
CD82 and Gangliosides Tune CD81 Membrane Behavior

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
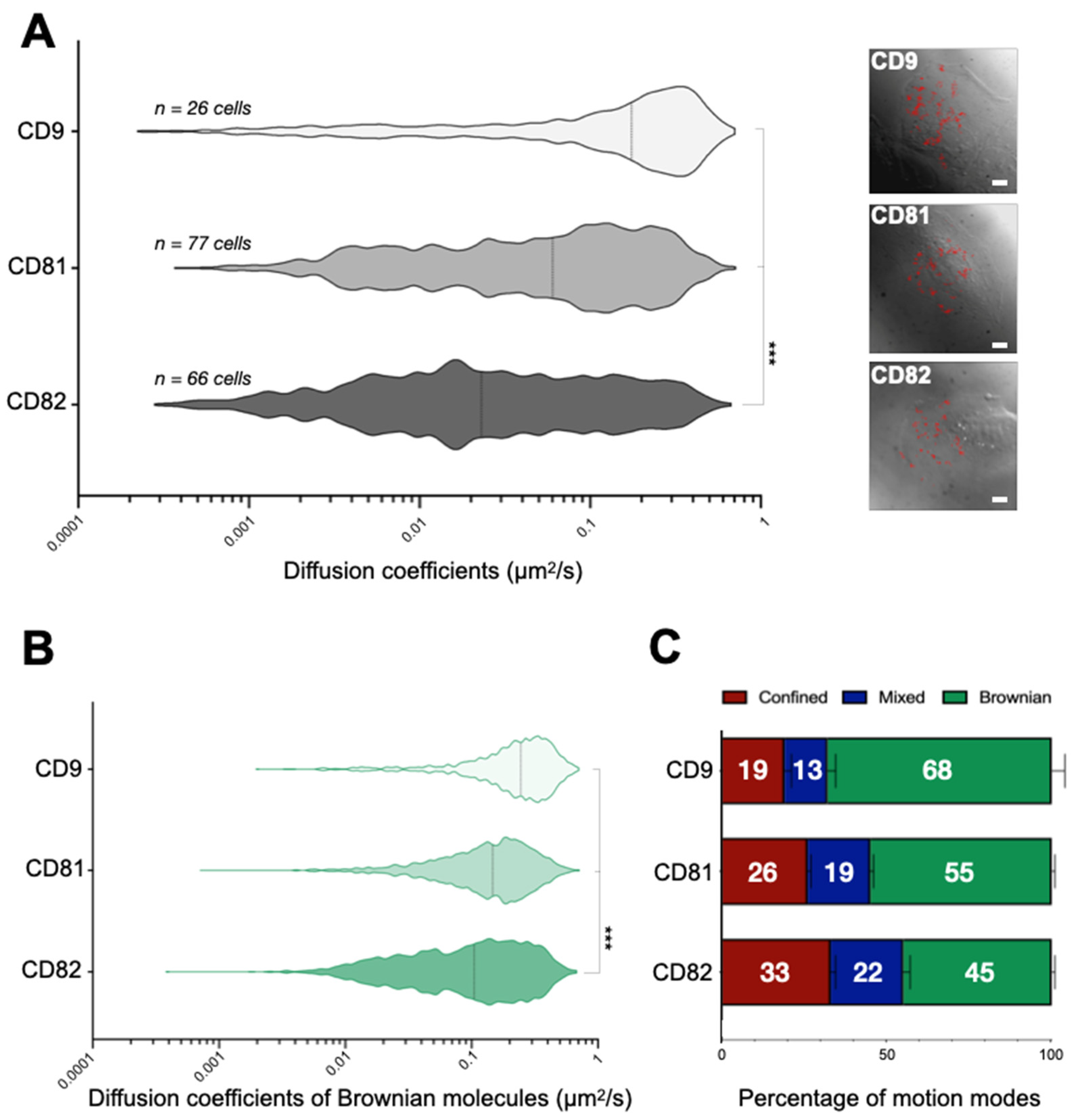
2.1. Tetraspanins Display Different Dynamics at the Plasma Membrane of HB2 Cells
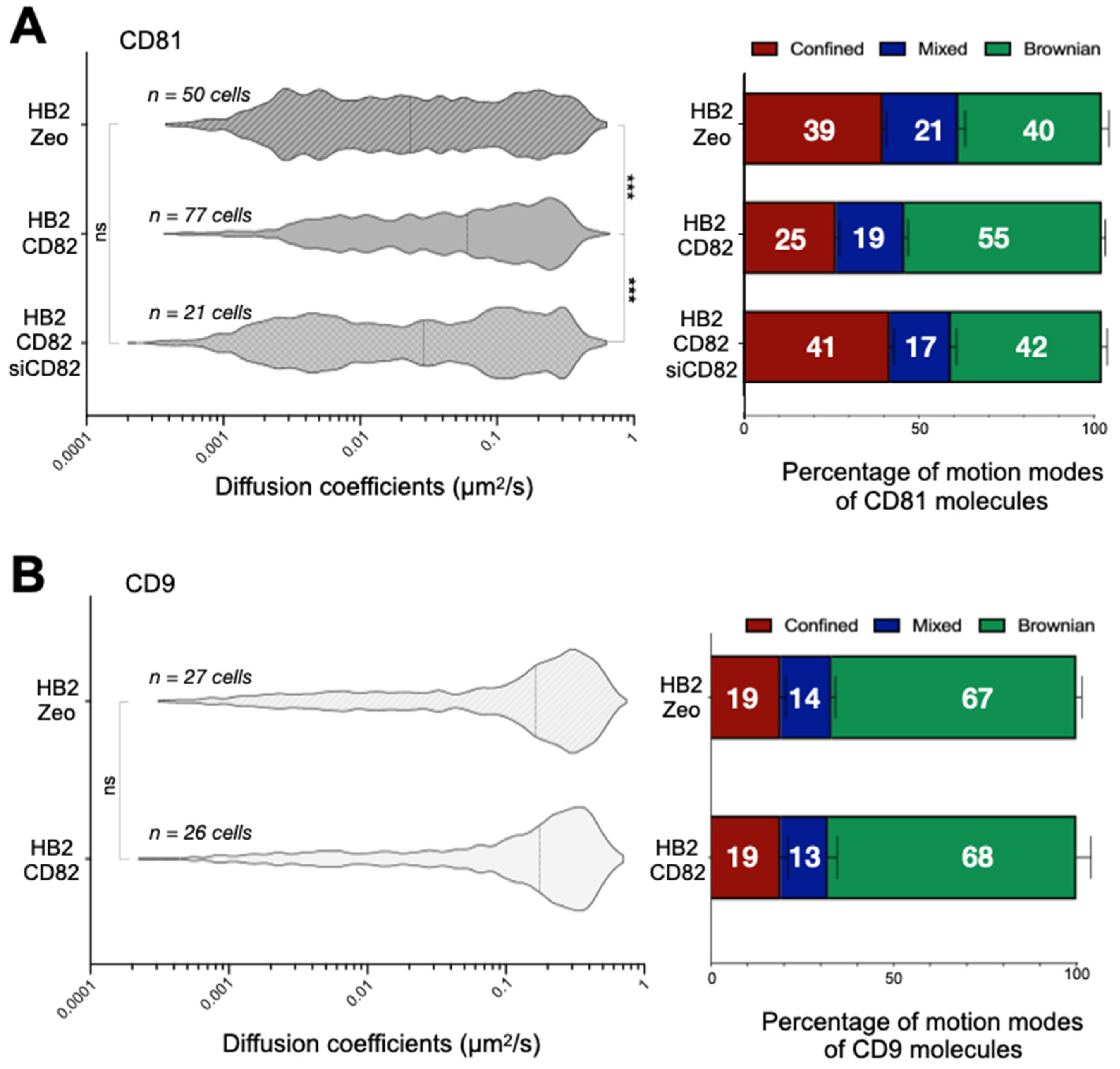
2.2. CD82 Specifically Increases CD81 Dynamics at the Plasma Membrane of HB2 Cells
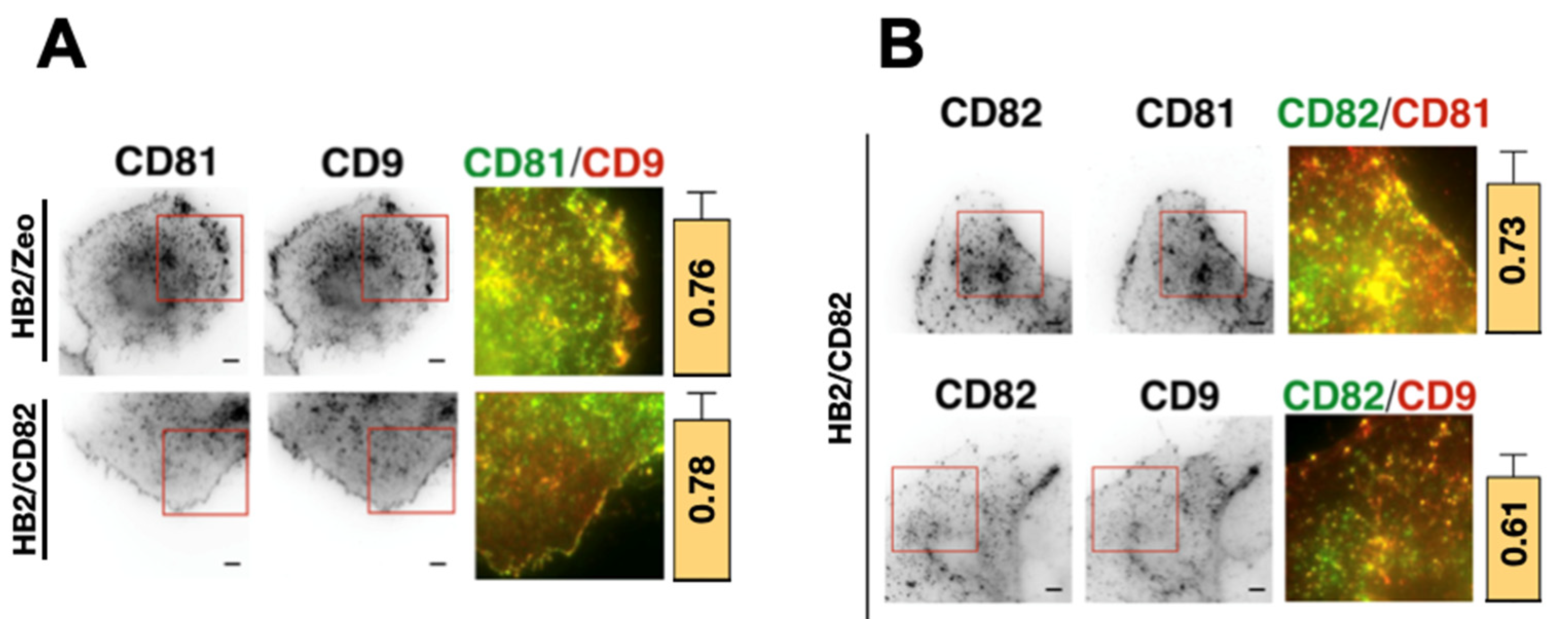
2.3. CD82 Modulates the Localization of CD81 at the Plasma Membrane
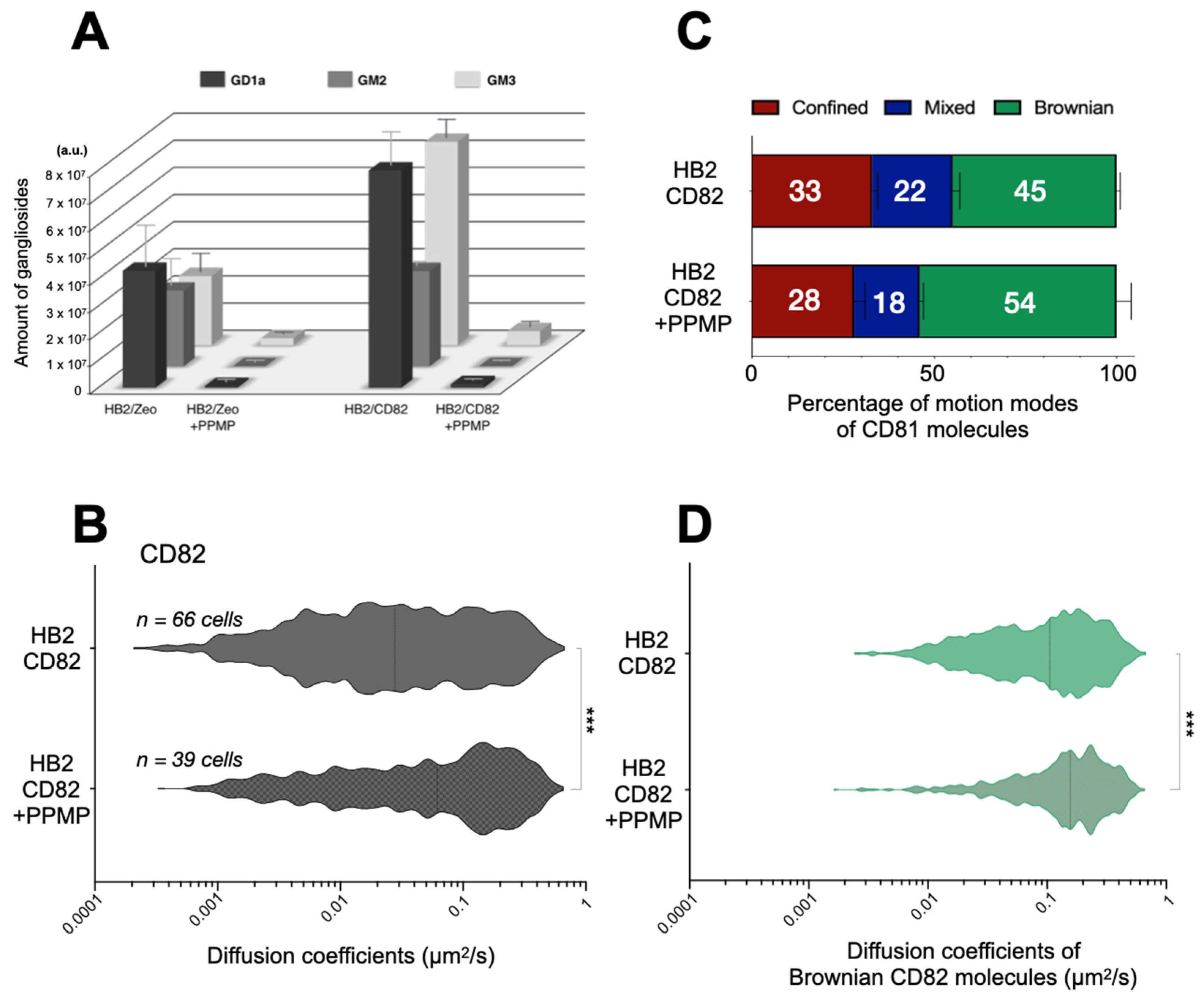
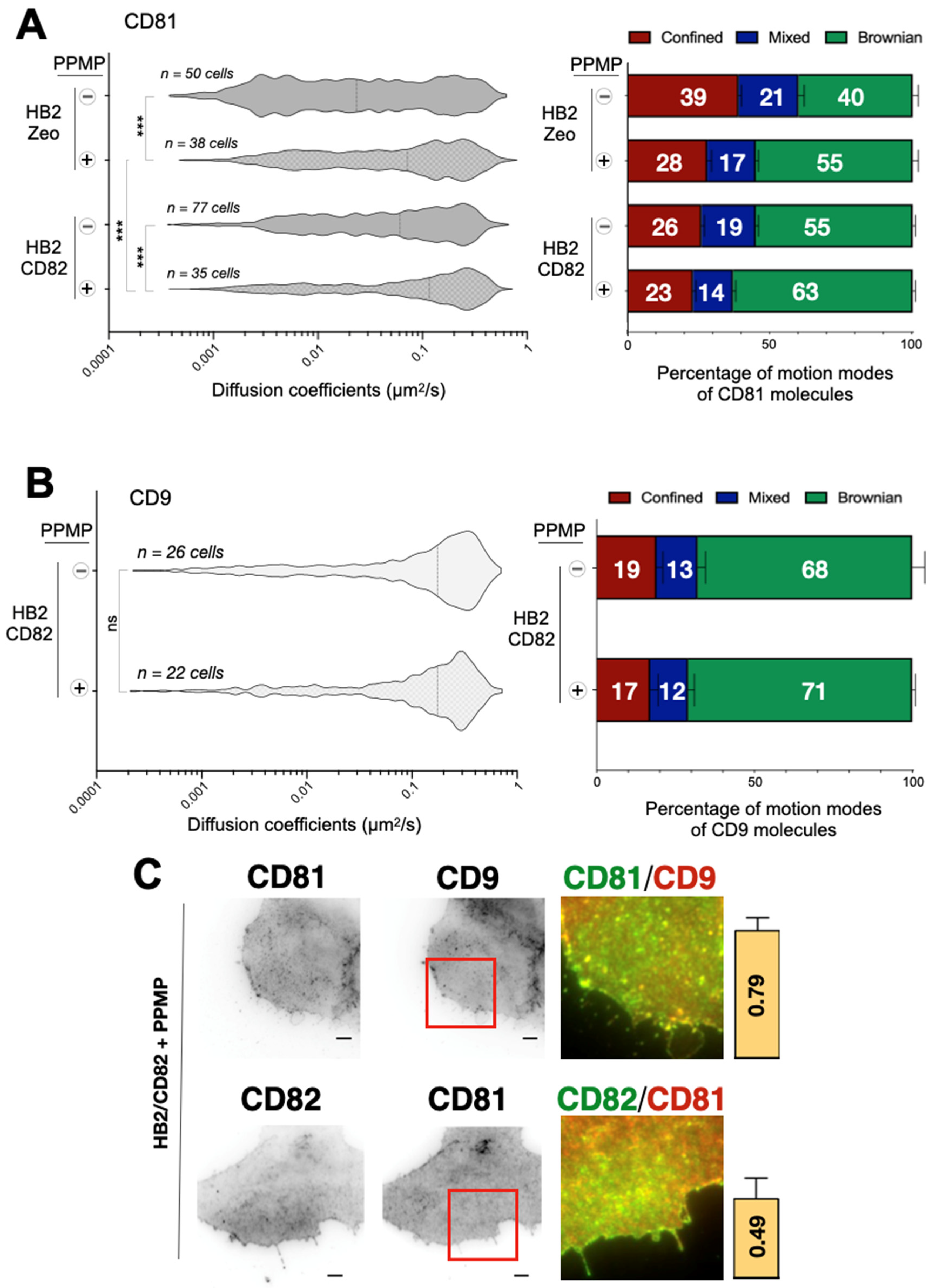
2.4. Effect of Gangliosides on the Dynamics and Organization of Tetraspanins
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Cell Culture and Treatments
4.3. SiRNA Experiments
4.4. Single-Molecule Tracking
4.5. Flow Cytometry Experiments
4.6. Dual-Color Immunofluorescence Using TIRF Microscopy
4.7. Mass Spectrometry Analysis of Lipids
4.8. Immunoprecipitations of Tetraspanins and Partners
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zimmerman, B.; Kelly, B.; McMillan, B.J.; Seegar, T.C.; Dror, R.O.; Kruse, A.C.; Blacklow, S.C. Crystal Structure of a Full-Length Human Tetraspanin Reveals a Cholesterol-Binding Pocket. Cell 2016, 167, 1041–1051.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seigneuret, M.; Delaguillaumie, A.; Lagaudriere-Gesbert, C.; Conjeaud, H. Structure of the Tetraspanin Main Extracellular Domain: A Partially Conserved Fold with a Structurally Variable Domain Insertion. J. Biol. Chem. 2001, 276, 40055–40064. [Google Scholar] [CrossRef] [Green Version]
- Hemler, M.E. Tetraspanin Proteins Mediate Cellular Penetration, Invasion, and Fusion Events and Define a Novel Type of Membrane Microdomain. Annu. Rev. Cell Dev. Biol. 2003, 19, 397–422. [Google Scholar] [CrossRef] [PubMed]
- Charrin, S.; Jouannet, S.; Boucheix, C.; Rubinstein, E. Tetraspanins at a glance. J. Cell Sci. 2014, 127, 3641–3648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemler, M.E. Tetraspanin proteins promote multiple cancer stages. Nat. Rev. Cancer 2014, 14, 49–60. [Google Scholar] [CrossRef]
- Feng, J.; Huang, C.; Wren, J.; Wang, D.-W.; Yan, J.; Zhang, J.; Sun, Y.; Han, X.; Zhang, X.A. Tetraspanin CD82: A suppressor of solid tumors and a modulator of membrane heterogeneity. Cancer Metastasis Rev. 2015, 34, 619–633. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Weissman, A.M. Dissecting the diverse functions of the metastasis suppressor CD82/KAI1. FEBS Lett. 2011, 585, 3166–3173. [Google Scholar] [CrossRef] [Green Version]
- Claas, C.; Stipp, C.S.; Hemler, M.E. Evaluation of Prototype Transmembrane 4 Superfamily Protein Complexes and Their Relation to Lipid Rafts. J. Biol. Chem. 2001, 276, 7974–7984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrin, S.; Le Naour, F.; Silvie, O.; Milhiet, P.-E.; Boucheix, C.; Rubinstein, E. Lateral organization of membrane proteins: Tetraspanins spin their web. Biochem. J. 2009, 420, 133–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez-Mó, M.; Barreiro, O.; Gordon, M.; Sala-Valdés, M.; Sánchez-Madrid, F. Tetraspanin-enriched microdomains: A functional unit in cell plasma membranes. Trends Cell Biol. 2009, 19, 434–446. [Google Scholar] [CrossRef]
- Barreiro, O.; Zamai, M.; Yáñez-Mó, M.; Tejera, E.; López-Romero, P.; Monk, P.; Gratton, E.; Caiolfa, V.R.; Sánchez-Madrid, F. Endothelial adhesion receptors are recruited to adherent leukocytes by inclusion in preformed tetraspanin nanoplatforms. J. Cell Biol. 2008, 183, 527–542. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.H.; Mirchev, R.; Deng, X.; Yacono, P.; Yang, H.L.; Golan, D.E.; Hemler, M.E. CD151 restricts α6 integrin diffusion mode. J. Cell Sci. 2012, 125, 1478–1487. [Google Scholar] [CrossRef] [Green Version]
- Espenel, C.; Margeat, E.; Dosset, P.; Arduise, C.; Le Grimellec, C.; Royer, C.; Boucheix, C.; Rubinstein, E.; Milhiet, P.-E. Single-molecule analysis of CD9 dynamics and partitioning reveals multiple modes of interaction in the tetraspanin web. J. Cell Biol. 2008, 182, 765–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krementsov, D.; Rassam, P.; Margeat, E.; Roy, N.H.; Schneider-Schaulies, J.; Milhiet, P.-E.; Thali, M. HIV-1 Assembly Differentially Alters Dynamics and Partitioning of Tetraspanins and Raft Components. Traffic 2010, 11, 1401–1414. [Google Scholar] [CrossRef] [Green Version]
- Dahmane, S.; Rubinstein, E.; Milhiet, P.-E. Viruses and Tetraspanins: Lessons from Single Molecule Approaches. Viruses 2014, 6, 1992–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danglot, L.; Chaineau, M.; Dahan, M.; Gendron, M.-C.; Boggetto, N.; Perez, F.; Galli, T. Role of TI-VAMP and CD82 in EGFR cell-surface dynamics and signaling. J. Cell Sci. 2010, 123, 723–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Claas, C.; Kraeft, S.-K.; Chen, L.B.; Wang, Z.; Kreidberg, J.A.; Hemler, M.E. Palmitoylation of Tetraspanin Proteins: Modulation of CD151 Lateral Interactions, Subcellular Distribution, and Integrin-dependent Cell Morphology. Mol. Biol. Cell 2002, 13, 767–781. [Google Scholar] [CrossRef] [Green Version]
- Miranti, C. Controlling cell surface dynamics and signaling: How CD82/KAI1 suppresses metastasis. Cell. Signal. 2009, 21, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Handa, K.; Withers, D.A.; Hakomori, S.-I. Glycosylation Effect on Membrane Domain (GEM) Involved in Cell Adhesion and Motility: A Preliminary Note on Functional α3, α5-CD82 Glycosylation Complex in ldlD 14 Cells. Biochem. Biophys. Res. Commun. 2000, 279, 744–750. [Google Scholar] [CrossRef]
- Lee, J.H.; Seo, Y.-W.; Park, S.R.; Kim, Y.J.; Kim, K.K. Expression of a splice variant of KAI1, a tumor metastasis suppressor gene, influences tumor invasion and progression. Cancer Res. 2003, 63, 7247–7255. [Google Scholar]
- Ruseva, Z.; Geiger, P.X.C.; Hutzler, P.; Kotzsch, M.; Luber, B.; Schmitt, M.; Gross, E.; Reuning, U. Tumor suppressor KAI1 affects integrin αvβ3-mediated ovarian cancer cell adhesion, motility, and proliferation. Exp. Cell Res. 2009, 315, 1759–1771. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, E.; Sugiura, T.; Berditchevski, F. Attenuation of EGF receptor signaling by a metastasis suppressor, the tetraspanin CD82/KAI-1. Curr. Biol. 2000, 10, 1009–1012. [Google Scholar] [CrossRef] [Green Version]
- Odintsova, E.; Voortman, J.; Gilbert, E.; Berditchevski, F. Tetraspanin CD82 regulates compartmentalisation and ligand-induced dimerization of EGFR. J. Cell Sci. 2003, 116, 4557–4566. [Google Scholar] [CrossRef] [Green Version]
- Hakomori, S.-I. The glycosynapse. Proc. Natl. Acad. Sci. USA 2002, 99, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huang, X.; Zhang, J.; Li, Y.; Ma, K. Synergistic inhibition of cell migration by tetraspanin CD82 and gangliosides occurs via the EGFR or cMet-activated Pl3K/Akt signalling pathway. Int. J. Biochem. Cell Biol. 2013, 45, 2349–2358. [Google Scholar] [CrossRef]
- Schmidt, T.; Schutz, G.J.; Baumgartner, W.; Gruber, H.J.; Schindler, H. Imaging of single molecule diffusion. Proc. Natl. Acad. Sci. USA 1996, 93, 2926–2929. [Google Scholar] [CrossRef] [Green Version]
- Kusumi, A.; Fujiwara, T.; Tsunoyama, T.A.; Kasai, R.; Liu, A.; Hirosawa, K.M.; Kinoshita, M.; Matsumori, N.; Komura, N.; Ando, H.; et al. Defining raft domains in the plasma membrane. Traffic 2019, 21, 106–137. [Google Scholar] [CrossRef]
- Abraham, L.; Lu, H.Y.; Falcão, R.C.; Scurll, J.; Jou, T.; Irwin, B.; Tafteh, R.; Gold, M.R.; Coombs, D. Limitations of Qdot labelling compared to directly-conjugated probes for single particle tracking of B cell receptor mobility. Sci. Rep. 2017, 7, 11379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosset, P.; Rassam, P.; Fernandez, L.; Espenel, C.; Rubinstein, E.; Margeat, E.; Milhiet, P.-E. Automatic detection of diffusion modes within biological membranes using back-propagation neural network. BMC Bioinform. 2016, 17, 197. [Google Scholar] [CrossRef] [Green Version]
- Potel, J.; Rassam, P.; Montpellier, C.; Kaestner, L.; Werkmeister, E.; Tews, B.A.; Couturier, C.; Popescu, C.-I.; Baumert, T.F.; Rubinstein, E.; et al. EWI-2wint promotes CD81 clustering that abrogates Hepatitis C Virus entry. Cell. Microbiol. 2013, 15, 1234–1252. [Google Scholar] [CrossRef]
- Harris, H.J.; Clerte, C.; Farquhar, M.J.; Goodall, M.; Hu, K.; Rassam, P.; Dosset, P.; Wilson, G.K.; Balfe, P.; Ijzendoorn, S.C.; et al. Hepatoma polarization limits CD 81 and hepatitis C virus dynamics. Cell. Microbiol. 2012, 15, 430–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berditchevski, F. Complexes of tetraspanins with integrins: More than meets the eye. J. Cell Sci. 2001, 114, 4143–4151. [Google Scholar] [CrossRef]
- Park, S.-Y.; Yoon, S.-J.; Freire-De-Lima, L.; Kim, J.-H.; Hakomori, S.-I. Control of cell motility by interaction of gangliosides, tetraspanins, and epidermal growth factor receptor in A431 versus KB epidermoid tumor cells. Carbohydr. Res. 2009, 344, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Inokuchi, J.-I.; Jimbo, M.; Shimeno, H.; Nagamatsu, A.; Shayman, J.A.; Shukla, G.S.; Radin, N.S. Improved Inhibitors of Glucosylceramide Synthase1. J. Biochem. 1992, 111, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, E.; Butters, T.D.; Monti, E.; Sprong, H.; van Meer, G.; Berditchevski, F. Gangliosides play an important role in the organization of CD82-enriched microdomains. Biochem. J. 2006, 400, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Delaguillaumie, A.; Harriague, J.; Kohanna, S.; Bismuth, G.; Rubinstein, E.; Seigneuret, M.; Conjeaud, H. Tetraspanin CD82 controls the association of cholesterol-dependent microdomains with the actin cytoskeleton in T lymphocytes: Relevance to co-stimulation. J. Cell Sci. 2004, 117, 5269–5282. [Google Scholar] [CrossRef] [Green Version]
- Delaguillaumie, A.; Lagaudrière-Gesbert, C.; Popoff, M.R.; Conjeaud, H. Rho GTPases Link Cytoskeletal Rearrangements and Activation Processes Induced via the Tetraspanin CD82 in T Lymphocytes. J. Cell Sci. 2002, 115, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Saffman, P.G.; Delbruck, M. Brownian motion in biological membranes. Proc. Natl. Acad. Sci. USA 1975, 72, 3111–3113. [Google Scholar] [CrossRef] [Green Version]
- Charrin, S.; Manié, S.; Thiele, C.; Billard, M.; Gerlier, D.; Boucheix, C.; Rubinstein, E. A physical and functional link between cholesterol and tetraspanins. Eur. J. Immunol. 2003, 33, 2479–2489. [Google Scholar] [CrossRef]
- Umeda, R.; Satouh, Y.; Takemoto, M.; Nakada-Nakura, Y.; Liu, K.; Yokoyama, T.; Shirouzu, M.; Iwata, S.; Nomura, N.; Sato, K.; et al. Structural insights into tetraspanin CD9 function. Nat. Commun. 2020, 11, 1606. [Google Scholar] [CrossRef] [Green Version]
- Weiss, L.E.; Milenkovic, L.; Yoon, J.; Stearns, T.; Moerner, W.E. Motional dynamics of single Patched1 molecules in cilia are controlled by Hedgehog and cholesterol. Proc. Natl. Acad. Sci. USA 2019, 116, 5550–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falk, J.; Thoumine, O.; Dequidt, C.; Choquet, D.; Faivre-Sarrailh, C. NrCAM Coupling to the Cytoskeleton Depends on Multiple Protein Domains and Partitioning into Lipid Rafts. Mol. Biol. Cell 2004, 15, 4695–4709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berditchevski, F.; Odintsova, E. Characterization of Integrin–Tetraspanin Adhesion Complexes: Role of Tetraspanins in Integrin Signaling. J. Cell Biol. 1999, 146, 477–492. [Google Scholar] [CrossRef] [PubMed]
- Jouannet, S.; Saint-Pol, J.; Fernandez, L.; Nguyen, V.; Charrin, S.; Boucheix, C.; Brou, C.; Milhiet, P.-E.; Rubinstein, E. TspanC8 tetraspanins differentially regulate the cleavage of ADAM10 substrates, Notch activation and ADAM10 membrane compartmentalization. Cell. Mol. Life Sci. 2016, 73, 1895–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuidscherwoude, M.; Göttfert, F.; Dunlock, V.-M.; Figdor, C.; Bogaart, G.V.D.; Van Spriel, A.B. The tetraspanin web revisited by super-resolution microscopy. Sci. Rep. 2015, 5, 12201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todeschini, A.R.; Dos Santos, J.N.; Handa, K.; Hakomori, S.-I. Ganglioside GM2/GM3 complex affixed on silica nanospheres strongly inhibits cell motility through CD82/cMet-mediated pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 1925–1930. [Google Scholar] [CrossRef] [Green Version]
- Todeschini, A.R.; Dos Santos, J.N.; Handa, K.; Hakomori, S.-I. Ganglioside GM2-Tetraspanin CD82 Complex Inhibits Met and Its Cross-talk with Integrins, Providing a Basis for Control of Cell Motility through Glycosynapse. J. Biol. Chem. 2007, 282, 8123–8133. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.G.; Ando, H.; Komura, N.; Konishi, M.; Imamura, A.; Ishida, H.; Kiso, M.; Fujiwara, T.; Kusumi, A. Revealing the Raft Domain Organization in the Plasma Membrane by Single-Molecule Imaging of Fluorescent Ganglioside Analogs. Methods Enzymol. 2018, 598, 267–282. [Google Scholar] [CrossRef]
- Charrin, S.; Le Naour, F.; Labas, V.; Billard, M.; Le Caer, J.-P.; Emile, J.-F.; Petit, M.-A.; Boucheix, C.; Rubinstein, E. EWI-2 is a new component of the tetraspanin web in hepatocytes and lymphoid cells. Biochem. J. 2003, 373, 409–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalaoui, S.; Zougbédé, S.; Charrin, S.; Silvie, O.; Arduise, C.; Farhati, K.; Boucheix, C.; Mazier, D.; Rubinstein, E.; Froissard, P. Hepatocyte Permissiveness to Plasmodium Infection Is Conveyed by a Short and Structurally Conserved Region of the CD81 Large Extracellular Domain. PLoS Pathog. 2008, 4, e1000010. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Sheetz, M.; Elson, E. Single particle tracking. Analysis of diffusion and flow in two-dimensional systems. Biophys. J. 1991, 60, 910–921. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, L.; Malrieu, M.; Bénistant, C.; Dosset, P.; Rubinstein, E.; Odintsova, E.; Berditchevski, F.; Milhiet, P.-E. CD82 and Gangliosides Tune CD81 Membrane Behavior. Int. J. Mol. Sci. 2021, 22, 8459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168459
Fernandez L, Malrieu M, Bénistant C, Dosset P, Rubinstein E, Odintsova E, Berditchevski F, Milhiet P-E. CD82 and Gangliosides Tune CD81 Membrane Behavior. International Journal of Molecular Sciences. 2021; 22(16):8459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168459
Chicago/Turabian StyleFernandez, Laurent, Morgane Malrieu, Christine Bénistant, Patrice Dosset, Eric Rubinstein, Elena Odintsova, Fedor Berditchevski, and Pierre-Emmanuel Milhiet. 2021. "CD82 and Gangliosides Tune CD81 Membrane Behavior" International Journal of Molecular Sciences 22, no. 16: 8459. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168459