
Insights into the Role of Membrane Lipids in the Structure, Function and Regulation of Integral Membrane Proteins

Abstract
:1. Introduction
2. Lipids and Respiratory Complexes
3. Secondary Active Transporters
3.1. CitS
3.2. UapA and ScBOR1p
3.3. Monoamine Transporters, hSERT and hDAT, and the Homologue LeuT
3.4. The Betaine Transporter, BetP
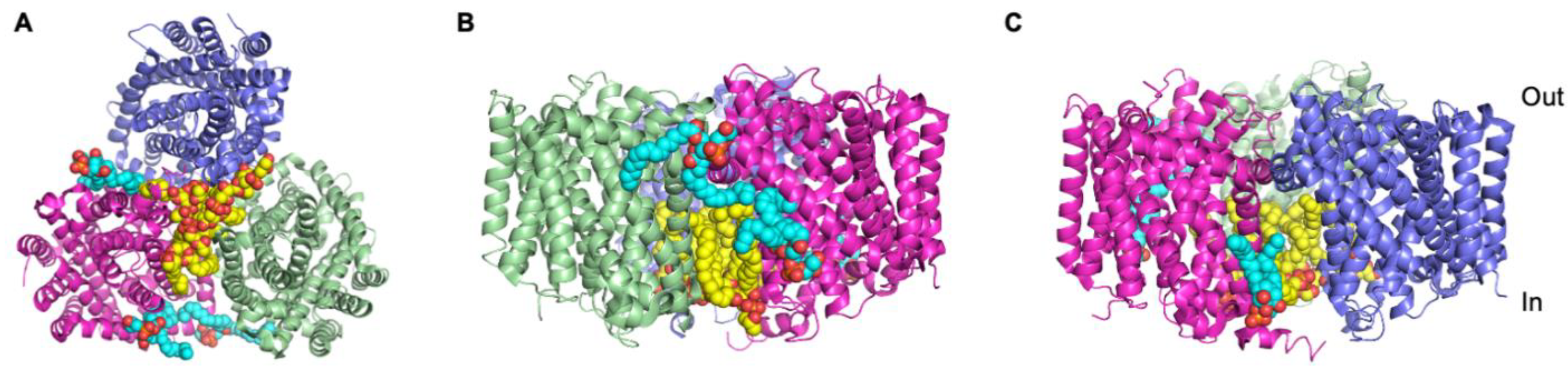
3.5. The Cationic Amino Acid Transporter, GkApcT
3.6. The Major Facilitator Superfamily Sugar Transporters, LacY and XylE
4. ATP Dependent Pumps and Transporters
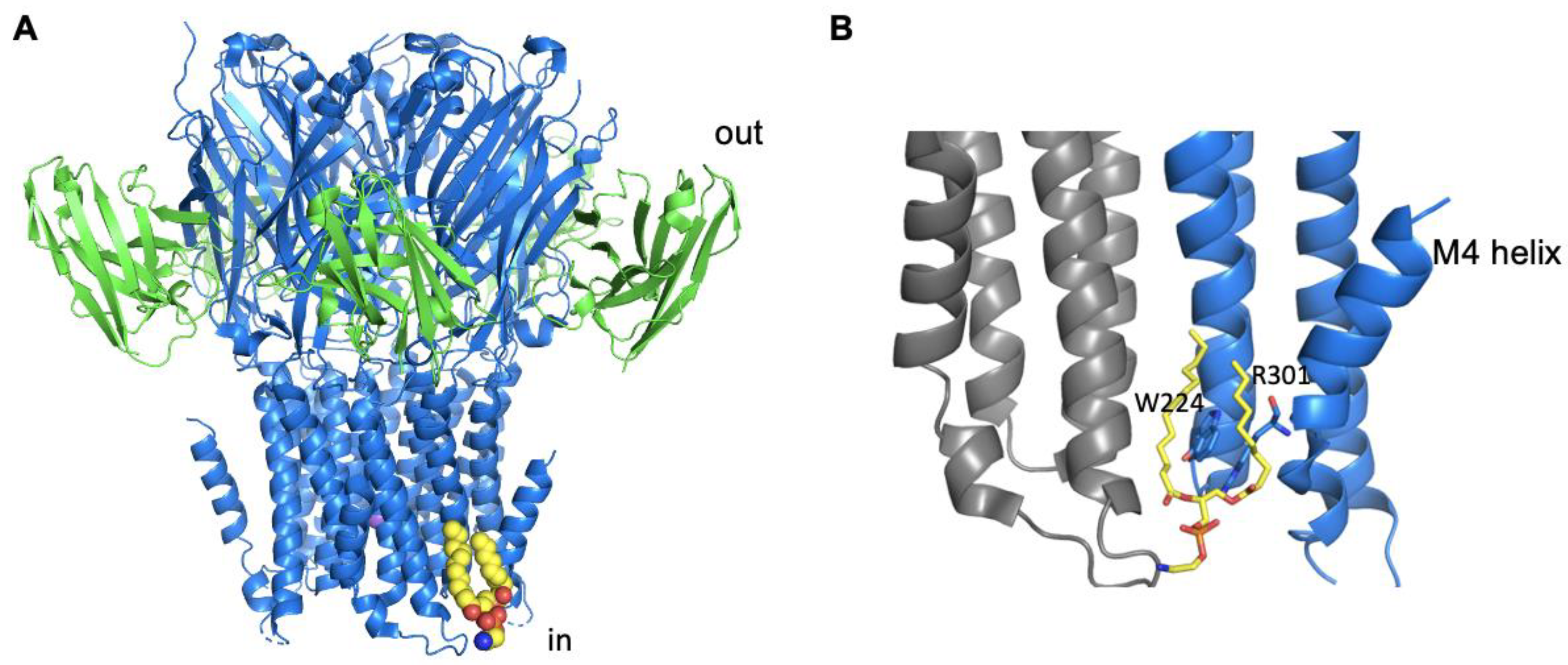
5. Channels
6. G-Protein Coupled Receptors
6.1. GPCRs and Cholesterol
6.2. GPCRs and Phospholipids
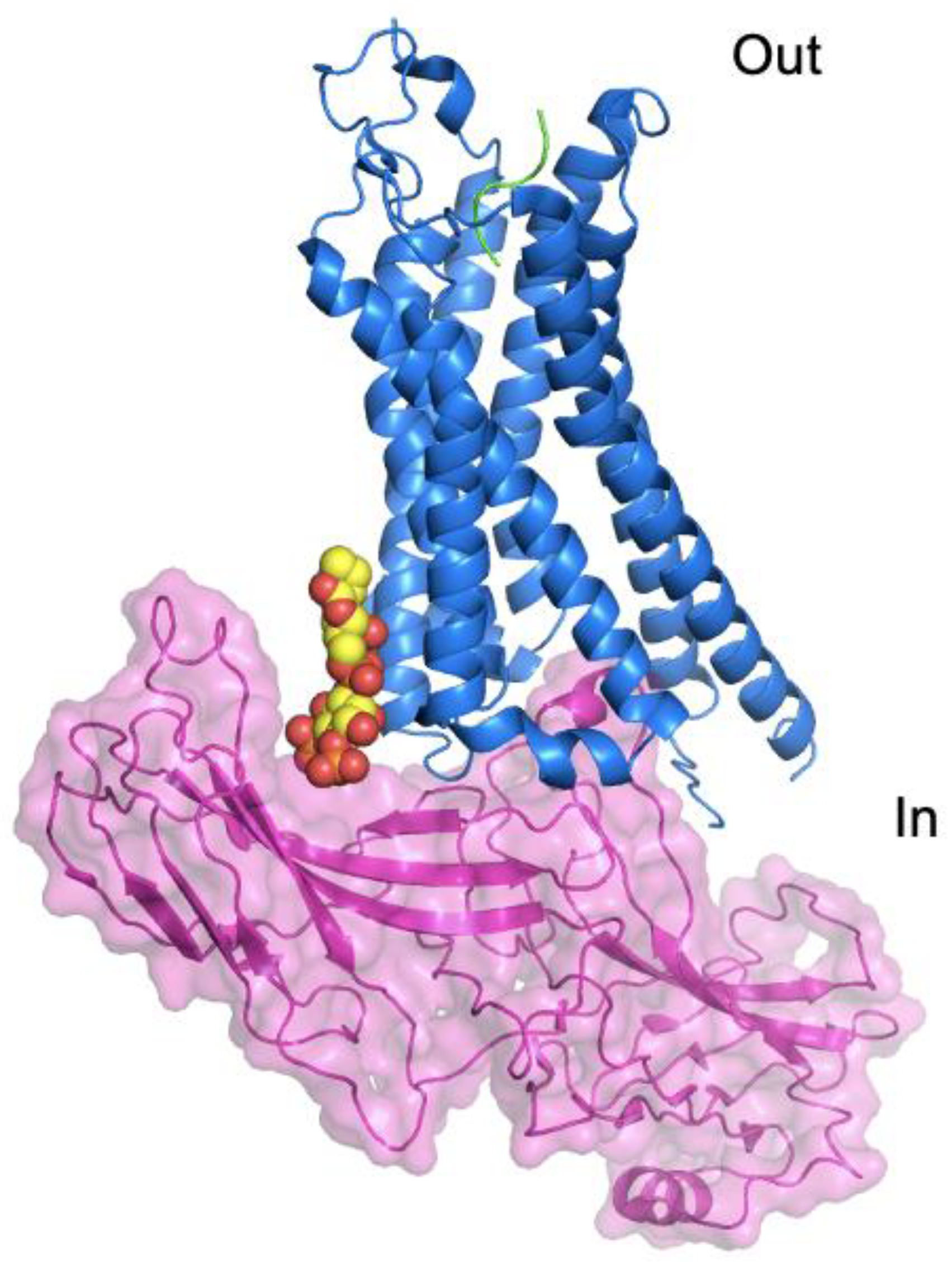
6.3. GPCR Complexes and Phospholipids
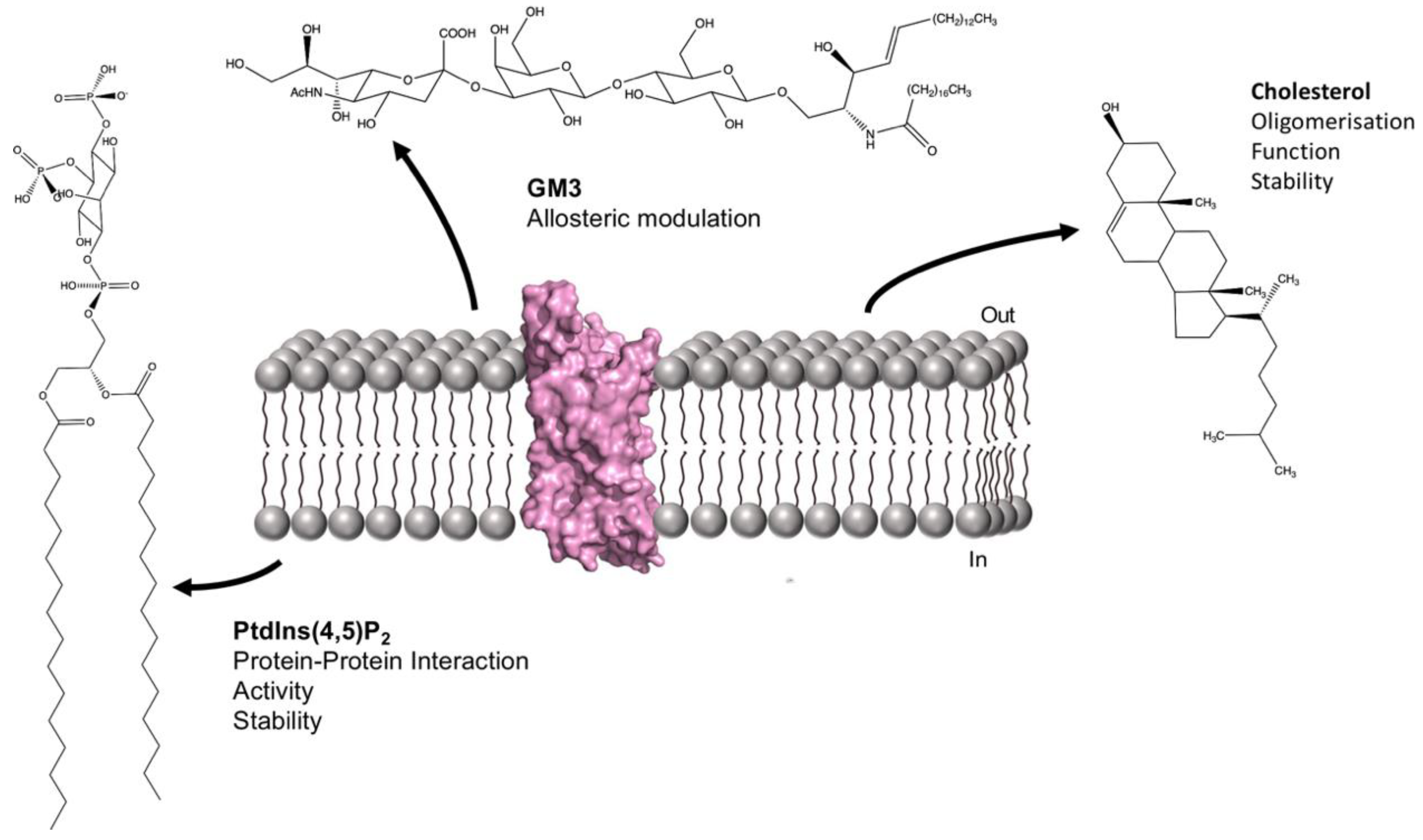
6.4. GPCRs and Sphingolipids and Glycolipids
7. Membrane Lipids and Other Membrane Proteins
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singer, S.J.; Nicolson, G.L. The Fluid Mosaic Model of the Structure of Cell Membranes. Science 1972, 175, 720–731. [Google Scholar] [CrossRef]
- Kalappurakkal, J.M.; Sil, P.; Mayor, S. Towards a New Picture of the Living Plasma Membrane. Protein Sci. 2020, 29, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Gerl, M.J. Revitalizing Membrane Rafts: New Tools and Insights. Nat. Rev. Mol. Cell Biol. 2010, 11, 688–699. [Google Scholar] [CrossRef]
- Devaux, P.F.; Morris, R. Transmembrane Asymmetry and Lateral Domains in Biological Membranes. Traffic 2004, 5, 241–246. [Google Scholar] [CrossRef]
- Laganowsky, A.; Reading, E.; Allison, T.M.; Ulmschneider, M.B.; Degiacomi, M.T.; Baldwin, A.J.; Robinson, C.V. Membrane Proteins Bind Lipids Selectively to Modulate Their Structure and Function. Nature 2014, 510, 172–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Cang, Z.; Yao, J.; Kim, M.; Deans, E.; Wei, G.; Kang, S.; Hong, H. Structural Cavities Are Critical to Balancing Stability and Activity of a Membrane-Integral Enzyme. Proc. Natl. Acad. Sci. USA 2020, 117, 22146–22156. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y. Be Cautious with Crystal Structures of Membrane Proteins or Complexes Prepared in Detergents. Crystals 2020, 10, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjerregaard, H.; Severinsen, K.; Said, S.; Wiborg, O.; Sinning, S. A Dualistic Conformational Response to Substrate Binding in the Human Serotonin Transporter Reveals a High Affinity State for Serotonin. J. Biol. Chem. 2015, 290, 7747–7755. [Google Scholar] [CrossRef] [Green Version]
- Laursen, L.; Severinsen, K.; Kristensen, K.B.; Periole, X.; Overby, M.; Müller, H.K.; Schiøtt, B.; Sinning, S. Cholesterol Binding to a Conserved Site Modulates the Conformation, Pharmacology, and Transport Kinetics of the Human Serotonin Transporter. J. Biol. Chem. 2018, 293, 3510–3523. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Lev, B.; Hossain, K.R.; Gorman, A.; Diaz, D.; Pham, T.H.N.; Cornelius, F.; Allen, T.W.; Clarke, R.J. Cholesterol Depletion Inhibits Na+, K+-ATPase Activity in a near-Native Membrane Environment. J. Biol. Chem. 2019, 294, 5956–5969. [Google Scholar] [CrossRef]
- Sharom, F.J. Complex Interplay between the P-Glycoprotein Multidrug Efflux Pump and the Membrane: Its Role in Modulating Protein Function. Front. Oncol. 2014, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.K.; Rosenhouse-Dantsker, A.; Nichols, C.G.; Enkvetchakul, D.; Levitan, I. Direct Regulation of Prokaryotic Kir Channel by Cholesterol. J. Biol. Chem. 2009, 284, 30727–30736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Friedberg, L.; Bose-Boyd, R.D.; Long, T.; Li, X. Sterols in an Intramolecular Channel of Smoothened Mediate Hedgehog Signaling. Nat. Chem. Biol. 2020, 16, 1368–1375. [Google Scholar] [CrossRef]
- Yeliseev, A.; Iyer, M.R.; Joseph, T.T.; Coffey, N.J.; Cinar, R.; Zoubak, L.; Kunos, G.; Gawrisch, K. Cholesterol as a Modulator of Cannabinoid Receptor CB2 Signaling. Sci. Rep. 2021, 11, 3706–3718. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Pearsall, E.A.; Hurst, D.P.; Zhang, Y.; Chu, J.; Zhou, Y.; Reggio, P.H.; Loh, H.H.; Law, P.-Y. Palmitoylation and Membrane Cholesterol Stabilize μ-Opioid Receptor Homodimerization and G Protein Coupling. BMC Cell Biol. 2012, 13, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Gimpl, G. Interaction of G Protein Coupled Receptors and Cholesterol. Chem. Phys. Lipids 2016, 199, 61–73. [Google Scholar] [CrossRef]
- Waltenspühl, Y.; Schöppe, J.; Ehrenmann, J.; Kummer, L.; Plückthun, A. Crystal Structure of the Human Oxytocin Receptor. Sci. Adv. 2020, 6, eabb5419. [Google Scholar] [CrossRef]
- Sejdiu, B.I.; Tieleman, D.P. Lipid-Protein Interactions Are a Unique Property and Defining Feature of G Protein-Coupled Receptors. Biophys. J. 2020, 118, 1887–1900. [Google Scholar] [CrossRef]
- Cherezov, V.; Rosenbaum, D.M.; Hanson, M.A.; Rasmussen, S.G.F.; Thian, F.S.; Kobilka, T.S.; Choi, H.J.; Kuhn, P.; Weis, W.I.; Kobilka, B.K.; et al. High-Resolution Crystal Structure of an Engineered Human 2-Adrenergic G Protein Coupled Receptor. Science 2007, 318, 1258–1265. [Google Scholar] [CrossRef] [Green Version]
- Manglik, A.; Kruse, A.C.; Kobilka, T.S.; Thian, F.S.; Mathiesen, J.M.; Sunahara, R.K.; Pardo, L.; Weis, W.I.; Kobilka, B.K.; Granier, S. Crystal Structure of the Μ-Opioid Receptor Bound to a Morphinan Antagonist. Nature 2012, 485, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Manglik, A.; Venkatakrishnan, A.J.; Laeremans, T.; Feinberg, E.N.; Sanborn, A.L.; Kato, H.E.; Livingston, K.E.; Thorsen, T.S.; Kling, R.C.; et al. Structural Insights into Μ-Opioid Receptor Activation. Nature 2015, 524, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Gahbauer, S.; Böckmann, R.A. Membrane-Mediated Oligomerization of G Protein Coupled Receptors and Its Implications for GPCR Function. Front. Physiol. 2016, 7, 494. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, X.; Mohole, M.; Chattopadhyay, A.; Sengupta, D. Role of Cholesterol-Mediated Effects in GPCR Heterodimers. Chem. Phys. Lipids 2020, 227, 104852. [Google Scholar] [CrossRef]
- Schrecke, S.; Zhu, Y.; McCabe, J.W.; Bartz, M.; Packianathan, C.; Zhao, M.; Zhou, M.; Russell, D.; Laganowsky, A. Selective Regulation of Human TRAAK Channels by Biologically Active Phospholipids. Nat. Chem. Biol. 2021, 17, 89–95. [Google Scholar] [CrossRef]
- Yen, H.-Y.; Hoi, K.K.; Liko, I.; Hedger, G.; Horrell, M.R.; Song, W.; Wu, D.; Heine, P.; Warne, T.; Lee, Y.; et al. PtdIns(4,5)P2 Stabilizes Active States of GPCRs and Enhances Selectivity of G-Protein Coupling. Nature 2018, 559, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Pyle, E.; Kalli, A.C.; Amillis, S.; Hall, Z.; Lau, A.M.; Hanyaloglu, A.C.; Diallinas, G.; Byrne, B.; Politis, A. Structural Lipids Enable the Formation of Functional Oligomers of the Eukaryotic Purine Symporter UapA. Cell Chem. 2018, 25, 840–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyle, E.; Guo, C.; Hofmann, T.; Schmidt, C.; Ribiero, O.; Politis, A.; Byrne, B. Protein–lipid Interactions Stabilize the Oligomeric State of BOR1p from Saccharomyces Cerevisiae. Anal. Chem. 2019, 91, 13071–13079. [Google Scholar] [CrossRef] [PubMed]
- Martens, C.; Shekhar, M.; Borysik, A.J.; Lau, A.M.; Reading, E.; Tajkhorshid, E.; Booth, P.J.; Politis, A. Direct Protein–lipid Interactions Shape the Conformational Landscape of Secondary Transporters. Nat. Commun. 2018, 9, 4151. [Google Scholar] [CrossRef] [PubMed]
- Hénault, C.M.; Govaerts, C.; Spurny, R.; Brams, M.; Estrada-Mondragon, A.; Lynch, J.; Bertrand, D.; Pardon, E.; Evans, G.L.; Woods, K.; et al. A Lipid Site Shapes the Agonist Response of a Pentameric Ligand-Gated Ion Channel. Nat. Chem. Biol. 2019, 15, 1156–1164. [Google Scholar] [CrossRef]
- Dawaliby, R.; Trubbia, C.; Delporte, C.; Masureel, M.; Antwerpen, P.V.; Kobilka, B.K.; Govaerts, C. Allosteric Regulation of G Protein–Coupled Receptor Activity by Phospholipids. Nat. Chem. Biol. 2016, 12, 35–39. [Google Scholar] [CrossRef]
- Strohman, M.J.; Maeda, S.; Hilger, D.; Masureel, M.; Du, Y.; Kobilka, B.K. Local Membrane Charge Regulates β2 Adrenergic Receptor Coupling to Gi3. Nat. Commun. 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dijkman, P.M.; Watts, A. Lipid Modulation of Early G Protein-Coupled Receptor Signalling Events. BBA Biomembr. 2015, 1848, 2889–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-J.; Jeong, H.; Hyun, J.; Ryu, B.; Park, K.; Lim, H.-H.; Yoo, J.; Woo, J.-S. Cryo-EM Structure of Human Cx31.3/GJC3 Connexin Hemichannel. Sci. Adv. 2020, 6, eaba4996. [Google Scholar] [CrossRef] [PubMed]
- Gahbauer, S.; Böckmann, R.A. Comprehensive Characterization of Lipid-Guided G Protein-Coupled Receptor Dimerization. J. Phys. Chem. B 2020, 124, 2823–2834. [Google Scholar] [CrossRef]
- Staus, D.P.; Hu, H.; Robertson, M.J.; Kleinhenz, A.L.W.; Wingler, L.M.; Capel, W.D.; Latorraca, N.R.; Lefkowitz, R.J.; Skiniotis, G. Structure of the M2 Muscarinic Receptor-β-Arrestin Complex in a Lipid Nanodisc. Nature 2020, 579, 297–302. [Google Scholar] [CrossRef]
- Koshy, C.; Schweikhard, E.S.; Gärtner, R.M.; Perez, C.; Yildiz, Ö.; Ziegler, C. Structural Evidence for Functional Lipid Interactions in the Betaine Transporter BetP. EMBO J. 2013, 32, 3096–3105. [Google Scholar] [CrossRef] [Green Version]
- Tong, A.; II, J.T.P.; Hsu, F.-F.; Schmidpeter, P.A.; Nimigean, C.M.; Sharp, L.; Brannigan, G.; Cheng, W.W. Direct Binding of Phosphatidylglycerol at Specific Sites Modulates Desensitization of a Ligand-Gated Ion Channel. ELife 2019, 8, e50766. [Google Scholar] [CrossRef]
- Zhang, M.; Gui, M.; Wang, Z.-F.; Gorgulla, C.; Yu, J.J.; Wu, H.; Sun, Z.-Y.J.; Klenk, C.; Merklinger, L.; Morstein, L.; et al. Cryo-EM Structure of an Activated GPCR-G Protein Complex in Lipid Nanodiscs. Nat. Struct. Mol. Biol. 2021, 28, 258–267. [Google Scholar] [CrossRef]
- Bruzzese, A.; Dalton, J.A.R.; Giraldo, J. Insights into Adenosine A2A Receptor Activation through Cooperative Modulation of Agonist and Allosteric Lipid Interactions. PLoS Comput. Biol. 2020, 16, e1007818. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, A.; Hofmaier, T.; Klotzsch, E.; Kudlacek, O.; Stockner, T.; Sitte, H.H.; Schütz, G.J. Direct PIP2 Binding Mediates Stable Oligomer Formation of the Serotonin Transporter. Nat. Commun. 2017, 8, 14089. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, P.J.; Belovich, A.N.; Khelashvili, G.; Saunders, C.; Erreger, K.; Javitch, J.A.; Sitte, H.H.; Weinstein, H.; Matthies, H.J.G.; Galli, A. PIP2 Regulates Psychostimulant Behaviors through Its Interaction with a Membrane Protein. Nat. Chem. Biol. 2014, 10, 582–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, A.L.; Corey, R.A.; Sansom, M.S.P. Defining How Multiple Lipid Species Interact with Inward Rectifier Potassium (Kir2) Channels. Proc. Natl. Acad. Sci. USA 2020, 117, 7803–7813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGoldrick, L.L.; Singh, A.K.; Demirkhanyan, L.; Lin, T.-Y.; Casner, R.G.; Zakharian, E.; Sobolevsky, A.I. Structure of the Thermo-Sensitive TRP Channel TRP1 from the Alga Chlamydomonas Reinhardtii. Nat. Commun. 2019, 10, 4180. [Google Scholar] [CrossRef]
- Gao, Y.; Cao, E.; Julius, D.; Cheng, Y. TRPV1 Structures in Nanodiscs Reveal Mechanisms of Ligand and Lipid Action. Nature 2016, 534, 347–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damian, M.; Louet, M.; Gomes, A.A.S.; M’Kadmi, C.; Denoyelle, S.; Cantel, S.; Mary, S.; Bisch, P.M.; Fehrentz, J.-A.; Catoire, L.J.; et al. Allosteric Modulation of Ghrelin Receptor Signaling by Lipids. Nat. Commun. 2021, 12, 3938. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Masureel, M.; Qu, Q.; Janetzko, J.; Inoue, A.; Kato, H.E.; Robertson, M.J.; Nguyen, K.C.; Glenn, J.S.; Skiniotis, G.; et al. Structure of the Neurotensin Receptor 1 in Complex with β-Arrestin 1. Nature 2020, 579, 303–308. [Google Scholar] [CrossRef]
- Ansell, T.B.; Song, W.; Sansom, M.S.P. The Glycosphingolipid GM3 Modulates Conformational Dynamics of the Glucagon Receptor. Biophys. J. 2020, 119, 300–313. [Google Scholar] [CrossRef]
- Paila, Y.D.; Ganguly, S.; Chattopadhyay, A. Metabolic Depletion of Sphingolipids Impairs Ligand Binding and Signaling of Human Serotonin 1A Receptors. Biochemistry 2010, 49, 2389–2397. [Google Scholar] [CrossRef]
- Prasanna, X.; Jafurulla, M.; Sengupta, D.; Chattopadhyay, A. The Ganglioside GM1 Interacts with the Serotonin1A Receptor via the Sphingolipid Binding Domain. BBA Biomembr. 2016, 1858, 2818–2826. [Google Scholar] [CrossRef]
- Jormakka, M.; Törnroth, S.; Byrne, B.; Iwata, S. Molecular Basis of Proton Motive Force Generation: Structure of Formate Dehydrogenase-N. Science 2002, 295, 1863–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, K.; Donlan, J.A.C.; Hopper, J.T.S.; Uzdavinys, P.; Landreh, M.; Struwe, W.B.; Drew, D.; Baldwin, A.J.; Stansfeld, P.J.; Robinson, C.V. The Role of Interfacial Lipids in Stabilizing Membrane Protein Oligomers. Nature 2017, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bolla, J.R.; Sauer, J.B.; Wu, D.; Mehmood, S.; Allison, T.M.; Robinson, C.V. Direct Observation of the Influence of Cardiolipin and Antibiotics on Lipid II Binding to MurJ. Nat. Chem. 2018, 10, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Mizumura, T.; Kondo, K.; Kurita, M.; Kofuku, Y.; Natsume, M.; Imai, S.; Shiraishi, Y.; Ueda, T.; Shimada, I. Activation of Adenosine A2A Receptor by Lipids from Docosahexaenoic Acid Revealed by NMR. Sci. Adv. 2020, 6, eaay8544. [Google Scholar] [CrossRef] [Green Version]
- Leonard, A.N.; Lyman, E. Activation of G-Protein-Coupled Receptors Is Thermodynamically Linked to Lipid Solvation. Biophys. J. 2021, 120, 1777–1787. [Google Scholar] [CrossRef]
- Javanainen, M.; Enkavi, G.; Guixà-Gonzaléz, R.; Kulig, W.; Martinez-Seara, H.; Levental, I.; Vattulainen, I. Reduced Level of Docosahexaenoic Acid Shifts GPCR Neuroreceptors to Less Ordered Membrane Regions. PLoS Comput. Biol. 2019, 15, e1007033. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Johnston, J.M.; Wang, H.; Khelashvili, G.; Filizola, M.; Weinstein, H. Membrane Driven Spatial Organization of GPCRs. Sci. Rep. 2013, 3, 2909. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, F.; Saito, N.; Tanimoto, Y.; Okada, K.; Morigaki, K.; Seno, K.; Maekawa, S. Raftophilic Rhodopsin-Clusters Offer Stochastic Platforms for G Protein Signalling in Retinal Discs. Commun. Biol. 2019, 2, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Benlekbir, S.; Venkatakrishnan, P.; Wang, Y.; Hong, S.; Hosler, J.; Tajkhorshid, E.; Rubinstein, J.L.; Gennis, R.B. Structure of the Alternative Complex III in a Supercomplex with Cytochrome Oxidase. Nature 2018, 557, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.J.; Finka, R.; Finka, R.; Smith, C.; Smith, C.; Lin, Y.-P.; Dafforn, T.; Dafforn, T.; Overduin, M. Membrane Proteins Solubilized Intact in Lipid Containing Nanoparticles Bounded by Styrene Maleic Acid Copolymer. J. Am. Chem. Soc. 2009, 131, 7484–7485. [Google Scholar] [CrossRef]
- Palsdottir, H.; Hunte, C. Lipids in Membrane Protein Structures. Biochim. Biophys. Acta Biomembr. 2004, 1666, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Lange, C.; Nett, J.H.; Trumpower, B.L.; Hunte, C. Specific Roles of Protein—Phospholipid Interactions in the Yeast Cytochrome Bc1 Complex Structure. EMBO J. 2001, 20, 6591–6600. [Google Scholar] [CrossRef] [Green Version]
- Palsdottir, H.; Lojero, C.G.; Trumpower, B.L.; Hunte, C. Structure of the Yeast Cytochrome Bc1 Complex with a Hydroxyquinone Anion Qo Site Inhibitor Bound. J. Biol. Chem. 2003, 278, 31303–31311. [Google Scholar] [CrossRef] [Green Version]
- Pinke, G.; Zhou, L.; Sazanov, L.A. Cryo-EM Structure of the Entire Mammalian F-Type ATP Synthase. Nat. Struct. Mol. Biol. 2020, 27, 1077–1085. [Google Scholar] [CrossRef]
- Wöhlert, D.; Grötzinger, M.J.; Kühlbrandt, W.; Yildiz, Ö. Mechanism of Na(+)—Dependent Citrate Transport from the Structure of an Asymmetrical CitS Dimer. ELife 2015, 4, e09375. [Google Scholar] [CrossRef]
- Alguel, Y.; Amillis, S.; Leung, J.; Lambrinidis, G.; Capaldi, S.; Scull, N.J.; Craven, G.; Iwata, S.; Armstrong, A.; Mikros, E.; et al. Structure of Eukaryotic Purine/H(+) Symporter UapA Suggests a Role for Homodimerization in Transport Activity. Nat. Commun. 2016, 7, 11336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolla, J.R.; Agasid, M.T.; Mehmood, S.; Robinson, C.V. Membrane Protein–Lipid Interactions Probed Using Mass Spectrometry. Annu. Rev. Biochem. 2019, 88, 85–111. [Google Scholar] [CrossRef] [PubMed]
- Thurtle-Schmidt, B.H.; Stroud, R.M. Structure of Bor1 Supports an Elevator Transport Mechanism for SLC4 Anion Exchangers. Proc. Natl. Acad. Sci. USA 2016, 113, 10542–10546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.K.; Kudlacek, O.; Baumgart, F.; Jaentsch, K.; Stockner, T.; Sitte, H.H.; Schütz, G.J. Dopamine Transporter Forms Stable Dimers in the Live Cell Plasma Membrane in a Phosphatidylinositol 4,5-Bisphosphate-Independent Manner. J. Biol. Chem. 2019, 294, 5632–5642. [Google Scholar] [CrossRef] [Green Version]
- Penmatsa, A.; Wang, K.H.; Gouaux, E. X-Ray Structure of Dopamine Transporter Elucidates Antidepressant Mechanism. Nature 2013, 503, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.H.; Penmatsa, A.; Gouaux, E. Neurotransmitter and Psychostimulant Recognition by the Dopamine Transporter. Nature 2015, 521, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.A.; Green, E.M.; Gouaux, E. X-Ray Structures and Mechanism of the Human Serotonin Transporter. Nature 2016, 532, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.C.; Amara, S.G. Membrane Cholesterol Modulates the Outward Facing Conformation of the Dopamine Transporter and Alters Cocaine Binding. J. Biol. Chem. 2010, 285, 32616–32626. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.T.; Zhen, J.; Reith, M.E.A. Importance of Cholesterol in Dopamine Transporter Function. J. Neurochem. 2012, 123, 700–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechara, C.; Robinson, C.V. Different Modes of Lipid Binding to Membrane Proteins Probed by Mass Spectrometry. J. Am. Chem. Soc. 2015, 137, 5240–5247. [Google Scholar] [CrossRef] [PubMed]
- Bolla, J.R.; Corey, R.A.; Sahin, C.; Gault, J.; Hummer, A.; Hopper, J.T.S.; Lane, D.P.; Drew, D.; Allison, T.M.; Stansfeld, P.J.; et al. A Mass-Spectrometry-Based Approach to Distinguish Annular and Specific Lipid Binding to Membrane Proteins. Angew. Chem. Int. Ed. 2020, 59, 3523–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungnickel, K.E.J.; Parker, J.L.; Newstead, S. Structural Basis for Amino Acid Transport by the CAT Family of SLC7 Transporters. Nat. Commun. 2018, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pearson, A.; Page, S.R.F.; Jorgenson, T.L.; Fischer, W.W.; Higgins, M.B. Novel Hopanoid Cyclases from the Environment. Environ. Microbiol. 2007, 9, 2175–2188. [Google Scholar] [CrossRef]
- Mistarz, U.H.; Brown, J.M.; Haselmann, K.F.; Rand, K.D. Probing the Binding Interfaces of Protein Complexes Using Gas-Phase H/D Exchange Mass Spectrometry. Struct. Fold. Des. 2016, 24, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Konermann, L.; Pan, J.; Liu, Y.-H. Hydrogen Exchange Mass Spectrometry for Studying Protein Structure and Dynamics. Chem. Soc. Rev. 2011, 40, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Esmaili, M.; Eldeeb, M.A.; Moosavi-Movahedi, A.A. Current Developments in Native Nanometric Discoidal Membrane Bilayer Formed by Amphipathic Polymers. Nanomaterials 2021, 11, 1771. [Google Scholar] [CrossRef]
- Farrelly, M.D.; Martin, L.L.; Thang, S.H. Polymer Nanodiscs and Their Bioanalytical Potential. Chem. Eur. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Reading, E.; Hall, Z.; Martens, C.; Haghighi, T.; Findlay, H.; Ahdash, Z.; Politis, A.; Booth, P.J. Interrogating Membrane Protein Conformational Dynamics within Native Lipid Compositions. Angew. Chem. Int. Ed. 2017, 129, 15860–15863. [Google Scholar] [CrossRef]
- Chorev, D.S.; Baker, L.A.; Wu, D.; Beilsten-Edmands, V.; Rouse, S.L.; Zeev-Ben-Mordehai, T.; Jiko, C.; Samsudin, F.; Gerle, C.; Khalid, S.; et al. Protein Assemblies Ejected Directly from Native Membranes Yield Complexes for Mass Spectrometry. Science 2018, 362, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinoda, T.; Ogawa, H.; Cornelius, F.; Toyoshima, C. Crystal Structure of the Sodium-Potassium Pump at 2.4 A Resolution. Nature 2009, 459, 446–450. [Google Scholar] [CrossRef]
- Eckford, P.; Sharom, F.J. ABC Efflux Pump-Based Resistance to Chemotherapy Drugs. Chem. Rev. 2009, 109, 2989–3011. [Google Scholar] [CrossRef] [PubMed]
- Thangapandian, S.; Kapoor, K.; Tajkhorshid, E. Probing Cholesterol Binding and Translocation in P-Glycoprotein. BBA Biomembr. 2020, 1862, 183090. [Google Scholar] [CrossRef]
- Duncan, A.L.; Song, W.; Sansom, M.S.P. Lipid-Dependent Regulation of Ion Channels and G Protein–Coupled Receptors: Insights from Structures and Simulations. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 31–50. [Google Scholar] [CrossRef]
- Hansen, S.B.; Tao, X.; MacKinnon, R. Structural Basis of PIP2 Activation of the Classical Inward Rectifier K+ Channel Kir2.2. Nature 2011, 477, 495–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.W.L.; D’Avanzo, N.; Doyle, D.A.; Nichols, C.G. Dual-Mode Phospholipid Regulation of Human Inward Rectifying Potassium Channels. Biophys. J. 2011, 100, 620–628. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Wang, S.; Borschel, W.; Heyman, S.; Gyore, J.; Nichols, C.G. Secondary Anionic Phospholipid Binding Site and Gating Mechanism in Kir2.1 Inward Rectifier Channels. Nat. Commun. 2013, 4, 2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Hu, M.; Yang, Y.; Xu, H. Organellar TRP Channels. Nat. Struct. Mol. Biol. 2018, 25, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Nadezhdin, K.D.; Neuberger, A.; Trofimov, Y.A.; Krylov, N.A.; Sinica, V.; Kupko, N.; Vlachova, V.; Zakharian, E.; Efremov, R.G.; Sobolevsky, A.I. Structural Mechanism of Heat-Induced Opening of a Temperature-Sensitive TRP Channel. Nat. Struct. Mol. Biol. 2021, 28, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Glukhova, A.; Thal, D.M.; Nguyen, A.T.; Vecchio, E.A.; Jörg, M.; Scammells, P.J.; May, L.T.; Sexton, P.M.; Christopoulos, A. Structure of the Adenosine A1 Receptor Reveals the Basis for Subtype Selectivity. Cell 2017, 168, 867–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucadyil, T.J.; Chattopadhyay, A. Role of Cholesterol in the Function and Organization of G-Protein Coupled Receptors. Prog. Lipid Res. 2006, 45, 295–333. [Google Scholar] [CrossRef]
- Paila, Y.D.; Chattopadhyay, A. The Function of G-Protein Coupled Receptors and Membrane Cholesterol: Specific or General Interaction? Glycoconj. J. 2008, 26, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Salas-Estrada, L.A.; Leioatts, N.; Romo, T.D.; Grossfield, A. Lipids Alter Rhodopsin Function via Ligand-like and Solvent-like Interactions. Biophys. J. 2018, 114, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, M.A.; Cherezov, V.; Griffith, M.T.; Roth, C.B.; Jaakola, V.-P.; Chien, E.Y.T.; Velasquez, J.; Kuhn, P.; Stevens, R.C. A Specific Cholesterol Binding Site Is Established by the 2.8 Å Structure of the Human Β2-Adrenergic Receptor. Structure 2008, 16, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Taghon, G.J.; Rowe, J.B.; Kapolka, N.J.; Isom, D.G. Predictable Cholesterol Binding Sites in GPCRs Lack Consensus Motifs. Structure 2021, 29, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Lemel, L.; Nieścierowicz, K.; García-Fernández, M.D.; Darré, L.; Durroux, T.; Busnelli, M.; Pezet, M.; Rébeillé, F.; Jouhet, J.; Mouillac, B.; et al. The Ligand-Bound State of a G Protein-Coupled Receptor Stabilizes the Interaction of Functional Cholesterol Molecules. J. Lipid Res. 2021, 62, 100059. [Google Scholar] [CrossRef]
- Marino, K.A.; Wade, R.C.; Prada-Gracia, D.; Provasi, D.; Filizola, M. Impact of Lipid Composition and Receptor Conformation on the Spatio-Temporal Organization of μ-Opioid Receptors in a Multi-Component Plasma Membrane Model. PLoS Comput. Biol. 2016, 12, e1005240. [Google Scholar] [CrossRef]
- Velazhahan, V.; Ma, N.; Pándy-Szekeres, G.; Kooistra, A.J.; Lee, Y.; Gloriam, D.E.; Vaidehi, N.; Tate, C.G. Structure of the Class D GPCR Ste2 Dimer Coupled to Two G Proteins. Nature 2021, 589, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Yen, H.-Y.; Robinson, C.V.; Sansom, M.S.P. State-Dependent Lipid Interactions with the A2a Receptor Revealed by MD Simulations Using In Vivo-Mimetic Membranes. Structure 2019, 27, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Dijkman, P.M.; Castell, O.K.; Goddard, A.D.; Munoz-Garcia, J.C.; de Graaf, C.; Wallace, M.I.; Watts, A. Dynamic Tuneable G Protein-Coupled Receptor Monomer-Dimer Populations. Nat. Commun. 2018, 9, 1710. [Google Scholar] [CrossRef] [PubMed]
- Ferré, S.; Casadó, V.; Devi, L.A.; Filizola, M.; Jockers, R.; Lohse, M.J.; Milligan, G.; Pin, J.-P.; Guitart, X. G Protein–Coupled Receptor Oligomerization Revisited: Functional and Pharmacological Perspectives. Pharmacol. Rev. 2014, 66, 413–434. [Google Scholar] [CrossRef] [Green Version]
- Møller, T.C.; Hottin, J.; Clerté, C.; Zwier, J.M.; Durroux, T.; Rondard, P.; Prézeau, L.; Royer, C.A.; Pin, J.-P.; Margeat, E.; et al. Oligomerization of a G Protein-Coupled Receptor in Neurons Controlled by Its Structural Dynamics. Sci. Rep. 2018, 8, 10414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.J.Y.; Gabriel, F.; Tandale, A.; Nietlispach, D. Structure and Dynamics of GPCRs in Lipid Membranes: Physical Principles and Experimental Approaches. Molecules 2020, 25, 4729. [Google Scholar] [CrossRef]
- Soubias, O.; Teague, W.E.; Hines, K.G.; Gawrisch, K. The Role of Membrane Curvature Elastic Stress for Function of Rhodopsin-like G Protein-Coupled Receptors. Biochimie 2014, 107, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Smith, J.C. Biological Membrane Organization and Cellular Signaling. Chem. Rev. 2019, 119, 5849–5880. [Google Scholar] [CrossRef]
- Rajagopal, S.; Shenoy, S.K. GPCR Desensitization: Acute and Prolonged Phases. Cell. Signal. 2018, 41, 9–16. [Google Scholar] [CrossRef]
- Luttrell, L.M.; Wang, J.; Plouffe, B.; Smith, J.S.; Yamani, L.; Kaur, S.; Jean-Charles, P.-Y.; Gauthier, C.; Lee, M.-H.; Pani, B.; et al. Manifold Roles of β-Arrestins in GPCR Signaling Elucidated with SiRNA and CRISPR/Cas9. Sci. Signal. 2018, 11, eaat7650. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Sarkar, P.; Chattopadhyay, A. Metabolic Depletion of Sphingolipids Reduces Cell Surface Population of the Human Serotonin 1A Receptor Due to Impaired Trafficking. ACS Chem. Neurosci. 2021, 12, 1189–1196. [Google Scholar] [CrossRef]
- Shaik, F.A.; Chelikani, P. Differential Effects of Membrane Sphingomyelin and Cholesterol on Agonist-Induced Bitter Taste Receptor T2R14 Signaling. Mol. Cell. Biochem. 2020, 463, 57–66. [Google Scholar] [CrossRef]
- Ingólfsson, H.I.; Melo, M.N.; van Eerden, F.J.; Arnarez, C.; Lopez, C.A.; Wassenaar, T.A.; Periole, X.; de Vries, A.H.; Tieleman, D.P.; Marrink, S.J. Lipid Organization of the Plasma Membrane. J. Am. Chem. Soc. 2014, 136, 14554–14559. [Google Scholar] [CrossRef]
- Sengupta, D.; Prasanna, X.; Mole, M.; Chattopadhyay, A. Exploring GPCR−Lipid Interactions by Molecular Dynamics Simulations: Excitements, Challenges, and the Way Forward. J. Phys. Chem. B 2018, 122, 5727–5737. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Dichlberger, A.; Martinez-Seara, H.; Nyholm, T.K.M.; Li, S.; Kim, Y.A.; Vattulainen, I.; Ikonen, E.; Blom, T. A Ceramide-Regulated Element in the Late Endosomal Protein LAPTM4B Controls Amino Acid Transporter Interaction. ACS Central Sci. 2018, 4, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.; Isas, J.M.; Langen, R. Annexin B12 Trimer Formation Is Governed by a Network of Protein–protein and Protein–lipid Interactions. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.A.; Haddad, B.G.; Dolan, K.A.; Myers, J.B.; Yoshioka, C.C.; Copperman, J.; Zuckerman, D.M.; Reichow, S.L. Connexin-46/50 in a Dynamic Lipid Environment Resolved by CryoEM at 1.9 Å. Nat. Commun. 2020, 11, 4331. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Membrane Protein | |||||||
|---|---|---|---|---|---|---|---|
| Respiratory Complexes | 2° Active Transporters | ATP-Dependent Pumps/Transporters | Channels | GPCRs | Other | ||
| Lipid entity | Cholesterol | Activity regulation [8,9] | Activity and changes in conformation [10], Modulation of substrate binding affinity [11] | Channel inhibition [12] | Activation [13], Signalling [14,15], Stability [16,17], Allosteric regulation [18], Lower affinity ligand binding [17], Oligomerisation [19,20,21,22,23] | ||
| PA | Activity [24] | Stabilisation [25] | |||||
| PE | Dimer formation and function [26], Dimer formation [27], conformer stability [28] | Conformer stabilisation and channel desensitisation [29] | Agonist and antagonist binding affinities [30], Increase G protein coupling [31,32] | Protein assembly [33] | |||
| PC | Dimerisation [34], β-arrestin interaction and function [35], Increase G protein coupling [31] | ||||||
| PG | Oligomerisation and function [36] | Conformer stabilisation and channel desensitisation [37] | Increase G protein coupling [30,38], Active conformer stability [39], β-arrestin interaction and function [35], Decrease G protein coupling [31] | ||||
| PS | Dimer formation [27], | Stabilisation [25], Dimerisation [34], Decrease G protein coupling [31] | |||||
| PI | Dimer formation and function [26], Possible stabilisation [27] | ||||||
| PtdIns(4,5)P2 | Possible dimer stabilisation [40,41], | Activation [42,43], Inactive conformer stability [44] | Active conformer stability [25,45], Increase G protein interaction [25], β-arrestin interaction and complex stability [46], Important for G protein recruitment [47] | ||||
| Glycolipids and sphingolipids | Ligand binding [48,49], Signalling [48], Allostery [47] | ||||||
| Cardiolipin | Oligomer stabilisation [50] | Oligomerisation [51], Allosteric regulation [52] | |||||
| DHA and unsaturated tails | Active conformer stability [53,54], Increase G protein coupling [53], Partitioning to lipid rafts [55], Oligomerisation [34,56] | ||||||
| Saturated tails | Dimerisation and function [57] | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renard, K.; Byrne, B. Insights into the Role of Membrane Lipids in the Structure, Function and Regulation of Integral Membrane Proteins. Int. J. Mol. Sci. 2021, 22, 9026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169026
Renard K, Byrne B. Insights into the Role of Membrane Lipids in the Structure, Function and Regulation of Integral Membrane Proteins. International Journal of Molecular Sciences. 2021; 22(16):9026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169026
Chicago/Turabian StyleRenard, Kenta, and Bernadette Byrne. 2021. "Insights into the Role of Membrane Lipids in the Structure, Function and Regulation of Integral Membrane Proteins" International Journal of Molecular Sciences 22, no. 16: 9026. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22169026