
Effects of HMGB1 on Tricellular Tight Junctions via TGF-β Signaling in Human Nasal Epithelial Cells

 , , and
, , and
Abstract
:1. Introduction
2. Results
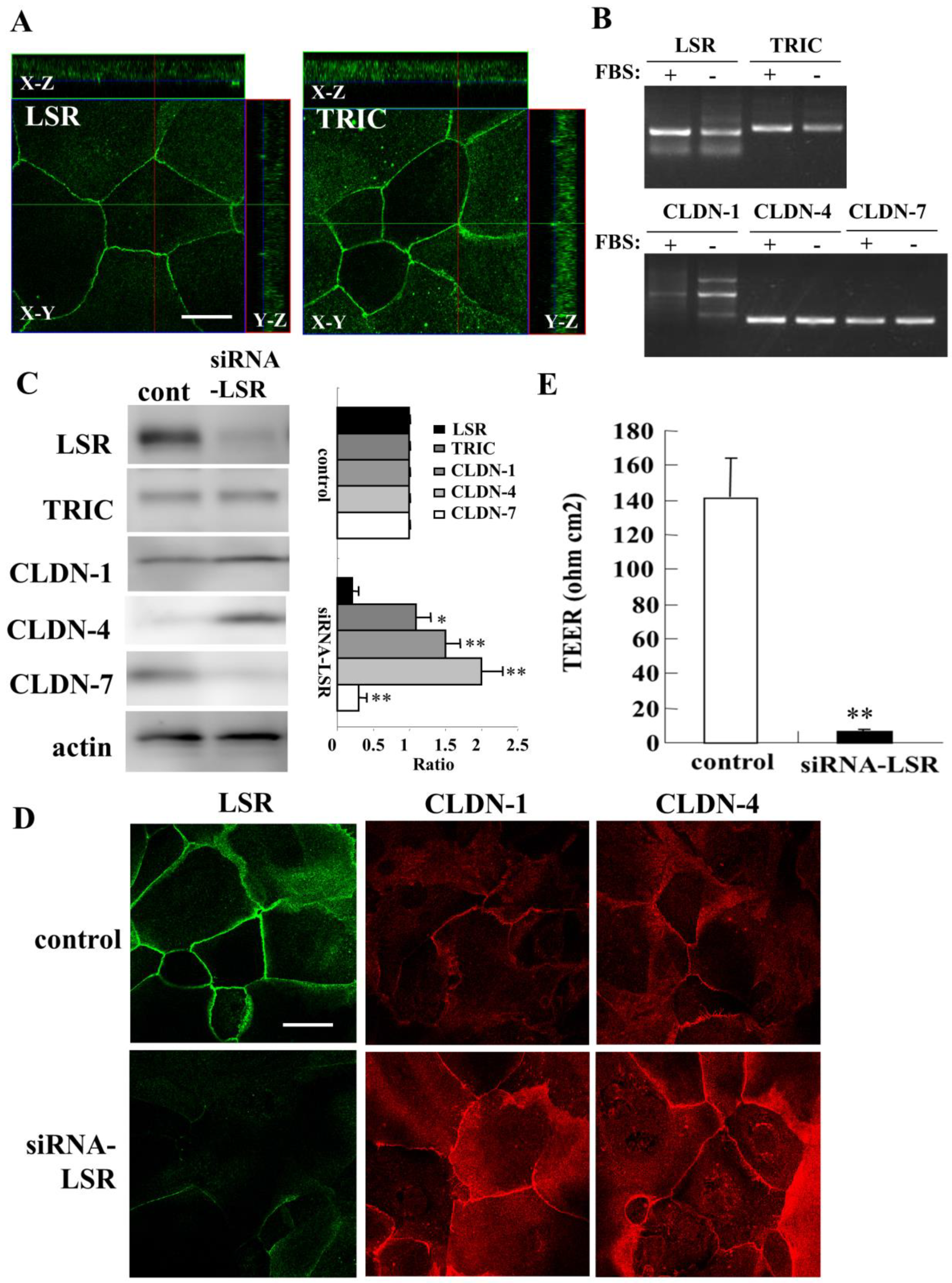
2.1. Knockdown of Angulin-1/LSR Downregulates Claudin-7 and Epithelial Barrier Function and Upregulates of CLDN-1 and -4 in Primary Cultured Human Nasal Epithelial Cells (HNECs)
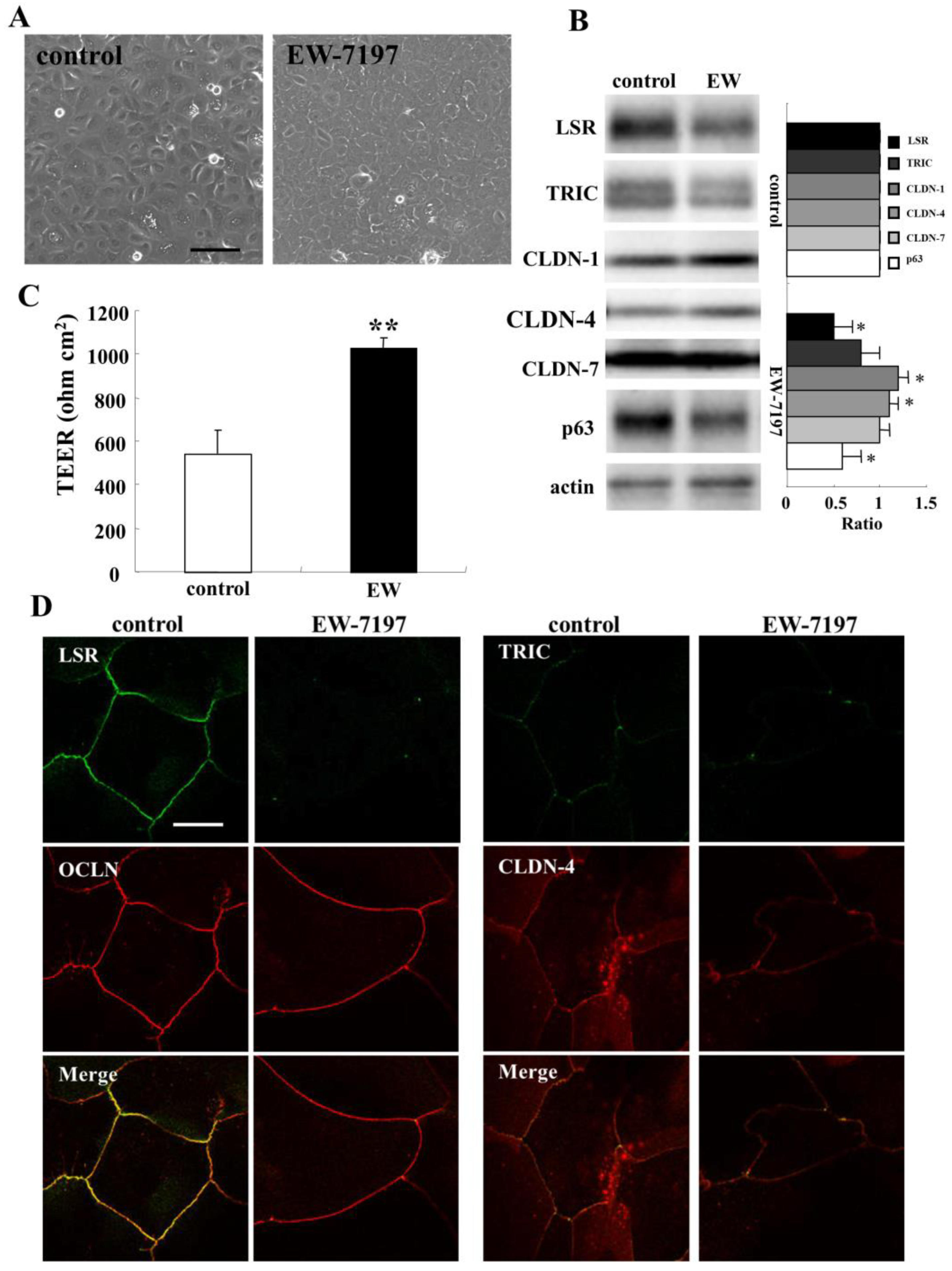
2.2. TGF-β Receptor Type 1 Inhibitor EW-7197 Concentrates Expression of Angulin-1/LSR and TRIC at tTJs from bTJs and Increases Epithelial Barrier in HNECs
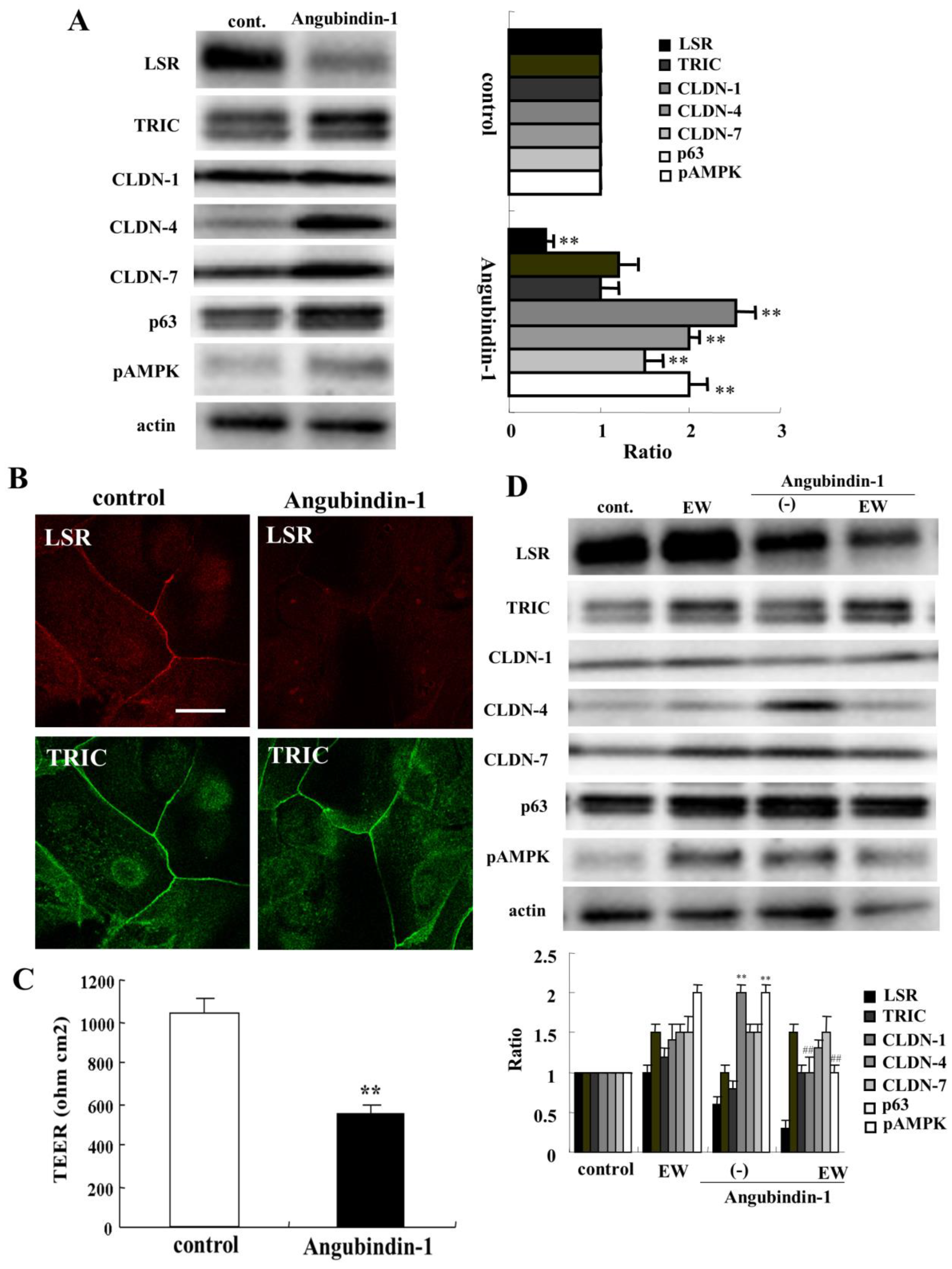
2.3. A Binder to Angulin-1/LSR Angubindin-1 Decreases Anglin-1/LSR and Epithelial Barrier Function in HNECs
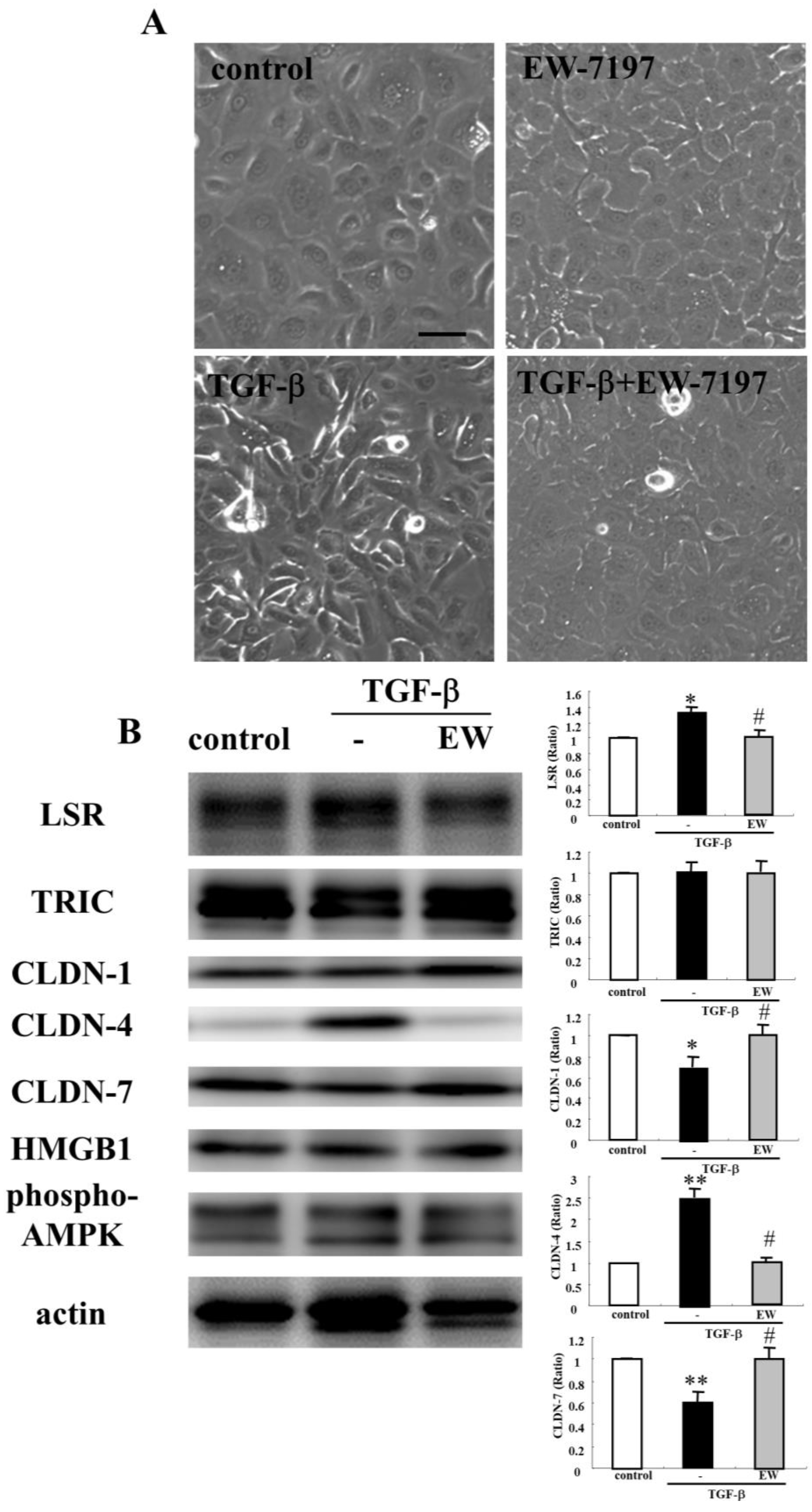
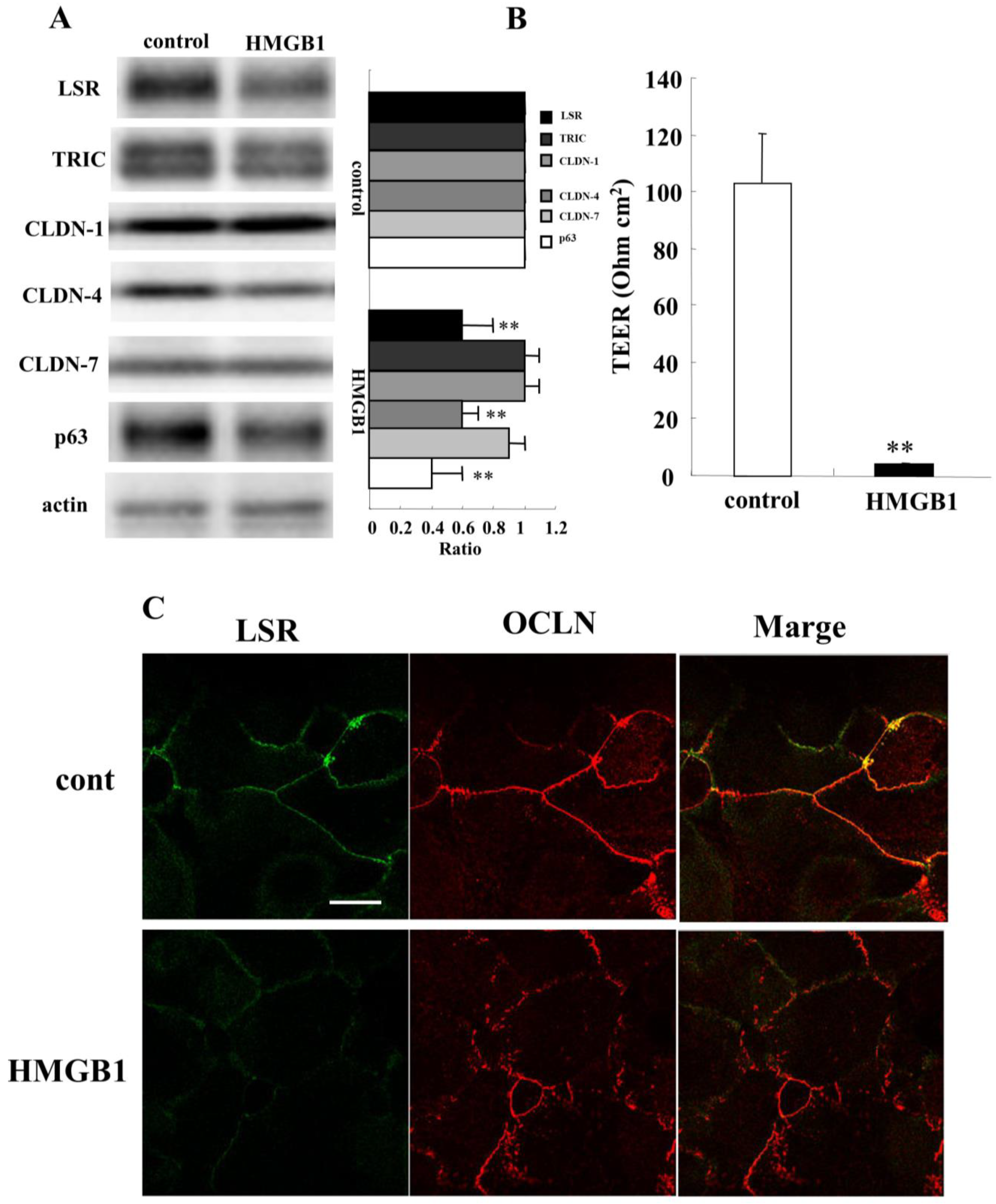
2.4. HMGB1 Decreases Angulin-1/LSR Expression and Epithelial Barrier Function in HNECs
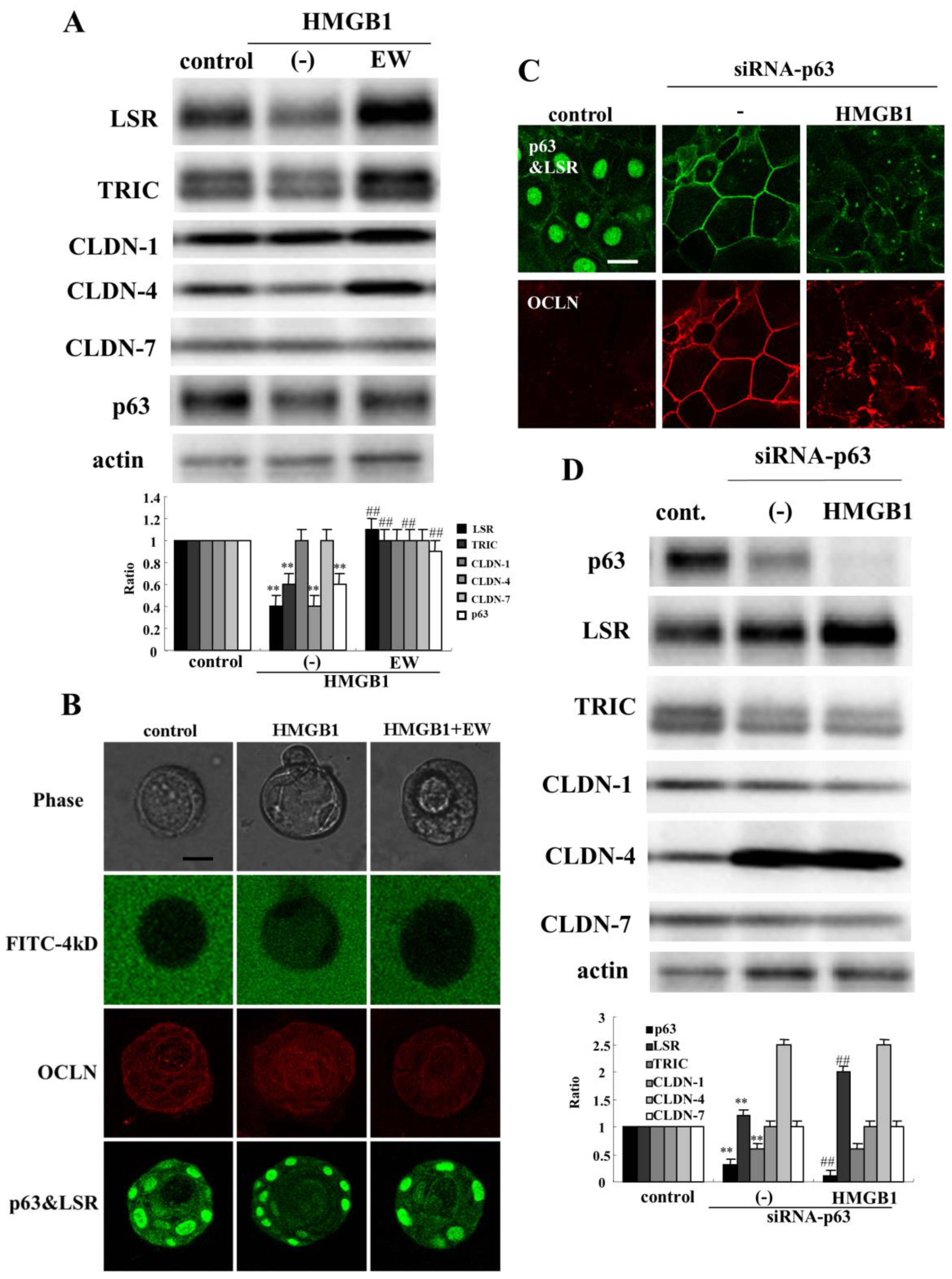
2.5. TGF-β Receptor Type 1 Inhibitor EW-7197 and Knockdown of p63 Suppressed the Effects of HMGB1 in HNECs
2.6. Knockdown of p63 Prevents Downregulation of Angulin-1/LSR Induced by HMGB1 in HNECs
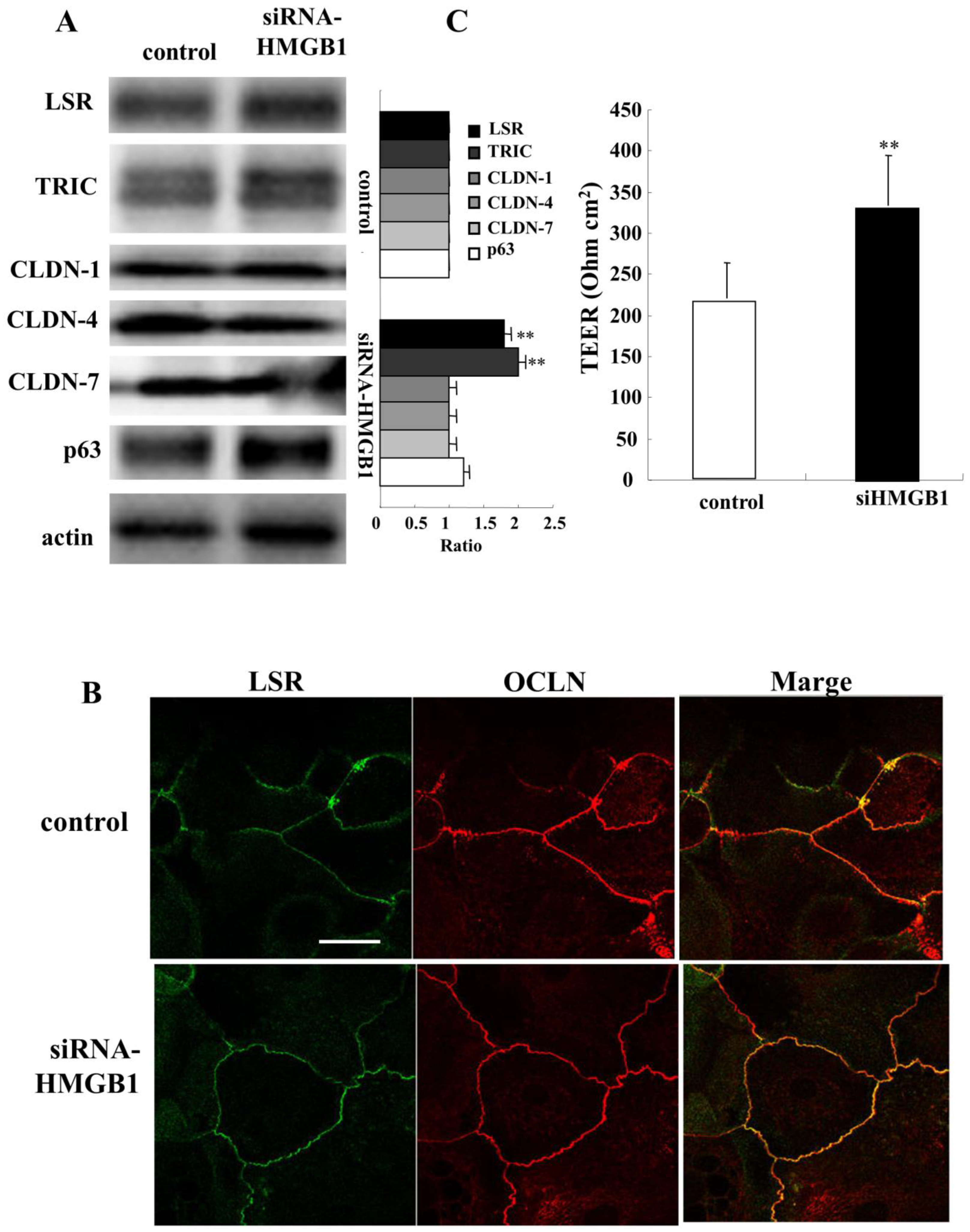
2.7. Knockdown of HMGB1 Induces Angulin-1/LSR and Enhances Epithelial Barrier Function in HNECs
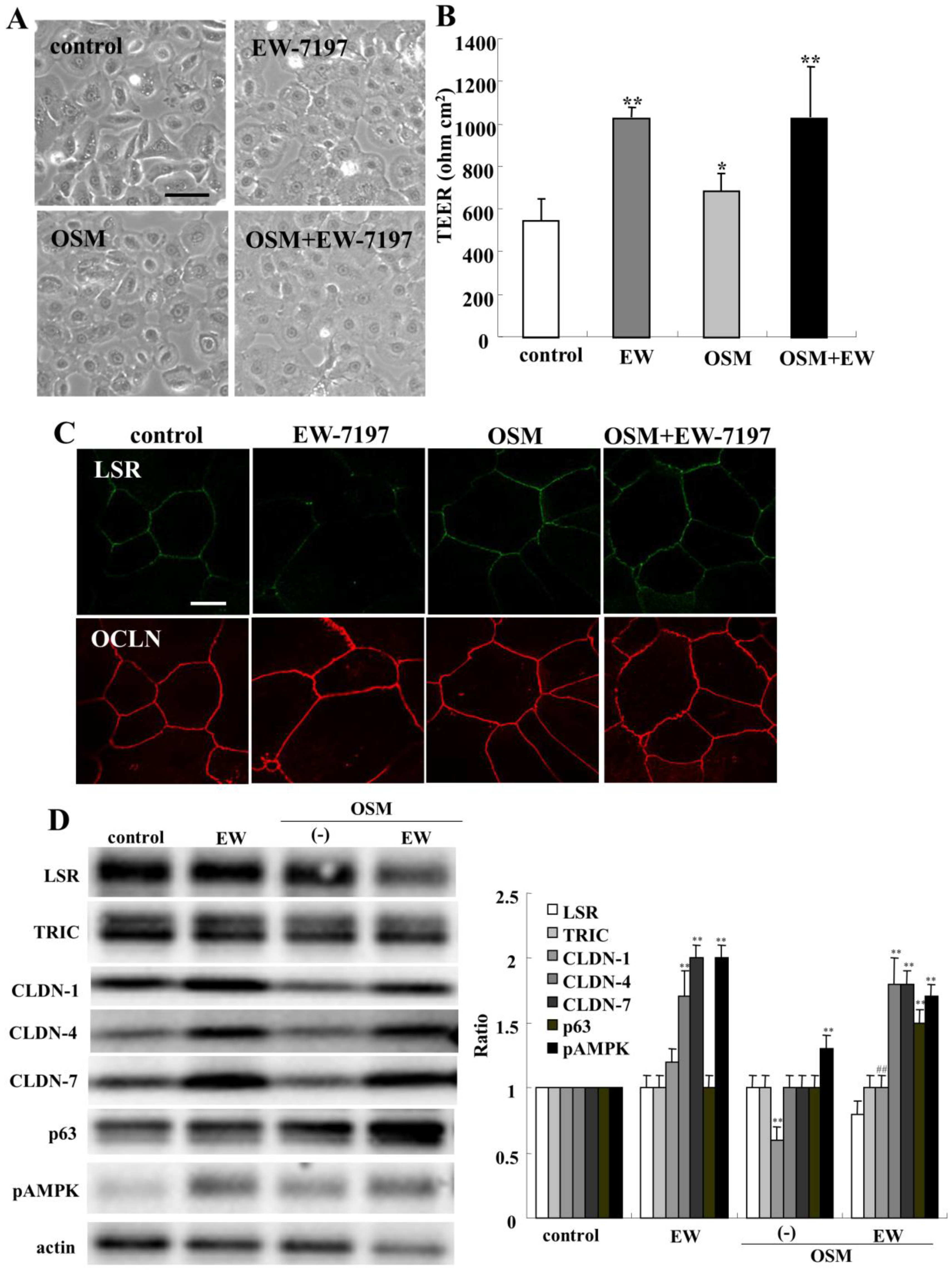
2.8. Effects of Oncostatin M on Epithelial Barrier and TJ Molecules in HNECs
2.9. Changes of COVID-19 Infection-Related Genes Induced by Treatment with HMGB1 and OSM in HNECs
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. Preparation of Angubindin-1
4.3. GeneChip Analysis
4.4. Immunohistochemical Analysis
4.5. Cell Culture and Treatments
4.6. 2.5-Dimensional (2.5D) Matrigel Culture
4.7. siRNA Experiment
4.8. RNA Isolation and Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
4.9. Western Blot Analysis
4.10. Immunocytochemistry
4.11. Measurement of Transepithelial Electrical Resistance (TEER)
4.12. Fluorescein Isothiocyanate (FITC) Permeability Assay
4.13. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethics Statement
Abbreviations
| HMGB1 | high-mobility group box 1 |
| bTJ | bicellular tight junction |
| tTJ | tricellular tight junction |
| LSR | lipolysis-stimulated lipoprotein receptor |
| TRIC | tricellulin |
| CLDNs | claudins |
| OCLN | occludin |
| ILDR | immunoglobulin-like domain-containing receptor |
| DAMP | danger associated molecular pattern |
| TLRs | toll-like receptors |
| RAGE | advanced glycation endproducts |
| AR | allergic rhinitis |
| CRS | chronic rhinosinusitis |
| ECRS | eosinophilic chronic rhinosinusitis |
| EMT | epithelial mesenchymal transition |
| hTERT | human telomerase reverse transcriptase |
| HNECs | human nasal epithelial cells |
References
- Kojima, T.; Go, M.; Takano, K.; Kurose, M.; Ohkuni, T.; Koizumi, J.; Kamekura, R.; Ogasawara, N.; Masaki, T.; Fuchimoto, J.; et al. Regulation of tight junctions in upper airway epithelium. Biomed. Res. Int. 2013, 2013, 947072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, K.; Kojima, T.; Sawada, N.; Himi, T. Role of tight junctions in signal transduction: An update. EXCLI J. 2014, 13, 1145–1162. [Google Scholar] [PubMed]
- Wynne, M.; Atkinson, C.; Schlosser, R.J.; Mulligan, J.K. Contribution of epithelial cell dysfunction to the pathogenesis of chronic rhinosinusitis with nasal polyps. Am. J. Rhinol. Allergy 2019, 33, 782–790. [Google Scholar] [CrossRef]
- Jiao, J.; Wang, C.; Zhang, L. Epithelial physical barrier defects in chronic rhinosinusitis. Expert Rev. Clin. Immunol. 2019, 15, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Siti Sarah, C.O.; Md Shukri, N.; Mohd Ashari, N.S.; Wong, K.K. Zonula occludens and nasal epithelial barrier integrity in allergic rhinitis. PeerJ 2020, 8, e9834. [Google Scholar] [CrossRef]
- Rinaldi, A.O.; Korsfeldt, A.; Ward, S.; Burla, D.; Dreher, A.; Gautschi, M.; Stolpe, B.; Tan, G.; Bersuch, E.; Melin, D.; et al. Electrical impedance spectroscopy for the characterization of skin barrier in atopic dermatitis. Allergy 2021, 1–14. [Google Scholar] [CrossRef]
- Inoue, H.; Akimoto, K.; Homma, T.; Tanaka, A.; Sagara, H. Airway epithelial dysfunction in asthma: Relevant to epidermal growth factor receptors and airway epithelial cells. J. Clin. Med. 2020, 9, 3698. [Google Scholar] [CrossRef]
- Sawada, N. Tight junction-related human diseases. Pathol. Int. 2013, 63, 1–12. [Google Scholar] [CrossRef]
- Furuse, M.; Itoh, M.; Hirase, T.; Nagafuchi, A.; Yonemura, S.; Tsukita, S.; Tsukita, S. Direct association of occludin with ZO-1 and its possible involvement in the localization of occludin at tight junctions. J. Cell. Biol. 1994, 127, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, U.; Schuetz, A. Structural features of tight-Junction proteins. Int. J. Mol. Sci. 2019, 20, 6020. [Google Scholar] [CrossRef] [Green Version]
- Higashi, T.; Tokuda, S.; Kitajiri, S.; Masuda, S.; Nakamura, H.; Oda, Y.; Furuse, M. Analysis of the ‘angulin’ proteins LSR, ILDR1 and ILDR2-tricellulin recruitment, epithelial barrier function and implication in deafness pathogenesis. J. Cell Sci. 2013, 126, 966–977. [Google Scholar] [CrossRef] [Green Version]
- Konno, T.; Kohno, T.; Kikuchi, S.; Shimada, H.; Satohisa, S.; Saito, T.; Kondoh, M.; Kojima, T. Epithelial barrier dysfunction and cell migration induction via JNK/cofilin/actin by angubindin-1. Tissue Barriers 2020, 8, 1695475. [Google Scholar] [CrossRef]
- Shimada, H.; Abe, S.; Kohno, T.; Satohisa, S.; Konno, T.; Takahashi, S.; Hatakeyama, T.; Arimoto, C.; Kakuki, T.; Kaneko, Y.; et al. Loss of tricellular tight junction protein LSR promotes cell invasion and migration via upregulation of TEAD1/AREG in human endometrial cancer. Sci. Rep. 2017, 7, 37049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohno, T.; Konno, T.; Kojima, T. Role of tricellular tight junction protein lipolysis-stimulated lipoprotein receptor (LSR) in cancer cells. Int. J. Mol. Sci. 2019, 20, 3555. [Google Scholar] [CrossRef] [Green Version]
- Nagahama, M.; Umezaki, M.; Oda, M.; Kobayashi, K.; Tone, S.; Suda, T.; Ishidoh, K.; Sakurai, J. Clostridium perfringens iota-toxin b induces rapid cell necrosis. Infect. Immun. 2011, 79, 4353–4360. [Google Scholar] [CrossRef] [Green Version]
- Krug, S.M.; Hayaishi, T.; Iguchi, D.; Watari, A.; Takahashi, A.; Fromm, M.; Nagahama, M.; Takeda, H.; Okada, Y.; Sawasaki, T.; et al. Angubindin-1, a novel paracellular absorption enhancer acting at the tricellular tight junction. J. Control. Release 2017, 260, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zeniya, S.; Kuwahara, H.; Daizo, K.; Watari, A.; Kondoh, M.; Yoshida-Tanaka, K.; Kaburagi, H.; Asada, K.; Nagata, T.; Nagahama, M.; et al. Angubindin-1 opens the blood-brain barrier in vivo for delivery of antisense oligonucleotide to the central nervous system. J. Control. Release 2018, 283, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.; Tachibana, K.; Krug, S.M.; Kunisawa, J.; Fromm, M.; Kondoh, M. Potential for tight junction protein-directed drug development using claudin binders and angubindin-1. Int. J. Mol. Sci. 2019, 20, 4016. [Google Scholar] [CrossRef] [Green Version]
- Kyuno, T.; Kyuno, D.; Kohno, T.; Konno, T.; Kikuchi, S.; Arimoto, C.; Yamaguchi, H.; Imamura, M.; Kimura, Y.; Kondoh, M.; et al. Tricellular tight junction protein LSR/angulin-1 contributes to the epithelial barrier and malignancy in human pancreatic cancer cell line. Histochem. Cell. Biol. 2020, 153, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Nishibori, M. The Role of high mobility group box-1 in epileptogenesis. Acta Med. Okayama 2019, 73, 383–386. [Google Scholar]
- Yun, B.H.; Kim, S.; Chon, S.J.; Kim, G.H.; Choi, Y.S.; Cho, S.; Lee, B.S.; Seo, S.K. High mobility group box-1 promotes inflammation in endometriotic stromal cells through Toll-like receptor 4/nuclear factor-kappa B. Am. J. Transl. Res. 2021, 13, 1400–1410. [Google Scholar] [PubMed]
- Watanabe, H.; Son, M. The immune tolerance role of the HMGB1-RAGE axis. Cells 2021, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Liu, Q.; Zhao, J.; Tang, H.; Sun, J. SIRT1 attenuates murine allergic rhinitis by downregulated HMGB 1/TLR4 pathway. Scand. J. Immunol. 2018, 87, e12667. [Google Scholar] [CrossRef] [Green Version]
- Bellussi, L.M.; Cocca, S.; Passali, G.C.; Passali, D. HMGB1 in the pathogenesis of nasal inflammatory diseases and its inhibition as new therapeutic approach: A review from the literature. Int. Arch. Otorhinolaryngol. 2017, 21, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.R.; Xu, J.; Lee, S.; Yeon, S.H.; Park, S.K.; Rha, K.S.; Kim, Y.M. Chloroquine treatment suppresses mucosal inflammation in a mouse model of eosinophilic chronic rhinosinusitis. Allergy Asthma Immunol. Res. 2020, 12, 994–1011. [Google Scholar] [CrossRef]
- Zheng, J.; Wei, X.; Zhan, J.B.; Jiang, H.Y. High mobility group box1 contributes to hypoxia-induced barrier dysfunction of nasal epithelial cells. J. Clin. Otorhinolaryngol. Head Neck Surg. 2017, 31, 1178–1181. [Google Scholar]
- Chen, R.; Huang, Y.; Quan, J.; Liu, J.; Wang, H.; Billiar, T.R.; Lotze, M.T.; Zeh, H.J.; Kang, R.; Tang, D. HMGB1 as a potential biomarker and therapeutic target for severe COVID-19. Heliyon 2020, 6, e05672. [Google Scholar] [CrossRef]
- Togami, K.; Yamaguchi, K.; Chono, S.; Tada, H. Evaluation of permeability alteration and epithelial-mesenchymal transition induced by transforming growth factor-β1 in A549, NCI-H441, and Calu-3 cells: Development of an in vitro model of respiratory epithelial cells in idiopathic pulmonary fibrosis. J. Pharmacol. Toxicol. Methods 2017, 86, 19–27. [Google Scholar] [CrossRef]
- Ohta, H.; Chiba, S.; Ebina, M.; Furuse, M.; Nukiwa, T. Altered expression of tight junction molecules in alveolar septa in lung injury and fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L193–L205. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.; Sun, J.; You, W.; Wei, Y.; Tian, H.; Jiang, S. Glycyrrhizin suppresses epithelial-mesenchymal transition by inhibiting high-mobility group box1 via the TGF-β1/Smad2/3 pathway in lung epithelial cells. PeerJ 2020, 8, e8514. [Google Scholar] [CrossRef] [Green Version]
- Kodera, Y.; Kohno, T.; Konno, T.; Arai, W.; Tsujiwaki, M.; Shindo, Y.; Chiba, H.; Miyakawa, M.; Tanaka, H.; Sakuma, Y.; et al. HMGB1 enhances epithelial permeability via p63/TGF-β signaling in lung and terminal bronchial epithelial cells. Tissue Barriers 2020, 8, 1805997. [Google Scholar] [CrossRef]
- Miyakawa, M.; Konno, T.; Kohno, T.; Kikuchi, S.; Tanaka, H.; Kojima, T. Increase in epithelial permeability and cell metabolism by high mobility group box 1, inflammatory cytokines and TPEN in Caco-2 cells as a novel model of inflammatory bowel disease. Int. J. Mol. Sci. 2020, 21, 8434. [Google Scholar] [CrossRef]
- Binabaj, M.M.; Asgharzadeh, F.; Avan, A.; Rahmani, F.; Soleimani, A.; Parizadeh, M.R.; Ferns, G.A.; Ryzhikov, M.; Khazaei, M.; Hassanian, S.M. EW-7197 prevents ulcerative colitis-associated fibrosis and inflammation. J. Cell. Physiol. 2019, 234, 11654–11661. [Google Scholar] [CrossRef] [PubMed]
- Koster, M.I.; Roop, D.R. The role of p63 in development and differentiation of the epidermis. J. Dermatol. Sci. 2004, 34, 3–9. [Google Scholar] [CrossRef]
- Kaneko, Y.; Kohno, T.; Kakuki, T.; Takano, K.-I.; Ogasawara, N.; Miyata, R.; Kikuchi, S.; Konno, T.; Ohkuni, T.; Yajima, R.; et al. The role of transcriptional factor p63 in regulation of epithelial barrier and ciliogenesis of human nasal epithelial cells. Sci. Rep. 2017, 7, 10935. [Google Scholar] [CrossRef] [PubMed]
- Kurose, M.; Kojima, T.; Koizumi, J.; Kamekura, R.; Ninomiya, T.; Murata, M.; Ichimiya, S.; Osanai, M.; Chiba, H.; Himi, T.; et al. Induction of claudins in passaged hTERT-transfected human nasal epithelial cells with an extended life span. Cell Tissue Res. 2007, 330, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Konno, T.; Kohno, T.; Kakuki, T.; Miyata, R.; Ohkuni, T.; Kakiuchi, A.; Yajima, R.; Ohwada, K.; Kurose, M.; et al. Induction of airway progenitor cells via p63 and KLF11 by Rho-kinase inhibitor Y27632 in hTERT-human nasal epithelial cells. Am. J. Transl. Res. 2019, 11, 599–611. [Google Scholar] [PubMed]
- Miyata, R.; Nomura, K.; Kakuki, T.; Takano, K.; Kohno, T.; Konno, T.; Sawada, N.; Himi, T.; Kojima, T. Irsogladine maleate regulates gap junctional intercellular communication-dependent epithelial barrier in human nasal epithelial cells. J. Membr. Biol. 2015, 248, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Dzaman, K.; Zagor, M.; Molinska-Glura, M.; Krzeski, A. High motility group box 1 (HMGB1) protein and its receptor for advanced glycation end products (RAGE) expression in chronic rhinosinusitis without nasal polyps. Folia Histochem. Cytobiol. 2015, 53, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Chen, S.; Zhong, G.; Kong, W.; Wang, Y. Agonist of PPAR-γ reduced epithelial-mesenchymal transition in eosinophilic chronic rhinosinusitis with nasal polyps via inhibition of high mobility group box 1. Int. J. Med. Sci. 2019, 16, 1631–1641. [Google Scholar] [CrossRef]
- Kodera, Y.; Chiba, H.; Konno, T.; Kohno, T.; Takahashi, H.; Kojima, T. HMGB1-downregulated angulin-1/LSR induces epithelial barrier disruption via claudin-2 and cellular metabolism via AMPK in airway epithelial Calu-3 cells. Biochem. Biophys. Res. Commun. 2020, 527, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Pothoven, K.L.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Harris, K.E.; Biyasheva, A.; Welch, K.; Shintani-Smith, S.; Conley, D.B.; Liu, M.C.; et al. Neutrophils are a major source of the epithelial barrier disrupting cytokine oncostatin M in patients with mucosal airways disease. J. Allergy Clin. Immunol. 2017, 139, 1966–1978. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Oda, Y.; Sasaki, H.; Ikenouchi, J.; Higashi, T.; Akashi, M.; Nishi, E.; Furuse, M. LSR defines cell corners for tricellular tight junction formation in epithelial cells. J. Cell Sci. 2011, 124, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Min, H.J.; Choe, J.W.; Kim, K.S.; Yoon, J.H.; Kim, C.H. High-mobility group box 1 protein induces epithelialmesenchymal transition in upper airway epithelial cells. Rhinology 2020, 58, 495–505. [Google Scholar] [CrossRef]
- Yoon, S.H.; Kim, K.Y.; Wang, Z.; Park, J.H.; Bae, S.M.; Kim, S.Y.; Song, H.Y.; Jeon, J.Y. EW-7197, a transforming growth factor-beta type I receptor kinase inhibitor, ameliorates acquired lymphedema in a mouse tail model. Lymphat. Res. Biol. 2020, 18, 433–438. [Google Scholar] [CrossRef]
- Ikai, K.; Sakai, M.; Minagi, H.O.; Gojo, N.; Sakai, T. ΔNp63 is upregulated during salivary gland regeneration following duct ligation and irradiation in mice. FEBS Lett. 2020, 594, 3216–3226. [Google Scholar] [CrossRef]
- Li, C.W.; Shi, L.; Zhang, K.K.; Li, T.Y.; Lin, Z.B.; Lim, M.K.; McKeon, F.; Xian, W.; Wang, D.Y. Role of p63/p73 in epithelial remodeling and their response to steroid treatment in nasal polyposis. J. Allergy Clin. Immunol. 2011, 127, e1–e2. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.P.; Utomo, L.; Rudnik-Jansen, I.; Du, J.; Zuithoff, N.P.A.; Krouwels, A.; van Osch, G.J.V.M.; Creemers, L.B. Association between oncostatin m expression and inflammatory phenotype in experimental arthritis models and osteoarthritis patients. Cells 2021, 10, 508. [Google Scholar] [CrossRef]
- Tian, T.; Zi, X.; Peng, Y.; Wang, Z.; Hong, H.; Yan, Y.; Guan, W.; Tan, K.S.; Liu, J.; Ong, H.H.; et al. H3N2 influenza virus infection enhances oncostatin M expression in human nasal epithelium. Exp. Cell Res. 2018, 371, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Pothoven, K.L.; Norton, J.E.; Hulse, K.E.; Suh, L.A.; Carter, R.G.; Rocci, E.; Harris, K.E.; Shintani-Smith, S.; Conley, D.B.; Chandra, R.K.; et al. Oncostatin M promotes mucosal epithelial barrier dysfunction, and its expression is increased in patients with eosinophilic mucosal disease. J. Allergy Clin. Immunol. 2015, 136, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, C.P.; Fernandes, D.E.; Casimiro, F.; da Mata, G.F.; Passos, M.T.; Varela, P.; Mastroianni-Kirsztajn, G.; Pesquero, J.B. Cathepsin L in COVID-19: From pharmacological evidences to genetics. Front. Cell. Infect. Microbiol. 2020, 10, 589505. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prasoon, P.; Kumari, C.; Pareek, V.; Faiq, M.A.; Narayan, R.K.; Kulandhasamy, M.; Kant, K. SARS-CoV-2-specific virulence factors in COVID-19. J. Med. Virol. 2021, 93, 1343–1350. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Gene | Control | HMGB1 + OSM | Fold (HMGB1 + OSM/Control) |
|---|---|---|---|---|
| NM_0218042 | ACE2 | 3 | 3 | 1.06 |
| NM_1536093 | TMPRSS6 | 1 | 3 | 2.75 |
| NM_001289823.1 | FURIN | 9 | 15 | 1.66 |
| XM_011518263.1 | CTSL | 438 | 881 | 2.01 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| angulin-1/LSR | CAGGACCTCAGAAGCCCCTGA | AACAGCACTTGTCTGGGCAGC |
| tricellulin | AGGCAGCTCGGAGACATAGA | TCACAGGGTATTTTGCCACA |
| claudin-1 | AACGCGGGGCTGCAGCTGTTG | GGATAGGGCCTTGGTGTTGGGT |
| claudin-4 | AGCCTTCCAGGTCCTCAACT | AGCAGCGAGTCGTACACCTT |
| claudin-7 | AGGCATAATTTTCATCGTGG | GAGTTGGACTTAGGGTAAGAGCG |
| G3PDH | ACCACAGTCCATGCCATCAC | TCCACCACCCTGTTGCTGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohwada, K.; Konno, T.; Kohno, T.; Nakano, M.; Ohkuni, T.; Miyata, R.; Kakuki, T.; Kondoh, M.; Takano, K.; Kojima, T. Effects of HMGB1 on Tricellular Tight Junctions via TGF-β Signaling in Human Nasal Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 8390. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168390
Ohwada K, Konno T, Kohno T, Nakano M, Ohkuni T, Miyata R, Kakuki T, Kondoh M, Takano K, Kojima T. Effects of HMGB1 on Tricellular Tight Junctions via TGF-β Signaling in Human Nasal Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(16):8390. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168390
Chicago/Turabian StyleOhwada, Kizuku, Takumi Konno, Takayuki Kohno, Masaya Nakano, Tsuyoshi Ohkuni, Ryo Miyata, Takuya Kakuki, Masuo Kondoh, Kenichi Takano, and Takashi Kojima. 2021. "Effects of HMGB1 on Tricellular Tight Junctions via TGF-β Signaling in Human Nasal Epithelial Cells" International Journal of Molecular Sciences 22, no. 16: 8390. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168390