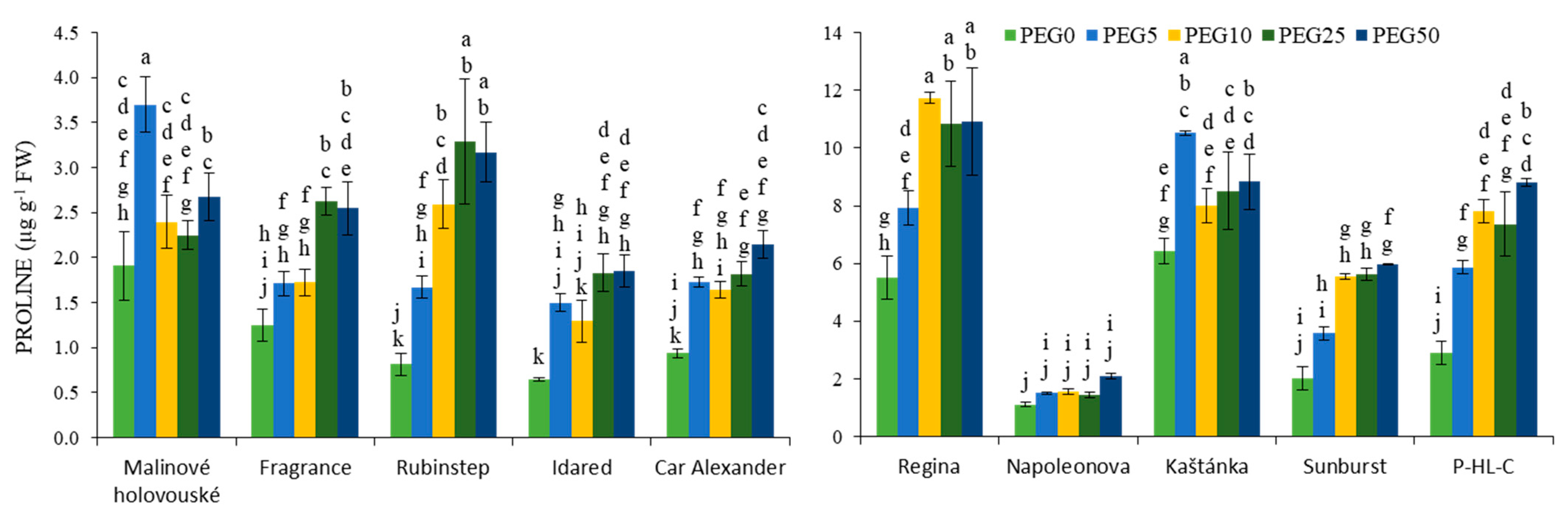
2.3.1. Proline Content
2.3.2. Simple Sugars and Sugar Alcohols Content
2.4. Determination of Antioxidant Capacity and Antioxidant Enzymes Activity
2.5. Determination of Abscisic Acid Content
2.6. Statistical Analysis
3. Results
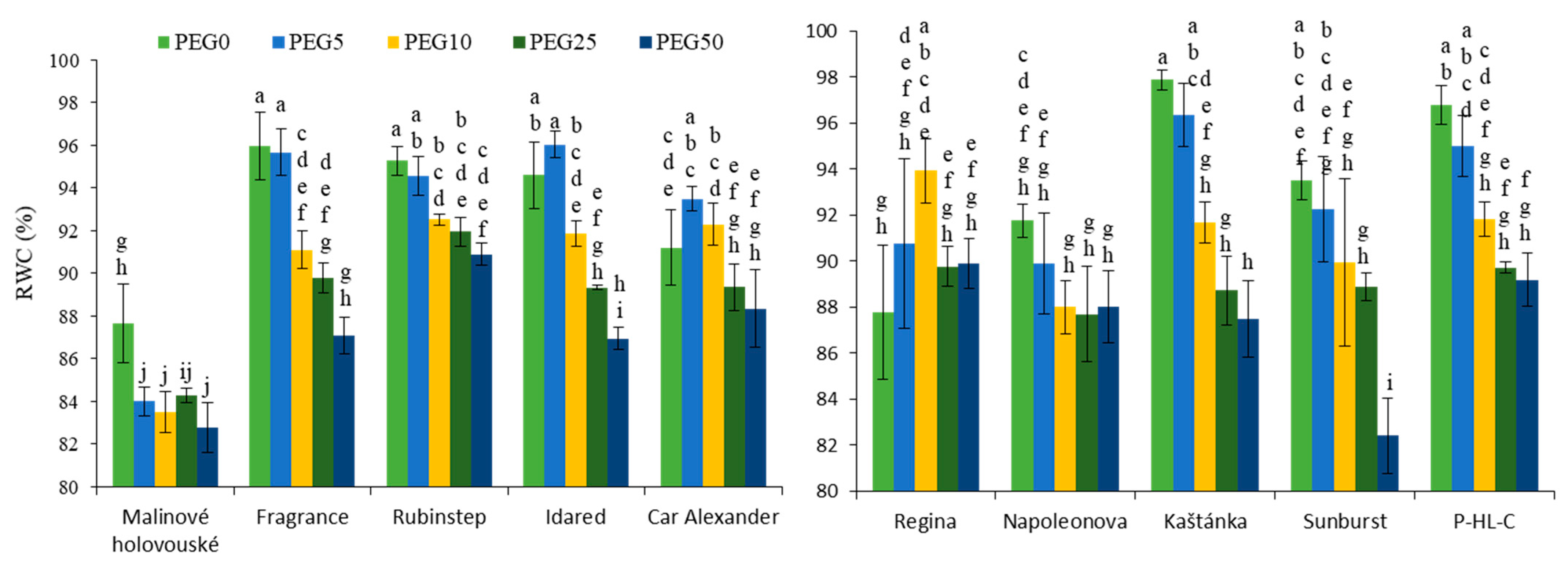
3.1. Relative Water Content (RWC)
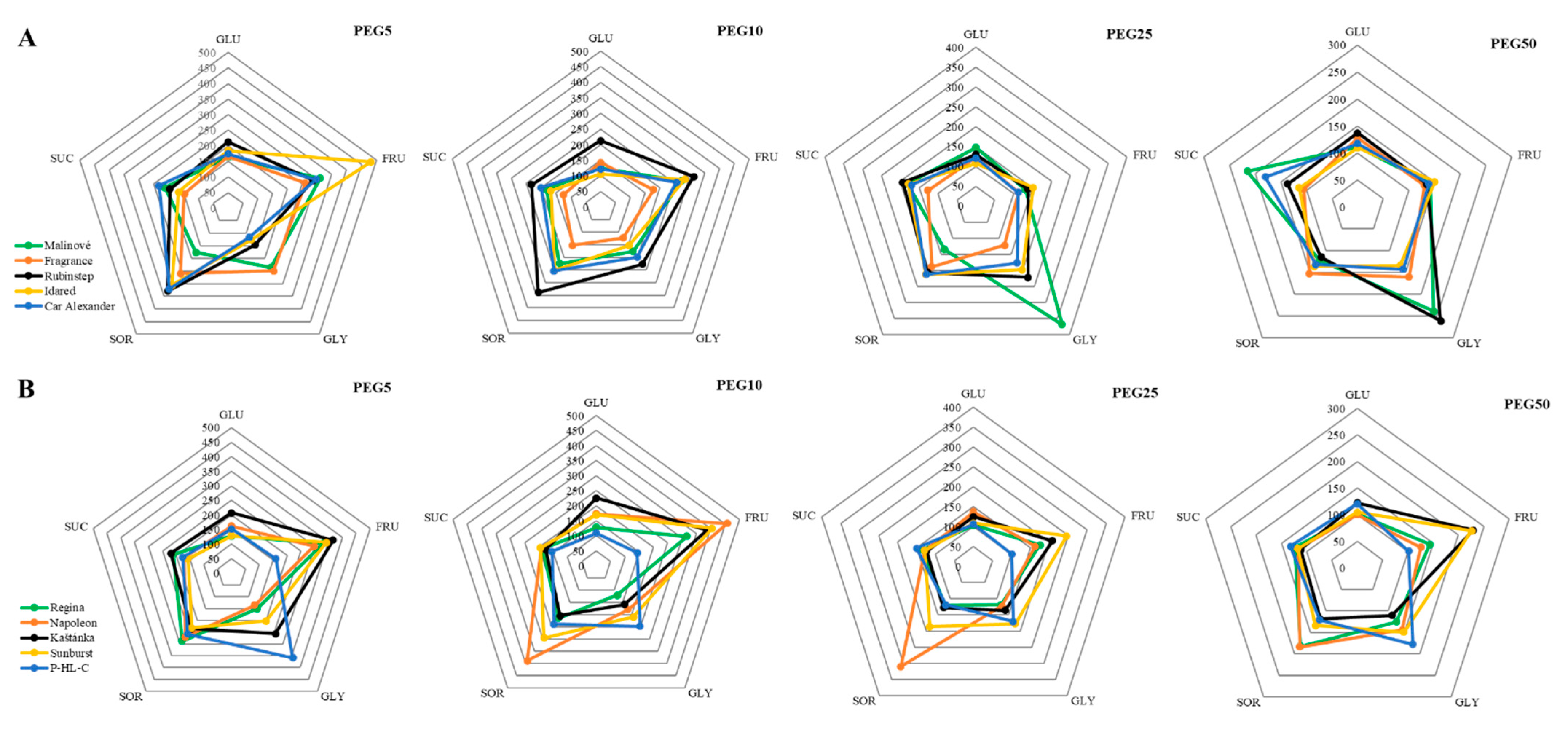
3.2. Osmotic Regulating Compounds
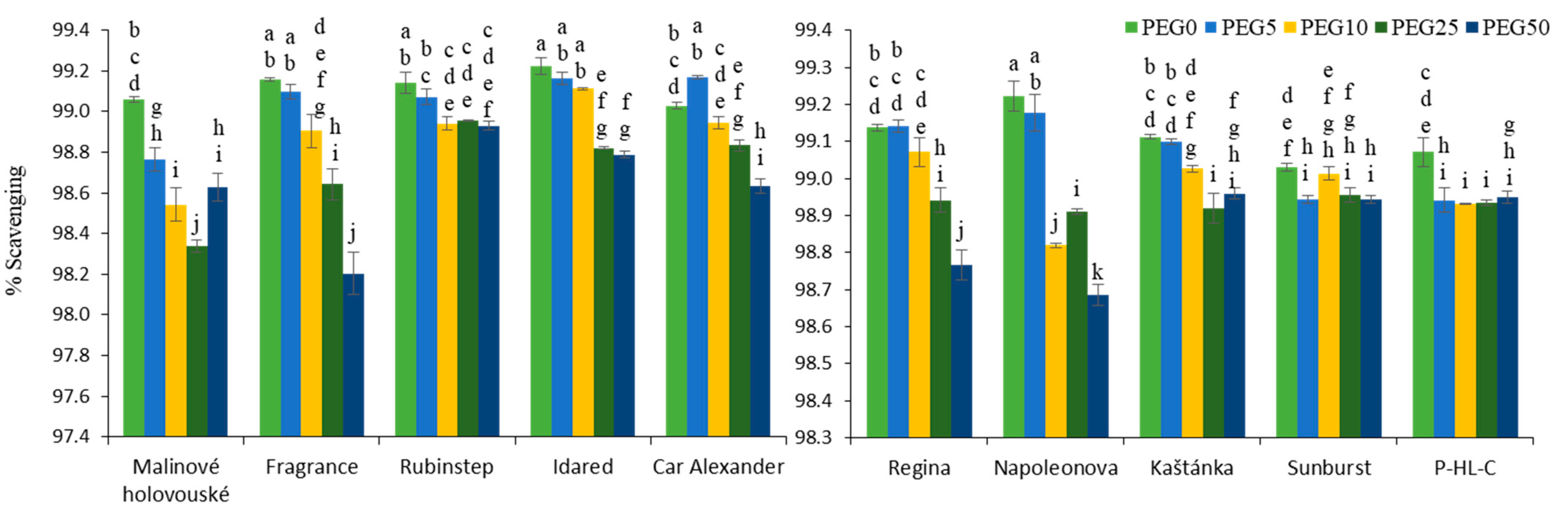
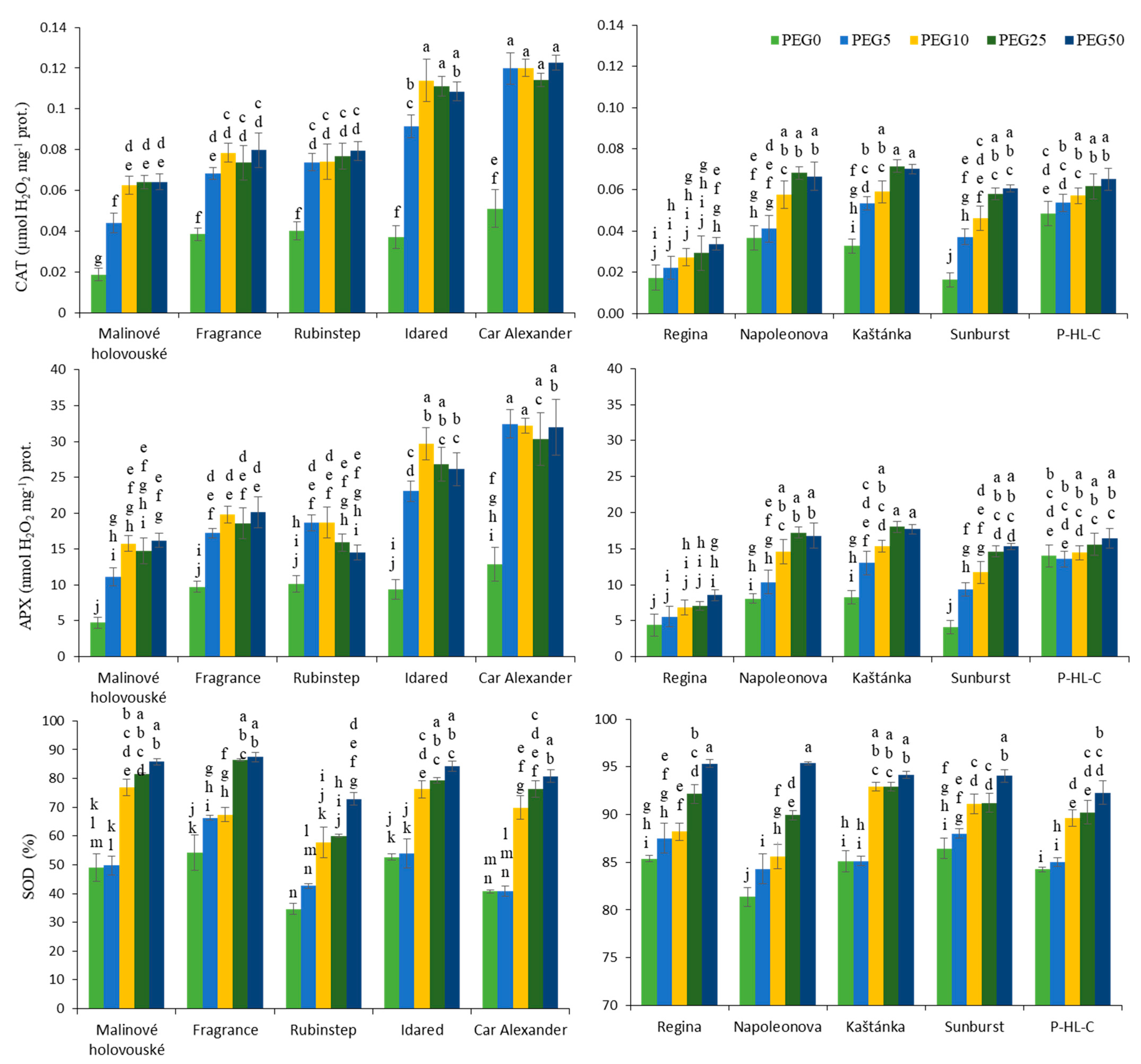
3.3. Antioxidant Capacity and Antioxidant Enzymes Activity
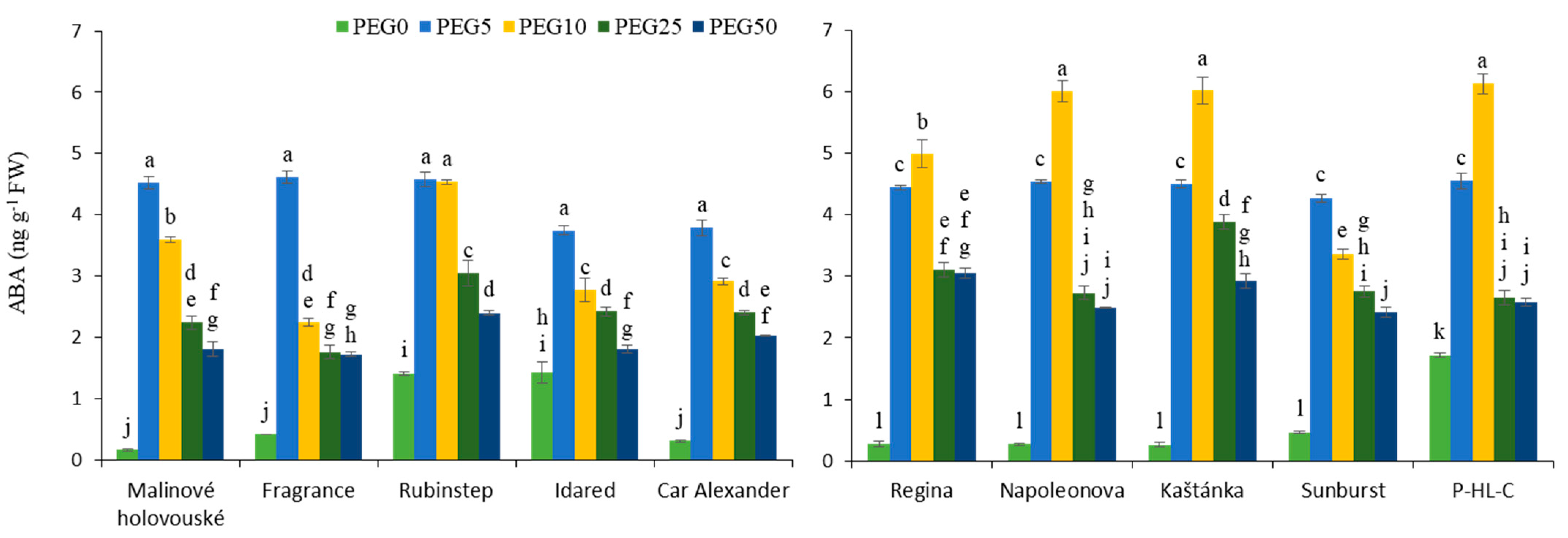
3.4. Abscisic Acid Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santander, C.; Aroca, R.; Ruiz-Lozano, J.M.; Olave, J.; Cartes, P.; Borie, F.; Cornejo, P. Arbuscular Mycorrhiza Effects on Plant Performance under Osmotic Stress. Mycorrhiza 2017, 27, 639–657. [Google Scholar] [CrossRef]
- Swapna, S.; Shylaraj, K.S. Screening for Osmotic Stress Responses in Rice Varieties under Drought Condition. Rice Sci. 2017, 24, 253–263. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant Salt-Tolerance Mechanism: A Review. Biochem. Biophys. Res. Commun. 2018, 496, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Amini, S.; Ghobadi, C.; Yamchi, A. Proline Accumulation and Osmotic Stress: An Overview of P5CS Gene in Plants. J. Plant Mol. Breed. 2015, 3, 44–55. [Google Scholar]
- Saddhe, A.A.; Manuka, R.; Penna, S. Plant Sugars: Homeostasis and Transport under Abiotic Stress in Plants. Physiol. Plant. 2020, 171, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Beszterda, M.; Goliński, P. Nonenzymatic Antioxidants in Plants. In Oxidative Damage to Plants; Elsevier: Amsterdam, The Netherlands, 2014; pp. 201–234. ISBN 978-0-12-799963-0. [Google Scholar]
- Zhang, C.; Shi, S. Physiological and Proteomic Responses of Contrasting Alfalfa (Medicago Sativa L.) Varieties to PEG-Induced Osmotic Stress. Front. Plant Sci. 2018, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide Dismutase—Mentor of Abiotic Stress Tolerance in Crop Plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef]
- Leung, D.W.M. Studies of Catalase in Plants under Abiotic Stress. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 27–39. ISBN 978-3-319-75087-3. [Google Scholar]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [Green Version]
- Asghar, M.A.; Li, Y.; Jiang, H.; Sun, X.; Ahmad, B.; Imran, S.; Yu, L.; Liu, C.; Yang, W.; Du, J. Crosstalk between Abscisic Acid and Auxin under Osmotic Stress. Agron. J. 2019, 111, 2157–2162. [Google Scholar] [CrossRef]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of Polyamines, Abscisic Acid and Proline under Osmotic Stress in the Leaves of Wheat Plants. Sci. Rep. 2018, 8, 12839. [Google Scholar] [CrossRef] [PubMed]
- Shivakrishna, P.; Ashok Reddy, K.; Manohar Rao, D. Effect of PEG-6000 Imposed Drought Stress on RNA Content, Relative Water Content (RWC), and Chlorophyll Content in Peanut Leaves and Roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar]
- Zhang, Z.; Igathinathane, C.; Li, J.; Cen, H.; Lu, Y.; Flores, P. Technology Progress in Mechanical Harvest of Fresh Market Apples. Comput. Electron. Agric. 2020, 175, 105606. [Google Scholar] [CrossRef]
- Acero, N.; Gradillas, A.; Beltran, M.; García, A.; Muñoz Mingarro, D. Comparison of Phenolic Compounds Profile and Antioxidant Properties of Different Sweet Cherry (Prunus avium L.) Varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef]
- Blažek, J. Odrůda jabloně Rubinstep. Vědecké Práce Ovocnářské 2001, 17, 163–165. [Google Scholar]
- Boček, S. Ovocné Dřeviny v Krajině: Pilotní vzdělávací Program, Hostětín 2007/8: Sborník Přednášek a Seminárních Prací; ZO ČSOP Veronica: Brno, Czech Republic, 2008; ISBN 978-80-904109-2-3. [Google Scholar]
- Nove Odrudy Ovoce = New Cultivars of Fruit; Vyzkumny a slechtitelsky ustav ovocnarsky Holovousy: Holovousy, Czech Republic, 2007; ISBN 978-80-87030-04-2.
- Kutina, J.; Holeček, S. Pomologický Atlas 2; Brázda: Praha, Czech Republic, 1992; ISBN 978-80-209-0192-7. [Google Scholar]
- Lane, W.D.; Schmid, H. Lapins and Sunburst sweet cherry. Can. J. Plant Sci. 1984, 64, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Blažková, J. Pěstování Třešní na Slabě Rostoucích Podnožích; Výzkumný a šlechtitelský Ústav Ovocnářský Holovousy: Holovousy, Czech Republic, 2005; ISBN 978-80-902636-8-0. [Google Scholar]
- Kutina, J. Pomologický Atlas 1; Brázda: Praha, Czech Republic, 1991; ISBN 80-209-0089-6. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A Re-Examination of the Relative Turgidity Techniques for Estimating Water Deficits in Leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E.; Zakaria, Z.A. Reactivity of Phenolic Compounds towards Free Radicals under In Vitro Conditions. J. Food Sci. Technol. 2015, 52, 5790–5798. [Google Scholar] [CrossRef] [Green Version]
- Kováčik, J.; Babula, P.; Hedbavny, J. Comparison of Vascular and Non-Vascular Aquatic Plant as Indicators of Cadmium Toxicity. Chemosphere 2017, 180, 86–92. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dobrev, P.I.; Vankova, R. Quantification of Abscisic Acid, Cytokinin, and Auxin Content in Salt-Stressed Plant Tissues. In Plant Salt Tolerance; Shabala, S., Cuin, T.A., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 251–261. ISBN 978-1-61779-985-3. [Google Scholar]
- Alizadeh, V.; Shokri, V.; Soltani, A.; Yousefi, M.A. Effects of Climate Change and Drought-Stress on Plant Physiology. Int. J. Adv. Biol. Biomed. Res. 2015, 3, 38–42. [Google Scholar]
- Wang, W.; Liang, D.; Li, C.; Hao, Y.; Ma, F.; Shu, F. Influence of drought stress on the cellular ultrastructure and antioxidant system in leaves of drought-tolerant and drought-sensitive apple rootstocks. Plant Physiol. Biochem. 2012, 51, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Yadollahi, A.; Nazari-Moghadam, R.; Imani, A.; Arzani, K. Changes in some physiological and osmotic parameters of several pistachio genotypes under drought stress. J. Biol. Environ. Sci. 2012, 6, 263–270. [Google Scholar]
- Khoyerdi, F.F.; Shamshiri, M.H.; Estaji, A. In vitro Screening of Almond (Prunus dulcis (Mill.)) Genotypes for Drought Tolerance. Sci. Hortic. 2016, 198, 44–51. [Google Scholar] [CrossRef]
- Akbarpour, E.; Imani, A.; Yeganeh, S.F. Physiological and Morphological Responses of Almond Cultivars under In Vitro Drought Stress. J. Nuts 2017, 8, 61–72. [Google Scholar]
- Jiménez, S.; Dridi, J.; Gutiérrez, D.; Moret, D.; Irigoyen, J.J.; Moreno, M.A.; Gogorcena, Y. Physiological, biochemical and molecular responses in four Prunus rootstocks submitted to drought stress. Tree Physiol. 2013, 33, 1061–1075. [Google Scholar] [CrossRef]
- Sivritepe, N.; Erturk, U.; Yerlikaya, C.; Turkan, I.; Bor, M.; Ozdemir, F. Response of the cherry rootstock to water stress induced in vitro. Biol. Plant. 2008, 52, 573–576. [Google Scholar] [CrossRef]
- Karimi, S.; Hojati, S.; Eshghi, S.; Moghaddam, R.N.; Jandoust, S. Magnetic exposure improves tolerance of fig ‘Sabz’ explants to drought stress induced in vitro. Sci. Hortic. 2012, 137, 95–97. [Google Scholar] [CrossRef]
- Turkan, I.; Bor, M.; Ozdemir, F.; Koca, H. Differential responses of lipid peroxidation and antioxidants in the leaves of drought—Tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci. 2005, 168, 223–231. [Google Scholar] [CrossRef]
- Říha, J. České Ovoce–díl III. Jablka; České nakladatelství: Praha, Czech Republic, 1919. [Google Scholar]
- Zhong, Y.-P.; Li, Z.; Bai, D.-F.; Qi, X.-J.; Chen, J.-Y.; Wei, C.-G.; Lin, M.-M.; Fang, J.-B. In Vitro Variation of Drought Tolerance in Five Actinidia Species. J. Am. Soc. Hortic. Sci. 2018, 143, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.; Yin, C.; Li, C. Differences in some morphological, physiological, and biochemical responses to drought stress in two contrasting populations of Populus przewalskii. Physiol. Plant. 2006, 127, 182–191. [Google Scholar] [CrossRef]
- Kautz, B.; Noga, G.; Hunsche, M. PEG and drought cause distinct changes in biochemical, physiological and morphological parameters of apple seedlings. Acta Physiol. Plant. 2015, 37, 162. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Piwowarczyk, B.; Kamińska, I.; Rybiński, W. Influence of PEG Generated Osmotic Stress on Shoot Regeneration and Some Biochemical Parameters in Lathyrus Culture. Czech J. Genet. Plant Breed. 2014, 50, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Thippeswamy, M.; Chandraobulreddy, P.; Sinilal, B.; Kumar, M.S.; Sudhakar, C. Proline accumulation and the expression of Δ1-pyrroline-5-carboxylate synthetase in two safflower cultivars. Biol. Plant. 2010, 54, 386–390. [Google Scholar] [CrossRef]
- Molinari, H.B.C.; Marur, C.J.; Daros, E.; Campos, M.K.F.; Carvalho, J.F.P.R.; Bespalhok Filho, J.C.; Pereira, L.F.P.; Vieira, L.G.E. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): Osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiol. Plant. 2007, 130, 218–229. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Li, C.; Zhang, Z.; Ma, F.; Li, M. Response of sugar metabolism in apple leaves subjected to short-term drought stress. Plant Physiol. Biochem. 2019, 141, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Šircelj, H.; Tausz, M.; Grill, D.; Batič, F. Detecting different levels of drought stress in apple trees (Malus domestica Borkh.) with selected biochemical and physiological parameters. Sci. Hortic. 2007, 113, 362–369. [Google Scholar] [CrossRef]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- De Campos, M.K.F.; de Carvalho, K.; de Souza, F.S.; Marur, C.J.; Pereira, L.F.P.; Filho, J.C.B.; Vieira, L.G.E. Drought tolerance and antioxidant enzymatic activity in transgenic ‘Swingle’ citrumelo plants over-accumulating proline. Environ. Exp. Bot. 2011, 72, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Akram, N.A.; Al-Qurainy, F.; Foolad, M.R. Drought tolerance: Roles of organic osmolytes, growth regulators, and mineral nutrientes. Adv. Agron. 2011, 111, 249–296. [Google Scholar]
- Dar, N.A.; Amin, I.; Wani, V.; Wani, S.A.; Shikari, A.B.; Wani, S.H.; Masoodi, K.Z. Abscisic acid: A key regulator of abiotic stress tolerance in plants. Plant Gene 2017, 11, 106–111. [Google Scholar] [CrossRef]
- Kowitcharoen, L.; Wongs-Aree, C.; Setha, S.; Komkhuntod, R.; Srilaong, V.; Kondo, S. Changes in abscisic acid and antioxidant activity in sugar apples underdrought conditions. Sci. Hortic. 2015, 193, 1–6. [Google Scholar] [CrossRef]
- Perin, E.C.; da Silva Messias, R.; Borowski, J.M.; Crizel, R.L.; Schott, I.B.; Carvalho, I.R.; Rombaldi, C.V.; Galli, V. ABA-dependent salt and drought stress improve strawberry fruit quality. Food Chem. 2019, 271, 516–526. [Google Scholar] [CrossRef]
- Bielsa, B.; Leida, C.; Rubio-Cabetas, M.J. Physiological characterization of drought stress response and expression of two transcription factors and two LEA genes in three Prunus genotypes. Sci. Hortic. 2016, 213, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Tworkoski, T.; Fazio, G.; Glenn, D.M. Apple rootstock resistance to drought. Sci. Hortic. 2016, 204, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Ismail, M.R.; Davies, W.J.; Awad, M.H. Leaf growth and stomatal sensitivity to ABA in droughted pepper plants. Sci. Hortic. 2002, 96, 313–327. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Li, N.; Chen, Z.; Xu, K. Comparative transcriptome analysis of the regulation of ABA signaling genes in different rootstock grafted tomato seedlings under drought stress. Environ. Exp. Bot. 2019, 166, 103814. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiroutova, P.; Kovalikova, Z.; Toman, J.; Dobrovolna, D.; Andrys, R. Complex Analysis of Antioxidant Activity, Abscisic Acid Level, and Accumulation of Osmotica in Apple and Cherry In Vitro Cultures under Osmotic Stress. Int. J. Mol. Sci. 2021, 22, 7922. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157922
Jiroutova P, Kovalikova Z, Toman J, Dobrovolna D, Andrys R. Complex Analysis of Antioxidant Activity, Abscisic Acid Level, and Accumulation of Osmotica in Apple and Cherry In Vitro Cultures under Osmotic Stress. International Journal of Molecular Sciences. 2021; 22(15):7922. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157922
Chicago/Turabian StyleJiroutova, Petra, Zuzana Kovalikova, Jakub Toman, Dominika Dobrovolna, and Rudolf Andrys. 2021. "Complex Analysis of Antioxidant Activity, Abscisic Acid Level, and Accumulation of Osmotica in Apple and Cherry In Vitro Cultures under Osmotic Stress" International Journal of Molecular Sciences 22, no. 15: 7922. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22157922