
Activation of Cx43 Hemichannels Induces the Generation of Ca2+ Oscillations in White Adipocytes and Stimulates Lipolysis


Abstract
:1. Introduction
2. Results
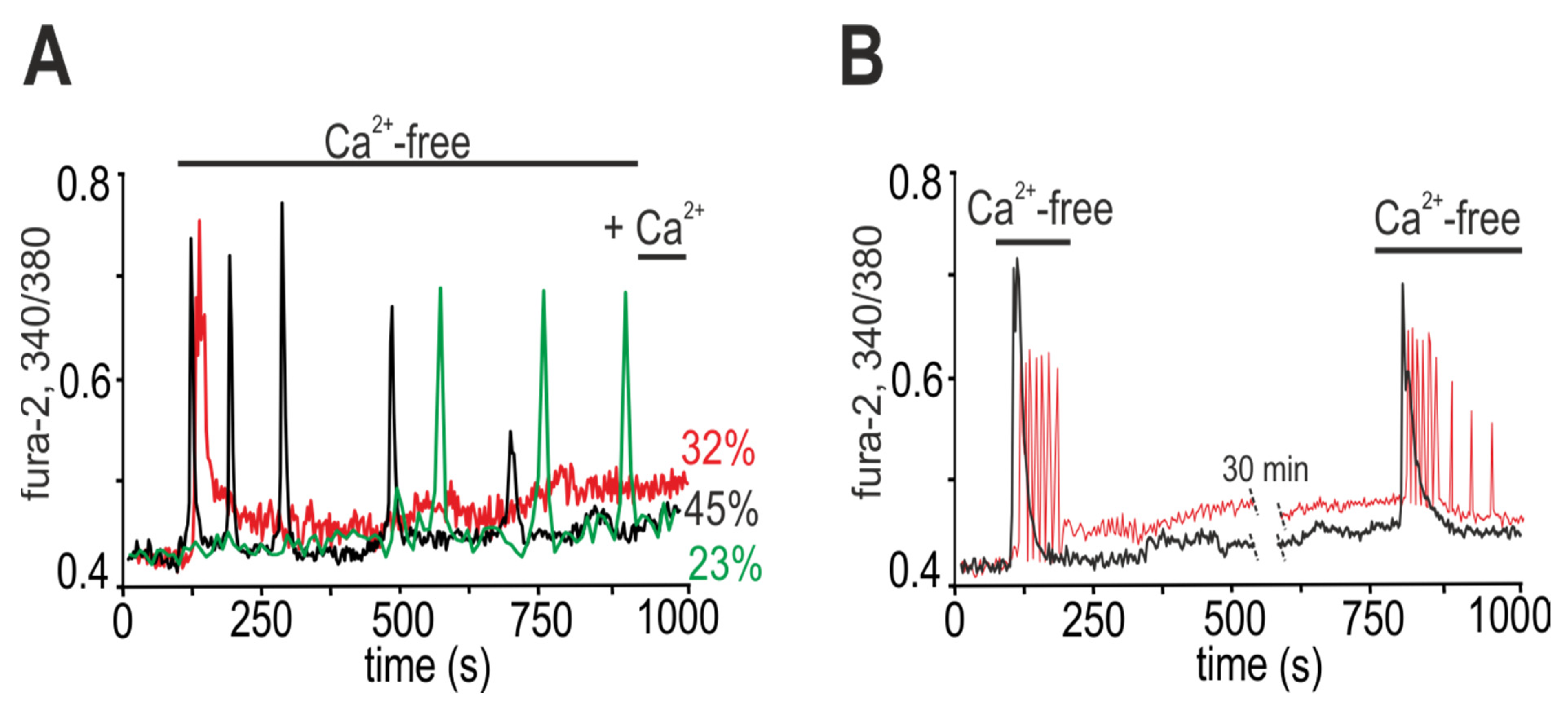
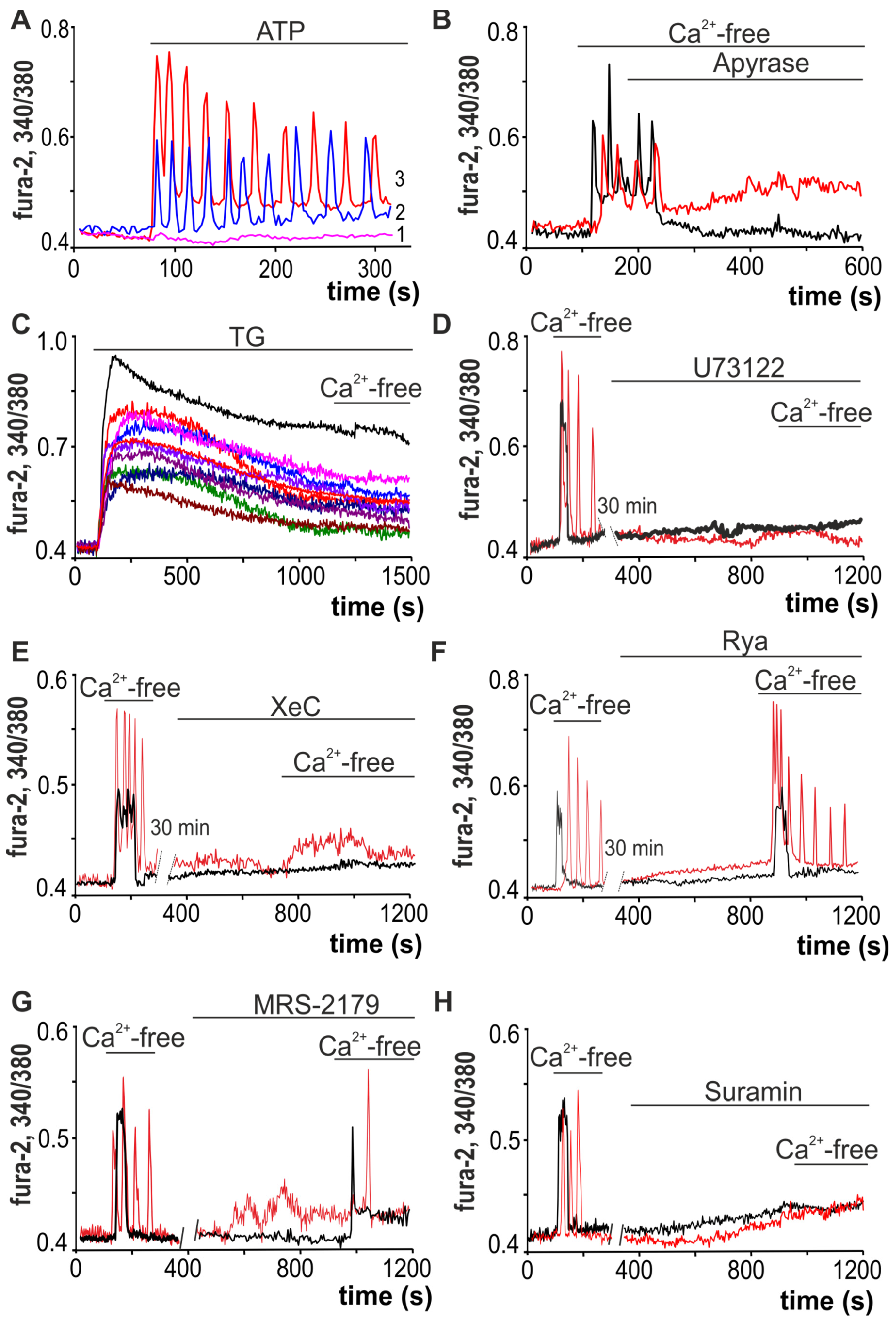
2.1. Ca2+ Responses of White Adipocytes to Ca2+-Free Medium Application
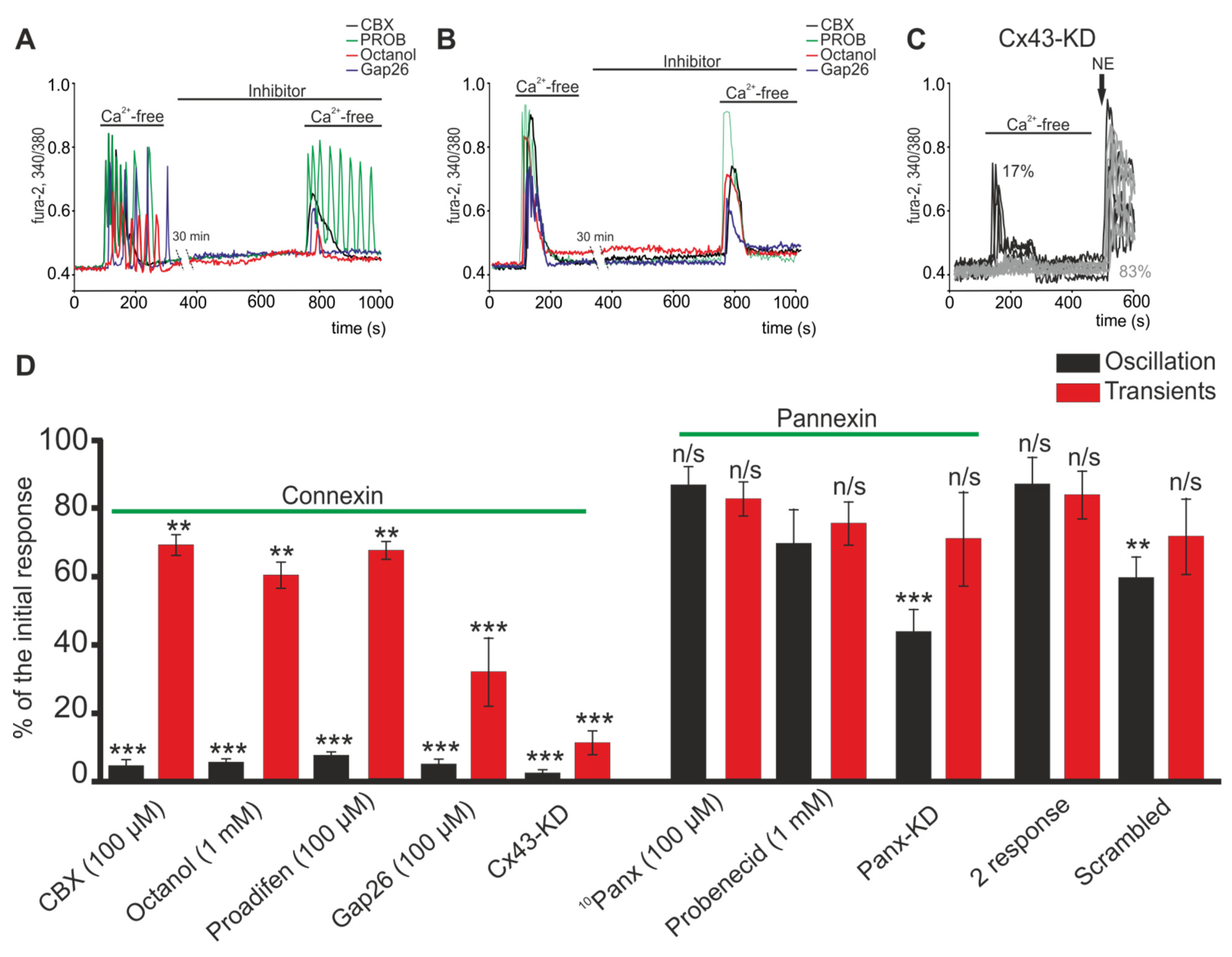
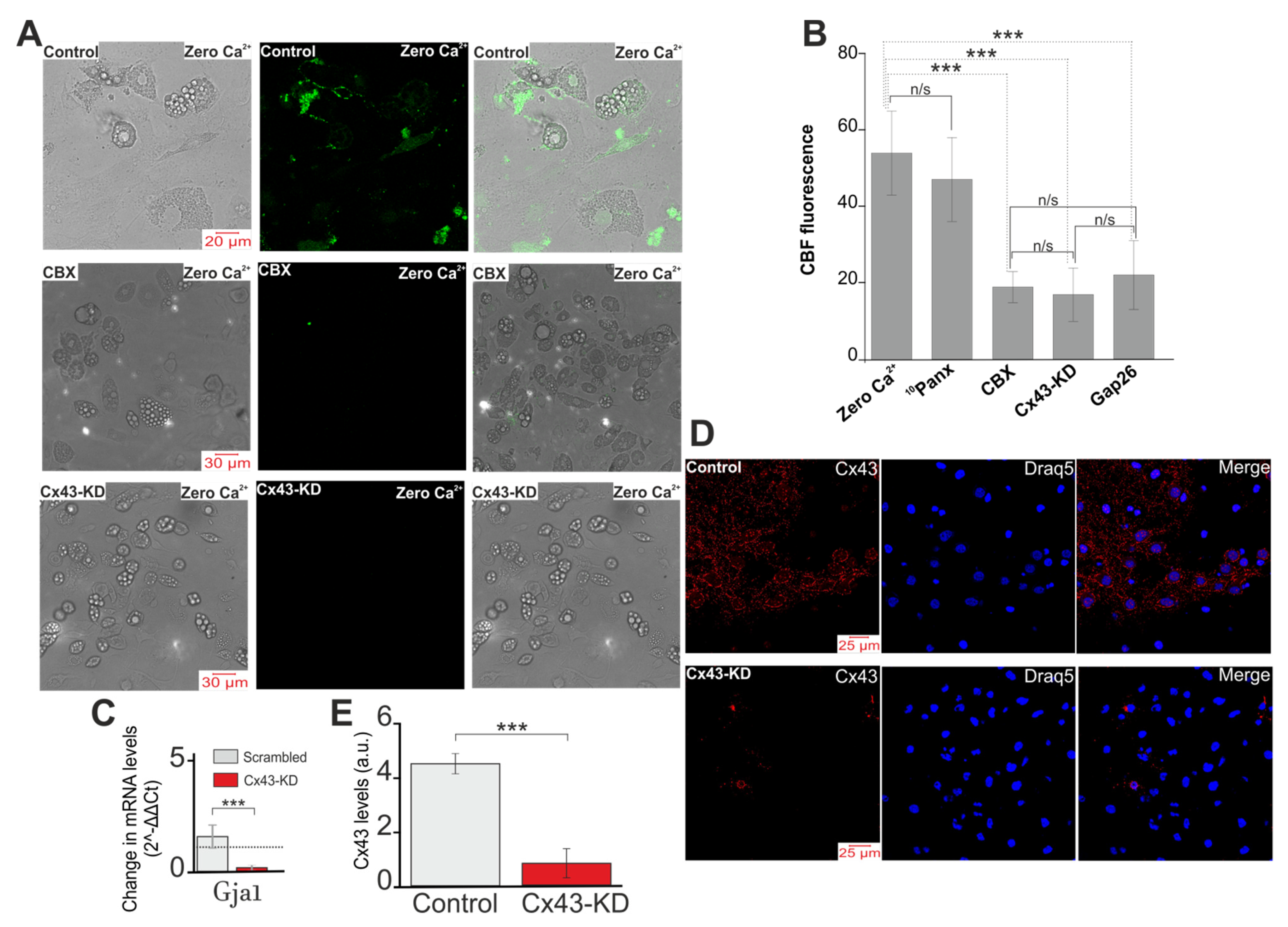
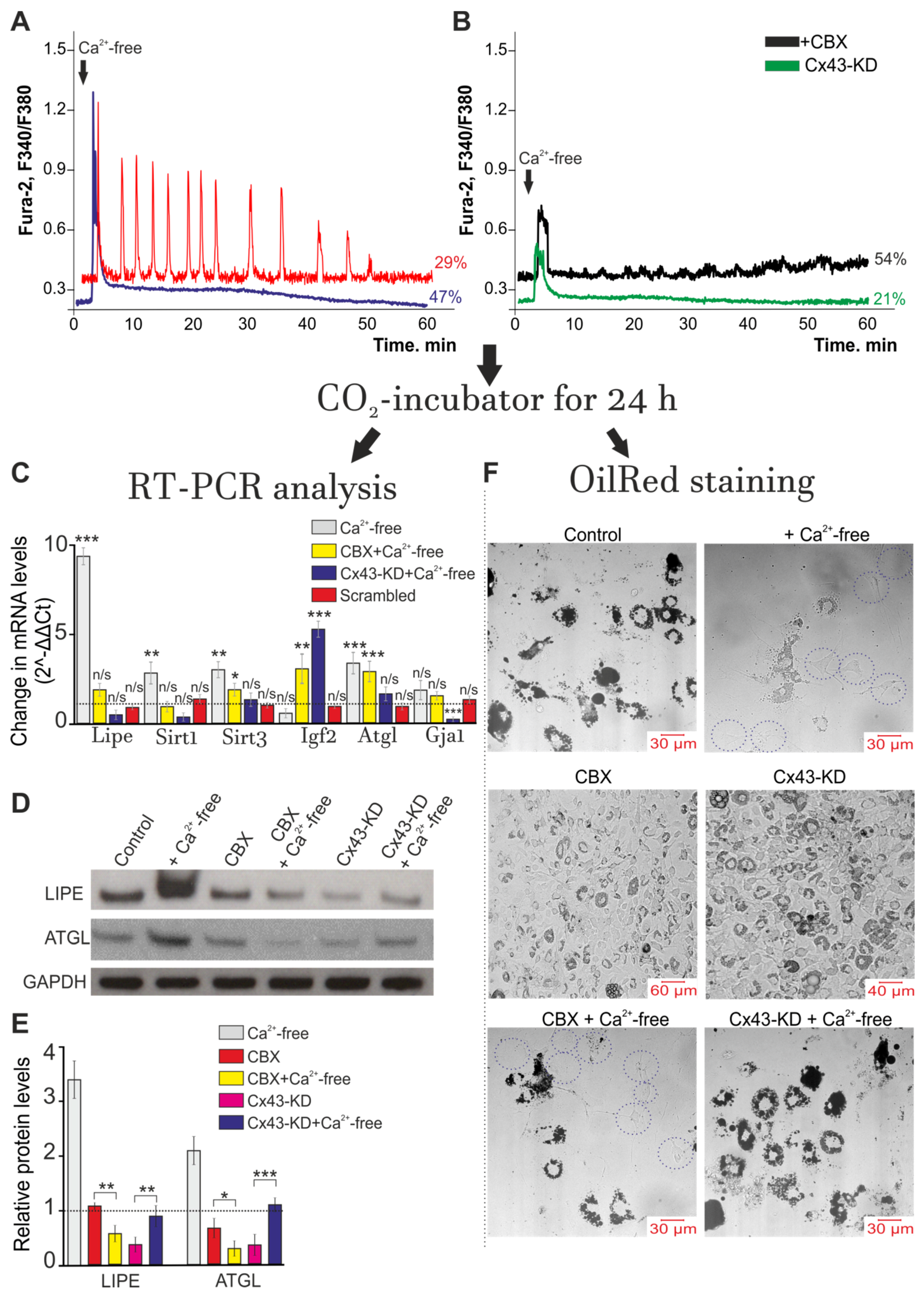
2.2. Activation of Cx43 Connexin Hemichannels Promotes Generation of Ca2+ Oscillations in Response to a Low Extracellular Ca2+ Level but This Has No Effect on Ca2+ Transients
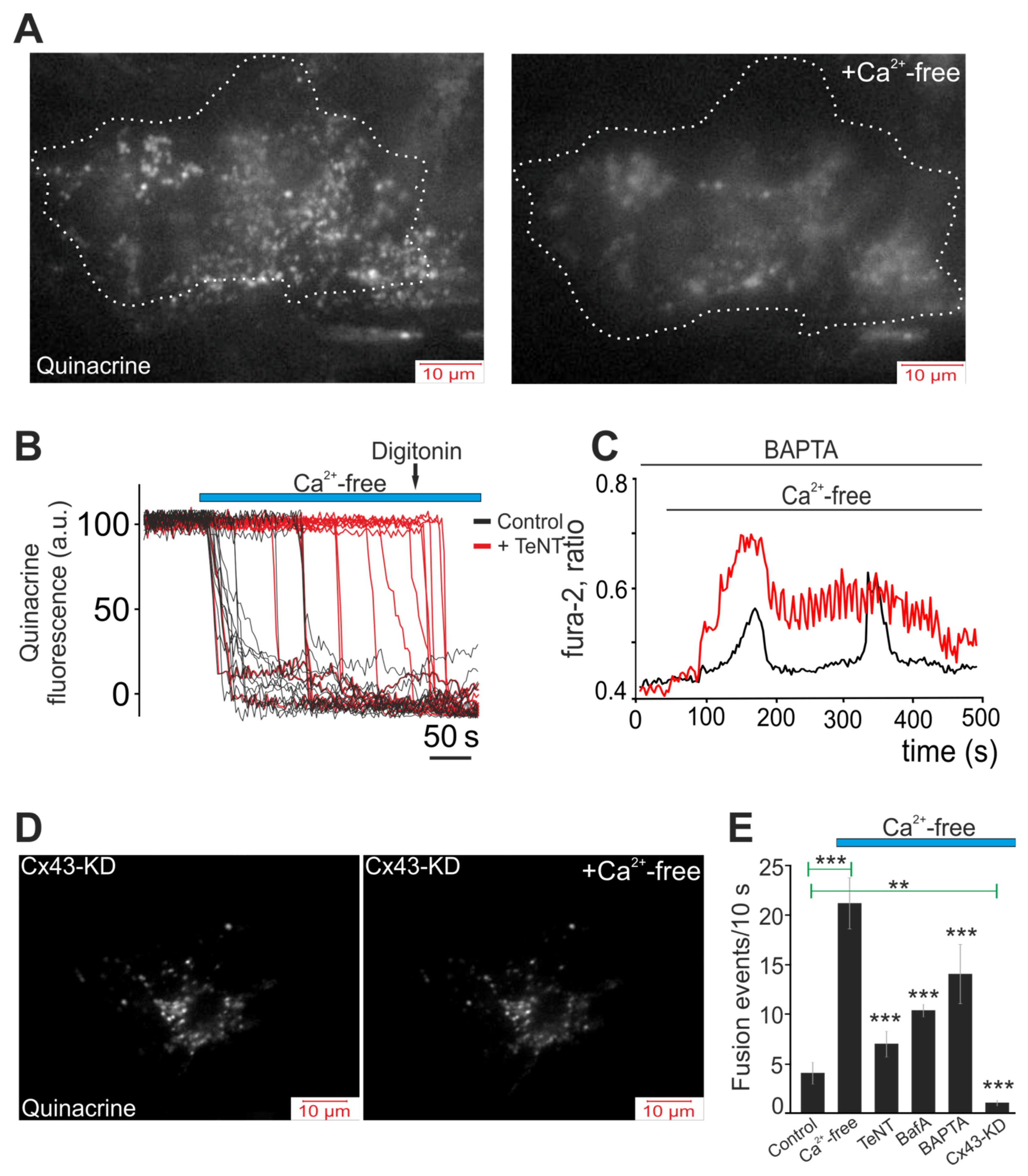
2.3. Decrease in Extracellular Ca2+ Ions Induces Vesicular ATP Release by White Adipocytes
2.4. Activation of Cx43 in Response to a Decrease in [Ca2+]ex Contributes to Phosphoinositide Signaling Pathway and Activation of G-proteins
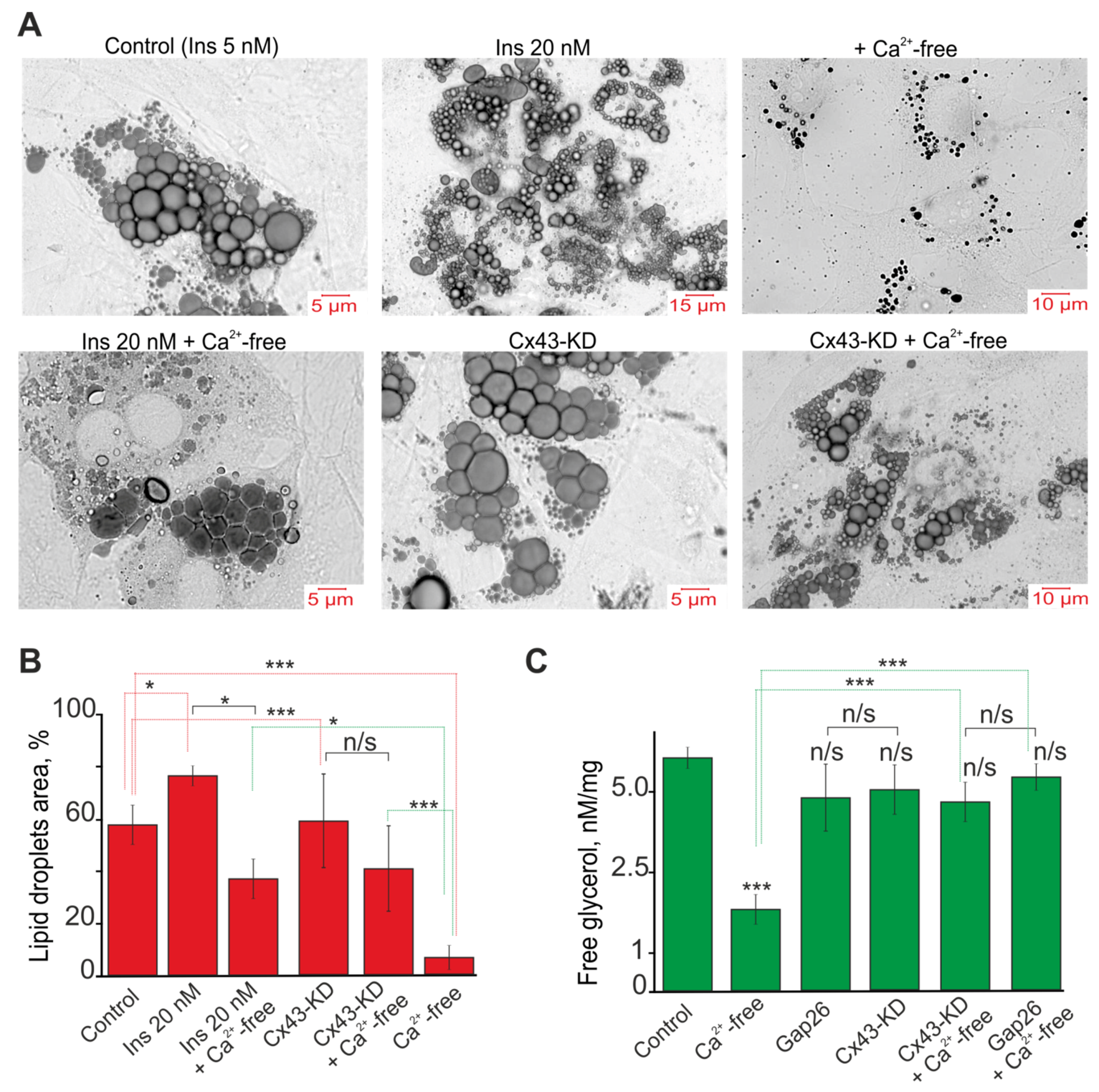
2.5. A Decrease in [Ca2+]ex Stimulates Lipolysis and Correlates with Generation of Ca2+ Oscillations in White Adipocytes
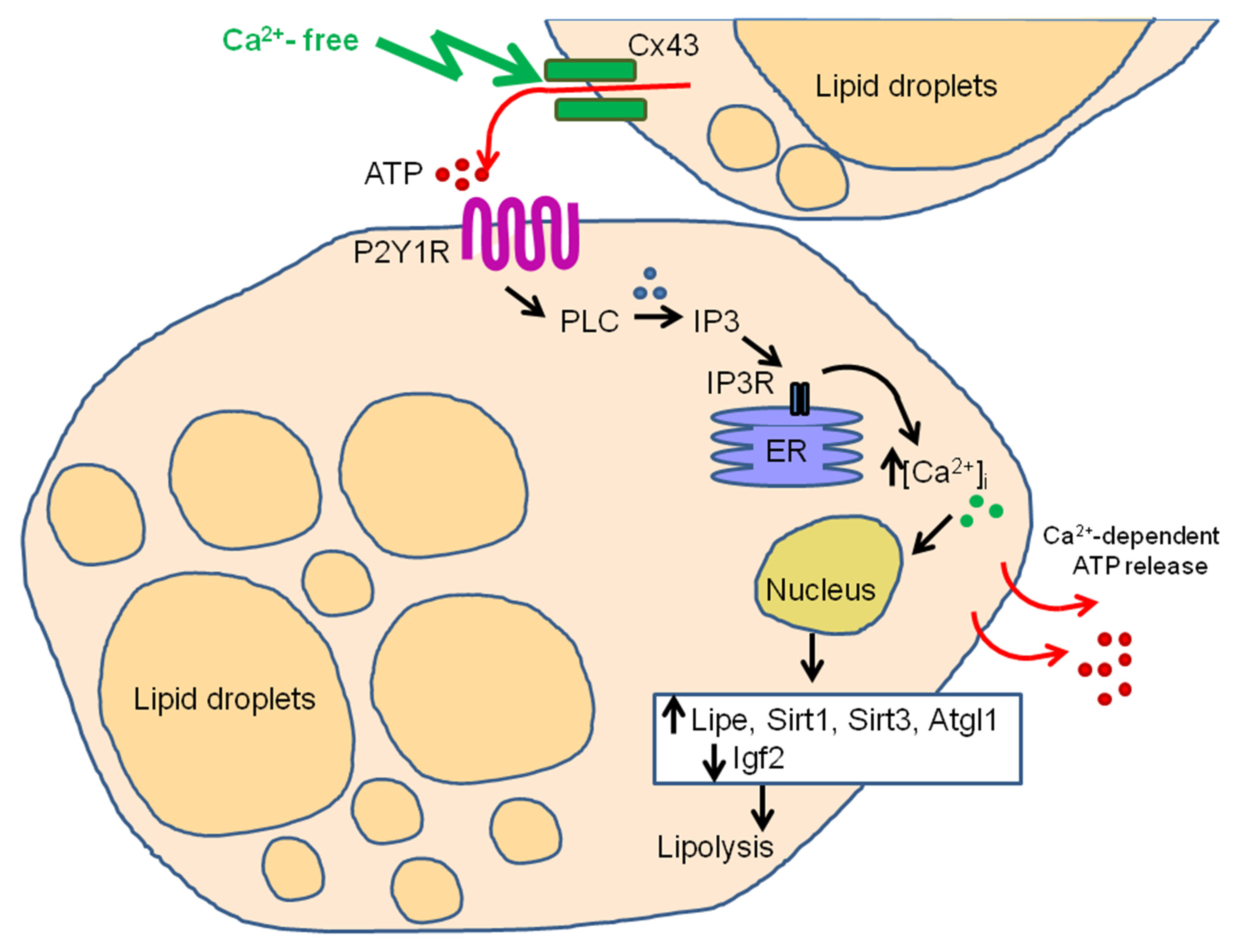
3. Discussion
4. Materials and Methods
4.1. Isolation of Primary White Preadipocytes
4.2. Cultures of Primary White Preadipocytes
4.3. Transfection with Small Interfering RNA (siRNA)
4.4. Immunocytochemical Staining of Adipocytes
4.5. The Measurement of Cytosolic Calcium Concentration
4.6. Assessment of Hemichannel Open Probability by Dye Loading
4.7. Quinacrine Staining
4.8. Total Internal Reflection Fluorescence (TIRF) Microscopy
4.9. Total RNA Isolation
4.10. Real-Time Polymerase Chain Reaction (RT-qPCR)
4.11. Western Blot
4.12. Lipid Droplets Assessment
4.13. Free Glycerol Release Assay
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Compliance with Ethical Standards
References
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-induced changes in adipose tissue microenvironment and their impact on cardiovascular disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Gao, Y.; Tao, C.; Shao, M.; Zhao, S.; Huang, W.; Yao, T.; Johnson, J.A.; Liu, T.; Cypess, A.M.; et al. Connexin 43 mediates white adipose tissue beiging by facilitating the propagation of sympathetic neuronal signals. Cell Metab. 2016, 24, 420–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collaboration NRF. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4.4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Kusminski, C.M.; Bickel, P.E.; Scherer, P.E. Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat. Rev. Drug Discov. 2016, 15, 639–660. [Google Scholar] [CrossRef]
- Suchacki, K.J.; Tavares, A.S.; Mattiucci, D.; Scheller, E.L.; Papanastasiou, G.; Gray, C.; Sinton, M.C.; Ramage, L.E.; McDougald, W.A.; Lovdel, A. Bone marrow adipose tissue is a unique adipose subtype with distinct roles in glucose homeostasis. Nat. Commun. 2020, 11, 3097. [Google Scholar] [CrossRef]
- Gburcik, V.; Cawthorn, W.P.; Nedergaard, J.; Timmons, J.A.; Cannon, B. An essential role for Tbx15 in the differentiation of brown and “brite” but not white adipocytes. Am. J. Physiol. Endocrinol. Metab. 2012, 303, 1053–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; De Vuyst, E.; Ponsaerts, R.; Boengler, K.; Palacios-Prado, N.; Wauman, J.; Lai, C.P.; De Bock, M.; Decrock, E.; Bol, M.; et al. Selective inhibition of Cx43 hemichannels by Gap19 and its impact on myocardial ischemia/reperfusion injury. Basic Res. Cardiol. 2013, 108, 309. [Google Scholar] [CrossRef] [Green Version]
- Bays, H.E.; González-Campoy, J.M.; Bray, G.A.; Kitabchi, A.E.; Bergman, D.A.; Schorr, A.B.; Rodbard, H.W.; Henry, R.R. Pathogenic potential of adipose tissue and metabolic consequences of adipocyte hypertrophy and increased visceral adiposity. Expert. Rev. Cardiovasc. Ther. 2008, 6, 343–368. [Google Scholar] [CrossRef] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, E.; Westermark, P.O.; Spalding, K.L.; Britton, T.; Rydén, M.; Frisén, J.; Bernard, S.; Arner, P. Adipocyte turnover: Relevance to human adipose tissue morphology. Diabetes 2010, 59, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Bays, H.E.; Laferrère, B.; Dixon, J.; Aronne, L.; González-Campoy, J.M.; Apovian, C.; Wolfe, B.M. Adiposopathy and bariatric surgery: Is ‘sick fat’ a surgical disease? Int. J. Clin. Pract. 2009, 63, 1285–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Wong, J.C.; Fiscus, R.R. Essential roles of the nitric oxide (NO)/cGMP/protein kinase G type-Iα (PKG-Iα) signaling pathway and the atrial natriuretic peptide (ANP)/cGMP/PKG-Iα autocrine loop in promoting proliferation and cell survival of OP9 bone marrow stromal cells. J. Cell Biochem. 2011, 112, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Verselis, V.K.; Trelles, M.P.; Rubinos, C.; Bargiello, T.A.; Srinivas, M. Loop gating of connexin hemichannels involves movement of pore-lining residues in the first extracellular loop domain. J. Biol. Chem. 2009, 284, 4484–4493. [Google Scholar] [CrossRef] [Green Version]
- Zemel, M.B.; Shi, H.; Greer, B.; Dirienzo, D.; Zemel, P.C. Regulation of adiposity by dietary calcium. FASEB J. 2000, 14, 1132–1138. [Google Scholar] [CrossRef]
- Zemel, M.B. Calcium modulation of hypertension and obesity: Mechanisms and implications. J. Am. Coll. Nutr. 2001, 20, 428–435. [Google Scholar] [CrossRef]
- De Bock, M.; Wang, N.; Bol, M.; Decrock, E.; Ponsaerts, R.; Bultynck, G.; Dupont, G.; Leybaert, L. Connexin 43 hemichannels contribute to cytoplasmic Ca2+ oscillations by providing a bimodal Ca2+-dependent Ca2+ entry pathway. J. Biol. Chem. 2012, 287, 12250–12266. [Google Scholar] [CrossRef] [Green Version]
- Van Campenhout, R.; Gomes, A.R.; De Groof, T.W.M.; Muyldermans, S.; Devoogdt, N.; Vinken, M. Mechanisms underlying connexin hemichannel activation in disease. Int. J. Mol. Sci. 2021, 22, 3503. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Halvorsen, Y.D.; Ellis, P.N.; Wilkison, W.O.; Zemel, M.B. Role of intracellular calcium in human adipocyte differentiation. Physiol. Genom. 2000, 3, 75–82. [Google Scholar] [CrossRef]
- Szabo, E.; Qiu, Y.; Baksh, S.; Michalak, M.; Opas, M. Calreticulin inhibits commitment to adipocyte differentiation. J. Cell. Biol. 2008, 182, 103–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turovsky, E.A.; Turovskaya, M.V.; Dynnik, V.V. Deregulation of Ca2+-signaling systems in white adipocytes, manifested as the loss of rhythmic activity, underlies the development of multiple hormonal resistance at obesity and type 2 diabetes. Int. J. Mol. Sci. 2021, 22, 5109. [Google Scholar] [CrossRef]
- Guerrero-Hernandez, A.; Verkhratsky, A. Calcium signalling in diabetes. Cell. Calcium. 2014, 56, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Kawai, A. The role of calmodulin in hormone-stimulated lipolysis. Metabolism 1985, 34, 303–308. [Google Scholar] [CrossRef]
- Izawa, T.; Komabayashi, T. Ca2+ and lipolysis in adipocytes from exercise-trained rats. J. Appl. Physiol. 1994, 77, 2618–2624. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Suzumura, A. Gap junctions and hemichannels composed of connexins: Potential therapeutic targets for neurodegenerative diseases. Front. Cell. Neurosci. 2014, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Castellano, P.; Eugenin, E. A Regulation of gap junction channels by infectious agents and inflammation in the CNS. Front. Cell. Neurosci. 2014, 8, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Numano, R.; Yokota-Hashimoto, H.; Matsui Sho Kimura, N.; Takeuchi, H.; Kitamura, T. A central-acting connexin inhibitor, INI-0602, prevents high-fat diet-induced feeding pattern disturbances and obesity in mice. Mol. Brain 2018, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehberg, J.; Moraga-Amaro, R.; Salazar, C.; Becerra, A.; Echeverría, C.; Orellana, J.A.; Bultynck, G.; Ponsaerts, R.; Leybaert, L.; Simon, F.; et al. Release of gliotransmitters through astroglial connexin 43 hemichannels is necessary for fear memory consolidation in the basolateral amygdale. FASEB J. 2012, 26, 3649–3657. [Google Scholar] [CrossRef] [Green Version]
- Salmina, A.B. Neuron-glia interactions as therapeutic targets in neurodegeneration. J. Alzheimers Dis. 2009, 16, 485–502. [Google Scholar] [CrossRef]
- Azarashvili, T.; Baburina, Y.; Grachev, D.; Krestinina, O.; Evtodienko, Y.; Stricker, R.; Reiser, G. Calcium-induced permeability transition in rat brain mitochondria is promoted by carbenoxolone through targeting connexin43. Am. J. Physiol. Cell. Physiol. 2011, 300, 707–720. [Google Scholar] [CrossRef]
- Boengler, K.; Ungefug, E.; Heusch, G.; Leybaert, L.; Schulz, R. Connexin 43 impacts on mitochondrial potassium uptake. Front. Pharmacol. 2013, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Boengler, K.; Ruiz-Meana, M.; Gent, S.; Ungefug, E.; Soetkamp, D.; Miro-Casas, E.; Cabestrero, A.; Fernandez-Sanz, C.; Semenzato, M.; Di Lisa, F.; et al. Mitochondrial connexin 43 impacts on respiratory complex I activity and mitochondrial oxygen consumption. J. Cell. Mol. Med. 2012, 16, 1649–1655. [Google Scholar] [CrossRef]
- Rottlaender, D.; Boengler, K.; Wolny, M.; Michels, G.; Endres-Becker, J.; Motloch, L.; Schwaiger, A.; Buechert, A.; Schulz, R.; Heusch, G.; et al. Connexin 43 acts as a cytoprotective mediator of signal transduction by stimulating mitochondrial KATP channels in mouse cardiomyocytes. J. Clin. Investig. 2010, 120, 1441–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, J.A.; Richards, T.; Becker, D.L. Connexins and diabetes. Cardiol. Res. Pract. 2012, 2012, 496904. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, R.; Allagnat, F.; Haefliger, J.A.; Meda, P. Connexins, diabetes and the metabolic syndrome. Curr. Protein Pept. Sci. 2009, 10, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, M.; Berberich, O.; Hoefner, C.; Blunk, T.; Bauer-Kreisel, P. Gap junctional intercellular communication in adipose-derived stromal/stem cells is cell density-dependent and positively impacts adipogenic differentiation. J. Cell. Physiol. 2018, 233, 3315–3329. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.N.; Kwon, H.J.; Im, S.W.; Son, Y.H.; Akindehin, S.; Jung, Y.S.; Lee, S.J.; Rhyu, I.J.; Kim, I.Y.; Seong, J.K.; et al. Connexin 43 is required for the maintenance of mitochondrial integrity in brown adipose tissue. Sci. Rep. 2017, 7, 7159. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.R.; Saier, M.H., Jr. Gap junctional proteins of animals: The innexin/pannexin superfamily. Prog. Biophys. Mol. Biol. 2007, 94, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Barbe, M.T.; Monyer, H.; Bruzzone, R. Cell-cell communication beyond connexins: The pannexin channels. Physiology 2006, 21, 103–114. [Google Scholar] [CrossRef]
- Giaume, C.; Leybaert, L.; Naus, C.C.; Sáez, C.J. Connexin and pannexin hemichannels in brain glial cells: Properties, pharmacology, and roles. Front. Pharmacol. 2013, 4, 88. [Google Scholar] [CrossRef] [Green Version]
- Cone, A.C.; Ambrosi, C.; Scemes, E.; Martone, M.E.; Sosinsky, G.E. A comparative antibody analysis of pannexin1 expression in four rat brain regions reveals varying subcellular localizations. Front. Pharmacol. 2013, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Prochnow, N.; Abdulazim, A.; Kurtenbach, S.; Wildförster, V.; Dvoriantchikova, G.; Hanske, J.; Petrasch-Parwez, E.; Shestopalov, V.I.; Dermietzel, R.; Manahan-Vaughan, D.; et al. Pannexin1 stabilizes synaptic plasticity and is needed for learning. PLoS ONE 2012, 7, e51767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamson, S.E.; Meher, A.K.; Chiu, Y.-H.; Sandilos, J.K.; Oberholtzer, N.P.; Walker, N.N.; Hargett, S.R.; Seaman, S.A.; Peirce-Cottler, S.M.; Isakson, B.E.; et al. Pannexin 1 is required for full activation of insulin-stimulated glucose uptake in adipocytes. Mol. Metab. 2015, 4, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Turovsky, E.A.; Turovskaya, M.V.; Berezhnov, A.V.; Tolmacheva, A.V.; Kaimachnikov, N.P.; Dolgacheva, L.P.; Zinchenko, V.P.; Maevskii, E.I.; Dynnik, V.V. Convergence of Ca2+ signaling pathways in adipocytes. The role of L-arginine and protein kinase G in generation of transient and periodic Ca2+ signals. Biochem. Suppl. Ser. A 2012, 6, 35–44. [Google Scholar] [CrossRef]
- Meigh, L.; Greenhalgh, S.A.; Rodgers, T.L.; Cann, M.J.; Roper, D.I.; Dale, N. CO2 directly modulates connexin 26 by formation of carbamate bridges between subunits. Elife 2013, 2, e01213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameritsch, P.; Khandoga, N.; Pohl, U.; Pogoda, K. Gap junctional communication promotes apoptosis in a connexin-type-dependent manner. Cell. Death Dis. 2013, 4, 584. [Google Scholar] [CrossRef]
- Akopova, I.; Tatur, S.; Grygorczyk, M.; Luchowski, R.; Gryczynski, I.; Gryczynski, Z.; Borejdo, J.; Grygorczyk, R. Imaging exocytosis of ATP-containing vesicles with TIRF microscopy in lung epithelial A549 cells. Purinergic. Signal 2012, 8, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turovskaya, M.V.; Gaidin, S.G.; Vedunova, M.V.; Babaev, A.A.; Turovsky, E.A. BDNF overexpression enhances the preconditioning effect of brief episodes of hypoxia, promoting survival of GABAergic neurons. Neurosci. Bull. 2020, 36, 733–760. [Google Scholar] [CrossRef] [Green Version]
- Chung, W.-C.; Kermode, J.C. Suramin disrupts receptor-G protein coupling by blocking association of G protein alpha and betagamma subunits. J. Pharmacol. Exp. Ther. 2005, 313, 191–198. [Google Scholar] [CrossRef]
- Karlstad, J.; Sun, Y.; Brij, B. Ca2+ signaling: An outlook on the characterization of Ca2+ channels and their importance in cellular functions. Adv. Exp. Med. Biol. 2012, 740, 143–157. [Google Scholar]
- Chadjichristos, C.E.; Matter, C.M.; Roth, I.; Sutter, E.; Pelli, G.; Lüscher, T.F.; Chanson, M.; Kwak, B.R. Reduced connexin43 expression limits neointima formation after balloon distension injury in hypercholesterolemic mice. Circulation 2006, 113, 2835–2843. [Google Scholar] [CrossRef]
- Wang, Y.; Sato, M.; Guo, Y.; Bengtsson, T.; Nedergaard, J. Protein kinase a-mediated cell proliferation in brown preadipocytes is independent of Erk1/2, PI3K and mTOR. Exp. Cell. Res. 2014, 328, 143–155. [Google Scholar] [CrossRef]
- Davidson, J.O.; Green, C.R.; Nicholson, L.F.; Bennet, L.; Gunn, A.J. Deleterious effects of high dose connexin 43 mimetic peptide infusion after cerebral ischaemia in near-term fetal sheep. Int. J. Mol. Sci. 2012, 13, 6303–6319. [Google Scholar] [CrossRef] [Green Version]
- Martínez, A.D.; Sáez, J.C. Regulation of astrocyte gap junctions by hypoxia-reoxygenation. Brain Res. Rev. 2000, 32, 250–258. [Google Scholar] [CrossRef]
- De Pina-Benabou, M.H.; Szostak, V.; Kyrozis, A.; Rempe, D.; Uziel, D.; Urban-Maldonado, M.; Benabou, S.; Spray, D.C.; Federoff, H.J.; Stanton, P.K.; et al. Blockade of gap junctions in vivo provides neuroprotection after perinatal global ischemia. Stroke 2005, 36, 2232–2237. [Google Scholar] [CrossRef] [Green Version]
- Theodoric, N.; Bechberger, J.F.; Naus, C.C.; Sin, W.C. Role of gap junction protein connexin43 in astrogliosis induced by brain injury. PLoS ONE 2012, 7, e47311. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.; Nagajyothi, F.; Thi, M.M.; Hanani, M.; Scherer, P.E.; Tanowitz, H.B.; Spray, D.C. Adipocytes in both brown and white adipose tissue of adult mice are functionally connected via gap junctions: Implications for chagas disease. Microbes. Infect. 2014, 16, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.J.; Cuatrecasas, P. Adenosine triphosphate-dependent inhibition of insulin-stimulated glucose transport in fat cells. Possible role of membrane phosphorylation. J. Biol. Chem. 1974, 249, 3170–3180. [Google Scholar] [CrossRef]
- Matsuoka, I.; Zhou, Q.; Ishimoto, H.; Nakanishi, H. Extracellular ATP stimulates adenylyl cyclase and phospholipase C through distinct purinoceptors in NG108-15 cells. Mol. Pharmacol. 1995, 47, 855–862. [Google Scholar]
- Lee, H.; Jun, D.J.; Suh, B.C.; Choi, B.H.; Lee, J.H.; Do, M.S.; Suh, B.S.; Ha, H.; Kim, K.T. Dual roles of P2 purinergic receptors in insulin-stimulated leptin production and lipolysis in differentiated rat white adipocytes. J. Biol. Chem. 2005, 280, 28556–28563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seufert, J.; Kieffer, T.J.; Leech, C.A.; Holz, G.G.; Moritz, W.; Ricordi, C.; Habener, J.F. Leptin suppression of insulin secretion and gene expression in human pancreatic islets: Implications for the development of adipogenic diabetes mellitus. J. Clin. Endocrinol. Metab. 1999, 84, 670–676. [Google Scholar] [CrossRef]
- Westfall, D.P.; Todorov, L.D.; Mihaylova-Todorova, S.T. ATP as a cotransmitter in sympathetic nerves and its inactivation by releasable enzymes. J. Pharmacol. Exp. Ther. 2002, 303, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.A.; Monassier, L.; Freund, M.; Bousquet, P.; Gachet, C. The purinergic P2Y1 receptor supports leptin secretion in adipose tissue. Endocrinology 2010, 151, 2060–2070. [Google Scholar] [CrossRef] [Green Version]
- Madec, S.; Rossi, C.; Chiarugi, M.; Santini, E.; Salvati, A.; Ferrannini, E.; Solini, A. Adipocyte P2X7 receptors expression: A role in modulating inflammatory response in subjects with metabolic syndrome? Atherosclerosis 2011, 219, 552–558. [Google Scholar] [CrossRef]
- Alves, L.A.; de Melo Reis, R.A.; de Souza, C.A.M.; de Freitas, M.S.; Teixeira, P.C.N.; Ferreira, D.N.M.; Xavier, R.F. The P2X7 receptor: Shifting from a low- to a high-conductance channel—An enigmatic phenomenon? Biochim. Biophys. Acta 2014, 1838, 2578–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, S.M.; Pappone, P.A. P2 receptor modulation of voltage-gated potassium currents in brown adipocytes. J. Gen. Physiol. 1999, 113, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, H.H.; Grilc, S.; Zorec, R. Correlated ATP-induced changes in membrane area and membrane conductance in single rat adipocytes. Ann. N. Y. Acad. Sci. 2005, 1048, 281–286. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Turovskaya, M.V.; Dolgacheva, L.P.; Zinchenko, V.P.; Dynnik, V.V. Acetylcholine promotes Ca2+ and NO-oscillations in adipocytes implicating Ca2+→NO→cGMP→cADP-ribose→Ca2+ positive feedback loop. Modulatory effects of norepinephrine and atrial natriuretic peptide. PLoS ONE 2013, 8, e63483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turovsky, E.A.; Kaimachnikov, N.R.; Zinshenko, V.R. Agonist-specific participation of SOC and ARC channels and iPLA2 in the regulation of Ca2+ entry during oscillatory responses in adipocytes. Biochem. Suppl. Ser. 2014, 8, 136–143. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Kaǐmachnikov, N.P.; Turovskaya, M.V.; Berezhnov, A.V.; Dynnik, V.V.; Zinchenko, V.P. Two mechanisms of calcium oscillations in adipocytes. Biochem. Suppl. Ser. 2011, 28, 463–472. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Zinchenko, V.P.; Kaimachnikov, N.P. Attenuation of calmodulin regulation evokes Ca2+ oscillations: Evidence for the involvement of intracellular arachidonate-activated channels and connexons. Mol. Cell. Biochem. 2019, 456, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Sauer, H.; Sharifpanah, F.; Hatry, M.; Steffen, P.; Bartsch, C.; Heller, R.; Padmasekar, M.; Howaldt, H.P.; Bein, G.; Wartenberg, M. NOS inhibition synchronizes calcium oscillations in human adipose tissue-derived mesenchymal stem cells by increasing gap-junctional coupling. J. Cell. Physiol. 2011, 226, 1642–1650. [Google Scholar] [CrossRef]
- Thomas, A.P.; Bird, G.S.; Hajnóczky, G.; Robb-Gaspers, L.D.; Putney, J.W. Spatial and temporal aspects of cellular calcium signaling. FASEB J. 1996, 10, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Keizer, J.; Levine, L. Ryanodine receptor adaptation and Ca2+(−) induced Ca2+ release-dependent Ca2+ oscillations. Biophys. J. 1996, 71, 3477–3487. [Google Scholar] [CrossRef] [Green Version]
- Ghandour, R.A.; Giroud, M.; Vegiopoulos, A.; Herzig, S.; Ailhaud, G.; Amri, E.; Pisani, D.F. IP-receptor and PPARs trigger the conversion of human white to brite adipocyte induced by carbaprostacyclin. Biochim. Biophys. Acta 2016, 1861, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolgacheva, L.P.; Turovskaya, M.V.; Dynnik, V.V.; Zinchenko, V.P.; Goncharov, N.V.; Davletov, B.; Turovsky, E.A. Angiotensin II activates different calcium signaling pathways in adipocytes. Arch. Biochem. Biophys. 2016, 593, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Liang, X.; Sun, Q.; Yu, H.; Zhong, J.; Ni, Y.; Chen, J.; Zhao, Z.; Gao, P.; et al. Activation of TRPV1 channel by dietary capsaicin improves visceral fat remodeling through connexin43-mediated Ca2+ Influx. Cardiovasc. Diabetol. 2015, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolotto, C.; Rosillo, M.; Botti, J.C.; Constenla, B.F.; Anabel, S. “Synapse-like” connections between adipocytes and stem cells: Morphological and molecular features of human adipose tissue. J. Stem Cell Dev. Biol. 2018, 1, 100003. [Google Scholar]
- Shao, Q.; Esseltine, J.L.; Huang, T.; Novielli-Kuntz, N.; Ching, J.E.; Sampson, J.; Laird, D.W. Connexin43 is dispensable for early stage human mesenchymal stem cell adipogenic differentiation but is protective against cell senescence. Biomolecules 2019, 9, 474. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Kim, S.N.; Kim, Y.A.; Lee, Y.H. The contribution of arachidonate 15-lipoxygenase in tissue macrophages to adipose tissue remodeling. Cell. Death Dis. 2016, 7, e2285. [Google Scholar] [CrossRef]
- Mailloux, R.J.; Harper, M.E. Uncoupling proteins and the control of mitochondrial reactive oxygen species production. Free Radical. Biol. Med. 2011, 51, 1106–1115. [Google Scholar] [CrossRef]
- Armani, A.; Berry, A.; Cirulli, F.; Caprio, M. Molecular mechanisms underlying metabolic syndrome: The expanding role of the adipocyte. FASEB J. 2017, 31, 4240–4255. [Google Scholar] [CrossRef]
- Osuga, J.; Ishibashi, S.; Oka, T.; Yagyu, H.; Tozawa, R.; Fujimoto, A.; Shionoiri, F.; Yahagi, N.; Kraemer, F.B.; Tsutsumi, O.; et al. Targeted disruption of hormone-sensitive lipase results in male sterility and adipocyte hypertrophy, but not in obesity. Proc. Natl. Acad. Sci. USA 2000, 97, 787–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortier, M.; Wang, S.P.; Mauriege, P.; Semache, M.; Mfuma, L.; Li, H.; Levy, E.; Richard, D.; Mitchell, G.A. Hormone-sensitive lipase-independent adipocyte lipolysis during beta-adrenergic stimulation, fasting, and dietary fat loading. Am. J. Physiol. Endocrinol. Metab. 2004, 287, 282–288. [Google Scholar] [CrossRef]
- Haemmerle, G.; Zimmermann, R.; Hayn, M.; Theussl, C.; Waeg, G.; Wagner, E.; Sattler, W.; Magin, T.M.; Wagner, E.F.; Zechner, R. Hormone-sensitive lipase deficiency in mice causes diglyceride accumulation in adipose tissue, muscle, and testis. J. Biol. Chem. 2002, 277, 4806–4815. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.J.; Yu, Z.; Patel, S.; Jue, D.; Liu, L.F.; Kraemer, F.B. Hormone-sensitive lipase modulates adipose metabolism through PPARγ. Biochim. Biophys. Acta 2011, 1811, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Sekiya, M.; Osuga, J.I.; Okazaki, H.; Yahagi, N.; Harada, K.; Shen, W.J.; Tamura, Y.; Tomita, S.; Iizuka, Y.; Ohashi, K.; et al. Absence of hormone-sensitive lipase inhibits obesity and adipogenesis in Lepob/ob mice. J. Biol. Chem. 2004, 279, 15084–15090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, R.; Strauss, J.G.; Haemmerle, G.; Schoiswohl, G.; Birner-Gruenberger, R.; Riederer, M.; Lass, A.; Neuberger, G.; Eisenhaber, F.; Hermetter, A.; et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 2004, 306, 1383–1386. [Google Scholar] [CrossRef] [Green Version]
- Kurylowicz, A.; Owczarz, M.; Polosak, J.; Jonas, M.I.; Lisik, W.; Jonas, M.; Chmura, A.; Puzianowska-Kuznicka, M. Sirt1 and sirt7 expression in adipose tissues of obese and normal-weight individuals is regulated by microRNAs but not by methylation status. Int. J. Obes. 2016, 40, 1635–1642. [Google Scholar] [CrossRef]
- Jing, E.; Gesta, S.; Kahn, C.R. Sirt2 regulates adipocyte differentiation through FoxO1 acetylation/ deacetylation. Cell. Metab. 2007, 6, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado De Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1promotes fat mobilization in whiteadipocytes by repressing PPAR-gamma. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef]
- Puri, N.; Sodhi, K.; Haarstad, M.; Kim, D.H.; Bohinc, S.; Foglio, E.; Favero, G.; Abraham, N.G. Hemeinduced oxidative stress attenuates sirtuin1 and enhances adipogenesis inmesenchymal stem cells and mouse preadipocytes. J. Cell. Biochem. 2012, 113, 1926–1935. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, Z.; Zhang, W.; Hu, X.; Wei, H.; Peng, J.; Jiang, S. Sirt1 inhibits adipogenesis and promotes myogenic differentiation in C3H10T1/2 pluripotent cells by regulating Wnt signaling. Cell. Biosci. 2015, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Xu, S.; Maitland-Toolan, K.A.; Sato, K.; Jiang, B.; Ido, Y.; Lan, F.; Walsh, K.; Wierzbicki, K.; Verbeuren, T.J.; et al. Sirt1 regulates hepatocyte lipid metabolism through activating AMP-activated protein kinase. J. Biol. Chem. 2008, 283, 20015–20026. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.T.; Chi, K.T.; Lan, Y.W.; Chan, J.C.; Ma, Y.S.; Wei, Y.H. Depletion of Sirt3 leads to the impairment of adipogenic differentiation and insulin resistance via interfering mitochondrial function of adipose-derived human mesenchymal stem cells. Free Radic. Res. 2018, 52, 1398–1415. [Google Scholar] [CrossRef]
- Livingstone, C.; Borai, A. Insulin-like growth factor-II: Its role in metabolic and endocrine disease. Clin. Endocrinol. 2014, 80, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. Minireview: IGF, insulin, and cancer. Endocrinology 2011, 152, 2546–2551. [Google Scholar] [CrossRef] [Green Version]
- Haselbacher, G.; Humbel, R. Evidence for two species of insulin-like growth factor II (IGF II and “big” IGF II) in human spinal fluid. Endocrinology 1982, 110, 1822–1824. [Google Scholar] [CrossRef]
- Jeyaratnaganthan, N.; Hojlund, K.; Kroustrup, J.P. Circulating levels of insulin-like growth factor-II/mannose-6-phosphate receptor in obesity and type 2 diabetes. Growth Horm IGF Res. 2010, 20, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Xuan, L.; Ma, J.; Yu, M.; Yang, Z.; Huang, Y.; Guo, C.; Lu, Y.; Yan, L.; Shi, S. Insulin-like growth factor 2 promotes adipocyte proliferation, differentiation and lipid deposition in obese type 2 diabetes. J. Transl. Sci. 2019, 6, 1–7. [Google Scholar]
- Kessler, S.M.; Laggai, S.; Van Wonterg, E. Transient hepatic overexpression of insulin-like growth factor 2 induces free cholesterol and lipid droplet formation. Front. Physiol. 2016, 7, 147. [Google Scholar]
- Da Costa, T.H.; Williamson, D.H.; Ward, A.; Bates, P.; Fisher, R.; Richardson, L.; Hill, D.J.; Robinson, I.C.A.F.; Graham, C.F. High plasma insulin-like growth factor-II and low lipid content in transgenic mice: Measurements of lipid metabolism. J. Endocrinol. 1994, 143, 433–439. [Google Scholar] [CrossRef]
- Berra-Romani, R.; Guzmán-Silva, A.; Vargaz-Guadarrama, A.; Flores-Alonso, J.C.; Alonso-Romero, J.; Treviño, S.; Sánchez-Gómez, J.; Coyotl-Santiago, N.; García-Carrasco, M.; Moccia, F. Type 2 diabetes alters intracellular Ca2+ handling in native endothelium of excised rat aorta. Int. J. Mol. Sci. 2019, 21, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlugosová, K.; Weismann, P.; Bernátová, I.; Sotníková, R.; Slezák, J.; Okruhlicová, L. Omega-3 fatty acids and atorvastatin affect connexin 43 expression in the aorta of hereditary hypertriglyceridemic rats. Can. J. Physiol. Pharmacol. 2009, 87, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Shaw, S.G.; Abraham, D.J.; Baker, D.M.; Tsui, J. Cell signalling pathways leading to novel therapeutic strategies in cardiovascular disease. Cardiol. Res. Pract. 2012, 2012, 475094. [Google Scholar] [CrossRef] [Green Version]
- Turovsky, E.A.; Turovskaya, M.V.; Zinchenko, V.P.; Dynnik, V.V.; Dolgacheva, L.P. Insulin induces Ca2+ oscillations in white fat adipocytes via PI3K and PLC. Biochem. Moscow Suppl. Ser. A 2016, 10, 53–59. [Google Scholar] [CrossRef]
- Małodobra-Mazur, M.; Cierzniak, A.; Pawełka, D.; Kaliszewski, K.; Rudnicki, J.; Dobosz, T. Metabolic differences between subcutaneous and visceral adipocytes differentiated with an excess of saturated and monounsaturated fatty acids. Genes 2020, 11, 1092. [Google Scholar] [CrossRef]
- Ohlson, K.B.; Shabalina, I.G.; Lennström, K.; Backlund, E.C.; Mohell, N.; Bronnikov, G.E.; Lindahl, S.G.; Cannon, B.; Nedergaard, J. Inhibitory effects of halothane on the thermogenic pathway in brown adipocytes: Localization to adenylyl cyclase and mitochondrial fatty acid oxidation. Biochem. Pharmacol. 2004, 68, 463–477. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Braga, A.; Yu, Y.; Esteras, N.; Korsak, A.; Theparambil, S.M.; Hadjihambi, A.; Hosford, P.S.; Teschemacher, A.G.; Marina, N.; et al. Mechanosensory signalling in astrocytes. J. Neurosci. 2020, 40, 9364–9371. [Google Scholar] [CrossRef]
- Gaidin, S.G.; Turovskaya, M.V.; Mal’tseva, V.N.; Zinchenko, V.P.; Blinova, E.V.; Turovsky, E.A. A complex neuroprotective effect of alpha-2-adrenergic receptor agonists in a model of cerebral ischemia–reoxygenation in vitro. Biochem. Suppl. Ser. A 2019, 13, 319–333. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NM_001289726 GAPDH | Forward 5′-tccactcacggcaaattcaac-3′ Reverse 5′-cggcatcgaaggtggaagag-3′ |
| NM_010719 Lipe | Forward 5′-gagcactacaaacgcaacgagaca-3′ Reverse 5′-aaattcagccccacgcaactct-3′ |
| NM_019812 Sirt1 | Forward 5′-ctttcagaaccaccaaagcgga-3′ Reverse 5′-acagaaaccccagctccagtca-3′ |
| NM_022433 Sirt3 | Forward 5′-acctttgtaacagctacatgcacggt-3′ Reverse 5′-ccatcacatcagcccatatgtcttc-3′ |
| NM_010514 Igf2 | Forward 5′-cctcctggagacatactgtgccac-3′ Reverse 5′-tgtctccaggtgtcatattggaagaa-3′ |
| NM_010288 Gja1 | Forward 5′-cttcaatggctgctcctcacca-3′ Reverse 5′- gctcgctggcttgcttgttgt-3′ |
| NM_025802 Atgl | Forward 5′-tcattcctcctaccctccaag-3′ Reverse 5′-atcgaagtccatctctgtagc-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turovsky, E.A.; Varlamova, E.G.; Turovskaya, M.V. Activation of Cx43 Hemichannels Induces the Generation of Ca2+ Oscillations in White Adipocytes and Stimulates Lipolysis. Int. J. Mol. Sci. 2021, 22, 8095. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158095
Turovsky EA, Varlamova EG, Turovskaya MV. Activation of Cx43 Hemichannels Induces the Generation of Ca2+ Oscillations in White Adipocytes and Stimulates Lipolysis. International Journal of Molecular Sciences. 2021; 22(15):8095. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158095
Chicago/Turabian StyleTurovsky, Egor A., Elena G. Varlamova, and Maria V. Turovskaya. 2021. "Activation of Cx43 Hemichannels Induces the Generation of Ca2+ Oscillations in White Adipocytes and Stimulates Lipolysis" International Journal of Molecular Sciences 22, no. 15: 8095. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158095