
Retinoprotective Effects of PACAP Eye Drops in Microbead-Induced Glaucoma Model in Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
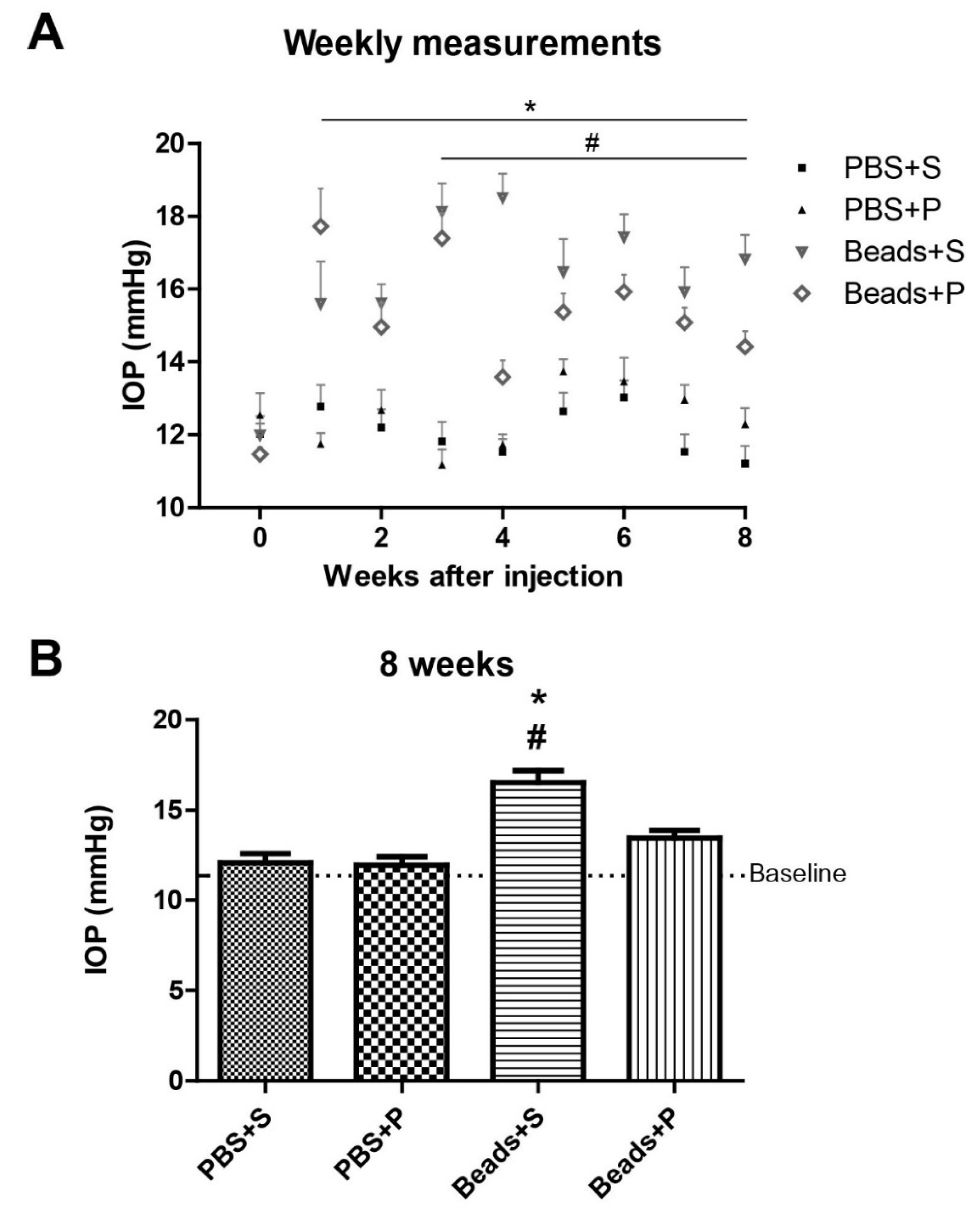
2.1. Effect of PACAP Eye Drops on IOP
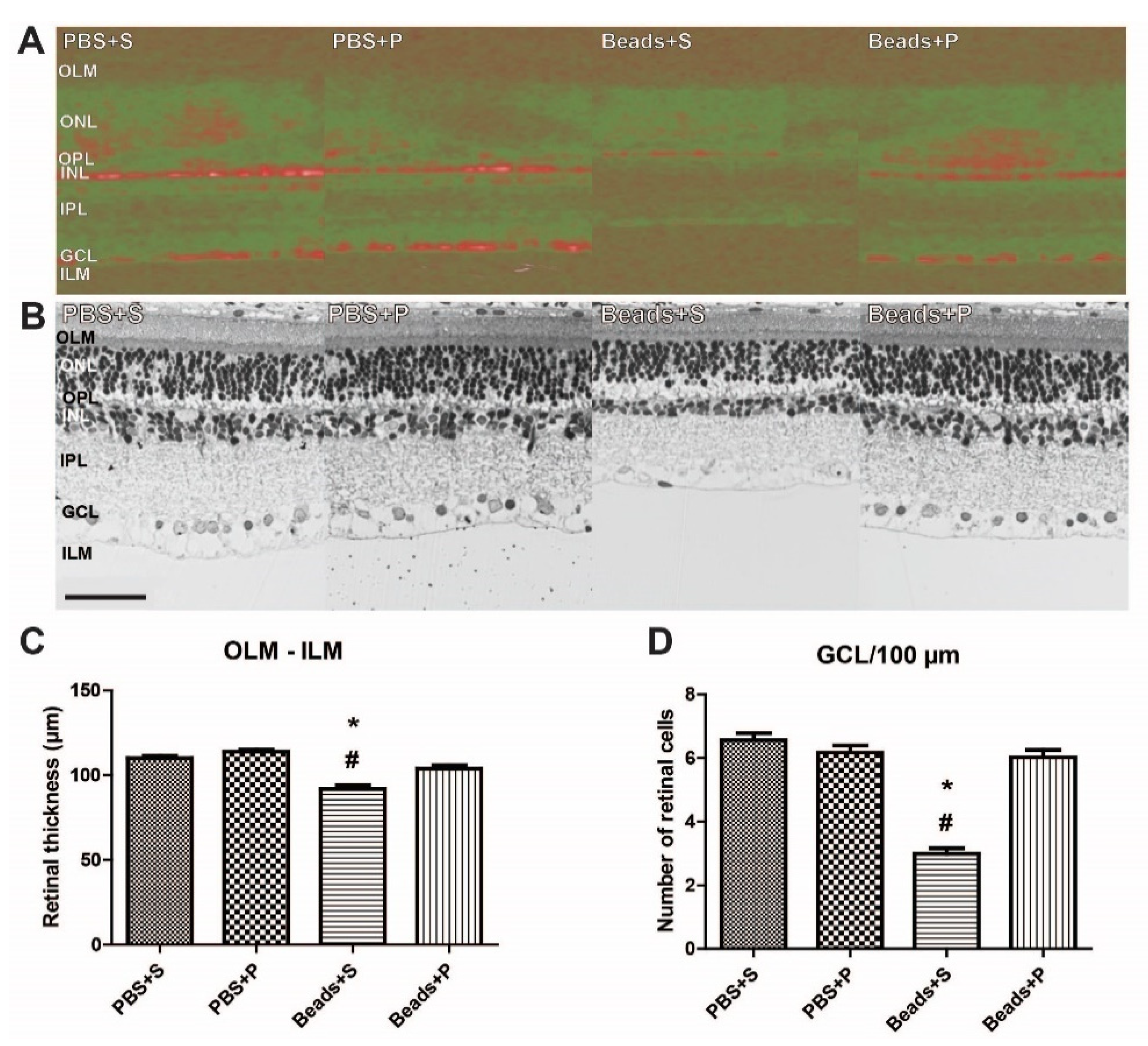
2.2. Effects of PACAP1-38 Eye Drops Treatment on Histological Changes of the Retina
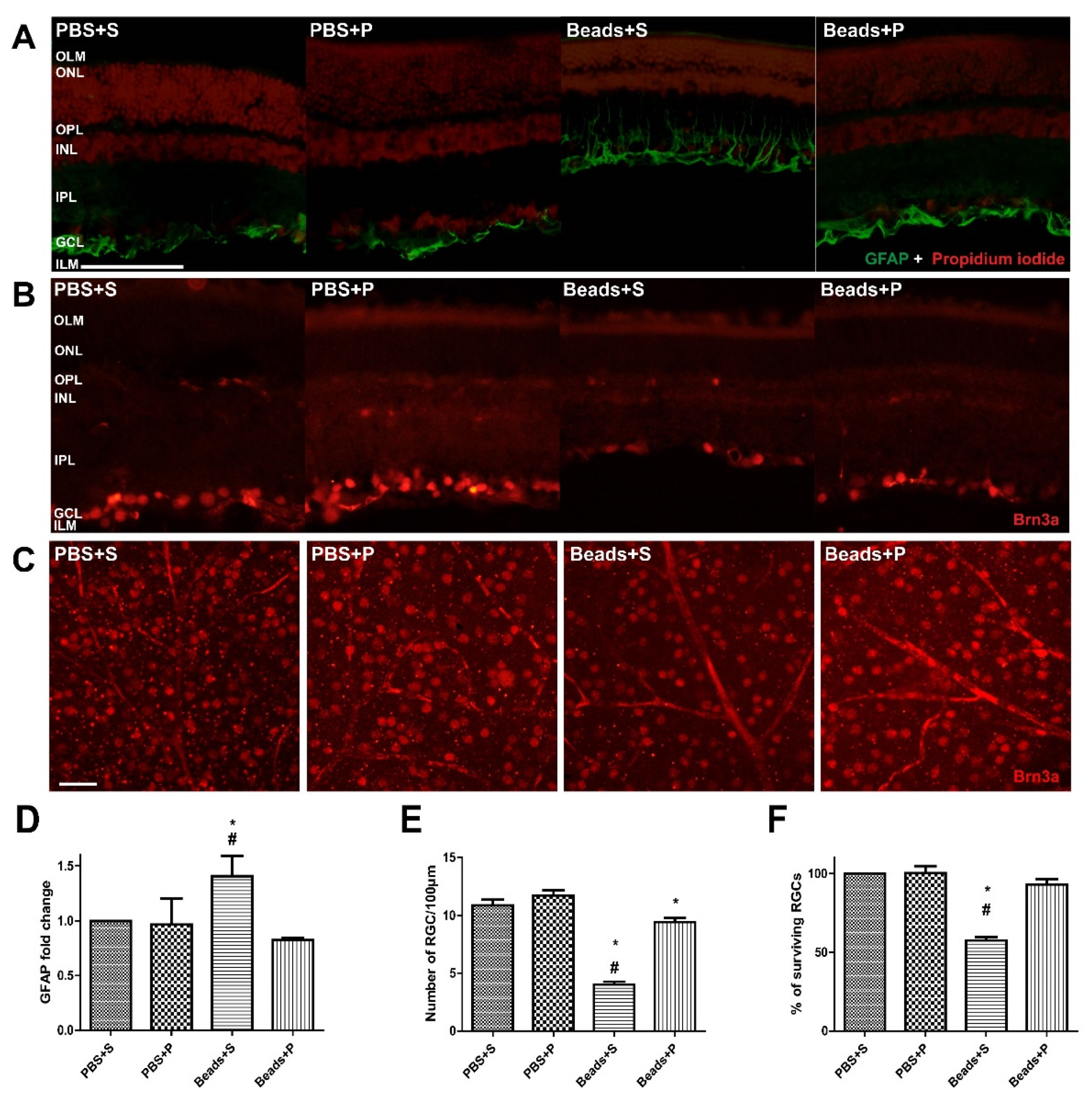
2.3. Effects of PACAP1-38 Treatment on Immunohistochemical Changes
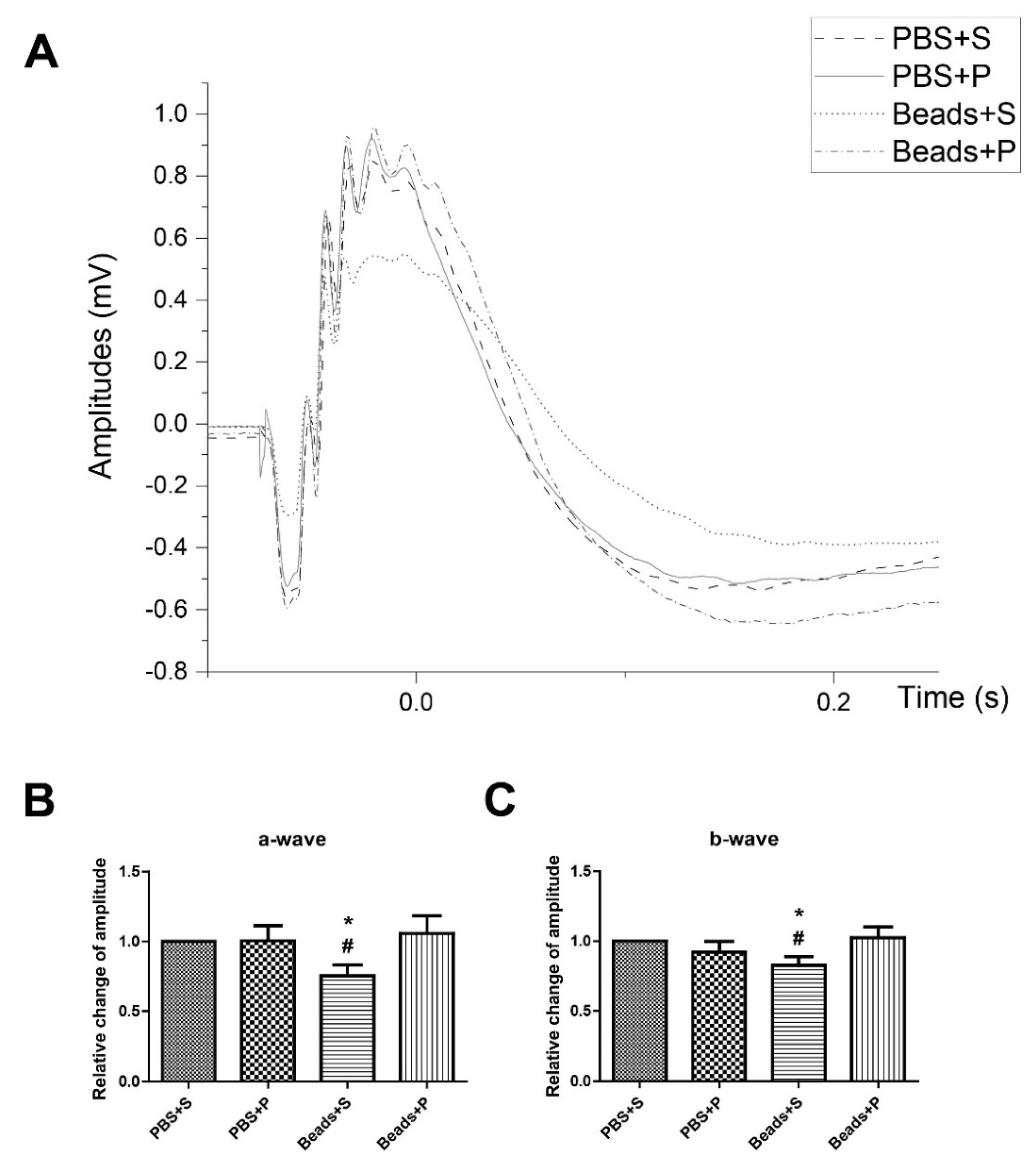
2.4. Protective Effect of PACAP1-38 Eye Drops on Visual Responses after Ocular Hypertension
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Administration of Microbeads
4.3. IOP Measurement
4.4. Eye Drops Treatment
4.5. Optical Coherence Tomography Examination
4.6. Morphological and Morphometric Analysis
4.7. Immunohistochemistry
4.8. Retinal Whole Mounts
4.9. Electroretinography
4.10. Statistical Analysis
5. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mantravadi, A.V.; Vadhar, N. Glaucoma. Prim. Care Clin. Off. Pract. 2015, 42, 437–449. [Google Scholar] [CrossRef]
- Kaufman, P.L.; Rasmussen, C.A. Advances in glaucoma treatment and management: Outflow drugs. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2495–2500. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Huang, P.; Yu, H.; Lin, Z.; Liu, X.; Shen, X.; Guo, L.; Zhong, Y. Extracellular signal-regulated kinase 1/2 pathway is insufficiently involved in the neuroprotective effect by hydrogen sulfide supplement in experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4346–4359. [Google Scholar] [CrossRef]
- Wei, X.; Cho, K.S.; Thee, E.F.; Jager, M.J.; Chen, D.F. Neuroimmflammation and microglia in glaucoma—Time for a paradigm shift. J. Neurosci. Res. 2019, 176, 100–106. [Google Scholar]
- Wang, M.; Zheng, Y. Oxidative stress and antioxidants in the trabecular meshwork. PeerJ 2019, 7, e8121. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.R.; Inman, D.M.; Calkins, D.J.; Horner, P.J.; Vetter, M.L. Microarray analysis of retinal gene expression in the DBA/2J model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2006, 47, 977–985. [Google Scholar] [CrossRef]
- Rieck, J. The pathogenesis of glaucoma in the interplay with the immune system. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2393–2409. [Google Scholar] [CrossRef] [Green Version]
- Atlasz, T.; Vaczy, A.; Werling, D.; Kiss, P.; Tamas, A.; Kovacs, K.; Fabian, E.; Kvarik, T.; Mammel, B.; Danyadi, B.; et al. Protective effects of PACAP in the retina. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP; Reglodi, D., Tamas, A., Eds.; Springer: Cham, Switzerland, 2016; pp. 501–527. [Google Scholar]
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 Years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Amin, F.M.; Schytz, H.W. Transport of the pituitary adenylate cyclase-activating polypeptide across the blood-brain barrier: Implications for migraine. J. Headache Pain 2018, 19, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Kiss, P.; Lubics, A.; Tamas, A. Review on the protective effects of PACAP in models of neurodegenerative diseases in vitro and in vivo. Curr. Pharm. Des. 2011, 17, 962–972. [Google Scholar] [CrossRef]
- Toth, D.; Tamas, A.; Reglodi, D. The neuroprotective and biomarker potential of PACAP in human traumatic brain injury. Int. J. Mol. Sci. 2020, 21, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamachi, T.; Matkovits, A.; Seki, T.; Shioda, S. Distribution and protective function of pituitary adenylate cyclase-activating polypeptide in the retina. Front. Endocrinol. 2012, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shioda, S.; Takenoya, F.; Wada, N.; Hirabayashi, T.; Seki, T.; Nakamachi, T. Pleiotropic and retinoprotective functions of PACAP. Anat. Sci. Int. 2016, 91, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.; Reglodi, D.; Banks, W.A.; Salameh, T.S.; Kovacs, K.; Kvarik, T.; Vaczy, A.; Kovacs, L.; Mayer, F.; Danyadi, B.; et al. Ocular delivery of PACAP1-27 protects the retina from ischemic damage in rodents. Investig. Opthalmol. Vis. Sci. 2016, 57, 6683–6691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaczy, A.; Reglodi, D.; Somoskeoy, T.; Kovacs, K.; Lokos, E.; Szabo, E.; Tamas, A.; Atlasz, T. The protective role of PAC1-receptor agonist maxadilan in BCCAO-induced retinal degeneration. J. Mol. Neurosci. 2016, 60, 186–194. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Bucolo, C.; D’Agata, V. Protective effect of PACAP-38 on retinal pigmented epithelium in an in vitro and in vivo model of diabetic retinopathy through EGFR-dependent mechanism. Peptides 2019, 119, 170108. [Google Scholar] [CrossRef] [PubMed]
- Vaczy, A.; Kovari, P.; Kovacs, K.; Farkas, K.; Szabo, E.; Kvarik, T.; Kocsis, B.; Fulop, B.; Atlasz, T.; Reglodi, D. Protective role of endogenous PACAP in inflammation-induced retinal degeneration. Curr. Pharm. Des. 2018, 24, 3534–3542. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, G.; D’Amico, A.G.; Amenta, A.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. Protective effect of PACAP against ultraviolet B radiation-induced human corneal endothelial cell injury. Neuropeptides 2020, 79, 101978. [Google Scholar] [CrossRef]
- Kvarik, T.; Mammel, B.; Reglodi, D.; Kovacs, K.; Werling, D.; Bede, B.; Vaczy, A.; Fabian, E.; Toth, G.; Kiss, P.; et al. PACAP is protective in a rat model of retinopathy of prematurity. J. Mol. Neurosci. 2016, 60, 179–185. [Google Scholar] [CrossRef]
- Szabadfi, K.; Atlasz, T.; Kiss, P.; Reglodi, D.; Szabo, A.; Kovacs, K.; Szalontai, B.; Setalo, G.; Banki, E.; Csanaky, K.; et al. Protective effects of the neuropeptide PACAP in diabetic retinopathy. Cell Tissue Res. 2012, 348, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.; Banks, W.; Salameh, T.; Kvarik, T.; Kovacs, L.; Vaczy, A.; Szabo, E.; Mayer, F.; Varga, R.; Tamas, A.; et al. Passage through the ocular barriers and beneficial effects in retinal ischemia of topical application of PACAP1-38 in rodents. Int. J. Mol. Sci. 2017, 18, 675. [Google Scholar] [CrossRef] [Green Version]
- Reglodi, D.; Atlasz, T.; Jungling, A.; Szabo, E.; Kovari, P.; Manavalan, S.; Tamas, A. Alternative routes of administration of the neuroprotective Pituitary Adenylate Cyclase Activating Polypeptide. Curr. Pharm. Des. 2019, 24, 3892–3904. [Google Scholar] [CrossRef] [PubMed]
- Urcola, J.H.; Hernández, M.; Vecino, E. Three experimental glaucoma models in rats: Comparison of the effects of intraocular pressure elevation on retinal ganglion cell size and death. Exp. Eye Res. 2006, 83, 429–437. [Google Scholar] [CrossRef]
- Sappington, R.M.; Carlson, B.J.; Crish, S.D.; Calkins, D.J. The microbead occlusion model: A paradigm for induced ocular hypertension in rats and mice. Investig. Ophthalmol. Vis. Sci. 2010, 51, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Atlasz, T.; Szabadfi, K.; Kiss, P.; Marton, Z.; Griecs, M.; Hamza, L.; Gaal, V.; Biro, Z.; Tamas, A.; Hild, G.; et al. Effects of PACAP in UV-A radiation-induced retinal degeneration models in rats. J. Mol. Neurosci. 2011, 43, 51–57. [Google Scholar] [CrossRef]
- Kvarik, T.; Reglodi, D.; Werling, D.; Vaczy, A.; Kovari, P.; Szabo, E.; Kovacs, K.; Hashimoto, H.; Ertl, T.; Gyarmati, J.; et al. The protective effects of endogenous PACAP in oxygen-induced retinopathy. J. Mol. Neurosci. 2021. [Google Scholar] [CrossRef]
- D’Amico, A.G.; Maugeri, G.; Musumeci, G.; Reglodi, D.; D’Agata, V. PACAP and NAP: Effect of two functionally related peptides in diabetic retinopathy. J. Mol. Neurosci. 2021, 71, 1525–1535. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Kanamori, A.; Nakamura, M.; Negi, A. Rat chronic glaucoma model induced by intracameral injection of microbeads suspended in sodium sulfate-sodium hyaluronate. Jpn. J. Ophthalmol. 2014, 58, 290–297. [Google Scholar] [CrossRef]
- Shim, M.S.; Kim, K.Y.; Ju, W.K. Role of cyclic AMP in the eye with glaucoma. BMB Rep. 2017, 50, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Meyer, D.K. The effects of PACAP on neural cell proliferation. Regul. Pept. 2006, 137, 50–57. [Google Scholar] [CrossRef]
- Rao, P.V.; Pattabiraman, P.P.; Kopczynski, C. Role of the Rho GTPase/Rho kinase signaling pathway in pathogenesis and treatment of glaucoma: Bench to bedside research. Exp. Eye Res. 2017, 158, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Hu, F.; Wang, M.; Gao, F.; Xu, P.; Xing, C.; Sun, X.; Zhang, S.; Wu, J. Comparative analysis of retinal ganglion cell damage in three glaucomatous rat models. Exp. Eye Res. 2018, 172, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Dai, L.; Xi, X.T.; Chen, Q.B.; An, M.X.; Li, Y. Sensitized heat shock protein 27 induces retinal ganglion cells apoptosis in rat glaucoma model. Int. J. Ophthalmol. 2020, 13, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Itoh, H.; Nakamachi, T.; Endo, K.; Wada, Y.; Nakamura, K.; Shioda, S. Suppression of rat retinal ganglion cell death by PACAP following transient ischemia induced by high intraocular pressure. J. Mol. Neurosci. 2011, 43, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Soto, I.; Pease, M.E.; Son, J.L.; Shi, X.; Quigley, H.A.; Marsh-Armstrong, N. Retinal ganglion cell loss in a rat ocular hypertension model is sectorial and involves early optic nerve axon loss. Investig. Opthalmol.Vis. Sci. 2011, 52, 434. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, M.; Seki, T.; Shinohara, Y.; Taki, C.; Nishimura, S.; Takaki, A.; Shioda, S. Pituitary adenylate cyclase-activating peptide (PACAP) stimulates production of interleukin-6 in rat Müller cells. Peptides 2006, 27, 1871–1876. [Google Scholar] [CrossRef]
- Smedowski, A.; Pietrucha-Dutczak, M.; Kaarniranta, K.; Lewin-Kowalik, J. A rat experimental model of glaucoma incorporating rapid-onset elevation of intraocular pressure. Sci. Rep. 2014, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Akopian, A.; Kumar, S.; Ramakrishnan, H.; Viswanathan, S.; Bloomfield, S.A. Amacrine cells coupled to ganglion cells via gap junctions are highly vulnerable in glaucomatous mouse retinas. J. Comp. Neurol. 2019, 527, 159–173. [Google Scholar] [CrossRef]
- Danyadi, B.; Szabadfi, K.; Reglodi, D.; Mihalik, A.; Danyadi, T.; Kovacs, Z.; Batai, I.; Tamas, A.; Kiss, P.; Toth, G.; et al. PACAP application improves functional outcome of chronic retinal ischemic injury in rats-evidence from electroretinographic measurements. J. Mol. Neurosci. 2014, 54, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, L.; Slaughter, M.M. Comparison of the waveforms of the ON bipolar neuron and the b -wave of the electroretinogram. Vision Res. 1993, 33, 2431–2435. [Google Scholar] [CrossRef]
- Kido, N.; Tanihara, H.; Honjo, M.; Inatani, M.; Tatsuno, T.; Nakayama, C.; Honda, Y. Neuroprotective effects of brain-derived neurotrophic factor in eyes with NMDA-induced neuronal death. Brain Res. 2000, 884, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Wójcik-Gryciuk, A.; Gajewska-Woźniak, O.; Kordecka, K.; Boguszewski, P.M.; Waleszczyk, W.; Skup, M. Neuroprotection of retinal ganglion cells with AAV2-BDNF pretreatment restoring normal TrkB receptor protein levels in glaucoma. Int. J. Mol. Sci. 2020, 21, 6262. [Google Scholar] [CrossRef] [PubMed]
- Zink, M.; Otto, C.; Zörner, B.; Zacher, C.; Schütz, G.; Henn, F.A.; Gass, P. Reduced expression of brain-derived neurotrophic factor in mice deficient for pituitary adenylate cyclase activating polypeptide type-I-receptor. Neurosci. Lett. 2004, 360, 106–108. [Google Scholar] [CrossRef]
- Brown, D.; Tamas, A.; Reglödi, D.; Tizabi, Y. PACAP protects against salsolinol-induced toxicity in dopaminergic SH-SY5Y cells: Implication for Parkinson’s Disease. J. Mol. Neurosci. 2013, 50, 600–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frechilla, D.; García-Osta, A.; Palacios, S.; Cenarruzabeitia, E.; Del Río, J. BDNF mediates the neuroprotective effect of PACAP-38 on rat cortical neurons. NeuroReport 2001, 12, 919–923. [Google Scholar] [CrossRef]
- Liu, L.-F.; Huang, C.-K.; Zhang, M.-Z. Reliability of Tonolab measurements in rats. Int. J. Ophthalmol. 2014, 7, 930–934. [Google Scholar] [PubMed]
- Pease, M.E.; Cone, F.E.; Gelman, S.; Son, J.L.; Quigley, H.A. Calibration of the TonoLab tonometer in mice with spontaneous or experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Jozsa, R.; Hollosy, T.; Tamas, A.; Toth, G.; Lengvari, I.; Reglodi, D. Pituitary adenylate cyclase activating polypeptide plays a role in olfactory memory formation in chicken. Peptides 2005, 26, 2344–2350. [Google Scholar] [CrossRef] [PubMed]
- Nair, G.; Kim, M.; Nagaoka, T.; Olson, D.E.; Thulé, P.M.; Pardue, M.T.; Duong, T.Q. Effects of common anesthetics on eye movement and electroretinogram. Doc. Ophthalmol. 2011, 122, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.E.; Tribble, J.R. Microbead models in glaucoma. Exp. Eye Res. 2015, 141, 9–14. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabo, E.; Patko, E.; Vaczy, A.; Molitor, D.; Csutak, A.; Toth, G.; Reglodi, D.; Atlasz, T. Retinoprotective Effects of PACAP Eye Drops in Microbead-Induced Glaucoma Model in Rats. Int. J. Mol. Sci. 2021, 22, 8825. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168825
Szabo E, Patko E, Vaczy A, Molitor D, Csutak A, Toth G, Reglodi D, Atlasz T. Retinoprotective Effects of PACAP Eye Drops in Microbead-Induced Glaucoma Model in Rats. International Journal of Molecular Sciences. 2021; 22(16):8825. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168825
Chicago/Turabian StyleSzabo, Edina, Evelin Patko, Alexandra Vaczy, Dorottya Molitor, Adrienne Csutak, Gabor Toth, Dora Reglodi, and Tamas Atlasz. 2021. "Retinoprotective Effects of PACAP Eye Drops in Microbead-Induced Glaucoma Model in Rats" International Journal of Molecular Sciences 22, no. 16: 8825. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168825