
Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals

Department of Animal Sciences, Faculty of Life and Environmental Science, Teikyo University of Science, Uenohara 409-0193, Yamanashi, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(15), 8311; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158311
Submission received: 2 July 2021
/
Revised: 28 July 2021
/
Accepted: 29 July 2021
/
Published: 2 August 2021
(This article belongs to the Special Issue Effects of Hormones on the Nervous System and Behavior)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In mammalian reproduction, sexually active males seek female conspecifics, while estrous females try to approach males. This sex-specific response tendency is called sexual preference. In small rodents, sexual preference cues are mainly chemosensory signals, including pheromones. In this article, we review the physiological mechanisms involved in sexual preference for opposite-sex chemosensory signals in well-studied laboratory rodents, mice, rats, and hamsters of both sexes, especially an overview of peripheral sensory receptors, and hormonal and central regulation. In the hormonal regulation section, we discuss potential rodent brain bisexuality, as it includes neural substrates controlling both masculine and feminine sexual preferences, i.e., masculine preference for female odors and the opposite. In the central regulation section, we show the substantial circuit regulating sexual preference and also the influence of sexual experience that innate attractants activate in the brain reward system to establish the learned attractant. Finally, we review the regulation of sexual preference by neuropeptides, oxytocin, vasopressin, and kisspeptin. Through this review, we clarified the contradictions and deficiencies in our current knowledge on the neuroendocrine regulation of sexual preference and sought to present problems requiring further study.
1. Introduction
While short-lived microbes have high mutation rates to obtain evolutional benefits against environmental pressure, long-life organisms, such as mammals, use diversity. This intraspecific diversity is achieved by sexual reproduction to disperse genes within conspecifics. To enable sexual reproduction, organisms have sexually differentiated their reproductive roles, resulting in the requirement of interaction with the opposite sex. To successfully breed offspring, animals, particularly small mammals, have evolved systems to find, attract, and approach the opposite sex, by chemosensory signals called pheromones.
The first attempts of systematic behavioral testing for opposite-sex selection or preference in laboratory rodents were reported by Slob et al. [1,2,3,4,5]. To determine the conspecifics an individual is attracted to, while simultaneously presenting various types of stimulus animals, it is possible to measure which animals the test subjects try to approach, investigate, and make contact with. The paired stimuli for simultaneous presentation would be opposing, such as estrous females vs. sexually active males, estrous females vs. anestrous females, and sexually active males vs. castrated males. In early studies, free moving rats were tested in a three-chamber apparatus with side chambers containing male and female stimulus rats unable to move beyond their chamber. In this context, male rats showed an opposite-sex preference by spending longer time in the chamber with estrous females than in that of males (same-sex) [4]. For female rats, however, it was more complicated. Insofar as males can copulate, receptive females prefer sexually inactive males rather than active ones [2], but when direct interactions were blocked by a wire-mesh barrier between them [1] or intromissions were prevented by vaginal occlusion [2], the receptive females showed a clear preference to spend longer time in the vicinity of sexually active male rats.
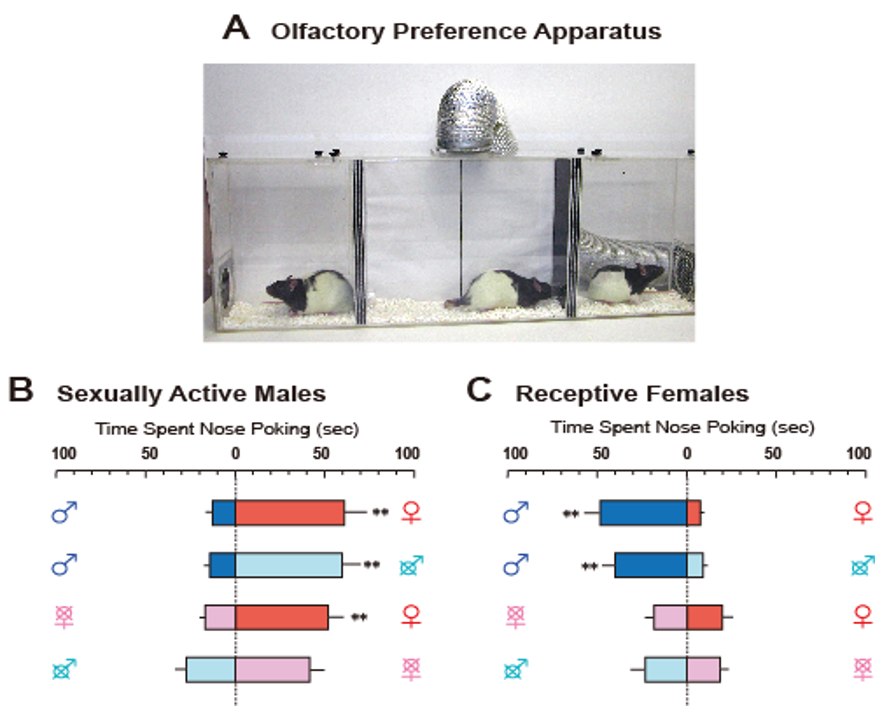
Therefore, sexual partner preferences, which appear to be simple phenomena, have been found to be influenced by many factors, and do not respond to a simple model of sexually dimorphic brain function. Therefore, we sought to develop a preference test for opposite-sex odors limited to airborne chemicals, components of sexual stimuli [6]. Figure 1A depicts our preference test apparatus in rats. The apparatus was divided into three chambers by spaced triplicate boards with a hole at different levels that enabled airflow. A fan connected to the ceiling of the center chamber helped move the airflow carrying the odors from the stimulus animals in the side chambers to subject rats in the center one. Odors led to subjects poking their noses into the air inlet to investigate them (for easier observation of nose-poking behavior, a transparent tube was attached to the inlets). This apparatus allowed us to obtain clear data on sex-specific odor preference patterns (Figure 1B,C). In this paper, we review recent findings on neural and hormonal preference regulation for conspecific odors in laboratory rodents, mice, rats, and hamsters.
2. Detection of Sexual Chemicals
The rodent nasal cavity has mainly two chemosensory sensory organs, the vomeronasal organ (VNO) and the olfactory epithelium (OE). However, recently, another chemosensory receptor in the end of the mouse nasal cavity, the Grueneberg ganglion, has been reported. It detects alarm pheromones and predator odors [7,8,9]. The VNO hitherto had been thought to be a chemical detector specialized for pheromones, and the OE a detector of airborne chemicals, handling the role of general olfaction. Furthermore, these receptors’ ligands have almost no overlap; olfactory neurons in the OE express a single type of G-protein-coupled receptor (GPCR), that is, detecting a single ligand, whereas some vomeronasal neurons in the VNO express multiple GPCR types, detecting multiple ligands [10].
Vomeronasal neurons express two classes of receptors: the V1R coupled with Gi2α and the V2R coupled with Goα proteins. In the mouse, vomeronasal sensory epithelium, neurons with V1R in the apical layer project their axons to the rostral part of the accessory olfactory bulb (AOB) and detect small, volatile molecules, while V2R, in the basal layer, project to the caudal AOB region and detect larger, nonvolatile molecules such as peptides and proteins. It has been reported that the use of these vomeronasal receptors differs between males and females [11]. Exposure to soiled bedding collected from sexually active males activates neurons with V2R in the basal layer in estrogen-primed female mice, whereas exposure to estrous female soiled bedding activates neurons with both V1R and V2R in both apical and basal layers in males, but not in estrogen-primed females. Thus, sex differences in V1R and V2R reactivity to the same social chemicals in the peripheral VNO may exist.
Sulfated estrogens are detected as pheromones by V1rj, a V1R subclass, and other unknown molecules in female mouse urine are detected by the V1re and V1rj subclasses, although each one alone has no effect on male mouse behavior. When combined, however, they strongly induce male courtship behavior in mice [12]. On the other hand, darcin (also known as a major urine protein, MUP20), a V2R ligand contained in male mouse urine that provokes aggressive behavior in male mice [13,14], can strongly attract receptive females to the intact male urine [15,16]. In addition, darcin also acts as a reinforcer; that is, when associated with airborne neutral odors in the urine, it produces the learned attraction to male odors in receptive females [15,16]. Exocrine gland-secreting peptide 1 (ESP1) has also been found in the male lacrimal gland as a V2R ligand [17]. Although it has been reported that contact with male mouse pheromone ESP1 enhanced lordosis behavior in receptive females [17], it is still uncertain whether this nonvolatile chemical contributes to the male sexual attraction of receptive female mice. Interestingly, proteome analysis in wild mice (Mus musculus) showed darcin and ESP1 also in saliva [18].
Though olfactory neurons express >900 canonical olfactory receptors in the mouse OE [19], trace amino-associated receptors (TAARs) have received attention as noncanonical olfactory receptors for pheromonal signals [20,21]. All TAARs found in 15 mouse genes and 17 genes in the rat function as olfactory receptors, except for TAAR1 [22]. Deficiency of TAARs 2–9 in male mice has been reported to abolish approach behavior toward receptive females [23], while exposure to isobutylamine, a volatile urinary amine considered to be a TAAR3 ligand, increased the activity in the brain regions related to male sexual behavior in wild type (WT) but not in TAARs 2–9 knock-out (KO) mice [23]. Trimethylamine, a TAAR5 ligand, abundantly contained in male mouse urine (>1000 times that of rat urine), works as an incentive for receptive female mice (but as a repellent for rats). TAAR5-KO females do not approach male mouse urine or trimethylamine [24]. All olfactory neurons expressing TAARs in the OE project their axons to specific (necklace) glomeruli surrounding the main olfactory bulb (MOB) [25,26] specialized in processing biologically significant signals, such as social cues, edible and rotten foods, and predator odors [27].
In addition, some epithelial neurons with canonical olfactory receptors may detect pheromones or other social signals. In fact, epithelial neurons express canonical olfactory receptors and transient receptor potential C2 (TRPc2), similar to VNO sensory neurons with V1R or V2R [28]. At present, the function of these neurons is unknown. What types of stimuli these neurons are sensing, and how they are involved in the perception of opposite-sex odors, are questions that warrant further research.
3. Sexual Stimuli Attributes
The studies on sensory receptors and their ligands, such as mentioned above, remind us that, while studying sexual preference behavior, we should always be aware of the attributes of the stimuli presented to experimental animals. This is because many studies on sexual preference often use a complex of natural stimuli rather than a single chemical as experimental stimulus. Early studies reported that the odor of the homogenized preputial gland attracts the opposite sex in rats [29] and mice [30]. In a later study in rats, a mixture of squalene extracted from the preputial gland and combined with 2-heptanone and 4-ethyl phenol extracted from male urine was attractive for female rats [31].
Rodent urine contains MUPs. While most mammals studied, such as pigs, cattle, cats, and dogs, have only a single MUP (humans have none), rodents have >20 genes encoding MUPs [32]. Some are highly diverse and used for social recognition as individual volatile odor signatures [33]. On the other hand, some MUPs directly act on vomeronasal receptors as pheromones, promoting puberty onset (Vandenbergh effect, [34]), inducing estrus in cyclic females (Whitten effect, [35]), etc. These pheromones have relatively high molecular weights and are nonvolatile, so they require direct nose contact to transmit the signals [36,37]. Furthermore, MUPs not only function as pheromones, but some act as pheromone transporter molecules [38,39,40]. For example, 2-sec-butyl-4,5-dihydrothiazole and 3,4-deydro-exo-brevicomin are transported by binding a MUP pocket, and cooperatively attract receptive female mice, although none is effective on its own [41,42].
In many studies examining sexual preference, soiled beddings collected from home cages of estrous females and sexually active males have been used as paired stimuli [43,44,45,46,47]. In such tests, however, it is not possible to determine whether the experimental subjects responded to either volatile or nonvolatile chemicals, as they could detect both volatile and nonvolatile chemicals close to bedding by poking their noses into the soiled bedding. Even when using filter paper soaked with stimulus urine, preventing direct contact with the filter paper is important for determining stimulus attributes, such as volatile vs. nonvolatile [48,49]. In addition, if a highly volatile chemical is a critical attractant, the time between soiled bedding or urine sample collection and presentation and preservation is important.
The attractant for opposite-sex conspecifics may not be released only in the urine. In fact, the olfactory preference apparatus introduced above presented the odor emitted from awake stimulus animals through airflow to subjects [6]. During the test, stimulus animals sufficiently acclimated to the apparatus did not always excrete urine and feces, but rather emitted their body odors. Some studies used anesthetized animals as stimuli for the preference test [45,50]. In fact, MUPs transporting volatile pheromones are not only produced in the liver and excreted from the kidney, but also expressed in exocrine glands such as the mammary, lacrimal, parotid, sublingual, and submandibular glands [51,52]. It is possible that secretions from these glands warmed by body temperature result in pheromone volatilization, and consequently become an attractive odor for opposite-sex conspecifics.
Our study found that when the odors of castrated and gonadally intact males were simultaneously presented in the preference test, sexually active male rats preferred that of the castrated one [53,54,55]. We considered it unlikely that the lack of testosterone (T) due to castration may newly produce an attractant for males, but that the lack of T disinhibits the negative feedback of the hypothalamus-pituitary-gonadal (HPG) axis, resulting in increased circulating levels of gonadotropin-releasing hormone (GnRH) and gonadotropins, and consequently, that those became male incentive odors. Therefore, we examined the effect of GnRH antagonist administration to castrated male rats, and found that the castrated male attractiveness decreased to levels comparable to gonadally intact males [56]. Furthermore, we compared the responses of sexually active male rats to the odors of castrated males with and without surgical removal of the pituitary gland (hypophysectomy, HPx) to remove the source of gonadotropin, demonstrating longer exploration of the odor of pituitary-intact mice than that of HPx males [56]. However, the odor of HPx males was still more attractive than that of gonadally intact males, indicating that the attractiveness of castrated males is not due only to elevated gonadotropin levels but also to increased GnRH. Furthermore, we examined the effect of administration of human and equine chorionic gonadotropins (hCG and eCG) on luteinizing hormone (LH) and follicle stimulating hormone (FSH) action in the rat, respectively; in HPx males, the recovery of attractiveness followed eCG, but not hCH, injection [56]. We thus concluded that the attractiveness of castrated males to sexually active male rats is due to elevated levels of both GnRH and FSH due to the lack of circulating T.
Male rats attracted to the odor of castrated males must be primed with estrogen or aromatizable androgens because males treated with aromatase inhibitors or castrated males treated with nonaromatizable dihydrotestosterone (DHT) did not prefer castrated males [57], indicating that the odor of castrated males must be explicitly different from the estrous odors attracting castrated males treated with not only estrogen but DHT. In real ecological environments, however, there is no opportunity for castrated male odor to attract sexually active males. Nevertheless, the existence of neurons specifically activated by castrated male odors in the mouse AOB has been reported [58,59]. Furthermore, glomerular activity maps for sexually mature female mouse urine overlapped with maps for gonadectomized urine of both males and females [59]. Our studies, therefore, suggest that the production of this attractive odor also occurs in females during estrus evoked by the surge of GnRH and gonadotropins. In the future, identifying the molecules responsible for the attractiveness of castrated males (also of estrous females) needs to be investigated.
4. Sexually Dimorphic Preference and Organizational Effects of Sex Steroids
As shown in Figure 1B, preferences for conspecific odors are distinctly sexually dimorphic; sexually mature active males prefer the odor of receptive females to that of males or ovariectomized anestrous females, while receptive females prefer the odors of sexually active males to that of females or castrated sexually inactive males [6]. These sex-specific preferences are due to sex differences in the brain resulting from the organizational effects of perinatal sex steroids, as administration of opposite-sex hormones after gonadectomy in adult animals could not reproduce a heterosexual preference pattern [57,60,61].
Rodents with a short gestation period, such as rats and mice, have the critical (sex-steroid sensitive) period for brain sexual differentiation during the perinatal period. Neonatal castration in male rats significantly reduced the preference for receptive females, recovered by consequent treatment with testosterone propionate (TP) but not DHT propionate [62]. In addition, administration of the antiestrogen nitromifene citrate to pregnant rat dams eliminated the adult masculine preference in male offspring, whereas prenatal administration of an antiandrogen, cyproterone acetate, had no effects [63]. Furthermore, male rats neonatally injected with the aromatase inhibitor 1,4,6-androstatriene-3,17-dione (ATD) also showed decreased preference [4,5,64,65], which was more effective than prenatal ATD administration [4]. These results, together, indicate that estrogenic action during the critical period in early development is important for masculinization of sexual preferences in the rat. However, exposure of gonadally intact male rats to excessive androgen appears counterproductive, as TP embryonic administration (ED17–19) to pregnant dams has been reported to produce adult male offspring who spend longer times with stimulus males rather than estrous females [66]. Additionally, chronic TP administration to male pups by subcutaneous implantation of a Silastic capsule containing TP after birth did not affect the time spent with females, but increased the time spent with males in adult preference tests, resulting in a loss of sexual preference [67].
On the other hand, sex hormones are not necessary for the normal development of the female brain. Postnatal chronic estradiol benzoate (EB) administration to female rat pups during lactation results in male preferences, i.e., preferring estrus females to males after sexual maturity [68]. Contrarily, pubertal sex hormones seem to promote female preferences. It has been reported that although T treatment in adult ovariectomized females did not lead to any sexual preference, chronic estrogen administration during postnatal days 30 to 90 made ovariectomized females show female preference after T treatments [69] (in our study, though, ovariectomized females showed female preference by implantation of a Silastic capsule containing T [6]).
In mice deficient for the estrogen receptor-α gene (ERα [70]), males decreased investigation time for estrous females in the preference test using anesthetized males and females as stimuli, which resulted in abolishing sexual preference. In contrast, in mice deficient for the estrogen receptor-β (ERβ [71]) gene, the males showed a normal level of masculine preference comparable to that of WT mice. Further, aromatase-KO (ARKO) male mice lost their preference for both volatile and nonvolatile odors of estrous females [72]. According to these results, it can be considered that the estradiol (E2) converted from circulating T secreted from the male testes by the aromatase expressed in the mouse brain acts on ERα –similar to rats– to masculinize the male brain and promote the display of male-typical preference.
On the other hand, there is evidence of the involvement of androgen receptors (AR) in the sexual differentiation of sexual preference. Tfm (the androgen-insensitivity strain, named from the human syndrome, “testicular feminization”) males made longer nose contact to male cage bedding than to estrous female bedding, and showed no preference in a Y-maze test where the goal boxes had sexually active male or receptive female stimulus mice [73]. Another study demonstrated that female mice injected with DHT at birth showed normal feminine sexual behavior but masculine preference in adulthood, i.e., spending longer time in the investigation of estrous soiled bedding and staying in the vicinity of estrous females rather than males, while E2 injection at birth failed to masculinize the adult preference for soiled bedding [46]. However, conditional ARKO male mice in the nervous system sparing hypodevelopment of the reproductive organs, preferred to investigate female rather than male soiled bedding, like WT males [74]. Together, the differentiation of mouse sexual preferences may be affected by both ER and AR, and the difference might be minute, such that masculinization might be affected by ER and feminization by AR.
In rodents, it has been considered that sex hormones, especially estrogen, are not required for female brain differentiation to display feminine preference. However, compared to WT females, ARKO female mice showed shorter sniffing time investigating both male and female volatile odors, resulting in no preference [50]. Later, it was found that progesterone (P) receptor expression transiently appears in the anteroventral periventricular nucleus and medial part of the medial preoptic nucleus, both important brain regions for sexual preference, during postnatal days 15 to 25 in normal female mice, whereas those regions in ARKO females did not express the P receptor during that period [75]. Therefore, they examined the effect of prepubertal EB treatment in ARKO females, demonstrating that estrogen action during postnatal days 15 to 25 plays an important role for the normal development of sexual preference in female mice [76]. In other words, estrogen might achieve brain masculinization and defeminization during the fetal period, and conversely promote feminization of the brain after birth but before puberty.
5. Hormonal Control for Opposite-Sex Preference
Because preference for opposite-sex chemosensory signals is measured by an approaching behavior toward sexual partners, a part of mating behavior, it depends—as with sexual behavior—on circulating sex steroids. Female sexual behavior strictly requires estrogen binding to ERα in the brain and is enhanced by the P action following estrogen [77]. On the other hand, it is well established that estrogen, converted from circulating T in the brain, is also critical for male sexual behavior, especially in rodents. Male mice deficient for ERα [78,79] or aromatase [80] show severely impaired intromission behavior, although the penile functions, both reflexive and noncontact penile erection [79,81], require AR for activation [82,83,84,85].
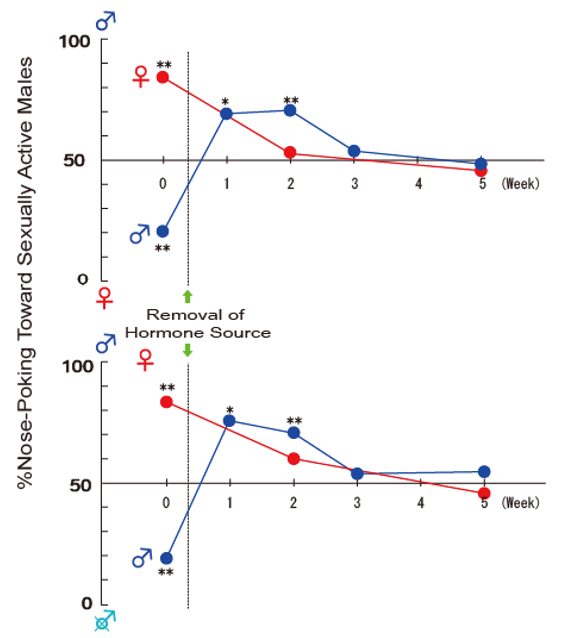
Similar to endocrine controls of sexual behavior, adult male rat castration eliminates masculine preference, which can be restored by T replacement therapy [6,62,64]. On the other hand, chronic treatment, but not a single injection [64,86], of either estrogen or DHT can induce masculine preference in castrated male rats [57]. Interestingly, male rat castration induces transient feminine preference, preferring male odors over estrous odor for one to two weeks followed by disappearing preference [6,57], while removal of the sex hormone source in female rats does eliminate sexual preference (Figure 2).
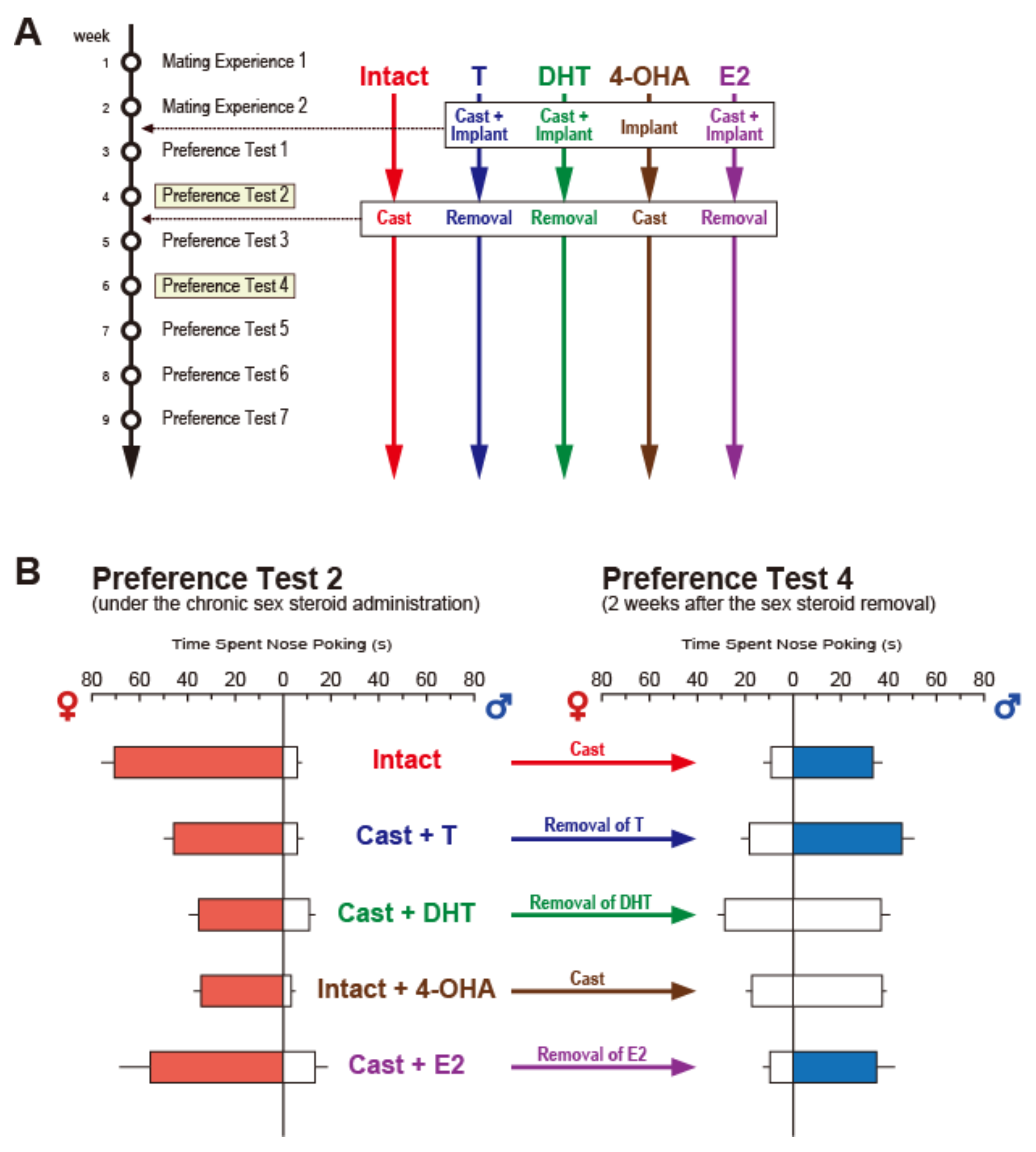
In order to examine which receptors, ER or AR, mediate this transient reversal of preference following castration, we prepared five experimental groups, each by two steps of hormonal manipulations: Intact group, first gonadally intact and then castration; T group, first castration and T implantation (subcutaneous implantation of a Silastic capsule containing T) and second, capsule removal; DHT group, first castration and DHT implantation and second, capsule removal; 4-OHA group: first gonadally intact and 4-OHA (4-hydroxyandrostenedione, steroidal aromatase inhibitor) implantation and second, castration; and E2 group, first castration and E2 implantation and second, capsule removal (Figure 3A). The olfactory preference tests were carried out twice after the first step and four times after the second [57]. In the preference test after the first step, all males displayed male-typical olfactory preferences, preferring estrous odor to male or anestrous odor, and castrated male odor to sexually active male odor. Following removal of the hormonal sources as the second step, males in Intact, T, and E2, but not DHT and 4-OHA, showed reversed female-typical preference, preferring male odor to estrous or castrated male odor, within 2 to 3 weeks after the second step (Figure 3B). Based on these results, we concluded that the transient reversal of sexual preference is mediated by ER, activated by estrogen, and converted from declined circulating T following castration [57].
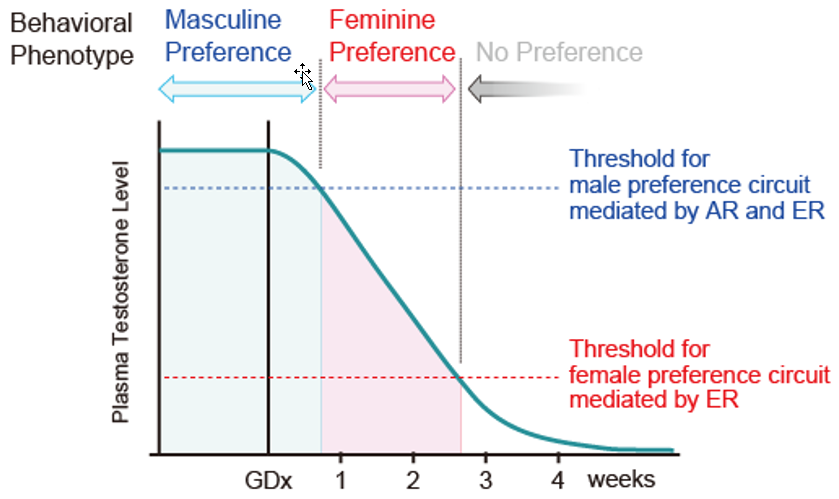
The model to explain this phenomenon is shown in Figure 4. This model assumes neural substrates for both masculine and feminine preferences in the rodent brain, and that masculine substrates have a much higher activation threshold by sex hormones via AR and ER than the feminine. In normal physiological states in males, the circulating androgen level is sufficiently high for activating the masculine substrate, prevailing over the feminine, and consequently determining the masculine preference phenotype. However, declining androgen levels after castration become insufficient to activate the masculine substrate but are still sufficient for the feminine substrates, making feminine preference the temporal phenotype, with both disappearing thereafter following further decreases in the androgen level. Chronical implantation of E2 capsules [57] or daily E2 injections [60] in castrated males increase the circulating androgen level beyond the masculine substrate threshold, consequently leading to a masculine preference, whereas a single E2 injection, insufficient for masculine but sufficient for feminine substrates, induces feminine preference in castrated males [6]. This model is also supported by the finding that semaphorin 7A gene deletion, which prevents migration of GnRH neurons born in the olfactory placode into the brain, resulting in hypoandrogenism, led to male mice showing feminine preference [87].
In female rats, the sexual preference for male volatile odors changes with estrous cycle, i.e., the preference is higher in the proestrus/estrus phase than in the diestrus phase, and diminishes with pregnancy [88]. This suggests that estrogen plays a critical role in the female preference for male odors, though chronic EB administration [1] or an E2 and P combination treatment [6,89] is required for a clear preference. In contrast, female rats showed preference for male soiled bedding independent of sexual experience and circulating sex hormones [90]. On the other hand, it has been reported that administration of 0.4 mg TP three times a week was insufficient for clear preference in female rats [1], whereas chronic T treatment by a Silastic capsule >2 weeks induces feminine preference [6,61]. In addition, chronic administration of a nonaromatizable androgen, DHT, and a synthetic and orally active anabolic androgen, methyltrienolone, also induce preference in female rats, indicating that the preference for male odors in female rats can be induced not only by ER activation but also potentially by AR activation [61].
As mentioned previously, female rat preference greatly depends on whether females are allowed to have direct mating with stimulus males [2]. This may be due to aversiveness of vaginal stimulation by males [1,2], but may be also affected by a female trait, the preference to stay with other females [91]. In mice, although estrous females prefer the volatile odors of sexually active males to those of females [53], virgin females show preference for female rather than male soiled bedding [92], having no aversive factor. Furthermore, the preference of female hamsters for male odors does not depend on sex hormones [93]. Further detailed analyses are needed for the elucidation of hormonal regulation in the feminine preference for male odors with consideration for species specificity, test conditions, mating experience, etc.
6. Regulation of Opposite-Sex Preference by Integrating the Main and Accessory Olfactory Systems
As mentioned previously, the rodent nasal cavity has mainly two chemosensory organs, the OE and the VNO. Because the latter is known as a pheromone receptor, extensive research has been carried out on the VNO function in a variety of social behaviors [94,95]. Although surgical VNO removal (VNOx) exerts a severe impact on sexual behavior in sexually naïve male hamsters [96], the effects on sexual behavior in rats [97,98,99] and mice [100,101] are weak and limited.
In contrast to sexual behavior, VNOx in female mice abolished the preference for male soiled bedding and decreased lordosis quotient (%displays of lordosis to male mounts), though it had no influence on the preference for volatile body or urine odors of males [45]. It also confirmed the ineffectiveness in the preference for airborne estrous female body odor in male rats [99]. On the other hand, TRPc2 gene deletion, which causes dysfunction in vomeronasal receptors, led to male mice displaying sexual instead of aggressive behavior toward males [102] and eliminated the preference for estrous female bedding [103,104]. Furthermore, female TRPc2-KO mice showed vigorous mounts and male sexual behavior, regardless of males and females [105]. However, surgical VNOx in female mice failed to promote mount behavior toward males [106]. This discrepancy in the effect between the gene KO and the surgical operation may be due to the broad distribution of the TRPc2 gene, densely expressed in vomeronasal neurons, but sparsely in erythroblasts, sperm, and the brain [107]. Dysfunction not only in the VNO but other central nervous activity caused by TRPc2-KO may have a different influence on sexual behavior.
OE destruction by zinc sulfate (ZnSO4) infusion into the nasal cavity did not completely eliminate male rat sexual behavior [99], but severely impaired male mouse sexual behavior [108] and female mouse lordosis [109], as well as eliminated the preference for volatile body odors [99,108,109] and for odors of directly contactable urine [108,109]. Furthermore, male mice with olfactory neuron loss, specifically in the dorsal OE and produced by targeted depletion of the cyclic nucleotide-gating olfactory channel combined with the diphtheria toxin fragment-A, had normal sniffing behavior and pheromonal detection but no preference for female urine [110]. In addition, ablation of α subunit of GO expressed in OE olfactory receptors induced structural changes in the MOB by reduced neurogenesis and increased cell death, subsequently decreasing the number of calbindin-positive interneurons and increasing that of thyrosine hydroxylase-positive interneurons in the MOB. As a result, male mice showed no preference for female soiled bedding and reduced anogenital investigation during mating, but still maintained the ability of food-finding [111]. In the rat, some males show no sexual behavior even after several mating sessions in a certain probability, interestingly; they have an olfactory deficit other than social odors and show no sexual preference [112,113].
Signals in the OE activate mitral cells in the MOB, whereas signals detected in the VNO activate mitral cells in the AOB. Optogenetical inhibition of mitral cells in the AOB significantly suppressed lordosis in ovariectomized, but EB and P primed, female mice, indicating that pheromones in the VON facilitate female sexual behavior in female mice [114]. In addition, male mice with lesions in the AOB showed a significant decrease in the time spent investigating estrous females, but still could discriminate between estrous and anestrous female odors or between male and female odors [115].
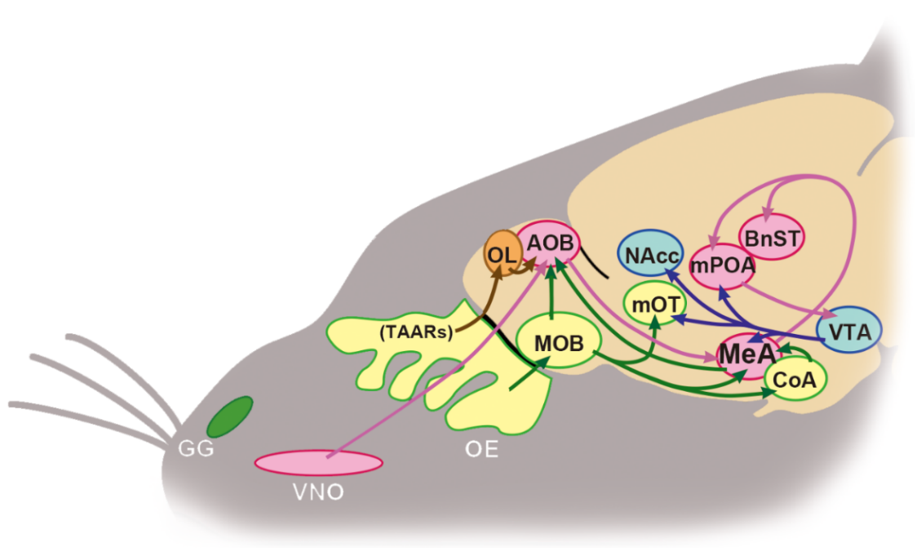
While the main and accessory olfactory systems have a parallel relationship in the projections, their interaction does not appear simple. ZnSO4 lesions in female OE mice prevented the increase of cFos-positive neurons in the MOB by exposure to the volatile odors in male urine but not in the AOB [109,116]. Conversely, VNOx suppressed increased cFos expression in AOB mitral cells after sexual behavior but not in the MOB [98]. Recently, it was shown that neurons in the MOB and the AOB have a complicated crosstalk in the bulb [117]. On the edge of the rostral AOB, i.e., between the dorsal part of the MOB and the AOB, there is a structure called the olfactory limbus, which receives afferents from a variety of OE receptors including TAARs. The olfactory limbus function is not yet properly characterized, but these input signals are likely aggregated to the large principal cells in the AOB and sent to the medial nucleus of the amygdala (MeA), which plays a critical role in social chemosensory signal processing [118].
It is also possible that the mutual interaction between the MOB and the AOB is mediated by the MeA, the secondary olfactory region. Optogenetic stimulation of mitral cells in the male mouse AOB expressing channelrhodopsin, and induced by Cre recombinase co-expressed with protocadherin 21, led to activation of main stream projection from the AOB to the MeA, and increased nose-poking to estrous female urine but not to male urine [119]. On the other hand, neurons in the MeA also receive afferent inputs from the MOB. Stimulation by volatile male urine odor in female mice induced cFos expression in mitral cells in the MOB, which send axons directly to the MeA [120]. Furthermore, injection of a retrograde neural tracer, cholera toxin B, into the AOB revealed that AOB neurons receive direct innervation from the MeA [121]. Mitral cells in the posteroventral MOB, which expresses an axon guidance molecule, Neuropilin 2 (Nrp2), send attractive social signals to the MeA. Mitral cell-specific Nrp2-KO abolished preference for estrous female urine in male mice and induced a loss of preference for castrated male urine containing female-attracting pheromone, (methylthio)methanethiol [122,123], in female mice [124].
Thus, it has become clear that the MeA, aggregating the main and accessory olfactory systems, plays an important role in the regulation of opposite-sex odor preference (Figure 5). A report comparing the effect of lesions in the anterior and posterior parts of the MeA in female mice demonstrated that both lesions severely suppressed lordosis, while only the posterior lesions eliminated preference for volatile and nonvolatile male urine odors [48]. In addition, silencing of MeA neurons by designer receptor exclusively activated by designer drug (DREADD) in female mice abolished the preference for volatile and nonvolatile male odors [125]. On the other hand, aromatase-positive MeA neurons receive afferent projections from the anterior part of the AOB, which relays pheromonal signals from V1R in the VNO. This projection from the AOB to the MeA is sexually dimorphic, much thicker in males than in females [126], and MeA neurons distribute social, sexual, and predator signals from the AOB to other regions, including the hypothalamic nuclei, in a sex-dependent fashion [127]. Dopamine receptor D1R-positive neurons in the posteroventral part of the MeA respond to both social and predator signals that mice should approach or avoid, respectively. When receiving dopamine (DA) signals from the midbrain ventral tegmental area (VTA), these D1R-positive neurons activate neurons in the bed nucleus of the stria terminalis (BnST), consequently leading mice to approach to the odor, whereas without DA signals from the VTA, these neurons activate others in the ventromedial hypothalamus, resulting in odor avoidance [128].
In contrast to this vomeronasal signal processing in the MeA, odor signals that drive innate behavior traveling via the MOB are conveyed to the cortical nucleus of the amygdala (CoA), and similarly to the MeA, sorted by whether to approach or avoid [129]. Optogenetic suppression of the projection form the MOB to the CoA lead to male mice that no longer avoid predator odors, and eliminated the approach to 2-phenylethanol, an innate mouse attractant [130]. In addition, simultaneous electrophysiological recordings of neuronal activities in the olfactory bulb and the amygdala in freely behaving female mice demonstrated synchronicity between the olfactory bulb and the MeA during male urine investigation, whereas synchronicity was found between the olfactory bulb and the posteromedial cortical amygdala (CoApm) during female urine investigation [131].
In male hamsters, a CoApm lesion has been reported to alter female investigation during mating but not to affect the preference for volatile odor of female bedding [132]. After injection of retrograde tracer, cholera toxin subunit B (CTB), into the anterior part of the MeA (MeAa) or the posterodorsal part of the MeA (MeApd) in male hamsters, activation of these labeled cells by cFos was examined when the injected animals were placed in a vacant female or male cage, suggesting the integration of social signaling in the MeAa and steroidal signaling in the MeApd [133,134]. Furthermore, combined unilateral lesions in the MeAa and MeApd impaired the preference for volatile odors in male hamsters when lesions were located contralaterally, but not ipsilaterally [134]. It should be noted that destruction of the nucleus of the lateral olfactory tract, a part of the MeAa, in male rats abolished avoidance of predator odors, decreased sexual behavior, and eliminated preference for estrous females [135].
In the neural regulation of male rat sexual behavior, estrous signals are transmitted to the MeA, processed, and transmitted to the BnST and medial preoptic area (mPOA) through the stria teminalis [136,137]. On the other hand, in male hamsters, combined lesions in the unilateral MeA and the contralateral mPOA impaired sexual behavior but not affected sexual preference, whereas combined lesions in the unilateral MeA and the contralateral BnST failed to suppress sexual behavior but did impair preference for volatile, but not nonvolatile, female odors [138]. These results suggest the importance of the BnST, more than the mPOA, in male hamster olfactory preference regulation. Indeed, bilateral lesions in the posterior BnST eliminated the preference for opposite-sex odors [139]. In contrast, mPOA lesions [140,141] or inactivation [142] completely suppressed olfactory preference for females in male rats, suggesting species-specific regulation of sexual preference even within rodents.
In female rats [143] and female hamsters [144], in contrast to males, mPOA destruction eliminated sexual preference for males. In female mice, mPOA neurons projecting to the VTA express neurotensin [145]. In response to male urine odor and E2, these neurons promote DA release in the nucleus accumbens (NAcc) [144], which plays an important role in motivational behavior as a part of the brain reward system. Indeed, DA release in the NAcc in sexually naïve female mice led to preference for male soiled bedding [146]. The medial olfactory tubercle (mOT), possibly involving reinforcement of odor learning and hedonic processing in drug abuse, is as another reward system driven by opposite-sex odors. The mOT receives direct inputs from not only the MOB [147] but also the MeA [148,149], and mOT lesions abolish preference for male bedding in female mice [150]. Following CTB injection into the female mouse mOT, exposure to male bedding showed a significant increase of cFos expression in MeA neurons and VTA neurons projecting to the mOT, whereas DREADD silencing in the mOT interrupted the preference for opposite-sex chemosignals in female mice [49].
On the other hand, serotonergic (5-HT) neurons in the midbrain raphe nuclei may also be involved in sexual preference. Male mice deficient in 5-HT neurons by co-expressing Cre with dPet1, a marker gene for 5-HT, combined with the floxed Lmx1b allele showed no preference for estrous female odor [44]. This was also confirmed in mice deficient for tryptophan hydroxylase 2, a synthetase mediating the first step of 5-HT biosynthesis [44]. Furthermore, female mice with 5-HT neuron loss by floxed Lmx1b showed transformed sexual preference, i.e., from estrous odors to male odors, and increased mounts toward receptive females [151]. However, there is a contradictory report that Tph2-KO male mice exhibit a normal masculine preference [152]. Because 5-HT neurons in the midbrain raphe nuclei have projections to the broad area in the whole forebrain with abundant types of receptors, exactly 14 including subtypes, each with its own characteristic function, a complete abolition in all 5-HT systems may have situation-dependent multiplex effects.
7. Experience and Sexual Preference
Male reproductive activity is strongly dependent on sexual experience, including sexual behavior. More than half of sexually naïve male rats, although interested in receptive females in the first mating session, would not accomplish a complete copulatory behavior. However, following weekly mating sessions, most males would begin sexual activity soon after the introduction of females. Such experience dependency is also observed in the noncontact erection of male rats, induced by estrous female odor [153]. Early studies showed that male rats without sexual experience show no preference for urine odors of estrous females [154], even though the pheromonal effect is an innate response [155]. In contrast, male mice [43,156] and male hamsters [157,158] show a preference for estrus female odor without sexual experience, although the gradual increase in time spent in olfactory investigation depends on sexual experience [53]. In hamsters, sexual experience affects the performance of sexual behavior after minor damage to the mPOA [159]. Herein, we attempt to discuss the effect of experience on preferences for opposite-sex odors in this section.
The olfactory environment during housing may exerts some influence on OE neurons. Sex-separated housed males from weaning to 6 months old exhibited more expressed genes in the OE, and a subset of these chemoreceptors showed altered expression frequencies depending on sex-separation and olfactory deprivation [160]. In addition, olfactory memory is formed at the olfactory bulb level, with estrogen facilitating odor learning in the MOB [161]; simultaneous olfactory inputs and noradrenaline release from the terminals of locus coeruleus neurons bring specific and long-lasting changes in the MOB, exerting influence on the preference for foods and urine odors [162]. In male mice, mating experience increases the densities of newborn cells in the AOB granule cells [163]. Furthermore, cFos expression in the piriform cortex, receiving direct projections from the MOB, coincided with sexual preferences before and after mating experience [164]. A study observing activities in mouse MeA neurons for several months by in vivo calcium imaging demonstrated that the MeApd exhibits persistent sex-specific changes mediated by oxytocin [165].
Sexual experience may exert physiological changes also in the mPOA. As mentioned above, excitotoxic lesions in the mPOA abolished the volatile odor preference of sexually inexperienced male hamsters, but had no effect on experienced males or nonvolatile chemosignals [159]. In mPOA, furthermore, nitrogen oxide synthase (NOS) expression in male rats [166] and AR, but not ERα, in male mice [167] increased by sexual behavior. Recently, Vgf gene expression was found to be increased in the rat mPOA including the sexually dimorphic nucleus of the preoptic area (SDN-POA) after sexual experience, while local Vgf knockdown in the mPOA by adeno-associated viral vector prevented experience-dependent enhancement of sexual preference [168]. Although the function of the Vgf gene is not well known at present, Vgf protein and its derived peptides are secretory and involved in reproductive functions, such as energy metabolism and penile erection [169,170], so further research is expected to clarify the relationship with sexual function and experience.
In addition to the innate and physiological effects of sexual experience described above, preference might be acquired and enhanced via conditioning. Sexually naïve male mice [43,156] and hamsters [171] can learn conditioned place preference (CPP) through exposure to estrous urine and bedding, meaning that those stimuli have a rewarding value. In male mice, CPP for such sexual stimuli could be acquired even after blocking vomeronasal inputs by surgical VNO removal [156], by lesion in the AOB, or after blocking olfactory epithelial inputs by ZnSO4 lesion in the nasal cavity, but not after blocking both [43]. When a male mouse encountered another male and estrous female, sexually naïve WT males attempted to investigate and mount the females, whereas TRPc2-KO mice showed comparable investigation and mounts toward both males and females [104]. At this time, DA release was found to increase in the NAcc of WT, but not TRPc2-KO mice [104]. Such activation of the reward system by DA may produce association learning with simultaneous sensory inputs, further reinforcing sexual preference [172,173].
In rat studies, sexually naïve males displaying no sexual preference showed not only increased cFos expression [174,175] but also increased DA release [176] in the NAcc in response to untouchable estrous females or estrous female bedding. Gaining sexual experience, however, significantly increased the number of cFos-immunoreactive cells [175] and DA release [176] in the male rat NAcc.
On the other hand, female mice, if able to directly contact male bedding or urine, exhibited preference for such nonvolatile male chemosignals without sexual experience, while if not able, they showed no preference for such male odor [177,178,179]. However, with repeated experience of direct contact with male bedding or urine, sexually inexperienced female mice started to show preference for volatile male odors [177,178,179]. In addition, sexually naïve females failed to acquire conditioned incentives by exposure to nonvolatile male odors in diestrus or after ovariectomy, implying its sex hormone dependency, whereas sexually experienced females successively acquired such conditioning even after ovariectomy, implying sex hormone independency [180]. Exposure to primary pheromones attracting sexually naïve female mice activates the basolateral nucleus of the amygdala (BLA) and the NAcc shell but not the VTA and the orbitofrontal cortex, whereas that of secondary attractants acquired via sexual experience activates not only the BLA but also the prefrontal cortex and the VTA [177]. Notably, the BLA role in connecting innate and learned attractants deserves attention.
In contrast to female mice, sexually inexperienced female rats show preference for male bedding consisting of both volatile and nonvolatile chemosignals [146,181,182] and for airborne male body odors [181]. Although exposure to male bedding in sexually inexperienced female rats did not change the number of cFos-immunoreactive cells in the NAcc [181], the increased DA release in the NAcc following male bedding stimulation was detected by microdialysis, and prevented by administration of a glutamate antagonist, resulting in decreased time spent investigating male bedding [146]. At present, where the neurons releasing DA in the NAcc project from is not known, but one such region may be the mOT. As mentioned in the previous section, because the mouse mOT receives direct innervation from the MOB and activates the reward system, it should be studied whether the rat mOT also involves in the mediation between chemosensory inputs and the reward system as well.
It is not that sexual experience has no effect on sexual preference in female rats. Following mating sessions with sexually active males where scented objects (a rubber duck with orange extract or a rubber bone with almond extract) were placed, female rats showed conditioned preference for those objects [183]. Namely, sexual experience acquires a reward value in sexual behavior, which makes possible associative odor learning. In addition, administration of selective androgen receptor modulator [9-chloro-2-ethyl-1-methyl-3-(2,2,2-trifluoroethyl)-3H-pyrrolo-[3,2-f]quinolin-7(6H)]-one after ovariectomy effectively induces sexual preference in sexually experienced, but not sexually naïve, female rats [184]. Furthermore, although no difference between sexually experienced and naïve female rats in cFos expression in the mPOA by sexual behavior was found, the increase in NOS expression was only detected in the mPOA of sexually experienced females [182]. Thus, sexual experience may induce physiological changes in female rat brain, affecting sexual preference.
8. Neuropeptidergic Regulation of Opposite-Sex Preference
Vassopressin (AVP) and oxytocin (OT) are posterior pituitary hormones, as well as neuroregulatory peptides, expressing in the main and accessory olfactory systems, the limbic systems, hypothalamic nuclei, etc. [185,186,187,188], and regulate a variety of olfactory-dependent social behaviors. For example, mice deficient for the OT gene showed a lack of social recognition [189,190,191], while mice deficient for the vasopressin receptor 1b (V1bR) could poorly discriminate familiar and novel females compared to WT mice [192].
Regarding the effect of preference for opposite-sex odor, the results seem to differ depending on stimulus presentation. In a three-chamber apparatus with side chambers having wire-mesh caged stimulus males or estrous females, WT male mice spent time in the female chamber longer than in the male chamber, whereas males deficient for the OT gene showed neither preference regardless of direct contact with the stimuli [53]. In contrast, OT-KO female mice did show a normal preference for male bedding as WT females [53]. However, another study reported that both male and female OT-KO mice exhibit a loss of preference for the odor of male and estrous female bodies [53]. In addition, conditioned oxytocin receptor (OTR)-KO in aromatase-positive neurons in the MeA abolished sexual preference in male mice [53], suggesting that chemosensory signals in the VNO and OE regulating sexual preference are processed in aromatase-positive oxytocin-sensitive neurons in the MeA. Pharmacological studies also demonstrated OT involvement in sexual preference. Oral administration of a non-peptide oxytocin antagonist in male mice impaired sexual preference, but did not affect social preference or social novelty preference [193]. In male rats, intraperitoneal injection of the above non-peptide oxytocin antagonist did not affect sexual preference per se, but significantly reduced time spent in investigation for estrous females [194].
OT also seems to enhance responses to social chemosensory signals via reward system activation. In estrogen-primed ovariectomized female mice, intracranial OT injection, although of little rewarding value, may have enhanced rewarding value if combined with social stimuli [195]. Male rats with intracranial injection of either OT or quinpirole, a D2 agonist, during repeated co-habituation with another male scented with almond extract came to prefer the scented male rather than estrous females [196]. These suggest a critical role of OT in the acquisition of attractive odors via the rewarding system.
On the other hand, there are not many studies on AVP in the regulation of sexual preference. Male mice with double depletion of two AVP receptor genes, V1aR and V1bR, showed a normal preference for estrous female bedding rather than that of males [197], and more preference for estrous body odor than that of males, comparable with that of WT males [197]. Nevertheless, the possible involvement of AVP in sexual preference may not be excluded, because of a report that male rats perinatally treated with ATD, an aromatase blocker, significantly increased the number of AVP-positive cells in the suprachiasmatic nucleus and became bisexual in the partner preference test [198]. In addition, WT and V1bR-KO male mice were subjected to preference tests with clean bedding vs. soiled bedding, revealing that the investigation time in WT males was higher for estrous female bedding than for that of males and higher for male or estrous female bedding than for a clean one, whereas V1bR-KO mice showed no preference at all [199]. Furthermore, it has also been reported that vasopressinergic projections from the MeA and BnST to the ventral pallidum (VP) in the rat are sexually dimorphic, much denser in males, and that local application of a V1aR antagonist into the VP impaired male preference for estrous females, but conversely enhanced female preference for males [200]. Further investigation is necessary to elucidate the involvement of the forebrain AVP system in the regulation of preference for opposite-sex odors.
There is another line of studies reporting that kisspeptin, which mediates between sex hormones and GnRH neurons in the HPG axis, is involved in the regulation of sexual preferences. Female mice exposure to male bedding or urine activated kisspeptin-immunoreactive cells in the rostral periventricular area of the third ventricle (RP3V) [201], and this activation was completely suppressed by VNO surgical removal but not by ZnSO4 lesion of the OE [202]. In fact, kisspeptin gene deletion abolished preference for male odors, while a compensatory treatment with kisspeptin recovered the preference [202]. Because RP3V kisspeptin neurons have direct projections to GnRH neurons, genetic disruption of Dicer in GnRH neurons, abolishing GnRH expression in GnRH::Cre; DicerloxP/loxP female mice produced a female-directed preference, but did not affect lordosis [202]. RP3V kisspeptin neurons have another downstream path, projecting to neuronal NOS (nNOS) in the ventrolateral part of the ventromedial hypothalamus (VMHvl). nNOS-KO female mice did not show a preference for male odors, while peripheral injection with a cocktail of NO donor and soluble guanylate cyclase agonist in nNOS-KO females recovered this preference for male odor [202].
On the other hand, kisspeptin may function in the regulation of male preference; specific activation of kisspeptin neurons in the MeApd by DREADD significantly facilitated investigatory behavior for estrous females during preference tests [203]. Local infusion of kisspeptin into the MeApd increased circulating LH and spontaneous penile erection, while infusion in the dorsal part of the MeApd produced only an increased level of circulating LH [204], suggesting that kisspeptin in the MeApd regulates sexual behavior separately to mediating the HPG axis. Presumably, the neural regulation of sexual preference by kisspeptin may be independent of its role in the HPG axis.
9. Future Outlook
In this review, we covered the available studies on the physiological mechanism of preference for chemosensory signals, which plays an important role in the social behavior of small rodents. This preference may be affected by several internal factors. That is, if animals show no preference at all when simultaneously presented with male and estrous female stimuli, the animals may have declined sexual motivation, lose the incentive value for the stimuli, or not be able to discriminate or detect those stimuli. The findings reviewed here include some requiring further analysis to determine which of these factors contribute to the result. In addition, we limited our review to the preference for chemosensory signals, but small rodents utilize other sensory modalities, such as ultrasonic vocalization, for courtship in mating. Social behavior in animals, including humans, is extremely complicated; therefore, further research is needed for their comprehensive understanding.
The sexual preference addressed here contains many elements as characteristics specific to each sex. This means that sex should be understood not as a dichotomy, but as a spectrum.
Author Contributions
Y.K. wrote the whole manuscript. H.H. contributed to drafting the manuscript by organizing literature especially on hormonal and peptidergic regulation of sexual preference and discussing them with Y.K. Both authors have read and agreed to publish the final version of the manuscript.
Funding
The work was supported by Grants-in Aid for Scientific Research from the Japan Society for the Promotion of Science (YK: 21K06274).
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors would like to express great appreciation to all the people, collaborators, and students concerned with this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Slob, A.; DeKlerk, L.; Brand, T. Homosexual and heterosexual partner preference in ovariectomized female rats: Effects of testosterone, estradiol and mating experience. Physiol. Behav. 1987, 41, 571–576. [Google Scholar] [CrossRef]
- Broekman, M.; De Bruin, M.; Smeenk, J.; Slob, A.K.; Van Der Schoot, P. Partner preference behavior of estrous female rats affected by castration of tethered male incentives. Horm. Behav. 1988, 22, 324–337. [Google Scholar] [CrossRef]
- Merkx, J.; Slob, A.; Bosch, J.V.D.W.T. Preference for an estrous female over a non-estrous female evinced by female rats requires dihydrotestosterone plus estradiol. Horm. Behav. 1989, 23, 466–472. [Google Scholar] [CrossRef]
- Brand, T.; Kroonen, J.; Mos, J.; Slob, A. Adult partner preference and sexual behavior of male rats affected by perinatal endocrine manipulations. Horm. Behav. 1991, 25, 323–341. [Google Scholar] [CrossRef] [Green Version]
- Bakker, J.; Van Ophemert, J.; Eijskoot, F.; Slob, A. A semiautomated test apparatus for studying partner preference behavior in the rat. Physiol. Behav. 1994, 56, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Kondo, Y.; Sakuma, Y. Sex-specific effects of gonadal steroids on conspecific odor preference in the rat. Horm. Behav. 2004, 46, 356–361. [Google Scholar] [CrossRef]
- Brechbühl, J.; Klaey, M.; Broillet, M.-C. Grueneberg Ganglion Cells Mediate Alarm Pheromone Detection in Mice. Science 2008, 321, 1092–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brechbühl, J.; Moine, F.; Klaey, M.; Nenniger-Tosato, M.; Hurni, N.; Sporkert, F.; Giroud, C.; Broillet, M.-C. Mouse alarm pheromone shares structural similarity with predator scents. Proc. Natl. Acad. Sci. USA 2013, 110, 4762–4767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brechbuhl, J.; Klaey, M.; Moine, F.; Bovay, E.; Hurni, N.; Nenniger-Tosato, M.; Broillet, M.C. Morphological and physiological species-dependent characteristics of the rodent Grueneberg ganglion. Front. Neuroanat. 2014, 8, 87. [Google Scholar]
- Stowers, L.; Kuo, T.-H. Mammalian pheromones: Emerging properties and mechanisms of detection. Curr. Opin. Neurobiol. 2015, 34, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Halem, H.A.; Baum, M.J.; Cherry, J.A. Sex Difference and Steroid Modulation of Pheromone-Induced Immediate Early Genes in the Two Zones of the Mouse Accessory Olfactory System. J. Neurosci. 2001, 21, 2474–2480. [Google Scholar] [CrossRef] [Green Version]
- Haga-Yamanaka, S.; Ma, L.; He, J.; Qiu, Q.; Lavis, L.D.; Looger, L.L.; Yu, C.R. Integrated action of pheromone signals in promoting courtship behavior in male mice. eLife 2014, 3, e03025. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.W.; Ackels, T.; Kuo, T.-H.; Cichy, A.; Dey, S.; Hays, C.; Kateri, M.; Logan, D.; Marton, T.F.; Spehr, M.; et al. Murine Pheromone Proteins Constitute a Context-Dependent Combinatorial Code Governing Multiple Social Behaviors. Cell 2014, 157, 676–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamero, P.; Marton, T.F.; Logan, D.; Flanagan, K.; Cruz, J.R.; Saghatelian, A.; Cravatt, B.F.; Stowers, L. Identification of protein pheromones that promote aggressive behaviour. Nature 2007, 450, 899–902. [Google Scholar] [CrossRef]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; McLean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Demir, E.; Li, K.; Bobrowski-Khoury, N.; Sanders, J.I.; Beynon, R.J.; Hurst, J.L.; Kepecs, A.; Axel, R. The pheromone darcin drives a circuit for innate and reinforced behaviours. Nature 2020, 578, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Haga, S.; Hattori, T.; Sato, T.; Sato, K.; Matsuda, S.; Kobayakawa, R.; Sakano, H.; Yoshihara, Y.; Kikusui, T.; Touhara, K. The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature 2010, 466, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Stopka, P.; Kuntová, B.; Klempt, P.; Havrdová, L.; Černá, M.; Stopková, R. On the saliva proteome of the Eastern European house mouse (Mus musculus musculus) focusing on sexual signalling and immunity. Sci. Rep. 2016, 6, 32481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niimura, Y. Olfactory Receptor Multigene Family in Vertebrates: From the Viewpoint of Evolutionary Genomics. Curr. Genom. 2012, 13, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberles, S.D. Trace Amine-associated Receptors Are Olfactory Receptors in Vertebrates. Ann. N. Y. Acad. Sci. 2009, 1170, 168–172. [Google Scholar] [CrossRef]
- Li, Q.; Liberles, S.D. Aversion and Attraction through Olfaction. Curr. Biol. 2015, 25, R120–R129. [Google Scholar] [CrossRef] [Green Version]
- Liberles, S.D. Trace amine-associated receptors: Ligands, neural circuits, and behaviors. Curr. Opin. Neurobiol. 2015, 34, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmeier, A.; Meyer, C.A.; Staempfli, A.; Casagrande, F.; Petrinovic, M.M.; Zhang, Y.-P.; Künnecke, B.; Iglesias, A.; Höner, O.P.; Hoener, M.C. How Female Mice Attract Males: A Urinary Volatile Amine Activates a Trace Amine-Associated Receptor That Induces Male Sexual Interest. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Korzan, W.J.; Ferrero, D.M.; Chang, R.B.; Roy, D.S.; Buchi, M.; Lemon, J.K.; Kaur, A.W.; Stowers, L.; Fendt, M.; et al. Synchronous Evolution of an Odor Biosynthesis Pathway and Behavioral Response. Curr. Biol. 2013, 23, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifico, R.; Dewan, A.; Cawley, D.; Guo, C.; Bozza, T. An Olfactory Subsystem that Mediates High-Sensitivity Detection of Volatile Amines. Cell Rep. 2012, 2, 76–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.A.; Tsai, L.; Roy, D.S.; Valenzuela, D.H.; Mosley, C.; Magklara, A.; Lomvardas, S.; Liberles, S.D.; Barnea, G. Neurons expressing trace amine-associated receptors project to discrete glomeruli and constitute an olfactory subsystem. Proc. Natl. Acad. Sci. USA 2012, 109, 13410–13415. [Google Scholar] [CrossRef] [Green Version]
- Kobayakawa, K.; Kobayakawa, R.; Matsumoto, H.; Oka, Y.; Imai, T.; Ikawa, M.; Okabe, M.; Ikeda, T.; Itohara, S.; Kikusui, T.; et al. Innate versus learned odour processing in the mouse olfactory bulb. Nature 2007, 450, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Omura, M.; Mombaerts, P. Trpc2-Expressing Sensory Neurons in the Main Olfactory Epithelium of the Mouse. Cell Rep. 2014, 8, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Orsulak, P.J.; Gawienowski, A.M. Olfactory Preferences for the Rat Preputial Gland1. Biol. Reprod. 1972, 6, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Bronson, F.H.; Caroom, D. Preputial gland of the male mouse: Attractant function. Reproduction 1971, 25, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Sun, L.; Zhang, J.H.; Feng, Z.Y. Sex- and gonad-affecting scent compounds and 3 male pheromones in the rat. Chem. Senses 2008, 33, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Logan, D.W.; Marton, T.F.; Stowers, L. Species Specificity in Major Urinary Proteins by Parallel Evolution. PLoS ONE 2008, 3, e3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, S.A.; Prescott, M.C.; Davidson, A.J.; McLean, L.; Beynon, R.J.; Hurst, J.L. Individual odour signatures that mice learn are shaped by involatile major urinary proteins (MUPs). BMC Biol. 2018, 16, 48. [Google Scholar] [CrossRef] [PubMed]
- Mucignat-Caretta, C.; Caretta, A.; Cavaggioni, A. Acceleration of puberty onset in female mice by male urinary proteins. J. Physiol. 1995, 486, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Marchlewska-Koj, A.; Cavaggioni, A.; Mucignat-Caretta, C.; Olejniczak, P. Stimulation of Estrus in Female Mice by Male Urinary Proteins. J. Chem. Ecol. 2000, 26, 2355–2366. [Google Scholar] [CrossRef]
- Pankevich, D.E.; Cherry, J.A.; Baum, M.J. Effect of vomeronasal organ removal from male mice on their preference for and neural Fos responses to female urinary odors. Behav. Neurosci. 2006, 120, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ricós, J.; Agustín-Pavón, C.; Lanuza, E.; Martínez-García, F. Intraspecific Communication Through Chemical Signals in Female Mice: Reinforcing Properties of Involatile Male Sexual Pheromones. Chem. Senses 2006, 32, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Ganfornina, M.D.; Gutiérrez, G.; Bastiani, M. A Phylogenetic Analysis of the Lipocalin Protein Family. Mol. Biol. Evol. 2000, 17, 114–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timm, D.E.; Baker, L.; Mueller, H.; Zidek, L.; Novotny, M.V. Structural basis of pheromone binding to mouse major urinary protein (MUP-I). Protein Sci. 2001, 10, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, S.D.; Robertson, D.H.L.; Cheetham, S.A.; Hurst, J.L.; Beynon, R.J. Structural and functional differences in isoforms of mouse major urinary proteins: A male-specific protein that preferentially binds a male pheromone. Biochem. J. 2005, 391, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jemiolo, B.; Alberts, J.; Sochinski-Wiggins, S.; Harvey, S.; Novotny, M. Behavioural and endocrine responses of female mice to synthetic analogues of volatile compounds in male urine. Anim. Behav. 1985, 33, 1114–1118. [Google Scholar] [CrossRef]
- Asaba, A.; Hattori, T.; Mogi, K.; Kikusui, T. Sexual attractiveness of male chemicals and vocalizations in mice. Front. Neurosci. 2014, 8, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korzan, W.J.; Freamat, M.; Johnson, A.G.; Cherry, J.A.; Baum, M.J. Either main or accessory olfactory system signaling can mediate the rewarding effects of estrous female chemosignals in sexually naive male mice. Behav. Neurosci. 2013, 127, 755–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Jiang, Y.; Si, Y.; Kim, J.-Y.; Chen, Z.-F.; Rao, Y. Molecular regulation of sexual preference revealed by genetic studies of 5-HT in the brains of male mice. Nature 2011, 472, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.; Pierman, S.; Douhard, Q.; Baum, M.J.; Bakker, J. The vomeronasal organ is required for the expression of lordosis behaviour, but not sex discrimination in female mice. Eur. J. Neurosci. 2006, 23, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodo, C.; Rissman, E.F. The Androgen Receptor Is Selectively Involved in Organization of Sexually Dimorphic Social Behaviors in Mice. Endocrinology 2008, 149, 4142–4150. [Google Scholar] [CrossRef] [PubMed]
- Meerts, S.H.; Clark, A.S. Stimulus animal characteristics do not modulate the expression of partner preference by female rats. Physiol. Behav. 2006, 89, 623–626. [Google Scholar] [CrossRef]
- DiBenedictis, B.T.; Ingraham, K.L.; Baum, M.J.; Cherry, J.A. Disruption of urinary odor preference and lordosis behavior in female mice given lesions of the medial amygdala. Physiol. Behav. 2012, 105, 554–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiBenedictis, B.T.; Olugbemi, A.O.; Baum, M.J.; Cherry, J.A. DREADD-Induced Silencing of the Medial Olfactory Tubercle Disrupts the Preference of Female Mice for Opposite-Sex Chemosignals. Eneuro 2015, 2. [Google Scholar] [CrossRef]
- Bakker, J.; Honda, S.-I.; Harada, N.; Balthazart, J. The Aromatase Knock-Out Mouse Provides New Evidence That Estradiol Is Required during Development in the Female for the Expression of Sociosexual Behaviors in Adulthood. J. Neurosci. 2002, 22, 9104–9112. [Google Scholar] [CrossRef]
- Gubits, R.M.; Lynch, K.R.; Kulkarni, A.B.; Dolan, K.P.; Gresik, E.W.; Hollander, P.; Feigelson, P. Differential regulation of alpha 2u globulin gene expression in liver, lachrymal gland, and salivary gland. J. Biol. Chem. 1984, 259, 12803–12809. [Google Scholar] [CrossRef]
- Shahan, K.; Denaro, M.; Gilmartin, M.; Shi, Y.; Derman, E. Expression of six mouse major urinary protein genes in the mammary, parotid, sublingual, submaxillary, and lachrymal glands and in the liver. Mol. Cell. Biol. 1987, 7, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, S.; Rai, D.; Terada, M.; Orikasa, C.; Nishimori, K.; Sakuma, Y.; Kondo, Y. Oxytocin is indispensable for conspecific-odor preference and controls the initiation of female, but not male, sexual behavior in mice. Neurosci. Res. 2019, 148, 34–41. [Google Scholar] [CrossRef]
- Achiraman, S.; Ponmanickam, P.; Ganesh, D.S.; Archunan, G. Detection of estrus by male mice: Synergistic role of olfactory–vomeronasal system. Neurosci. Lett. 2010, 477, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.C.; Nagata, K.; Sagoshi, S.; Ogawa, S. Estrogen and oxytocin involvement in social preference in male mice: A study using a novel long-term social preference paradigm with aromatase, estrogen receptor-α and estrogen receptor-β, oxytocin, and oxytocin receptor knockout male mice. Integr. Zoöl. 2018, 13, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Kumagai, R.; Kondo, Y. Why does castrated male odor attract sexually active male rats?–Attractivity induced by hypothalamus-pituitary-gonad axis block. Physiol. Behav. 2021, 230, 113288. [Google Scholar] [CrossRef]
- Xiao, K.; Chiba, A.; Sakuma, Y.; Kondo, Y. Transient reversal of olfactory preference following castration in male rats: Implication for estrogen receptor involvement. Physiol. Behav. 2015, 152, 161–167. [Google Scholar] [CrossRef]
- Luo, M.; Fee, M.S.; Katz, L.C. Encoding Pheromonal Signals in the Accessory Olfactory Bulb of Behaving Mice. Science 2003, 299, 1196–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammen, G.F.; Turaga, D.; Holy, T.E.; Meeks, J.P. Functional organization of glomerular maps in the mouse accessory olfactory bulb. Nat. Neurosci. 2014, 17, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, J.; Honda, S.; Harada, N.; Balthazart, J. Restoration of male sexual behavior by adult exogenous estrogens in male aromatase knockout mice. Horm. Behav. 2004, 46, 1–10. [Google Scholar] [CrossRef]
- Vega Matuszczyk, J.; Larsson, K. Role of androgen, estrogen and sexual experience on the female rat’s partner preference. Physiol Behav 1991, 50, 139–142. [Google Scholar] [CrossRef]
- Brand, T.; Slob, A. Neonatal organization of adult partner preference behavior in male rats. Physiol. Behav. 1991, 49, 107–111. [Google Scholar] [CrossRef]
- Matuszczyk, J.V.; Larsson, K. Sexual Preference and Feminine and Masculine Sexual Behavior of Male Rats Prenatally Exposed to Antiandrogen or Antiestrogen. Horm. Behav. 1995, 29, 191–206. [Google Scholar] [CrossRef]
- Bakker, J.; Brand, T.; van Ophemert, J.; Slob, A.K. Hormonal regulation of adult partner preference behavior in neonatally ATD-treated male rats. Behav. Neurosci. 1993, 107, 480–487. [Google Scholar] [CrossRef]
- Houtsmuller, E.; Brand, T.; de Jonge, F.; Joosten, R.; van de Poll, N.; Slob, A. SDN-POA volume, sexual behavior, and partner preference of male rats affected by perinatal treatment with ATD. Physiol. Behav. 1994, 56, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Cruz, C.D.; Pereira, O.C.M. Prenatal testosterone supplementation alters puberty onset, aggressive behavior, and partner preference in adult male rats. J. Physiol. Sci. 2012, 62, 123–131. [Google Scholar] [CrossRef]
- Henley, C.; Nunez, A.; Clemens, L. Exogenous androgen during development alters adult partner preference and mating behavior in gonadally intact male rats. Horm. Behav. 2010, 57, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henley, C.; Nunez, A.; Clemens, L. Estrogen treatment during development alters adult partner preference and reproductive behavior in female laboratory rats. Horm. Behav. 2009, 55, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, F.H.; Muntjewerff, J.W.; Louwerse, A.L.; Van De Poll, N.E. Sexual behavior and sexual orientation of the female rat after hormonal treatment during various stages of development. Horm. Behav. 1988, 22, 100–115. [Google Scholar] [CrossRef]
- Wersinger, S.R.; Rissman, E.F. Oestrogen receptor alpha is essential for female-directed chemo-investigatory behaviour but is not required for the pheromone-induced luteinizing hormone surge in male mice. J. Neuroendocr. 2000, 12, 103–110. [Google Scholar] [CrossRef]
- Bodo, C.; Rissman, E.F. New roles for estrogen receptor beta in behavior and neuroendocrinology. Front. Neuroendocrinol. 2006, 27, 217–232. [Google Scholar] [CrossRef]
- Bakker, J.; Honda, S.; Harada, N.; Balthazart, J. Sexual Partner Preference Requires a Functional Aromatase (Cyp19) Gene in Male Mice. Horm. Behav. 2002, 42, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Bodo, C.; Rissman, E.F. Androgen receptor is essential for sexual differentiation of responses to olfactory cues in mice. Eur. J. Neurosci. 2007, 25, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Raskin, K.; De Gendt, K.; Duittoz, A.; Liere, P.; Verhoeven, G.; Tronche, F.; Mhaouty-Kodja, S. Conditional Inactivation of Androgen Receptor Gene in the Nervous System: Effects on Male Behavioral and Neuroendocrine Responses. J. Neurosci. 2009, 29, 4461–4470. [Google Scholar] [CrossRef]
- Brock, O.; Douhard, Q.; Baum, M.J.; Bakker, J. Reduced Prepubertal Expression of Progesterone Receptor in the Hypothalamus of Female Aromatase Knockout Mice. Endocrinology 2010, 151, 1814–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, O.; Baum, M.J.; Bakker, J. The Development of Female Sexual Behavior Requires Prepubertal Estradiol. J. Neurosci. 2011, 31, 5574–5578. [Google Scholar] [CrossRef]
- Mazzucco, C.A.; Walker, H.A.; Pawluski, J.; Lieblich, S.E.; Galea, L.A. ERα, but not ERβ, mediates the expression of sexual behavior in the female rat. Behav. Brain Res. 2008, 191, 111–117. [Google Scholar] [CrossRef]
- Ogawa, S.; Lubahn, D.B.; Korach, K.; Pfaff, D.W. Behavioral effects of estrogen receptor gene disruption in male mice. Proc. Natl. Acad. Sci. USA 1997, 94, 1476–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imwalle, D.B.; Scordalakes, E.M.; Rissman, E.F. Estrogen Receptor α Influences Socially Motivated Behaviors. Horm. Behav. 2002, 42, 484–491. [Google Scholar] [CrossRef]
- Honda, S.; Harada, N.; Ito, S.; Takagi, Y.; Maeda, S. Disruption of sexual behavior in male aromatase-deficient mice lacking exons 1 and 2 of the cyp19 gene. Biochem. Biophys. Res. Commun. 1998, 252, 445–449. [Google Scholar] [CrossRef]
- Kondo, Y.; Tomihara, K.; Sakuma, Y. Sensory requirements for noncontact penile erection in the rat. Behav. Neurosci. 1999, 113, 1062–1070. [Google Scholar] [CrossRef]
- Bialy, M.; Nikolaev-Diak, A.; Kalata, U.; Nikolaev, E. Blockade of androgen receptor in the medial amygdala inhibits noncontact erections in male rats. Physiol. Behav. 2011, 103, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Bialy, M.; Sachs, B.D. Androgen Implants in Medial Amygdala Briefly Maintain Noncontact Erection in Castrated Male Rats. Horm. Behav. 2002, 42, 345–355. [Google Scholar] [CrossRef]
- Manzo, J.; Cruz, M.; Hernandez, M.E.; Pacheco, P.; Sachs, B. Regulation of Noncontact Erection in Rats by Gonadal Steroids. Horm. Behav. 1999, 35, 264–270. [Google Scholar] [CrossRef]
- Seo, S.I.; Kim, S.W.; Paick, J.S. The effects of androgen on penile reflex, erectile response to electrical stimulation and penile NOS activity in the rat. Asian J. Androl. 1999, 1, 169–174. [Google Scholar]
- Muroi, Y.; Ishii, T.; Komori, S.; Nishimura, M. A competitive effect of androgen signaling on male mouse attraction to volatile female mouse odors. Physiol. Behav. 2006, 87, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Schellino, R.; Trova, S.; Cimino, I.; Farinetti, A.; Jongbloets, B.C.; Pasterkamp, R.J.; Panzica, G.; Giacobini, P.; De Marchis, S.; Peretto, P. Opposite-sex attraction in male mice requires testosterone-dependent regulation of adult olfactory bulb neurogenesis. Sci. Rep. 2016, 6, 36063. [Google Scholar] [CrossRef]
- Eliasson, M.; Meyerson, B.J. Sexual preference in female rats during estrous cycle, pregnancy and lactation. Physiol. Behav. 1975, 14, 705–710. [Google Scholar] [CrossRef]
- Clark, A.S.; Kelton, M.C.; Guarraci, F.A.; Clyons, E.Q. Hormonal status and test condition, but not sexual experience, modulate partner preference in female rats. Horm. Behav. 2004, 45, 314–323. [Google Scholar] [CrossRef]
- Monchobogani, J.; Lanuza, E.; Lorente, M.; Martinez-Garcia, F. Attraction to male pheromones and sexual behaviour show different regulatory mechanisms in female mice. Physiol. Behav. 2004, 81, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Lukas, M.; Neumann, I.D. Social preference and maternal defeat-induced social avoidance in virgin female rats: Sex differences in involvement of brain oxytocin and vasopressin. J. Neurosci. Methods 2014, 234, 101–107. [Google Scholar] [CrossRef]
- Yao, S.; Bergan, J.; Lanjuin, A.; Dulac, C. Oxytocin signaling in the medial amygdala is required for sex discrimination of social cues. eLife 2017, 6, e31373. [Google Scholar] [CrossRef] [PubMed]
- Eidson, L.N.; Maras, P.M.; Epperson, E.; Petrulis, A. Female hamster preference for odors is not regulated by circulating gonadal hormones. Physiol. Behav. 2007, 91, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamero, P.; Leinders-Zufall, T.; Zufall, F. From genes to social communication: Molecular sensing by the vomeronasal organ. Trends Neurosci. 2012, 35, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, C.J.; Lepri, J.J. Consequences of removing the vomeronasal organ. J. Steroid Biochem. Mol. Biol. 1991, 39, 661–669. [Google Scholar] [CrossRef]
- Powers, J.; Winans, S. Vomeronasal organ: Critical role in mediating sexual behavior of the male hamster. Science 1975, 187, 961–963. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.R. Copulatory Behavior of Male Rats Paired with Natural Proestrous and Hormone-treated Ovariectomized Females. Exp. Anim. 1987, 36, 91–93. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Sudo, T.; Tomihara, K.; Sakuma, Y. Activation of accessory olfactory bulb neurons during copulatory behavior after deprivation of vomeronasal inputs in male rats. Brain Res. 2003, 962, 232–236. [Google Scholar] [CrossRef]
- Dhungel, S.; Masaoka, M.; Rai, D.; Kondo, Y.; Sakuma, Y. Both olfactory epithelial and vomeronasal inputs are essential for activation of the medial amygdala and preoptic neurons of male rats. Neuroscience 2011, 199, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Pankevich, D.E.; Baum, M.J.; Cherry, J.A. Olfactory Sex Discrimination Persists, Whereas the Preference for Urinary Odorants from Estrous Females Disappears in Male Mice after Vomeronasal Organ Removal. J. Neurosci. 2004, 24, 9451–9457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, S.K.J.; Martin, Y.H.; Salia, S.; Gamba, I.; Major, C.A.; Hassan, S.; Parsons, K.A.; Swift-Gallant, A. Puberty is a Critical Period for Vomeronasal Organ Mediation of Socio-sexual Behavior in Mice. Front. Behav. Neurosci. 2021, 14. [Google Scholar] [CrossRef]
- Stowers, L.; Holy, T.E.; Meister, M.; Dulac, C.; Koentges, G. Loss of Sex Discrimination and Male-Male Aggression in Mice Deficient for TRP2. Science 2002, 295, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Beny, Y.; Kimchi, T. Conditioned odor aversion induces social anxiety towards females in wild-type and TrpC2 knockout male mice. Genes Brain Behav. 2016, 15, 722–732. [Google Scholar] [CrossRef]
- Beny-Shefer, Y.; Zilkha, N.; Lavi-Avnon, Y.; Bezalel, N.; Rogachev, I.; Brandis, A.; Dayan, M.; Kimchi, T. Nucleus Accumbens Dopamine Signaling Regulates Sexual Preference for Females in Male Mice. Cell Rep. 2017, 21, 3079–3088. [Google Scholar] [CrossRef] [Green Version]
- Kimchi, T.; Xu, J.; Dulac, C. A functional circuit underlying male sexual behaviour in the female mouse brain. Nature 2007, 448, 1009–1014. [Google Scholar] [CrossRef]
- Martel, K.L.; Baum, M.J. Adult Testosterone Treatment But Not Surgical Disruption of Vomeronasal Function Augments Male-Typical Sexual Behavior in Female Mice. J. Neurosci. 2009, 29, 7658–7666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, B.A. Trpc2. Handb. Exp. Pharmacol. 2014, 222, 53–65. [Google Scholar]
- Keller, M.; Douhard, Q.; Baum, M.J.; Bakker, J. Sexual Experience Does Not Compensate for the Disruptive Effects of Zinc Sulfate—Lesioning of the Main Olfactory Epithelium on Sexual Behavior in Male Mice. Chem. Senses 2006, 31, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Douhard, Q.; Baum, M.J.; Bakker, J. Destruction of the Main Olfactory Epithelium Reduces Female Sexual Behavior and Olfactory Investigation in Female Mice. Chem. Senses 2006, 31, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Hattori, T.; Asaba, A.; Inoue, N.; Kanomata, N.; Kikusui, T.; Kobayakawa, R.; Kobayakawa, K. Genetic dissection of pheromone processing reveals main olfactory system-mediated social behaviors in mice. Proc. Natl. Acad. Sci. USA 2015, 112, E311–E320. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-M.; Kim, S.-S.; Choi, C.-I.; Cha, H.-L.; Oh, H.-H.; Ghil, S.; Lee, Y.-D.; Birnbaumer, L.; Suh-Kim, H. Development of the main olfactory system and main olfactory epithelium-dependent male mating behavior are altered in Go-deficient mice. Proc. Natl. Acad. Sci. USA 2016, 113, 10974–10979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimomi, Y.; Kondo, Y. Blunt olfaction in sexually sluggish male rats. Exp. Anim. Jpn. Assoc. Lab. Anim. Sci. 2020, 69, 441–447. [Google Scholar] [CrossRef]
- Portillo, W.; Paredes, R.G. Sexual incentive motivation, olfactory preference, and activation of the vomeronasal projection pathway by sexually relevant cues in non-copulating and naive male rats. Horm. Behav. 2004, 46, 330–340. [Google Scholar] [CrossRef]
- McCarthy, E.A.; Kunkhyen, T.; Korzan, W.J.; Naik, A.; Maqsudlu, A.; Cherry, J.A.; Baum, M.J. A comparison of the effects of male pheromone priming and optogenetic inhibition of accessory olfactory bulb forebrain inputs on the sexual behavior of estrous female mice. Horm. Behav. 2017, 89, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakupovic, J.; Kang, N.; Baum, M.J. Effect of bilateral accessory olfactory bulb lesions on volatile urinary odor discrimination and investigation as well as mating behavior in male mice. Physiol. Behav. 2008, 93, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, K.L.; Baum, M.J. Sexually dimorphic activation of the accessory, but not the main, olfactory bulb in mice by urinary volatiles. Eur. J. Neurosci. 2007, 26, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Barroso, V.; Ordaz-Sanchez, B.; Pena-Ortega, F.; Larriva-Sahd, J.A. Electrophysiological Evidence for a Direct Link between the Main and Accessory Olfactory Bulbs in the Adult Rat. Front. Neurosci. 2015, 9, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Barroso, V.; Peña-Ortega, F.; Larriva-Sahd, J.A. Olfaction and Pheromones: Uncanonical Sensory Influences and Bulbar Interactions. Front. Neuroanat. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunkhyen, T.; McCarthy, E.A.; Korzan, W.J.; Doctor, D.; Han, X.; Baum, M.J.; Cherry, J.A. Optogenetic Activation of Accessory Olfactory Bulb Input to the Forebrain Differentially Modulates Investigation of Opposite versus Same-Sex Urinary Chemosignals and Stimulates Mating in Male Mice. Eneuro 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, N.; Baum, M.J.; Cherry, J.A. A direct main olfactory bulb projection to the ‘vomeronasal’ amygdala in female mice selectively responds to volatile pheromones from males. Eur. J. Neurosci. 2009, 29, 624–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, K.L.; Baum, M.J. A centrifugal pathway to the mouse accessory olfactory bulb from the medial amygdala conveys gender-specific volatile pheromonal signals. Eur. J. Neurosci. 2009, 29, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Stowers, L.; Marton, T.F. What Is a Pheromone? Mammalian Pheromones Reconsidered. Neuron 2005, 46, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.Y.; Zhang, S.-Z.; Block, E.; Katz, L.C. Encoding social signals in the mouse main olfactory bulb. Nature 2005, 434, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Inokuchi, K.; Imamura, F.; Takeuchi, H.; Kim, R.; Okuno, H.; Nishizumi, H.; Bito, H.; Kikusui, T.; Sakano, H. Nrp2 is sufficient to instruct circuit formation of mitral-cells to mediate odour-induced attractive social responses. Nat. Commun. 2017, 8, 15977. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, E.A.; Maqsudlu, A.; Bass, M.; Georghiou, S.; Cherry, J.A.; Baum, M.J. DREADD-induced silencing of the medial amygdala reduces the preference for male pheromones and the expression of lordosis in estrous female mice. Eur. J. Neurosci. 2017, 46, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- Billing, A.; Correia, M.H.; Kelly, D.A.; Li, G.-L.; Bergan, J.F. Synaptic Connections of Aromatase Circuits in the Medial Amygdala Are Sex Specific. Eneuro 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Bergan, J.F.; Ben-Shaul, Y.; Dulac, C. Sex-specific processing of social cues in the medial amygdala. eLife 2014, 3, e02743. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.M.; Marcotulli, D.; Shen, A.; Zweifel, L.S. Divergent medial amygdala projections regulate approach–avoidance conflict behavior. Nat. Neurosci. 2019, 22, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Iurilli, G.; Datta, S.R. Population Coding in an Innately Relevant Olfactory Area. Neuron 2017, 93, 1180–1197. [Google Scholar] [CrossRef] [Green Version]
- Root, C.M.; Denny, C.A.; Hen, R.; Axel, R. The participation of cortical amygdala in innate, odour-driven behaviour. Nature 2014, 515, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Pardo-Bellver, C.; Martínez-Bellver, S.; Martínez-García, F.; Lanuza, E.; Teruel-Martí, V. Synchronized Activity in The Main and Accessory Olfactory Bulbs and Vomeronasal Amygdala Elicited by Chemical Signals in Freely Behaving Mice. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Maras, P.; Petrulis, A. The posteromedial cortical amygdala regulates copulatory behavior, but not sexual odor preference, in the male Syrian hamster (Mesocricetus auratus). Neuroscience 2008, 156, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maras, P.M.; Petrulis, A. The anterior medial amygdala transmits sexual odor information to the posterior medial amygdala and related forebrain nuclei. Eur. J. Neurosci. 2010, 32, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Maras, P.M.; Petrulis, A. Lesions that functionally disconnect the anterior and posterodorsal sub-regions of the medial amygdala eliminate opposite-sex odor preference in male Syrian hamsters (Mesocricetus auratus). Neuroscience 2010, 165, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Vaz, R.P.; Cardoso, A.; Sá, S.; Pereira, P.; Madeira, M.D. The integrity of the nucleus of the lateral olfactory tract is essential for the normal functioning of the olfactory system. Brain Struct. Funct. 2017, 222, 3615–3637. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Arai, Y. Functional association between the medial amygdala and the medial preoptic area in regulation of mating behavior in the male rat. Physiol. Behav. 1995, 57, 69–73. [Google Scholar] [CrossRef]
- Kondo, Y.; Yamanouchi, K. The possible involvement of the nonstrial pathway of the amygdala in neural control of sexual behavior in male rats. Brain Res. Bull. 1995, 38, 37–40. [Google Scholar] [CrossRef]
- Been, L.E.; Petrulis, A. Dissociated functional pathways for appetitive and consummatory reproductive behaviors in male Syrian hamsters. Horm. Behav. 2012, 61, 204–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Been, L.E.; Petrulis, A. Lesions of the posterior bed nucleus of the stria terminalis eliminate opposite-sex odor preference and delay copulation in male Syrian hamsters: Role of odor volatility and sexual experience. Eur. J. Neurosci. 2010, 32, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Dhungel, S.; Urakawa, S.; Kondo, Y.; Sakuma, Y. Olfactory preference in the male rat depends on multiple chemosensory inputs converging on the preoptic area. Horm. Behav. 2011, 59, 193–199. [Google Scholar] [CrossRef]
- Hurtazo, H.A.; Paredes, R.G. Olfactory preference and Fos expression in the accessory olfactory system of male rats with bilateral lesions of the medial preoptic area/anterior hypothalamus. Neuroscience 2005, 135, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Hurtazo, H.; Paredes, R.; Ågmo, A. Inactivation of the medial preoptic area/anterior hypothalamus by lidocaine reduces male sexual behavior and sexual incentive motivation in male rats. Neuroscience 2008, 152, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Guarraci, F.A.; Clark, A.S. Ibotenic acid lesions of the medial preoptic area disrupt the expression of partner preference in sexually receptive female rats. Brain Res. 2006, 1076, 163–170. [Google Scholar] [CrossRef]
- Martinez, L.A.; Petrulis, A. The medial preoptic area is necessary for sexual odor preference, but not sexual solicitation, in female Syrian hamsters. Horm. Behav. 2013, 63, 606–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHenry, J.A.; Otis, J.M.; Rossi, M.A.; Robinson, J.E.; Kosyk, O.; Miller, N.W.; McElligott, Z.A.; Budygin, E.A.; Rubinow, D.R.; Stuber, G.D. Hormonal gain control of a medial preoptic area social reward circuit. Nat. Neurosci. 2017, 20, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Catalán, M.J.; Orrico, A.; Hipólito, L.; Zornoza, T.; Polache, A.; Lanuza, E.; Martínez-García, F.; Granero, L.; Agustín-Pavón, C. Glutamate and Opioid Antagonists Modulate Dopamine Levels Evoked by Innately Attractive Male Chemosignals in the Nucleus Accumbens of Female Rats. Front. Neuroanat. 2017, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Wesson, D.W.; Wilson, D. Sniffing out the contributions of the olfactory tubercle to the sense of smell: Hedonics, sensory integration, and more? Neurosci. Biobehav. Rev. 2011, 35, 655–668. [Google Scholar] [CrossRef] [Green Version]
- Pardo-Bellver, C.; Cádiz-Moretti, B.; Novejarque, A.; Martínez-García, F.; Lanuza, E. Differential efferent projections of the anterior, posteroventral, and posterodorsal subdivisions of the medial amygdala in mice. Front. Neuroanat. 2012, 6, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiBenedictis, B.T.; Helfand, A.I.; Baum, M.J.; Cherry, J.A. A quantitative comparison of the efferent projections of the anterior and posterior subdivisions of the medial amygdala in female mice. Brain Res. 2014, 1543, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Agustín-Pavón, C.; Martinez-Garcia, F.; Lanuza, E. Focal lesions within the ventral striato-pallidum abolish attraction for male chemosignals in female mice. Behav. Brain Res. 2014, 259, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, Y.; Rao, Y. Serotonin signaling in the brain of adult female mice is required for sexual preference. Proc. Natl. Acad. Sci. USA 2013, 110, 9968–9973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angoa-Pérez, M.; Herrera-Mundo, N.; Kane, M.J.; Sykes, C.E.; Anneken, J.H.; Francescutti, D.M.; Kuhn, D.M. Brain Serotonin Signaling Does Not Determine Sexual Preference in Male Mice. PLoS ONE 2015, 10, e0118603. [Google Scholar] [CrossRef]
- Sachs, B.D. Erection evoked in male rats by airborne scent from estrous females. Physiol. Behav. 1997, 62, 921–924. [Google Scholar] [CrossRef]
- Lydell, K.; Doty, R.L. Male rat of odor preferences for female urine as a function of sexual experience, urine age, and urine source. Horm. Behav. 1972, 3, 205–212. [Google Scholar] [CrossRef]
- Olvera-Hernández, S.; Hernández, A.; Reyes, R.; Fernández-Guasti, A. Establishment of partner preference in male rats: Effect of prenatal letrozole and sexual experience. Horm. Behav. 2019, 109, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Pankevich, D.E.; Cherry, J.A.; Baum, M.J. Accessory olfactory neural Fos responses to a conditioned environment are blocked in male mice by vomeronasal organ removal. Physiol. Behav. 2006, 87, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Ballard, C.L.; Wood, R.I. Partner preference in male hamsters: Steroids, sexual experience and chemosensory cues. Physiol. Behav. 2007, 91, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maras, P.M.; Petrulis, A. Olfactory experience and the development of odor preference and vaginal marking in female Syrian hamsters. Physiol. Behav. 2008, 94, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Been, L.E.; Petrulis, A. The role of the medial preoptic area in appetitive and consummatory reproductive behaviors depends on sexual experience and odor volatility in male Syrian hamsters. Neuroscience 2010, 170, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Van Der Linden, C.; Jakob, S.; Gupta, P.; Dulac, C.; Santoro, S.W. Sex separation induces differences in the olfactory sensory receptor repertoires of male and female mice. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dillon, T.S.; Fox, L.C.; Han, C.; Linster, C. 17beta-estradiol enhances memory duration in the main olfactory bulb in CD-1 mice. Behav Neurosci 2013, 127, 923–931. [Google Scholar] [CrossRef] [Green Version]
- Shea, S.D.; Katz, L.C.; Mooney, R. Noradrenergic Induction of Odor-Specific Neural Habituation and Olfactory Memories. J. Neurosci. 2008, 28, 10711–10719. [Google Scholar] [CrossRef]
- Unda, N.M.; Eportillo, W.; Corona, R.; Paredes, R.G. Sexual Stimulation Increases the Survival of New Cells in the Accessory Olfactory Bulb of the Male Rat. Front. Neurosci. 2016, 10, 65. [Google Scholar] [CrossRef] [Green Version]
- Swaney, W.T.; Curley, J.P.; Champagne, F.A.; Keverne, E.B. The paternally expressed gene Peg3 regulates sexual experience-dependent preferences for estrous odors. Behav. Neurosci. 2008, 122, 963–973. [Google Scholar] [CrossRef]
- Li, Y.; Mathis, A.; Grewe, B.; Osterhout, J.A.; Ahanonu, B.; Schnitzer, M.J.; Murthy, V.; Dulac, C. Neuronal Representation of Social Information in the Medial Amygdala of Awake Behaving Mice. Cell 2017, 171, 1176–1190. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, J.M.; Brann, J.H.; Gil, M.; Hull, E.M. Sexual experience increases nitric oxide synthase in the medial preoptic area of male rats. Behav. Neurosci. 2006, 120, 1389–1394. [Google Scholar] [CrossRef] [Green Version]
- Swaney, W.; Dubose, B.N.; Curley, J.P.; Champagne, F.A. Sexual experience affects reproductive behavior and preoptic androgen receptors in male mice. Horm. Behav. 2012, 61, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maejima, S.; Abe, Y.; Yamaguchi, S.; Musatov, S.; Ogawa, S.; Kondo, Y.; Tsukahara, S. VGF in the Medial Preoptic Nucleus Increases Sexual Activity Following Sexual Arousal Induction in Male Rats. Endocrinology 2018, 159, 3993–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.E.; Brameld, J.M.; Jethwa, P.H. Neuroendocrine Role for VGF. Front. Endocrinol. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Succu, S.; Mascia, M.S.; Melis, T.; Sanna, F.; Melis, M.R.; Possenti, R.; Argiolas, A. Pro-VGF-derived peptides induce penile erection in male rats: Involvement of paraventricular nitric oxide. Neuropharmacology 2005, 49, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.R.; Meerts, S.H.; Sisk, C.L. Male Syrian hamsters demonstrate a conditioned place preference for sexual behavior and female chemosensory stimuli. Horm. Behav. 2010, 58, 410–414. [Google Scholar] [CrossRef] [Green Version]
- Gadziola, M.A.; Tylicki, K.A.; Christian, D.L.; Wesson, D.W. The Olfactory Tubercle Encodes Odor Valence in Behaving Mice. J. Neurosci. 2015, 35, 4515–4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, Q.; Wen, P.; Zhang, J.; Rao, X.; Zhou, Z.; Zhang, H.; He, X.; Li, J.; Zhou, Z.; et al. Activation of the dopaminergic pathway from VTA to the medial olfactory tubercle generates odor-preference and reward. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, H.H.; Ettenberg, A. Exposure to female rats produces differences in c-fos induction between sexually-naive and experienced male rats. Brain Res. 2002, 947, 57–66. [Google Scholar] [CrossRef]
- Hosokawa, N.; Chiba, A. Effects of sexual experience on conspecific odor preference and estrous odor-induced activation of the vomeronasal projection pathway and the nucleus accumbens in male rats. Brain Res. 2005, 1066, 101–108. [Google Scholar] [CrossRef]
- Fujiwara, M.; Chiba, A. Sexual odor preference and dopamine release in the nucleus accumbens by estrous olfactory cues in sexually naïve and experienced male rats. Physiol. Behav. 2018, 185, 95–102. [Google Scholar] [CrossRef]
- Moncho-Bogani, J.; Martinez-Garcia, F.; Novejarque, A.; Lanuza, E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala. Eur. J. Neurosci. 2005, 21, 2186–2198. [Google Scholar] [CrossRef]
- Moncho-Bogani, J.; Lanuza, E.; Hernandez, A.; Novejarque, A.; Martinez-Garcia, F. Attractive properties of sexual pheromones in mice: Innate or learned? Physiol. Behav. 2002, 77, 167–176. [Google Scholar] [CrossRef]
- Ramm, S.; Cheetham, S.A.; Hurst, J.L. Encoding choosiness: Female attraction requires prior physical contact with individual male scents in mice. Proc. R. Soc. B Biol. Sci. 2008, 275, 1727–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, E.A.; Naik, A.S.; Coyne, A.F.; Cherry, J.A.; Baum, M.J. Effect of Ovarian Hormones and Mating Experience on the Preference of Female Mice to Investigate Male Urinary Pheromones. Chem. Senses 2017, 43, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, N.; Chiba, A. Effects of sexual experience on conspecific odor preference and male odor-induced activation of the vomeronasal projection pathway and the nucleus accumbens in female rats. Brain Res. 2007, 1175, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Meerts, S.H.; Park, S.; Sekhawat, R. Sexual experience modulates partner preference and mPOA nitric oxide synthase in female rats. Behav. Neurosci. 2016, 130, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Piergies, A.M.; Hicks, M.E., Jr.; Schwartz, J.P.; Meerts, S.H. Sexually experienced, but not naïve, female rats show a conditioned object preference (COP) for mating after a single training trial. Physiol. Behav. 2019, 198, 42–47. [Google Scholar] [CrossRef]
- Kudwa, A.E.; López, F.J.; McGivern, R.F.; Handa, R.J. A Selective Androgen Receptor Modulator Enhances Male-Directed Sexual Preference, Proceptive Behavior, and Lordosis Behavior in Sexually Experienced, But Not Sexually Naive, Female Rats. Endocrinology 2010, 151, 2659–2668. [Google Scholar] [CrossRef] [PubMed]
- Mitre, M.; Marlin, B.J.; Schiavo, J.K.; Morina, E.; Norden, S.E.; Hackett, T.A.; Aoki, C.J.; Chao, M.V.; Froemke, R.C. A Distributed Network for Social Cognition Enriched for Oxytocin Receptors. J. Neurosci. 2016, 36, 2517–2535. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and Estrogen Receptor beta in the Brain: An Overview. Front. Endocrinol. 2015, 6, 160. [Google Scholar] [CrossRef] [Green Version]
- Vaccari, C.; Lolait, S.J.; Ostrowski, N.L. Comparative Distribution of Vasopressin V1b and Oxytocin Receptor Messenger Ribonucleic Acids in Brain1. Endocrinology 1998, 139, 5015–5033. [Google Scholar] [CrossRef] [PubMed]
- Wacker, D.; Ludwig, M. The role of vasopressin in olfactory and visual processing. Cell Tissue Res. 2018, 375, 201–215. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Young, L.J.; Hearn, E.F.; Matzuk, M.M.; Insel, T.R.; Winslow, J.T. Social amnesia in mice lacking the oxytocin gene. Nat. Genet. 2000, 25, 284–288. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Aldag, J.M.; Insel, T.R.; Young, L.J. Oxytocin in the Medial Amygdala is Essential for Social Recognition in the Mouse. J. Neurosci. 2001, 21, 8278–8285. [Google Scholar] [CrossRef]
- Oettl, L.-L.; Ravi, N.; Schneider, M.; Scheller, M.F.; Schneider, P.; Mitre, M.; da Silva Gouveia, M.; Froemke, R.; Chao, M.; Young, W.S.; et al. Oxytocin Enhances Social Recognition by Modulating Cortical Control of Early Olfactory Processing. Neuron 2016, 90, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Wersinger, S.R.; Ginns, E.I.; O’Carroll, A.M.; Lolait, S.J.; Young, W.S., 3rd. Vasopressin V1b receptor knockout reduces aggressive behavior in male mice. Mol. Psychiatry 2002, 7, 975–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Zerda, S.H.; Netser, S.; Magalnik, H.; Wagner, S. Impaired sex preference, but not social and social novelty preferences, following systemic blockade of oxytocin receptors in adult male mice. Psychoneuroendocrinology 2020, 116, 104676. [Google Scholar] [CrossRef]
- Blitzer, D.; Wells, T.; Hawley, W. Administration of an oxytocin receptor antagonist attenuates sexual motivation in male rats. Horm. Behav. 2017, 94, 33–39. [Google Scholar] [CrossRef]
- Kent, K.; Arientyl, V.; Khachatryan, M.M.; Wood, R.I. Oxytocin Induces a Conditioned Social Preference in Female Mice. J. Neuroendocr. 2013, 25, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Rio, R.T.-D.; Silvaran, M.B.T.; Díaz-Estrada, V.X.; Herrera-Covarrubias, D.; Corona-Morales, A.A.; Pfaus, J.G.; Coria-Avila, G.A. Conditioned same-sex partner preference in male rats is facilitated by oxytocin and dopamine: Effect on sexually dimorphic brain nuclei. Behav. Brain Res. 2015, 283, 69–77. [Google Scholar] [CrossRef]
- Shimizu, K.; Nakamura, K.; Yokosuka, M.; Kondo, Y. Modulation of male mouse sociosexual and anxiety-like behaviors by vasopressin receptors. Physiol. Behav. 2018, 197, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Swaab, D.; Slob, A.; Houtsmuller, E.; Brand, T.; Zhou, J. Increased number of vasopressin neurons in the suprachiasmatic nucleus (SCN) of ‘bisexual’ adult male rats following perinatal treatment with the aromatase blocker ATD. Dev. Brain Res. 1995, 85, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Wersinger, S.R.; Kelliher, K.R.; Zufall, F.; Lolait, S.J.; O’Carroll, A.M.; Young, W.S., 3rd. Social motivation is reduced in vasopressin 1b receptor null mice despite normal performance in an olfactory discrimination task. Horm. Behav. 2004, 46, 638–645. [Google Scholar] [CrossRef] [Green Version]
- DiBenedictis, B.T.; Cheung, H.K.; Nussbaum, E.R.; Veenema, A. Involvement of ventral pallidal vasopressin in the sex-specific regulation of sociosexual motivation in rats. Psychoneuroendocrinology 2020, 111, 104462. [Google Scholar] [CrossRef]
- Bakker, J.; Pierman, S.; Gonzales-Martinez, D. Effects of aromatase mutation (ArKO) on the sexual differentiation of kisspeptin neuronal numbers and their activation by same versus opposite sex urinary pheromones. Horm. Behav. 2010, 57, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Hellier, V.; Brock, O.; Candlish, M.; Desroziers, E.; Aoki, M.; Mayer, C.; Piet, R.; Herbison, A.; Colledge, W.H.; Prevot, V.; et al. Female sexual behavior in mice is controlled by kisspeptin neurons. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Adekunbi, D.; Li, X.F.; Lass, G.; Shetty, K.; Adegoke, O.A.; Yeo, S.H.; Colledge, W.H.; Lightman, S.L.; O’Byrne, K.T. Kisspeptin neurones in the posterodorsal medial amygdala modulate sexual partner preference and anxiety in male mice. J. Neuroendocr. 2018, 30, e12572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresham, R.; Li, S.; Adekunbi, D.; Hu, M.; Li, X.F.; O’Byrne, K.T. Kisspeptin in the medial amygdala and sexual behavior in male rats. Neurosci. Lett. 2016, 627, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
(A). The apparatus for olfactory preference test in rats, consisting of three compartments. Experimental rats were placed in the middle, and stimulus rats were placed in two neighboring side compartments. A fan connecting to the airduct at the ceiling of the middle compartment dispersed airflow from the two side compartments to convey the odors of stimulus rats. A transparent tube was placed on each air inlet in the middle compartment to facilitate observation of nose-poking. (B) and (C). Typical olfactory preference shown by sexually active male rats (B) and receptive female rats (C). Sex symbol characters on both sides of each column indicate the type of stimulus rat. Crosses over the sex symbol characters indicate gonadectomy of stimulus rat [6]. Reprinted with permission from Ref. [6]. Copyright 2004 Elsevier.
Figure 1.
(A). The apparatus for olfactory preference test in rats, consisting of three compartments. Experimental rats were placed in the middle, and stimulus rats were placed in two neighboring side compartments. A fan connecting to the airduct at the ceiling of the middle compartment dispersed airflow from the two side compartments to convey the odors of stimulus rats. A transparent tube was placed on each air inlet in the middle compartment to facilitate observation of nose-poking. (B) and (C). Typical olfactory preference shown by sexually active male rats (B) and receptive female rats (C). Sex symbol characters on both sides of each column indicate the type of stimulus rat. Crosses over the sex symbol characters indicate gonadectomy of stimulus rat [6]. Reprinted with permission from Ref. [6]. Copyright 2004 Elsevier.

Figure 2.
Effects of sex hormone ablation on sexual preference. The results in preference tests between sexually active male vs. estrous female and between sexually active male vs. castrated male are shown in the upper and lower panels, respectively. Ovariectomized female rats were primed with estrogen and progesterone in the preference test at week 0, and thereafter tested without sex hormone treatment. Male rats were gonadally intact and castrated immediately after the preference test at week 0. Male rats transiently displayed the feminine type of sexual preference during 2 weeks after castration [6].
Figure 2.
Effects of sex hormone ablation on sexual preference. The results in preference tests between sexually active male vs. estrous female and between sexually active male vs. castrated male are shown in the upper and lower panels, respectively. Ovariectomized female rats were primed with estrogen and progesterone in the preference test at week 0, and thereafter tested without sex hormone treatment. Male rats were gonadally intact and castrated immediately after the preference test at week 0. Male rats transiently displayed the feminine type of sexual preference during 2 weeks after castration [6].

Figure 3.
We examined the hormonal influence on the transient reversal of sexual preference observed after castration in male rats [57]. (A): in the study, all male rats received mating experience sessions twice weekly followed by seven preference tests. The intact group was castrated after the second preference test; T, DHT, and E2 groups were castrated a week before the first preference test and simultaneously implanted with Silastic capsules containing T, DHT, and E2, respectively. 4-OHA group received a subcutaneous implantation of a Silastic capsule containing a steroidal aromatase inhibitor, 4-OHA, a week before the first test and were castrated after the second preference test. (B) shows only the results in preference tests 2 and 4 as the most important points in this experiment. The DHT and 4-OHA groups showed a lack of estrogen after treatment, resulting in no reversed preference after steroid supply ablation. Colored bar in each column of (B) indicates significantly longer time in nose-poking toward females than males (red) or toward males than females (blue) (p < 0.05). Reprinted with permission from Ref. [57]. Copyright 2015 Elsevier.
Figure 3.
We examined the hormonal influence on the transient reversal of sexual preference observed after castration in male rats [57]. (A): in the study, all male rats received mating experience sessions twice weekly followed by seven preference tests. The intact group was castrated after the second preference test; T, DHT, and E2 groups were castrated a week before the first preference test and simultaneously implanted with Silastic capsules containing T, DHT, and E2, respectively. 4-OHA group received a subcutaneous implantation of a Silastic capsule containing a steroidal aromatase inhibitor, 4-OHA, a week before the first test and were castrated after the second preference test. (B) shows only the results in preference tests 2 and 4 as the most important points in this experiment. The DHT and 4-OHA groups showed a lack of estrogen after treatment, resulting in no reversed preference after steroid supply ablation. Colored bar in each column of (B) indicates significantly longer time in nose-poking toward females than males (red) or toward males than females (blue) (p < 0.05). Reprinted with permission from Ref. [57]. Copyright 2015 Elsevier.

Figure 4.
Model explaining the transient reversed preference observed after male rat castration. We hypothesize that the male rat brain has both circuits for expression of masculine and feminine preference. In gonadally intact males, their circulating testosterone is sufficient to activate the masculine circuit, displaying dominant masculine preferences, and obscuring feminine preferences. However, the declining level of circulating testosterone after castration becomes insufficient to activate the masculine circuit, while still activating the feminine circuit because of its lower threshold in steroid sensitivity, therefore leading to a temporal feminine preference as the level falls below the threshold.
Figure 4.
Model explaining the transient reversed preference observed after male rat castration. We hypothesize that the male rat brain has both circuits for expression of masculine and feminine preference. In gonadally intact males, their circulating testosterone is sufficient to activate the masculine circuit, displaying dominant masculine preferences, and obscuring feminine preferences. However, the declining level of circulating testosterone after castration becomes insufficient to activate the masculine circuit, while still activating the feminine circuit because of its lower threshold in steroid sensitivity, therefore leading to a temporal feminine preference as the level falls below the threshold.

Figure 5.
Schematic diagram of supposed chemosignals that modulate preference for opposite sex in mice. Red, green, brown, and blue arrows represent vomeronasal, main olfactory epithelial, TAARs, and DA rewarding signal streams, respectively. OE: olfactory epithelium, VNO: vomeronasal organ, GG: Grueneberg ganglion, TAAR: trace amine-associated receptor, MOB: main olfactory bulb, AOB: accessory olfactory bulb, OL: olfactory limbus, mOT: medial olfactory tubercle, CoA: cortical nucleus of amygdala, MeA, medial nucleus of amygdala, BnST: bed nucleus of stria terminalis, mPOA: medial preoptic area, VTA: ventral tegmental area, NAcc: nucleus accumbens.
Figure 5.
Schematic diagram of supposed chemosignals that modulate preference for opposite sex in mice. Red, green, brown, and blue arrows represent vomeronasal, main olfactory epithelial, TAARs, and DA rewarding signal streams, respectively. OE: olfactory epithelium, VNO: vomeronasal organ, GG: Grueneberg ganglion, TAAR: trace amine-associated receptor, MOB: main olfactory bulb, AOB: accessory olfactory bulb, OL: olfactory limbus, mOT: medial olfactory tubercle, CoA: cortical nucleus of amygdala, MeA, medial nucleus of amygdala, BnST: bed nucleus of stria terminalis, mPOA: medial preoptic area, VTA: ventral tegmental area, NAcc: nucleus accumbens.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kondo, Y.; Hayashi, H. Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals. Int. J. Mol. Sci. 2021, 22, 8311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158311
AMA Style
Kondo Y, Hayashi H. Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals. International Journal of Molecular Sciences. 2021; 22(15):8311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158311
Chicago/Turabian StyleKondo, Yasuhiko, and Himeka Hayashi. 2021. "Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals" International Journal of Molecular Sciences 22, no. 15: 8311. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22158311
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.