
Proteomic Characterization of Antibiotic Resistance in Listeria and Production of Antimicrobial and Virulence Factors

 , , ,
, , ,
Abstract
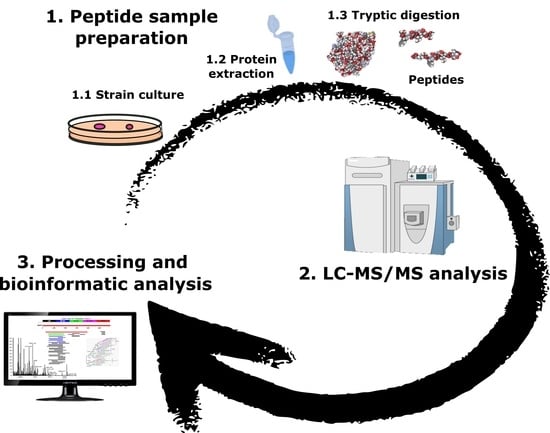
:
1. Introduction
2. Results
2.1. Listeria spp. Proteomics Data Repository
2.2. Proteins Involved in Bacterial Resistance to Antibiotics and Other Toxic Substances
2.3. Antibacterial Compounds and Proteins Involved in Antibacterial Production
2.4. Proteins Involved in Bacterial Toxicity
2.5. Proteins Involved in Host Colonization and Immune Evasion
2.6. Transporters Associated to Virulence Factors
2.7. Other Bacterial Virulence Factors
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Protein Extraction
4.3. Peptide Sample Preparation
4.4. Shotgun LC–ESI–MS/MS Analysis
4.5. LC–ESI–MS/MS Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [Green Version]
- Walsh, D.; Duffy, G.; Sheridan, J.J.; Blair, I.S.; McDowell, D.A. Antibiotic resistance among Listeria, including Listeria monocytogenes, in retail foods. J. Appl. Microbiol. 2001, 90, 517–522. [Google Scholar] [CrossRef]
- Rocha, C.E.; Mol, J.P.S.; Garcia, L.N.N.; Costa, L.F.; Santos, R.L.; Paixão, T.A. Comparative experimental infection of Listeria monocytogenes and Listeria ivanovii in bovine trophoblasts. PLoS ONE 2017, 12, e0176911. [Google Scholar] [CrossRef] [Green Version]
- Vygovska, L.M. Determination Listeria spp. (L. welshimeri, L. grayi, L. murrayi, L. innocua). J. Vet. Med. Biotechnol. Biosaf. 2018, 4, 33–36. [Google Scholar]
- Iannetti, L.; Schirone, M.; Neri, D.; Visciano, P.; Acciari, V.A.; Centorotola, G.; Mangieri, M.S.; Torresi, M.; Santarelli, G.A.; Di Marzio, V.; et al. Listeria monocytogenes in poultry: Detection and strain characterization along an integrated production chain in Italy. Food Microbiol. 2020, 91, 103533. [Google Scholar] [CrossRef]
- Arslan, S.; Baytur, S. Prevalence and antimicrobial resistance of Listeria species and subtyping and virulence factors of Listeria monocytogenes from retail meat. J. Food Saf. 2019, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of antibiotic resistance in Listeria monocytogenes isolated from food products: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [Green Version]
- Trost, M.; Wehmhöner, D.; Kärst, U.; Dieterich, G.; Wehland, J.; Jänsch, L. Comparative proteome analysis of secretory proteins from pathogenic and nonpathogenic Listeria species. Proteomics 2005, 5, 1544–1557. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Freitag, N.E. Regulation of Listeria monocytogenes Virulence. Microbiol. Spectr. 2019, 7, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Lee, S. Bacteriocins of Listeria monocytogenes and their potential as a virulence factor. Toxins 2020, 12, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, C.; Brown, L.; Maryam, M.; Vij, R.; Smith, D.F.Q.; Burnet, M.C.; Kyle, J.E.; Heyman, H.M.; Ramirez, J.; Prados-Rosales, R.; et al. Listeria monocytogenes virulence factors, including listeriolysin O, are secreted in biologically active extracellular vesicles. J. Biol. Chem. 2019, 294, 1202–1217. [Google Scholar] [CrossRef] [Green Version]
- Toledo-Arana, A.; Dussurget, O.; Nikitas, G.; Sesto, N.; Guet-Revillet, H.; Balestrino, D.; Loh, E.; Gripenland, J.; Tiensuu, T.; Vaitkevicius, K.; et al. The Listeria transcriptional landscape from saprophytism to virulence. Nature 2009, 459, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Healey, S.; Regan, P.; Laksanalamai, P.; Hu, Z. PCR-based methodologies for detection and characterization of Listeria monocytogenes and Listeria ivanovii in foods and environmental sources. Food Sci. Hum. Wellness 2017, 6, 39–59. [Google Scholar] [CrossRef]
- Soni, D.K.; Ahmad, R.; Dubey, S.K. Biosensor for the detection of Listeria monocytogenes: Emerging trends. Crit. Rev. Microbiol. 2018, 44, 590–608. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.R.; Shah, R.M.; Palombo, E.A. Maldi-tof ms: A rapid methodology for identifying and subtyping Listeria monocytogenes. Methods Mol. Biol. 2021, 2220, 17–29. [Google Scholar] [PubMed]
- Ojima-Kato, T.; Yamamoto, N.; Takahashi, H.; Tamura, H. Matrix-assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) can precisely discriminate the lineages of Listeria monocytogenes and species of Listeria. PLoS ONE 2016, 11, e0159730. [Google Scholar] [CrossRef] [Green Version]
- Varadarajan, A.R.; Goetze, S.; Pavlou, M.P.; Grosboillot, V.; Shen, Y.; Loessner, M.J.; Christian, H.A.; Wollcheid, B. A proteogenomic resource enabling integrated analysis of Listeria genotype-proteotype-phenotype relationships. J. Proteome Res. 2020, 19, 1647–1662. [Google Scholar] [CrossRef]
- Tiong, H.K.; Hartson, S.; Muriana, P.M. Comparison of five methods for direct extraction of surface proteins from Listeria monocytogenes for proteomic analysis by orbitrap mass spectrometry. J. Microbiol. Methods 2015, 110, 54–60. [Google Scholar] [CrossRef]
- Calvo, E.; Pucciarelli, M.G.; Bierne, H.; Cossart, P.; Albar, J.P.; García-Del Portillo, F. Analysis of the Listeria cell wall proteome by two-dimensional nanoliquid chromatography coupled to mass spectrometry. Proteomics 2005, 5, 433–443. [Google Scholar] [CrossRef]
- Esbelin, J.; Santos, T.; Ribière, C.; Desvaux, M.; Viala, D.; Chambon, C.; Hébraud, M. Comparison of three methods for cell surface proteome extraction of Listeria monocytogenes biofilms. OMICS J. Integr. Biol. 2018, 22, 779–787. [Google Scholar] [CrossRef] [Green Version]
- Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Rama, J.L.R.; Calo-Mata, P.; Sánchez-Pérez, A.; Villa, T.G. Proteomic characterization of antibiotic resistance, and production of antimicrobial and virulence factors in Streptococcus species associated with bovine mastitis. Could enzybiotics represent novel therapeutic agents against these pathogens? Antibiotics 2020, 9, 302. [Google Scholar] [CrossRef]
- Abril, A.G.; Ortea, I.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Cañas, B.; Villa, T.G.; Calo-Mata, P. Characterization biomarker peptides for the Identification of Bacillus species by LC-ESI-MS/MS. In Exploring Microorganisms: Recent Advances in Applied Microbiology; Mendez-Vilas, A., Ed.; Universal Publishers Inc.: Boca Raton, FL, USA, 2018; pp. 74–77. ISBN1 1-62734-623-6. ISBN2 978-1-62734-623-8. [Google Scholar]
- Carrera, M.; Böhme, K.; Gallardo, J.M.; Barros-Velázquez, J.; Cañas, B.; Calo-Mata, P. Characterization of foodborne strains of Staphylococcus aureus by shotgun proteomics: Functional networks, virulence factors and species-specific peptide biomarkers. Front. Microbiol. 2017, 8, 2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhamed, H.; Ramachandran, R.; Narayanan, L.; Ozdemir, O.; Cooper, A.; Olivier, A.K.; Karsi, A.; Lawrence, M.L. Contributions of a LysR transcriptional regulator to Listeria monocytogenes virulence and identification of its regulons. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Curtis, T.D.; Takeuchi, I.; Gram, L.; Knudsen, G.M. The influence of the Toxin/antitoxin mazEF on growth and survival of Listeria monocytogenes under stress. Toxins 2017, 9, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocourt, J.; Grimont, P.A.D. Listeria welshimeri sp. nov. and Listeria seeligeri sp. nov. Int. J. Syst. Bacteriol. 1983, 33, 866–869. [Google Scholar] [CrossRef] [Green Version]
- Dowson, C.G.; Hutchison, A.; Brannigan, J.A.; George, R.C.; Hansman, D.; Linares, J.; Tomasz, A.; Smith, J.M.; Spratt, B.G. Horizontal transfer of penicillin-binding protein genes in penicillin-resistant clinical isolates of Streptococcus pneumoniae. Proc. Natl. Acad. Sci. USA 1989, 86, 8842–8846. [Google Scholar] [CrossRef] [Green Version]
- Grove, A. Quick guide MarR family transcription factors. Curr. Biol. 2013, 23, R142–R143. [Google Scholar] [CrossRef] [Green Version]
- López-pelegrín, M.; Cerdà-costa, N.; Martínez-jiménez, F.; Cintas-pedrola, A.; Canals, A.; Peinado, J.R.; Marti-renom, M.A.; López-otín, C.; Arolas, J.L.; Gomis-rüth, F.X. A Novel family of soluble minimal scaffolds provides structural insight into the catalytic domains of integral membrane metallopeptidases. J. Biol. Chem. 2013, 288, 21279–21294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque-Sastre, L.; Arroyo, C.; Fox, E.M.; McMahon, B.J.; Bai, L.; Li, F.; Fanning, S. Antimicrobial resistance in Listeria species. In Antimicrobial Resistance in Bacteria from Livestock and Companion Animals; ASM Press: Washington, DC, USA, 2018; pp. 237–259. [Google Scholar]
- Fillgrove, K.L.; Pakhomova, S.; Schaab, M.R.; Newcomer, M.E.; Armstrong, R.N. Structure and mechanism of the genomically encoded fosfomycin resistance protein, FosX, from Listeria monocytogenes. Biochemistry 2007, 46, 8110–8120. [Google Scholar] [CrossRef] [PubMed]
- Vetting, M.W.; Luiz, L.P.; Yu, M.; Hegde, S.S.; Magnet, S.; Roderick, S.L.; Blanchard, J.S. Structure and functions of the GNAT superfamily of acetyltransferases. Arch. Biochem. Biophys. 2005, 433, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Blankenfeldt, W.; Kuzin, A.P.; Skarina, T.; Korniyenko, Y.; Tong, L.; Bayer, P.; Janning, P.; Thomashow, L.S.; Mavrodi, D.V. Structure and function of the phenazine biosynthetic protein PhzF from Pseudomonas fluorescens. Proc. Natl. Acad. Sci. USA 2004, 101, 16431–16436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbertson, L.; Nodwell, J.R. The TetR Family of Regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, M.; Depardieu, F.; Molinas, C.; Reynolds, P.; Courvalin, P. The vanZ gene of Tn1546 from Enterococcus faecium BM4147 confers resistance to teicoplanin. Gene 1995, 154, 87–92. [Google Scholar] [CrossRef]
- Matson, J.S.; Livny, J.; DiRita, V.J. A putative Vibrio cholerae two-component system controls a conserved periplasmic protein in response to the antimicrobial peptide polymyxin B. PLoS ONE 2017, 12, e0186199. [Google Scholar] [CrossRef] [Green Version]
- Iyer, L.M.; Abhiman, S.; Aravind, L. MutL homologs in restriction-modification systems and the origin of eukaryotic MORC ATPases. Biol. Direct 2008, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Suprenant, K.A.; Bloom, N.; Fang, J.; Lushington, G. The major vault protein is related to the toxic anion resistance protein (TelA) family. J. Exp. Biol. 2007, 210, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, C.N.; Harrop, S.J.; Boucher, Y.; Hassan, K.A.; Leo, R.D.; Xu, X.; Cui, H.; Savchenko, A.; Chang, C.; Labbate, M.; et al. Crystal structure of an integron gene cassette-associated protein from Vibrio cholerae identifies a cationic drug-binding module. PLoS ONE 2011, 6, e16934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, J.S.; Cooksey, D.A. Copper resistance in Pseudomonas syringae mediated by periplasmic and outer membrane proteins. Proc. Natl. Acad. Sci. USA 1991, 88, 8915–8919. [Google Scholar] [CrossRef] [Green Version]
- Hobman, J.L.; Wilkie, J.; Brown, N.L. A design for life: Prokaryotic metal-binding MerR family regulators. Biometals 2005, 18, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Domingos, R.M.; Teixeira, R.D.; Zeida, A.; Agudelo, W.A.; Alegria, T.G.P.; Da Silva Neto, J.F.; Vieira, P.S.; Murakami, M.T.; Farah, C.S.; Estrin, D.A.; et al. Substrate and product-assisted catalysis: Molecular aspects behind structural switches along organic hydroperoxide resistance protein catalytic cycle. ACS Catal. 2020, 10, 6587–6602. [Google Scholar] [CrossRef]
- Son, M.S.; Del Castilho, C.; Duncalf, K.A.; Carney, D.; Weiner, J.H.; Turner, R.J. Mutagenesis of SugE, a small multidrug resistance protein. Biochem. Biophys. Res. Commun. 2003, 312, 914–921. [Google Scholar] [CrossRef]
- Haubert, L.; Kremer, F.S.; da Silva, W.P. Whole-genome sequencing identification of a multidrug-resistant Listeria monocytogenes serotype 1/2a isolated from fresh mixed sausage in southern Brazil. Infect. Genet. Evol. 2018, 65, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Chastanet, A.; Derre, I.; Nair, S.; Msadek, T. clpB, a Novel Member of the Listeria monocytogenes CtsR regulon, is involved in virulence but not in general stress tolerance. J. Bacteriol. 2004, 186, 1165–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Delumeau, O.; Dutta, S.; Brigulla, M.; Kuhnke, G.; Hardwick, S.W.; Völker, U.; Yudkin, M.D.; Lewis, R.J. Functional and structural characterization of RsbU, a stress signaling protein phosphatase 2C. J. Biol. Chem. 2004, 279, 40927–40937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubbs, J.M.; Mongkolsuk, S. Peroxiredoxins in Bacterial Antioxidant Defense. In Subcellular Biochemistry; Springer: New York, NY, USA, 2007; Volume 44, pp. 143–193. [Google Scholar]
- Park, S.C.; Kwak, Y.M.; Song, W.S.; Hong, M.; Yoon, S. Il Structural basis of effector and operator recognition by the phenolic acid-responsive transcriptional regulator PadR. Nucleic Acids Res. 2017, 45, 13080–13093. [Google Scholar] [CrossRef] [Green Version]
- Rolhion, N.; Chassaing, B.; Nahori, M.-A.; de Bodt, J.; Moura, A.; Lecuit, M.; Dussurget, O.; Bérard, M.; Marzorati, M.; Fehlner-Peach, H.; et al. Specific targeting of intestinal Prevotella copri by a Listeria monocytogenes bacteriocin. bioRxiv 2019, 680801. [Google Scholar] [CrossRef] [Green Version]
- Mutschler, H.; Gebhardt, M.; Shoeman, R.L.; Meinhart, A. A Novel mechanism of programmed cell death in bacteria by toxin–antitoxin systems corrupts peptidoglycan synthesis. PLoS Biol. 2011, 9, e1001033. [Google Scholar] [CrossRef]
- Jørgensen, M.G.; Pandey, D.P.; Jaskolska, M.; Gerdes, K. HicA of Escherichia coli defines a novel family of translation-independent mRNA interferases in bacteria and archaea. J. Bacteriol. 2009, 191, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Grishin, N.V.; Koonin, E.V. The HicAB cassette, a putative novel, RNA-targeting toxin-antitoxin system in archaea and bacteria. Bioinformatics 2006, 22, 2581–2584. [Google Scholar] [CrossRef]
- Lima-Mendez, G.; Alvarenga, D.O.; Ross, K.; Hallet, B.; Van Melderen, L.; Varani, A.M.; Chandler, M. Toxin-antitoxin gene pairs found in Tn3 family transposons appear to be an integral part of the transposition module. MBio 2020, 11, e00452-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepanowski, R.; Braun, S.; Riedel, V.; Schneiker, S.; Krahn, I.; Pühler, A.; Schlüter, A. The 120 592 bp IncF plasmid pRSB107 isolated from a sewage-treatment plant encodes nine different antiobiotic-resistance determinants, two iron-acquisition systems and other putative virulence-associated functions. Microbiology 2005, 151, 1095–1111. [Google Scholar] [CrossRef] [Green Version]
- Santos, T.; Viala, D.; Chambon, C.; Esbelin, J.; Hébraud, M. Listeria monocytogenes biofilm adaptation to different temperatures seen through shotgun proteomics. Front. Nutr. 2019, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Bonazzi, M.; Lecuit, M.; Cossart, P. Listeria monocytogenes internalin and E-cadherin: From bench to bedside. Cold Spring Harb. Perspect. Biol. 2009, 1, a0030807. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, K.K.; Pier, D.B.; Goldmann, D.A.; Pier, G.B. The Teicoplanin-Associated Locus Regulator (TcaR) and the intercellular adhesin locus regulator (IcaR) are transcriptional inhibitors of the ica locus in Staphylococcus aureus. J. Bacteriol. 2004, 186, 2449–2456. [Google Scholar] [CrossRef] [Green Version]
- Molloy, S. Bacterial pathogenicity: A competent escape for Listeria. Nat. Rev. Microbiol. 2012, 10, 670. [Google Scholar] [CrossRef]
- Schubert, K.; Bichlmaier, A.M.; Mager, E.; Wolff, K.; Ruhland, G.; Fiedler, F. P45, an extracellular 45 kDa protein of Listeria monocytogenes with similarity to protein p60 and exhibiting peptidoglycan lyric activity. Arch. Microbiol. 2000, 173, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Jacobitz, A.W.; Kattke, M.D.; Wereszczynski, J.; Clubb, R.T. Sortase Transpeptidases: Structural biology and catalytic mechanism. In Advances in Protein Chemistry and Structural Biology; Academic Press Inc.: Cambridge, MA, USA, 2017; Volume 109, pp. 223–264. [Google Scholar]
- Herzberg, M.; Kaye, I.K.; Peti, W.; Wood, T.K. YdgG (TqsA) controls biofilm formation in Escherichia coli K-12 through autoinducer 2 transport. J. Bacteriol. 2006, 188, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Vollmer, W.; Tomasz, A. Peptidoglycan N-Acetylglucosamine deacetylase, a putative virulence factor in Streptococcus pneumoniae. Infect. Immun. 2002, 70, 7176–7178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.; Ogunniyi, A.D.; Choi, M.; Pyo, S.; Rhee, D.; Paton, J.C. The ClpP Protease of Streptococcus pneumoniae modulates virulence gene expression and protects against fatal Pneumococcal challenge. Infect. Immun. 2004, 72, 5646–5653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajfasz, J.K.; Martinez, A.R.; Rivera-ramos, I.; Abranches, J.; Koo, H.; Quivey, R.G. Role of Clp proteins in expression of virulence properties of Streptococcus mutans. J. Bacteriol. 2009, 191, 2060–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.W.; Metzger, J.F. Demonstration of a capsular structure on Listeria monocytogenes. Pathol. Microbiol. 1962, 25, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.R.; Stevens, R.W. Fine structure of Listeria monocytogenes. J. Bacteriol. 1963, 86, 414–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijek, T.M.; Mou, S.; Bachert, B.A.; Kuehl, K.A.; Williams, J.A.; Daye, S.P.; Worsham, P.L.; Bozue, J.A. The D-alanyl-D-alanine carboxypeptidase enzyme is essential for virulence in the Schu S4 strain of Francisella tularensis and a dacD mutant is able to provide protection against a pneumonic challenge. Microb. Pathog. 2019, 137, 103742. [Google Scholar] [CrossRef] [PubMed]
- Tomazi, T.; de Souza Filho, A.F.; Heinemann, M.B.; dos Santos, M.V. Molecular characterization and antimicrobial susceptibility pattern of Streptococcus agalactiae isolated from clinical mastitis in dairy cattle. PLoS ONE 2018, 13, e0199561. [Google Scholar] [CrossRef]
- Schmelcher, M.; Waldherr, F.; Loessner, M.J. Listeria bacteriophage peptidoglycan hydrolases feature high thermoresistance and reveal increased activity after divalent metal cation substitution. Appl. Microbiol. Biotechnol. 2012, 93, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Danevčič, T.; Borić Vezjak, M.; Tabor, M.; Zorec, M.; Stopar, D. Prodigiosin induces autolysins in actively grown Bacillus subtilis Cells. Front. Microbiol. 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A.; Bycroft, M. The structure of a LysM domain from E. coli membrane-bound lytic murein transglycosylase D (MltD). J. Mol. Biol. 2000, 299, 1113–1119. [Google Scholar] [CrossRef]
- McGowan, S.; Lucet, I.S.; Cheung, J.K.; Awad, M.M.; Whisstock, J.C.; Rood, J.I. The FxRxHrS motif: A conserved region essential for DNA binding of the VirR response regulator from Clostridium perfringens. J. Mol. Biol. 2002, 322, 997–1011. [Google Scholar] [CrossRef]
- Broxton, C.N.; Culotta, V.C. SOD Enzymes and microbial pathogens: Surviving the oxidative storm of infection. PLoS Pathog. 2016, 12, e1005295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribardo, D.A.; Lambert, T.J.; Mciver, K.S. Role of Streptococcus pyogenes Two-Component response regulators in the temporal control of Mga and the Mga-Regulated virulence gene emm. Infect. Immun. 2004, 72, 3668–3673. [Google Scholar] [CrossRef] [Green Version]
- Warne, B.; Harkins, C.P.; Harris, S.R.; Vatsiou, A.; Stanley-Wall, N.; Parkhill, J.; Peacock, S.J.; Palmer, T.; Holden, M.T.G. The Ess/Type VII secretion system of Staphylococcus aureus shows unexpected genetic diversity. BMC Genom. 2016, 17, 222. [Google Scholar] [CrossRef] [Green Version]
- Schulein, R.; Dehio, C. The VirB/VirD4 type IV secretion system of Bartonella is essential for establishing intraerythrocytic infection. Mol. Microbiol. 2002, 46, 1053–1067. [Google Scholar] [CrossRef]
- Pallen, M.J. The ESAT-6/WXG100 superfamily and a new Gram-positive secretion system? Trends Microbiol. 2002, 10, 209–212. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Wu, H.; Li, N.; Jiang, J.; Guo, Y.; Feng, Y.; Xiao, L. Characterization of a species-specific insulinase-like protease in Cryptosporidium parvum. Front. Microbiol. 2019, 10, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumrall, E.T.; Keller, A.P.; Shen, Y.; Loessner, M.J. Structure and function of Listeria teichoic acids and their implications. Mol. Microbiol. 2020, 113, 627–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.S.; Sim, J.R.; Yun, C.H.; Han, S.H. Lipoteichoic acids as a major virulence factor causing inflammatory responses via Toll-like receptor 2. Arch. Pharm. Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Lan, Z.; Fiedler, F.; Kathariou, S. A sheep in wolf’s clothing: Listeria innocua strains with teichoic acid-associated surface antigens and genes characteristic of Listeria monocytogenes serogroup 4. J. Bacteriol. 2000, 182, 6161–6168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murazumi, N.; Kumita, K.; Yoshio, A.; Ito, E. Partial purification and properties of UDP-N-Acetylmannosamine:N-Acetylgiucosaminyl Pyrophosphorylundecaprenol N-Acetylmannosaminyltransferase from Bacillus subtilis. J. Biochem. 1988, 104, 980–984. [Google Scholar] [CrossRef]
- Shen, Y.; Boulos, S.; Sumrall, E.; Gerber, B.; Julian-Rodero, A.; Eugster, M.R.; Fieseler, L.; Nyström, L.; Ebert, M.O.; Loessner, M.J. Structural and functional diversity in Listeria cell wall teichoic acids. J. Biol. Chem. 2017, 292, 17832–17844. [Google Scholar] [CrossRef] [Green Version]
- Loughman, J.A.; Caparon, M.G. Contribution of invariant residues to the function of Rgg family transcription regulators. J. Bacteriol. 2007, 189, 650–655. [Google Scholar] [CrossRef] [Green Version]
- Price, C.T.D.; Bukka, A.; Cynamon, M.; Graham, J.E.; Al, P.E.T.; Acteriol, J.B. Glycine Betaine uptake by the ProXVWZ ABC transporter contributes to the ability of Mycobacterium tuberculosis to initiate growth in human macrophages. J. Bacteriol. 2008, 190, 3955–3961. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.J.; Song, S.; Mason, K.; Pinkett, H.W. Selective substrate uptake: The role of ATP-binding cassette (ABC) importers in pathogenesis. BBA Biomembr. 2018, 1860, 868–877. [Google Scholar] [CrossRef]
- Naka, H.; Liu, M.; Crosa, J.H. Two ABC transporter systems participate in siderophore transport in the marine pathogen Vibrio anguillarum 775 (pJM1). FEMS Microbiol. Lett. 2013, 341, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Orelle, C.; Mathieu, K.; Jault, J.M. Multidrug ABC transporters in bacteria. Res. Microbiol. 2019, 170, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Greene, N.P.; Kaplan, E.; Crow, A.; Koronakis, V. Antibiotic resistance mediated by the MacB ABC transporter family: A structural and functional perspective. Front. Microbiol. 2018, 9, 950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Mukherjee, M.M.; Varela, M.F. Modulation of bacterial multidrug resistance efflux pumps of the major facilitator superfamily. Int. J. Bacteriol. 2013, 2013, 204141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimyo, M.; Hongo, E.; Hama-inaba, H.; Machida, I. Cloning and characterization of the mvrC gene of Escherichia coli K-12 which confers resistance against methyl viologen toxicity. Nucleic Acids Res. 1992, 20, 3159–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omote, H.; Hiasa, M.; Matsumoto, T.; Otsuka, M.; Moriyama, Y. The MATE proteins as fundamental transporters of metabolic and xenobiotic organic cations. Trends Pharmacol. Sci. 2006, 27, 587–593. [Google Scholar] [CrossRef]
- Hernandez-Mendoza, A.; Quinto, C.; Segovia, L.; Perez-Rueda, E. Ligand-binding prediction in the resistance-nodulation-cell division (RND) proteins. Comput. Biol. Chem. 2007, 31, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Craig, N.L.; Chandler, M.; Gellert, M.; Lambowitz, A.M.; Rice, P.A.; Sandmeyer, S.B. Mobile DNA III, 3rd ed.; John Wiley & Sons: Washington, DC, USA, 2020. [Google Scholar]
- Robinson, K.M.; Dunning Hotopp, J.C. Mobile elements and viral integrations prompt considerations for bacterial DNA integration as a novel carcinogen. Cancer Lett. 2014, 352, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yu, Y.; Tao, F.; Zhang, J.; Copetti, D.; Kudrna, D.; Talag, J.; Lee, S.; Wing, R.A.; Fan, C. DNA methylation changes facilitated evolution of genes derived from Mutator-like transposable elements. Genome Biol. 2016, 17, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, X.; Osmundson, T.; Shi, L.; Yan, H. Comparative Genomic Analysis of a Multidrug-Resistant Listeria monocytogenes ST477 Isolate. Foodborne Pathog. Dis. 2019, 16, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Koudelka, Š.; Gelbíčová, T.; Procházková, M.; Karpíšková, R. Lineage and serotype identification of Listeria monocytogenes by Matrix-assisted Laser Desorption Ionization-time of Flight Mass Spectrometry. Czech J. Food Sci. 2018, 36, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Promadej, N.; Kim, J.W.; Kathariou, S. Teichoic acid glycosylation mediated by gtcA is required for phage adsorption and susceptibility of Listeria monocytogenes serotype 4b. Appl. Environ. Microbiol. 2008, 74, 1653–1655. [Google Scholar] [CrossRef] [Green Version]
- Matereke, L.T.; Okoh, A.I. Listeria monocytogenes virulence, antimicrobial resistance and environmental persistence: A review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef]
- Dams, D.; Briers, Y. Enzybiotics: Enzyme-based antibacterials as therapeutics. Adv. Exp. Med. Biol. 2019, 1148, 233–253. [Google Scholar] [PubMed]
- Schmitz-Esser, S.; Anast, J.M.; Cortes, B.W. A large-scale sequencing-based survey of plasmids in Listeria monocytogenes reveals global dissemination of plasmids. Front. Microbiol. 2021, 12, 510. [Google Scholar] [CrossRef]
- Li, L.; Olsen, R.H.; Shi, L.; Ye, L.; He, J.; Meng, H. Characterization of a plasmid carrying cat, ermB and tetS genes in a foodborne Listeria monocytogenes strain and uptake of the plasmid by cariogenic Streptococcus mutans. Int. J. Food Microbiol. 2016, 238, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Nie, Q.; Wang, W.; Shi, L.; Yan, H. Characterization of a transferable bcrABC and cadAC genes-harboring plasmid in Listeria monocytogenes strain isolated from food products of animal origin. Int. J. Food Microbiol. 2016, 217, 117–122. [Google Scholar] [CrossRef]
- Yan, H.; Yu, R.; Li, D.; Shi, L.; Schwarz, S.; Yao, H.; Li, X.; Du, X. A novel multiresistance gene cluster located on a plasmid-borne transposon in Listeria monocytogenes. J. Antimicrob. Chemother. 2020, 75, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Böhme, K.; Fernández-No, I.C.; Barros-Velázquez, J.; Gallardo, J.M.; Cañas, B.; Calo-Mata, P. Rapid species identification of seafood spoilage and pathogenic Gram-positive bacteria by MALDI-TOF mass fingerprinting. Electrophoresis 2011, 32, 2951–2965. [Google Scholar] [CrossRef] [PubMed]
- Böhme, K.; Fernández-No, I.C.; Barros-Velázquez, J.; Gallardo, J.M.; Calo-Mata, P.; Cañas, B. Species differentiation of seafood spoilage and pathogenic gram-negative bacteria by MALDI-TOF mass fingerprinting. J. Proteome Res. 2010, 9, 3169–3183. [Google Scholar] [CrossRef]
- Carrera, M.; Cañas, B.; Gallardo, J.M. The sarcoplasmic fish proteome: Pathways, metabolic networks and potential bioactive peptides for nutritional inferences. J. Proteomics 2013, 78, 211–220. [Google Scholar] [CrossRef]
- Calo-mata, P.; Carrera, M.; Böhme, K.; Caamaño-Antelo, S.; Gallardo, J.M.; Barros-Velázquez, J.; Cañas, B. Novel peptide biomarker discovery for detection and identification of bacterial pathogens by LC-ESI-MS/MS. J. Anal. Bioanal. Tech. 2016, 7, 296. [Google Scholar]
- Käll, L.; Canterbury, J.D.; Weston, J.; Noble, W.S.; MacCoss, M.J. Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat. Methods 2007, 4, 923–925. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Sample | Species | Strain | Culture Collection Number | Source | Pathogenicity | Variety | NCBI Accession Number | Total Virulence Factor Peptides |
|---|---|---|---|---|---|---|---|---|
| Li1 | Listeria welshimeri | ATCC 35897 | CECT 919 | Decaying vegetation | Group 1 | Serovar 6a | NZ_LT906444.1 | 17 |
| Li2 | Listeria seeligeri | ATCC 35967 | CECT 917 | Soil | Avirulent Group 1 | Serovar 1/2b | DQ065845 NC_013891.1 | 116 |
| Li3 | Listeria innocua | ATCC 33090 | CECT 910 | Cow brain | Group 1 | Serovar 6a | X98527 | 55 |
| Li4 | Listeria monocytogenes | LIS FBUNI | USC | Dairy product | - | - | - | 73 |
| Li5 | Listeria monocytogenes | ATCC 15313 | CECT 4031 | Rabbit | Group 2 | Serovar 1/2 | AJ515512 NC_003210.1 | 15 |
| Li6 | Listeria monocytogenes | ATCC 19114 | CECT 934 | Brain of sheep with circling disease | Group 2 | Serovar 4a | JF967620.1 | 18 |
| Li7 | Listeria monocytogenes | ATCC 13932 | CECT 935 | Spinal fluid of child with meningitis | Group 2 | Serovar 4b | JF967617.1 | 33 |
| Li8 | Listeria monocytogenes | NCTC 11994 | CECT 4032 | Associated with case of meningitis after eating soft cheese | Group 2 | Serovar 4b | AJ508749.1 | 37 |
| Li9 | Listeria ivanovii | ATCC 19119 | CECT 913 | Sheep | Group 2 | Serovar 5 | X98528 NZ_CP009577.1 | 31 |
| Function | Protein |
|---|---|
| Antibiotic resistance | Beta-lactamase |
| Metallo-beta-lactamase | |
| PbpX beta-lactamase-like superfamily | |
| Bleomycin resistance protein | |
| Aminoglycoside N(3)-acetyltransferase | |
| Glyoxalase/bleomycin resistance protein/dioxygenase | |
| MarR family transcriptional regulator | |
| Peptidase M56 | |
| FosX/FosE/FosI family fosfomycin resistance thiol transferase | |
| GNAT family acetyltransferase | |
| PhzF family phenazine biosynthesis protein | |
| TetR | |
| VanZ family protein O | |
| YdeI (YjbR/CyaY-like superfamily) | |
| Tetronasin resistance transmembrane protein | |
| Penicillin-binding protein | |
| Additional resistances and tolerances | AIPR protein |
| Chaperone protein ClpB | |
| Chaperone protein DnaJ | |
| Tellurite resistance protein TelA | |
| Cass2 domain-containing protein | |
| Copper resistance protein | |
| MerR family transcriptional regulator | |
| Organic hydroperoxide resistance protein | |
| Two-component sensor histidine kinase response to antimicrobials | |
| Quaternary ammonium compound-resistance protein sugE | |
| Quinolone resistance protein norB | |
| Heat shock protein 70 | |
| Hsp20/alpha crystallin family protein | |
| General stress protein | |
| OsmC family protein | |
| PadR family transcriptional regulator | |
| RsbU protein | |
| Universal stress protein | |
| Antimicrobial compounds production | Lactococcin 972 family bacteriocin |
| Antibiotic biosynthesis monooxygenase | |
| Toxins | Toxin zeta |
| LXG domain-containing protein | |
| HicA toxin | |
| Type II toxin-antitoxin system RelE/ParE family toxin | |
| Type II toxin-antitoxin system PemK/MazF family toxin | |
| Host colonization and immune evasion | Autoinducer 2 transporter |
| Viral enhancin protein | |
| Invasion associated secreted endopeptidase | |
| Peptidase_C1B | |
| Aminopeptidase ysdC | |
| Beta-Ala-Xaa dipeptidase | |
| Peptidase M23 | |
| Peptidase M16 | |
| Peptidase M23 | |
| M23 family metallopeptidase | |
| Peptidoglycan DD-metalloendopeptidase family protein M22 | |
| Peptidoglycan DD-metalloendopeptidase family protein M23 | |
| Peptidase_M22 | |
| M42 family peptidase | |
| Enhancing factor (Viral) Peptidase M60 | |
| Peptidase M60 | |
| Peptidase T | |
| Peptidase T | |
| Peptidase_M75 | |
| Peptidase C60 sortase | |
| Membrane protein with peptidase activity M56 | |
| Tripeptide aminopeptidase M20 | |
| Peptidase SA1530 | |
| Peptidase SA1531 | |
| NLP/P60 family domain | |
| Invasion-associated endopeptidase p60 | |
| Peptidoglycan endopeptidase P60 | |
| Peptidase P60 | |
| Competence protein ComEC/Rec2 | |
| Protein ComGA | |
| Peptidoglycan O-acetyltransferase | |
| ClpX protease | |
| ClpP protease | |
| ClpY protease | |
| Internalin A | |
| Internalin | |
| Internalin | |
| Internalin-J | |
| Class 1 internalin InlJ | |
| Class 3 internalin InlC | |
| Internalin B | |
| Capsular polysaccharide synthesis enzyme | |
| Capsular biosynthesis protein | |
| Capsular exopolysaccharide family protein | |
| 1-phosphatidylinositol- specific phospholipase C (PI-PLC) | |
| Chemotaxis protein CheA | |
| Cna B-type domain-containing protein | |
| N-acetylmuramoyl-L-alanine amidase | |
| D-alanyl-D-alanine carboxypeptidase | |
| Muramidase-2; Autolysin | |
| LysM domain-containing protein | |
| LysR family transcriptional regulator | |
| Autolysin LytC | |
| Lysis protein | |
| Superoxide dismutase | |
| Type VI secretion protein | |
| Type VII secretion protein EssA | |
| Type VII secretion protein EssB | |
| Putative type IV secretion system VirB4 component | |
| Mga protein | |
| Collagen-binding protein (adhesin) | |
| Adhesion lipoprotein | |
| Adhesin | |
| Intercellular adhesion protein R | |
| ESAT-6-like protein | |
| Insulinase | |
| Rgg family transcriptional regulator | |
| Protein DltD D-alanyl-lipoteichoic acid biosynthesis protein DltD | |
| Putative glycosyl/glycerophosphate transferases involved in teichoic acid biosynthesis | |
| Wall teichoic acid glycosylation protein GtcA | |
| Teichoic acid biosynthesis domain-containing protein | |
| Teichoic acid/polysaccharide export protein | |
| ABC transporters | Fluoroquinolones export ATP-binding protein Rv2688c/MT2762 |
| Antibiotic ABC transporter ATP-binding protein | |
| Petrobactin ABC transporter substrate-binding protein YclQ | |
| Bacitracin ABC transporter BceA | |
| Antibiotic ABC_2_transport permease | |
| Multidrug ABC export ATP-binding/permease protein | |
| Multidrug ABC transporter permease/ATP-binding protein | |
| Multidrug resistance ABC transporter ATP-binding/permease protein YheI | |
| Multidrug ABC transporter ATP-binding protein | |
| Spermidine/putrescine ABC Transporter PotA | |
| Siderophore ABC transporter | |
| ABC transporter ATP-binding protein | |
| ABC transporter domain-containing protein | |
| ABC transporter permease | |
| ABC transporter substrate-binding protein | |
| ABC transporter permease | |
| ABC-2 family transporter protein | |
| Peptide ABC transporter | |
| Oligopeptide ABC transporter binding protein AppA | |
| ABC-type methionine transporter MetN | |
| Amino acid ABC transporter | |
| Di-tripeptide-proton ABC symporter | |
| Glutamine ABC transporter ATP-binding protein | |
| Glycine/betaine ABC transporter | |
| Amino acid ABC transporter ATP-binding protein | |
| Iron compound ABC transporter permease | |
| Macrolide ABC transporter | |
| Manganese ABC transporter | |
| Metal ABC transporter ATP-binding protein | |
| Zinc ABC transporter substrate-binding protein AdcA | |
| Putative high-affinity zinc ABC transporter (Zn(II)-binding lipoprotein) | |
| Pheromone ABC transporter | |
| Thiol reductant ABC exporter subunit CydC | |
| Other transporters | Arsenic transporter |
| Arsenical pump-driving ATPase | |
| Cadmium, zinc, and cobalt-transporting ATPase | |
| Chloramphenicol/florfenicol efflux MFS transporter | |
| EmrB multiple drug resistance efflux pump MFS transporter | |
| Multidrug efflux SMR transporter | |
| Multidrug export protein mepA | |
| Multidrug efflux MFS transporter | |
| RND superfamily drug exporter | |
| Putrescine importer PuuP | |
| Integrase | |
| Recombinase | |
| IS21 family transposase | |
| IS3 family transposase | |
| IS30 family transposase | |
| Transposase B of Tn554 | |
| Transposase B of Tn555 | |
| Tn3 family transposase | |
| IS200/IS605 family transposase | |
| IS30 family transposase | |
| Transposase IS116/IS110/IS902 family protein | |
| Mutator family transposase | |
| Plasmid recombination enzyme | |
| Plasmid pRiA4b ORF-3 family protein | |
| Plasmid pRiA4b ORF-3 family protein | |
| Plasmid mobilization relaxosome protein MobC | |
| Bacillus transposase protein | |
| Phage infection protein, YhgE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abril, A.G.; Carrera, M.; Böhme, K.; Barros-Velázquez, J.; Calo-Mata, P.; Sánchez-Pérez, A.; Villa, T.G. Proteomic Characterization of Antibiotic Resistance in Listeria and Production of Antimicrobial and Virulence Factors. Int. J. Mol. Sci. 2021, 22, 8141. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158141
Abril AG, Carrera M, Böhme K, Barros-Velázquez J, Calo-Mata P, Sánchez-Pérez A, Villa TG. Proteomic Characterization of Antibiotic Resistance in Listeria and Production of Antimicrobial and Virulence Factors. International Journal of Molecular Sciences. 2021; 22(15):8141. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158141
Chicago/Turabian StyleAbril, Ana G., Mónica Carrera, Karola Böhme, Jorge Barros-Velázquez, Pilar Calo-Mata, Angeles Sánchez-Pérez, and Tomás G. Villa. 2021. "Proteomic Characterization of Antibiotic Resistance in Listeria and Production of Antimicrobial and Virulence Factors" International Journal of Molecular Sciences 22, no. 15: 8141. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158141