
Somatostatin and Astroglial Involvement in the Human Limbic System in Alzheimer’s Disease

 , , , , , and
, , , , , and
Abstract
:
1. Introduction
2. Results
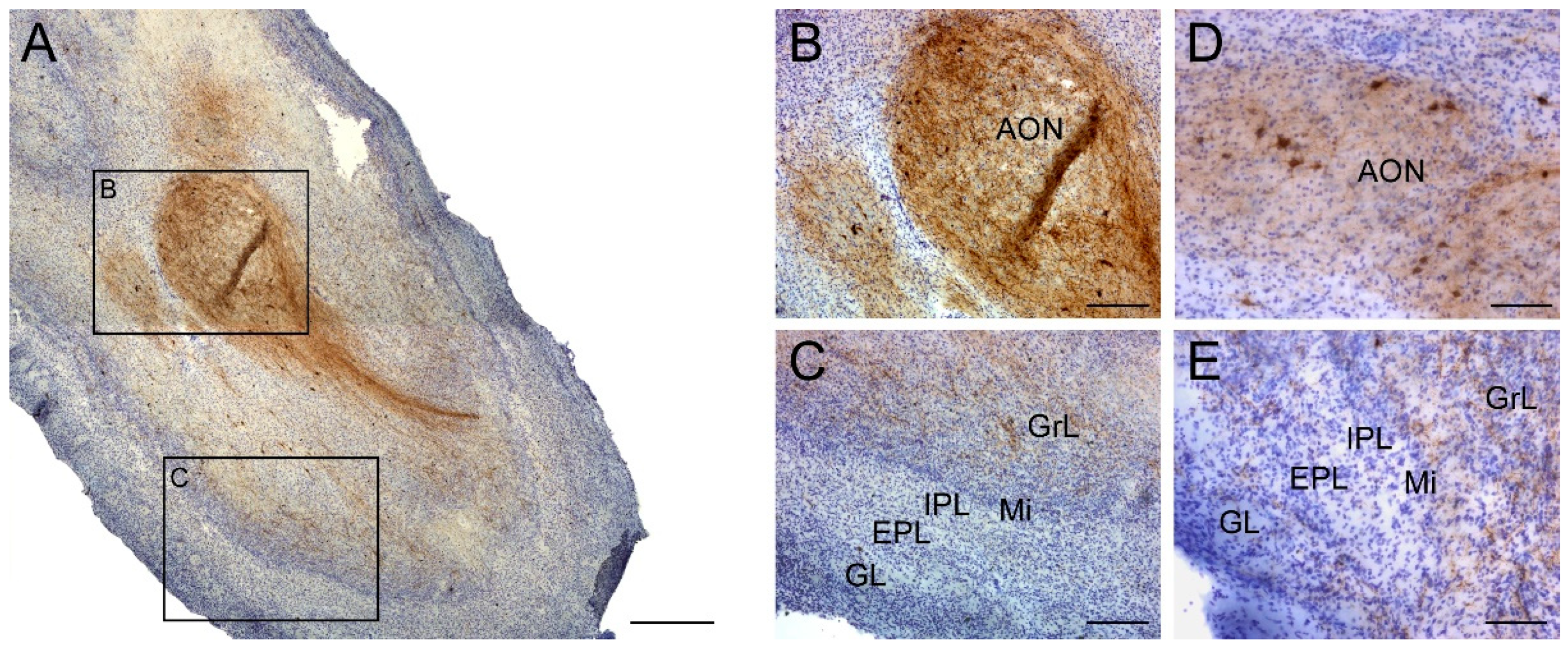
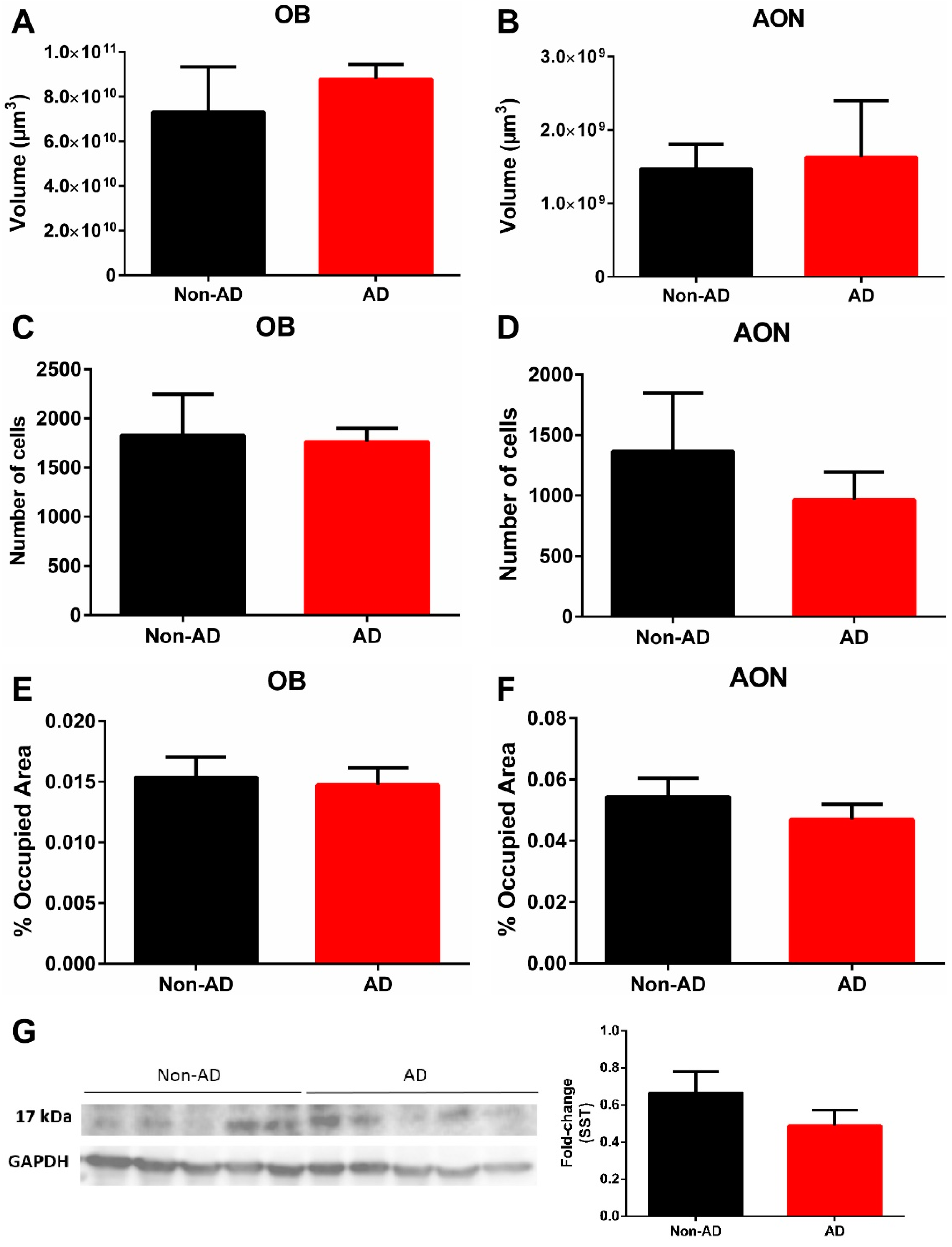
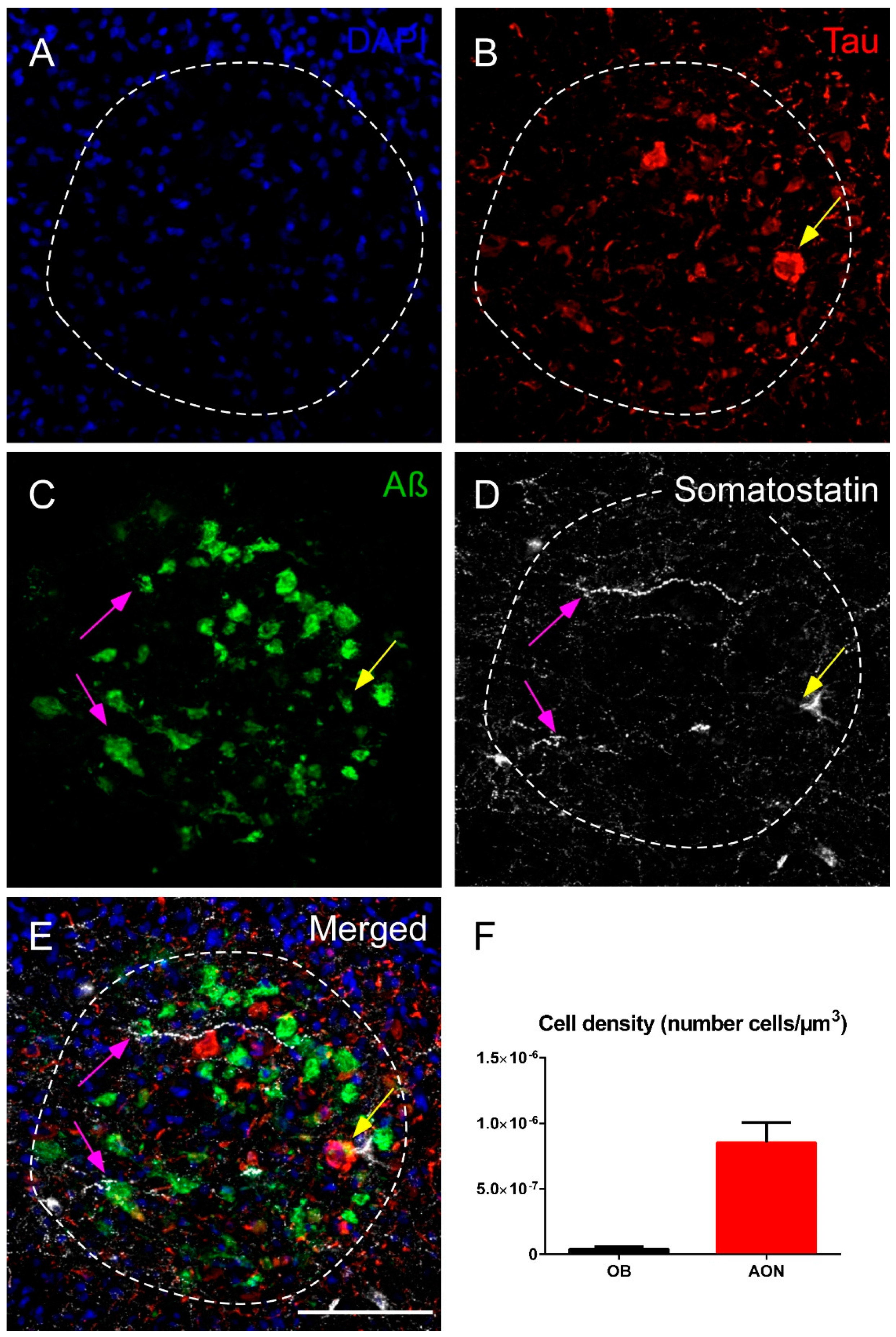
2.1. Somatostatin Expression in the Olfactory Bulb
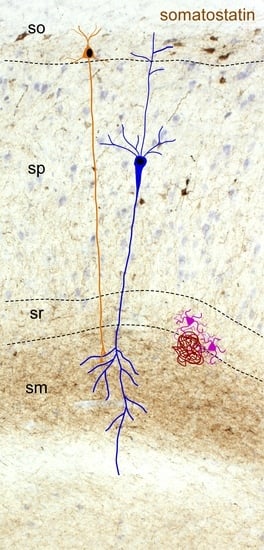
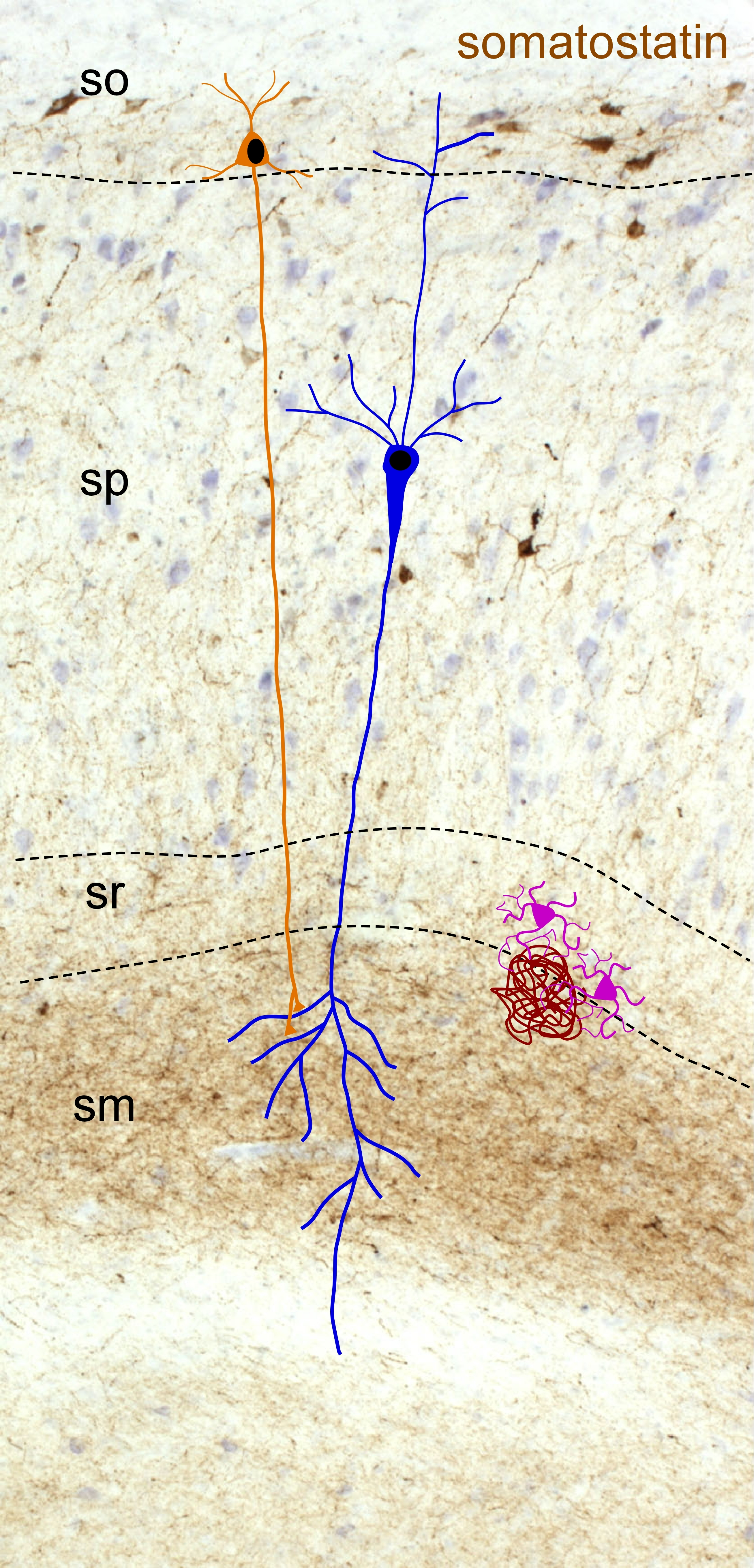
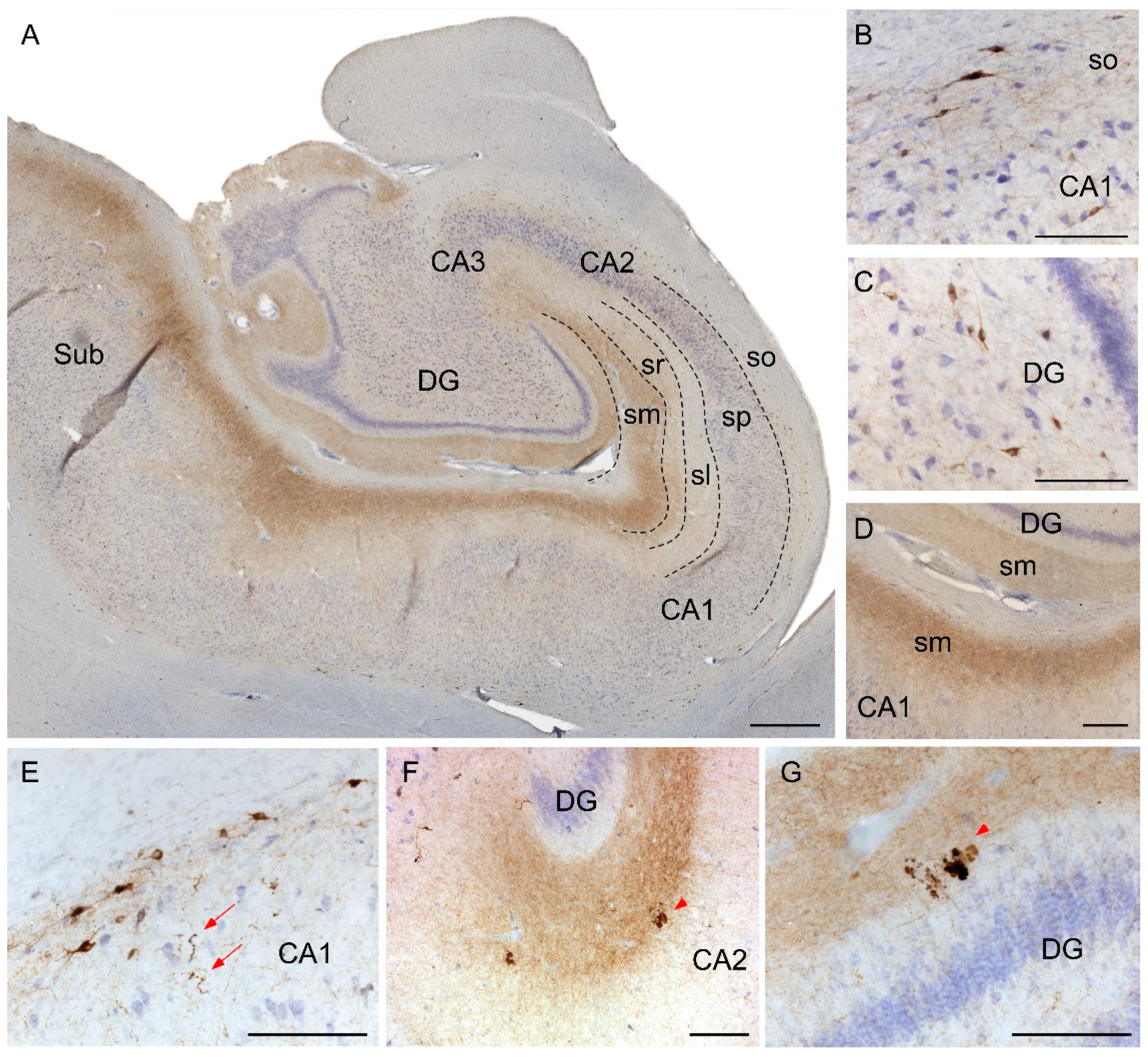
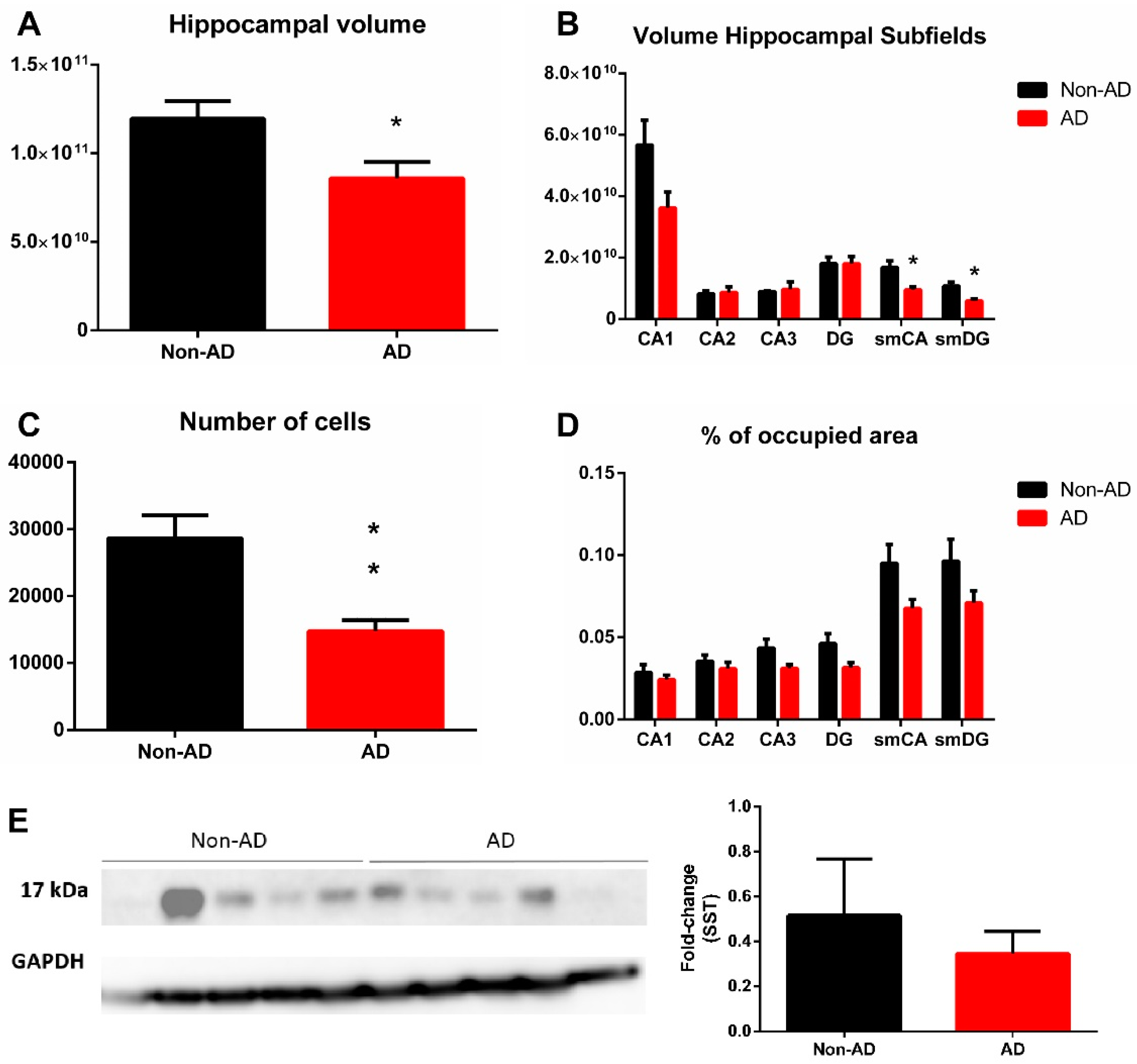
2.2. Somatostatin Expression in the Hippocampus
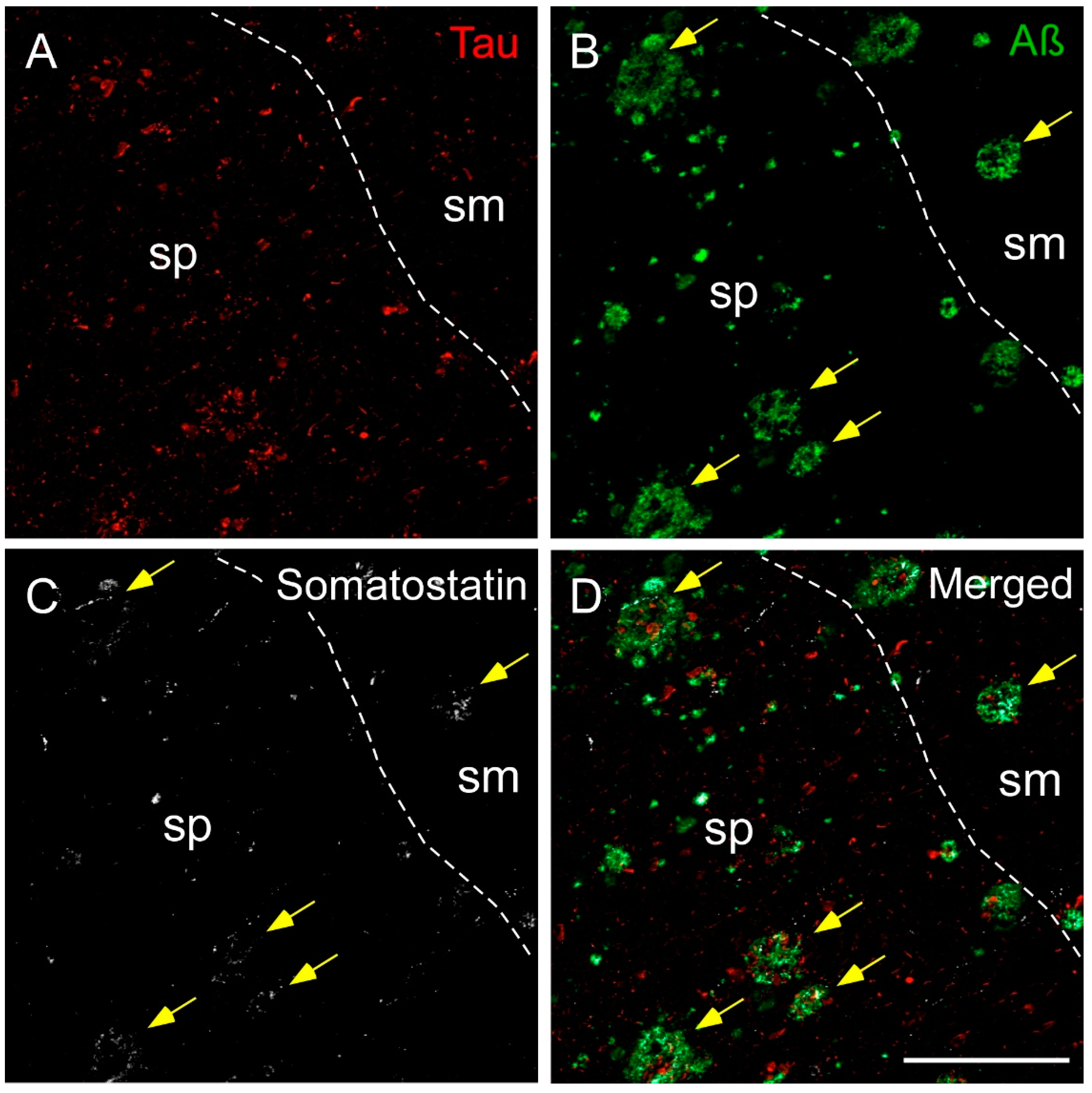
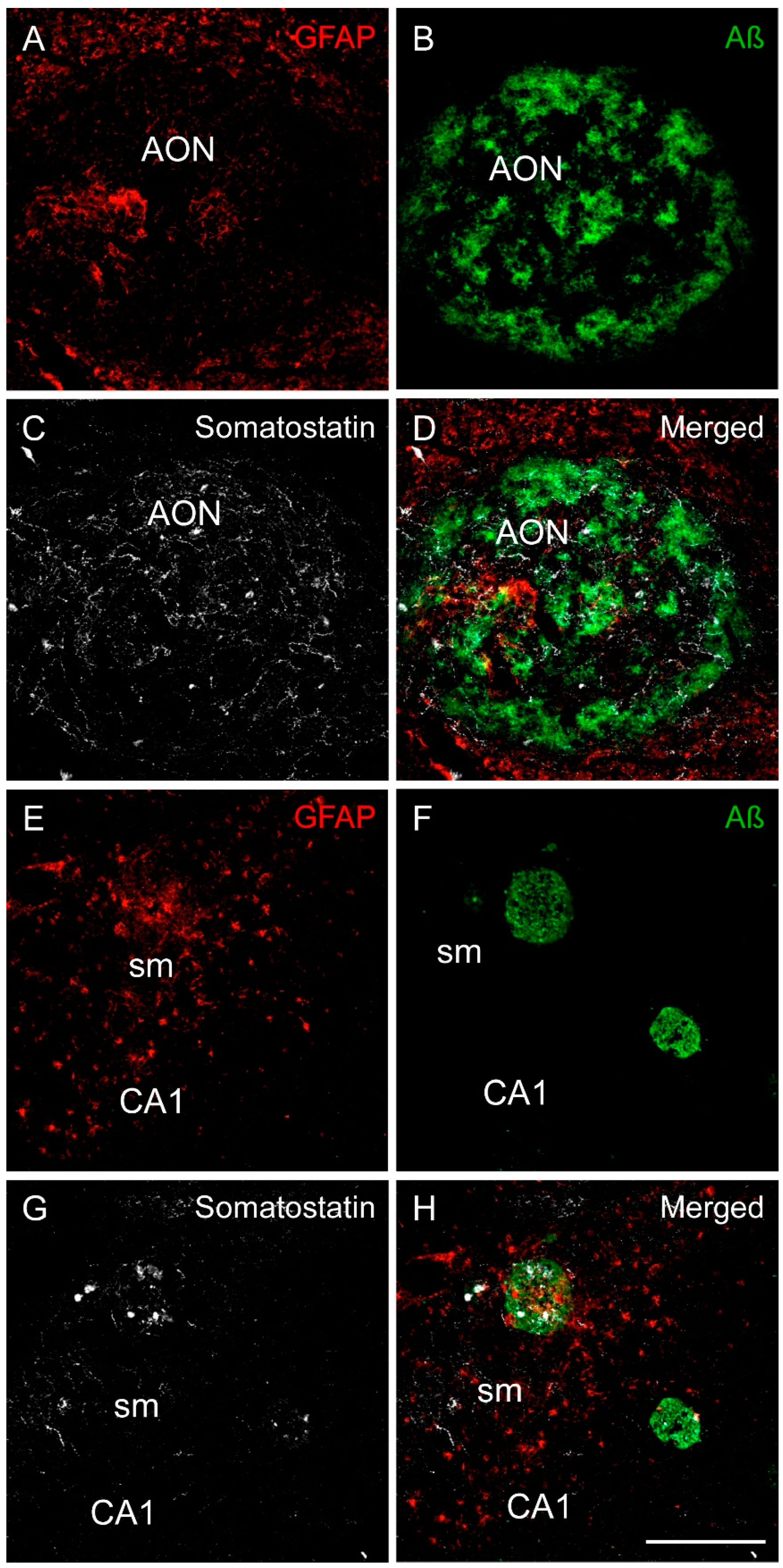
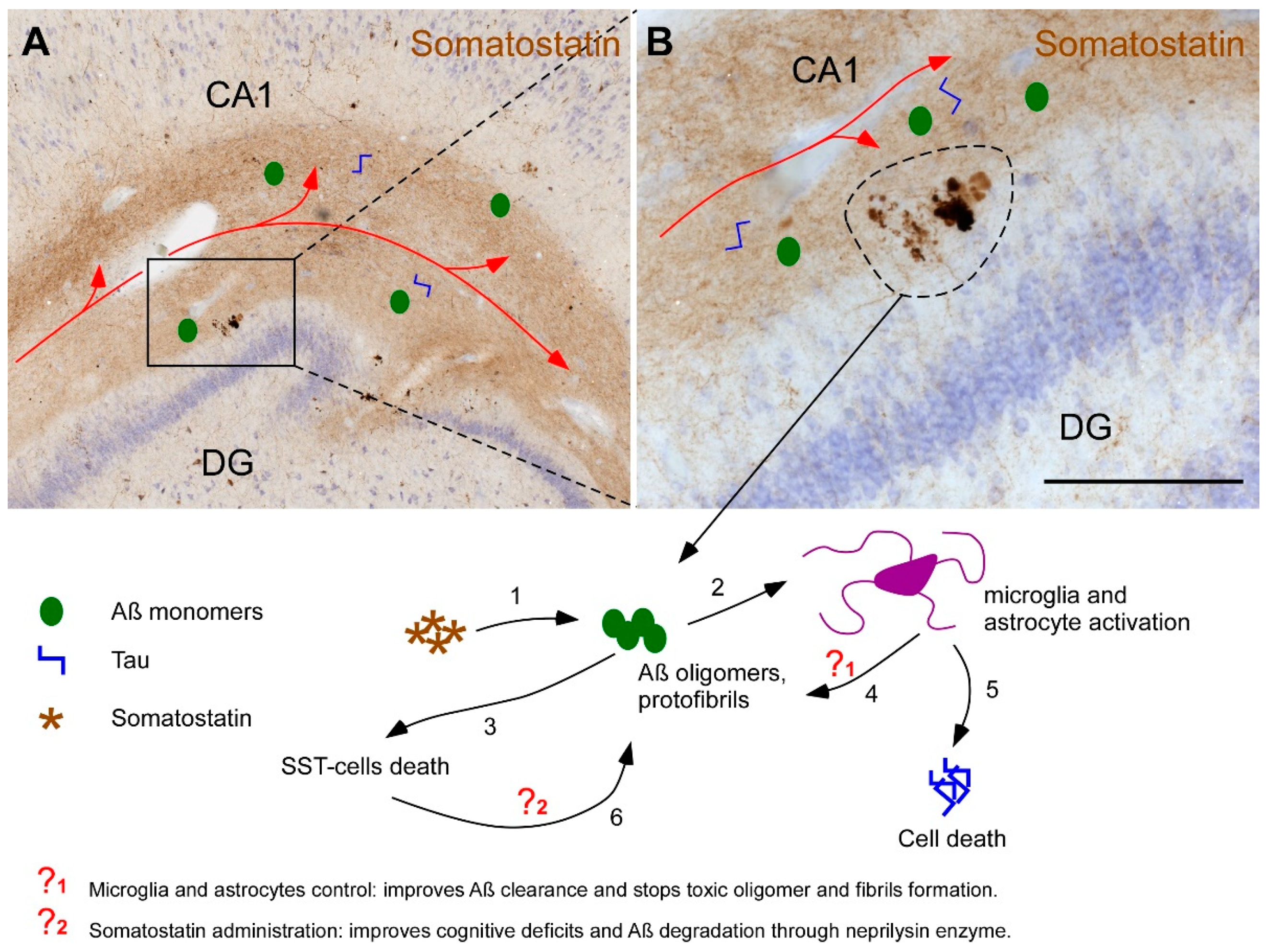
2.3. Somatostatin, Amyloid-β and Tau
2.4. Somatostatin, Amyloid-β and Astrocytes Distribution
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.2. Stereological Quantifications
4.3. Immunohistochemistry and Immunofluorescence
4.4. Western Blotting
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, R.S.; Arnold, S.E.; Schneider, J.A.; Boyle, P.A.; Buchman, A.S.; Bennett, D.A. Olfactory impairment in presymptomatic Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2009, 1170, 730–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballard, C.; Gauthier, S.; Corbett, A.; Brayne, C.; Aarsland, D.; Jones, E. Alzheimer’s disease. Lancet 2011, 377, 1019–1031. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Busche, M.A.; Hyman, B.T. Synergy between amyloid-beta and tau in Alzheimer’s disease. Nat. Neurosci. 2020, 23, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Attems, J.; Lintner, F.; Jellinger, K.A. Olfactory involvement in aging and Alzheimer’s disease: An autopsy study. J. Alzheimers Dis. 2005, 7, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C. Olfactory and other sensory impairments in Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 11–24. [Google Scholar] [CrossRef]
- Martel, G.; Dutar, P.; Epelbaum, J.; Viollet, C. Somatostatinergic systems: An update on brain functions in normal and pathological aging. Front. Endocrinol. 2012, 3, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavorska, I.; Wehr, M. Somatostatin-Expressing Inhibitory Interneurons in Cortical Circuits. Front. Neural Circuits 2016, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Viollet, C.; Lepousez, G.; Loudes, C.; Videau, C.; Simon, A.; Epelbaum, J. Somatostatinergic systems in brain: Networks and functions. Mol. Cell. Endocrinol. 2008, 286, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.H.; Yoon, J.; Lee, S.H. The role of neuropeptide somatostatin in the brain and its application in treating neurological disorders. Exp. Mol. Med. 2021, 53, 328–338. [Google Scholar] [CrossRef]
- Roberts, G.W.; Crow, T.J.; Polak, J.M. Location of neuronal tangles in somatostatin neurones in Alzheimer’s disease. Nature 1985, 314, 92–94. [Google Scholar] [CrossRef]
- Saito, T.; Iwata, N.; Tsubuki, S.; Takaki, Y.; Takano, J.; Huang, S.M.; Suemoto, T.; Higuchi, M.; Saido, T.C. Somatostatin regulates brain amyloid beta peptide Abeta42 through modulation of proteolytic degradation. Nat. Med. 2005, 11, 434–439. [Google Scholar] [CrossRef]
- Wang, H.; Muiznieks, L.D.; Ghosh, P.; Williams, D.; Solarski, M.; Fang, A.; Ruiz-Riquelme, A.; Pomes, R.; Watts, J.C.; Chakrabartty, A.; et al. Somatostatin binds to the human amyloid beta peptide and favors the formation of distinct oligomers. eLife 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Sanchez, D.; Ubeda-Banon, I.; Flores-Cuadrado, A.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Astillero-Lopez, V.; Martinez-Marcos, A. Somatostatin, Olfaction, and Neurodegeneration. Front. Neurosci. 2020, 14, 96. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Wan, C.Q.; Liu, Z. Astrocyte and Alzheimer’s disease. J. Neurol. 2017, 264, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Hickman, S.; Izzy, S.; Sen, P.; Morsett, L.; El Khoury, J. Microglia in neurodegeneration. Nat. Neurosci. 2018, 21, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Hussain, M.D.; Yan, L.J. Microglia, neuroinflammation, and beta-amyloid protein in Alzheimer’s disease. Int. J. Neurosci. 2014, 124, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Lackie, R.E.; Maciejewski, A.; Ostapchenko, V.G.; Marques-Lopes, J.; Choy, W.Y.; Duennwald, M.L.; Prado, V.F.; Prado, M.A.M. The Hsp70/Hsp90 Chaperone Machinery in Neurodegenerative Diseases. Front. Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreiser, R.P.; Wright, A.K.; Block, N.R.; Hollows, J.E.; Nguyen, L.T.; LeForte, K.; Mannini, B.; Vendruscolo, M.; Limbocker, R. Therapeutic Strategies to Reduce the Toxicity of Misfolded Protein Oligomers. Int. J. Mol. Sci. 2020, 21, 8651. [Google Scholar] [CrossRef] [PubMed]
- Li, J.G.; Chiu, J.; Ramanjulu, M.; Blass, B.E.; Pratico, D. A pharmacological chaperone improves memory by reducing Abeta and tau neuropathology in a mouse model with plaques and tangles. Mol. Neurodegener. 2020, 15, 1. [Google Scholar] [CrossRef]
- Beal, M.F.; Mazurek, M.F.; Svendsen, C.N.; Bird, E.D.; Martin, J.B. Widespread reduction of somatostatin-like immunoreactivity in the cerebral cortex in Alzheimer’s disease. Ann. Neurol. 1986, 20, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Winchester, R.L.; Martyn, K. Could Early Identification of Changes in Olfactory Function Be an Indicator of Preclinical Neurodegenerative Disease? A Systematic Review. Neurol. Ther. 2020, 9, 243–263. [Google Scholar] [CrossRef]
- Large, A.M.; Kunz, N.A.; Mielo, S.L.; Oswald, A.M. Inhibition by Somatostatin Interneurons in Olfactory Cortex. Front. Neural Circuits 2016, 10, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiz-Sanchez, D.; Ubeda-Banon, I.; de la Rosa-Prieto, C.; Argandona-Palacios, L.; Garcia-Munozguren, S.; Insausti, R.; Martinez-Marcos, A. Somatostatin, tau, and beta-amyloid within the anterior olfactory nucleus in Alzheimer disease. Exp. Neurol. 2010, 223, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Booker, S.A.; Vida, I. Morphological diversity and connectivity of hippocampal interneurons. Cell Tissue Res. 2018, 373, 619–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorens-Martin, M.; Blazquez-Llorca, L.; Benavides-Piccione, R.; Rabano, A.; Hernandez, F.; Avila, J.; DeFelipe, J. Selective alterations of neurons and circuits related to early memory loss in Alzheimer’s disease. Front. Neuroanat. 2014, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Drouet, V.; Wu, J.W.; Witter, M.P.; Small, S.A.; Clelland, C.; Duff, K. Trans-synaptic spread of tau pathology in vivo. PLoS ONE 2012, 7, e31302. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.A.; Devidze, N.; Verret, L.; Ho, K.; Halabisky, B.; Thwin, M.T.; Kim, D.; Hamto, P.; Lo, I.; Yu, G.Q.; et al. Transsynaptic progression of amyloid-beta-induced neuronal dysfunction within the entorhinal-hippocampal network. Neuron 2010, 68, 428–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, L.C.; Mittag, M.; Poll, S.; Steffen, J.; Wagner, J.; Geis, H.R.; Schwarz, I.; Schmidt, B.; Schwarz, M.K.; Remy, S.; et al. Dysfunction of Somatostatin-Positive Interneurons Associated with Memory Deficits in an Alzheimer’s Disease Model. Neuron 2016, 92, 114–125. [Google Scholar] [CrossRef] [Green Version]
- de Flores, R.; La Joie, R.; Chetelat, G. Structural imaging of hippocampal subfields in healthy aging and Alzheimer’s disease. Neuroscience 2015, 309, 29–50. [Google Scholar] [CrossRef] [Green Version]
- Hama, E.; Saido, T.C. Etiology of sporadic Alzheimer’s disease: Somatostatin, neprilysin, and amyloid beta peptide. Med. Hypotheses 2005, 65, 498–500. [Google Scholar] [CrossRef]
- Rofo, F.; Ugur Yilmaz, C.; Metzendorf, N.; Gustavsson, T.; Beretta, C.; Erlandsson, A.; Sehlin, D.; Syvanen, S.; Nilsson, P.; Hultqvist, G. Enhanced neprilysin-mediated degradation of hippocampal Abeta42 with a somatostatin peptide that enters the brain. Theranostics 2021, 11, 789–804. [Google Scholar] [CrossRef]
- Matos, M.; Bosson, A.; Riebe, I.; Reynell, C.; Vallee, J.; Laplante, I.; Panatier, A.; Robitaille, R.; Lacaille, J.C. Astrocytes detect and upregulate transmission at inhibitory synapses of somatostatin interneurons onto pyramidal cells. Nat. Commun. 2018, 9, 4254. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Kam, T.I.; Lee, S.; Park, H.; Oh, Y.; Kwon, S.H.; Song, J.J.; Kim, D.; Kim, H.; Jhaldiyal, A.; et al. Blocking microglial activation of reactive astrocytes is neuroprotective in models of Alzheimer’s disease. Acta Neuropathol. Commun. 2021, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Olabarria, M.; Noristani, H.N.; Yeh, C.Y.; Rodriguez, J.J. Astrocytes in Alzheimer’s disease. Neurotherapeutics 2010, 7, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Reyes, R.E.; Nava-Mesa, M.O.; Vargas-Sanchez, K.; Ariza-Salamanca, D.; Mora-Munoz, L. Involvement of Astrocytes in Alzheimer’s Disease from a Neuroinflammatory and Oxidative Stress Perspective. Front. Mol. Neurosci. 2017, 10, 427. [Google Scholar] [CrossRef] [Green Version]
- Arranz, A.M.; De Strooper, B. The role of astroglia in Alzheimer’s disease: Pathophysiology and clinical implications. Lancet Neurol. 2019, 18, 406–414. [Google Scholar] [CrossRef]
- Craft, S.; Asthana, S.; Newcomer, J.W.; Wilkinson, C.W.; Matos, I.T.; Baker, L.D.; Cherrier, M.; Lofgreen, C.; Latendresse, S.; Petrova, A.; et al. Enhancement of memory in Alzheimer disease with insulin and somatostatin, but not glucose. Arch. Gen. Psychiatry 1999, 56, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Mai, J.K.; Paxinos, G.; Voss, T. Atlas of the Human Brain; Academic Press: New York, NY, USA, 2007. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | DxAP | Sex | Age | PMD (h) | Brain Weight | Braak Stage | Disease Duration (Years) | Cause of Death | Treatment |
|---|---|---|---|---|---|---|---|---|---|
| 31 | AD | F | 90 | 12 | 920 | VI | - | Acute respiratory failure, respiratory infection and Alzheimer’s disease | Fresh-frozen |
| 32 | AD | F | 81 | 6 | 935 | VI | 15 | Cardiorespiratory arrest | Fresh-frozen |
| 33 | AD | F | 80 | 5 | 1060 | VI | 8.5 | Acute heart failure | Fresh-frozen |
| 34 | AD | M | 80 | 21 | 1300 | VI | - | Respiratory failure; bronchoaspiration | Fresh-frozen |
| 35 | AD | F | 76 | 11 | 900 | VI | - | Respiratory failure | Fresh-frozen |
| 36 | AD | F | 83 | 2 | 1000 | VI | 9 | Respiratory failure | Formalin-fixed |
| 37 | AD | F | 85 | 2 | 1150 | VI | 7 | Respiratory failure | Formalin-fixed |
| 38 | AD | M | 75 | 3 | 1050 | V | 4 | Multiorgan failure | Formalin-fixed |
| 39 | AD | F | 84 | 2 | 920 | V | 11 | - | Formalin-fixed |
| 40 | AD | M | 77 | 6 | 1060 | VI | 10 | Acute Respiratory infection | Formalin-fixed |
| 41 | Non-AD | F | 37 | 9 | 1200 | - | - | Septic Shock | Fresh-frozen |
| 42 | Non-AD | F | 83 | 7 | 1320 | - | - | - | Fresh-frozen |
| 43 | Non-AD | M | 68 | 4 | 1350 | - | - | Sepsis | Fresh-frozen |
| 44 | Non-AD | F | 82 | 4 | 800 | - | - | Cardiorespiratory arrest | Fresh-frozen |
| 45 | Non-AD | F | 71 | 7 | 975 | - | - | Respiratory failure | Fresh-frozen |
| 46 | Non-AD | F | 62 | 2 | 1050 | - | Multiorgan failure | Formalin-fixed | |
| 47 | Non-AD | M | 58 | 6 | 1500 | - | Cardiorespiratory arrest | Formalin-fixed | |
| 48 | Non-AD | M | 53 | 5 | 1300 | - | 2 | Respiratory failure | Formalin-fixed |
| 49 | Non-AD | M | 78 | 4 | 1100 | - | 1.5 | Respiratory failure | Formalin-fixed |
| 50 | Non-AD | M | 63 | 2 | 1400 | - | 1.5 | Respiratory failure | Formalin-fixed |
| Case | DxAP | Sex | Age | PMD (h) | Brain Weight | Braak Stage | Disease Duration (Years) | Cause of death | Treatment |
|---|---|---|---|---|---|---|---|---|---|
| 1 | AD | M | 78 | 5 | - | V | - | - | Fresh-frozen |
| 2 | AD | M | 85 | 3 | 1130 | VI | 4 | High digestive hemorrhage | Fresh-frozen |
| 3 | AD | M | 73 | 13 | 1290 | V | 2 | Cardiorespiratory arrest; severe sepsis respiratory focus | Fresh-frozen |
| 4 | AD | M | 82 | 4 | 1105 | V | 6 | Cardiorespiratory arrest; renal insufficiency | Fresh-frozen |
| 5 | AD | M | 75 | 6 | 1080 | VI | 10 | Respiratory infection (Bronchoaspiration) | Fresh-frozen |
| 6 | AD | F | 91 | 8 | - | V | nd | - | Fresh-frozen |
| 7 | AD | F | 80 | 5 | 1060 | VI | 8.5 | Acute heart failure | Fresh-frozen |
| 8 | AD | F | 79 | 15 | 1195 | V | 4 | Respiratory failure | Fresh-frozen |
| 9 | AD | F | 81 | 13 | 920 | VI | 20 | Multiorgan failure | Fresh-frozen |
| 10 | AD | F | 81 | 6 | 935 | VI | 15 | Cardiorespiratory arrest | Fresh-frozen |
| 11 | AD | F | 76 | 11 | - | VI | - | - | Formalin-fixed |
| 12 | AD | M | 69 | 2 | - | VI | - | Multiorgan failure | Formalin-fixed |
| 13 | AD | F | 67 | 4 | - | VI | - | Pneumonia (Bronchoaspiration) | Formalin-fixed |
| 14 | AD | M | 67 | 4 | - | VI | - | - | Formalin-fixed |
| 15 | AD | F | 78 | 8 | 970 | VI | - | - | Formalin-fixed |
| 16 | Non-AD | M | 68 | 4 | 1350 | - | - | Sepsis | Fresh-frozen |
| 17 | Non-AD | M | 88 | 0 | 1285 | - | - | - | Fresh-frozen |
| 18 | Non-AD | M | 40 | 5 | 1400 | - | - | - | Fresh-frozen |
| 19 | Non-AD | M | 46 | 3 | 610 | - | - | Respiratory infection | Fresh-frozen |
| 20 | Non-AD | M | 43 | 5 | 1412 | - | - | Septic shock secondary to pneumonia | Fresh-frozen |
| 21 | Non-AD | F | 49 | 2 | 1500 | - | - | Cardiorespiratory arrest | Fresh-frozen |
| 22 | Non-AD | F | 59 | 4 | 1500 | - | - | Respiratory failure | Fresh-frozen |
| 23 | Non-AD | F | 57 | 13 | 1335 | - | - | Respiratory failure | Fresh-frozen |
| 24 | Non-AD | F | 41 | 3 | 1295 | - | - | Renal insufficiency | Fresh-frozen |
| 25 | Non-AD | F | 71 | 7 | 975 | - | - | Cardiorespiratory arrest | Fresh-frozen |
| 26 | Non-AD | M | 77 | 10 | - | - | - | - | Formalin-fixed |
| 27 | Non-AD | F | 82 | 4 | - | - | - | Respiratory failure | Formalin-fixed |
| 28 | Non-AD | M | 68 | 4 | - | - | - | Cardiorespiratory arrest | Formalin-fixed |
| 29 | Non-AD | M | 83 | 13 | 1630 | - | - | Cardiorespiratory arrest | Formalin-fixed |
| 30 | Non-AD | M | 78 | 6 | - | I | - | - | Formalin-fixed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Rodriguez, M.; Astillero-Lopez, V.; Villanueva-Anguita, P.; Paya-Rodriguez, M.E.; Flores-Cuadrado, A.; Villar-Conde, S.; Ubeda-Banon, I.; Martinez-Marcos, A.; Saiz-Sanchez, D. Somatostatin and Astroglial Involvement in the Human Limbic System in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 8434. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168434
Gonzalez-Rodriguez M, Astillero-Lopez V, Villanueva-Anguita P, Paya-Rodriguez ME, Flores-Cuadrado A, Villar-Conde S, Ubeda-Banon I, Martinez-Marcos A, Saiz-Sanchez D. Somatostatin and Astroglial Involvement in the Human Limbic System in Alzheimer’s Disease. International Journal of Molecular Sciences. 2021; 22(16):8434. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168434
Chicago/Turabian StyleGonzalez-Rodriguez, Melania, Veronica Astillero-Lopez, Patricia Villanueva-Anguita, M. Eugenia Paya-Rodriguez, Alicia Flores-Cuadrado, Sandra Villar-Conde, Isabel Ubeda-Banon, Alino Martinez-Marcos, and Daniel Saiz-Sanchez. 2021. "Somatostatin and Astroglial Involvement in the Human Limbic System in Alzheimer’s Disease" International Journal of Molecular Sciences 22, no. 16: 8434. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168434