
Interaction between SCP3 and JAB1 Confers Cancer Therapeutic Resistance and Stem-like Properties through EGF Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
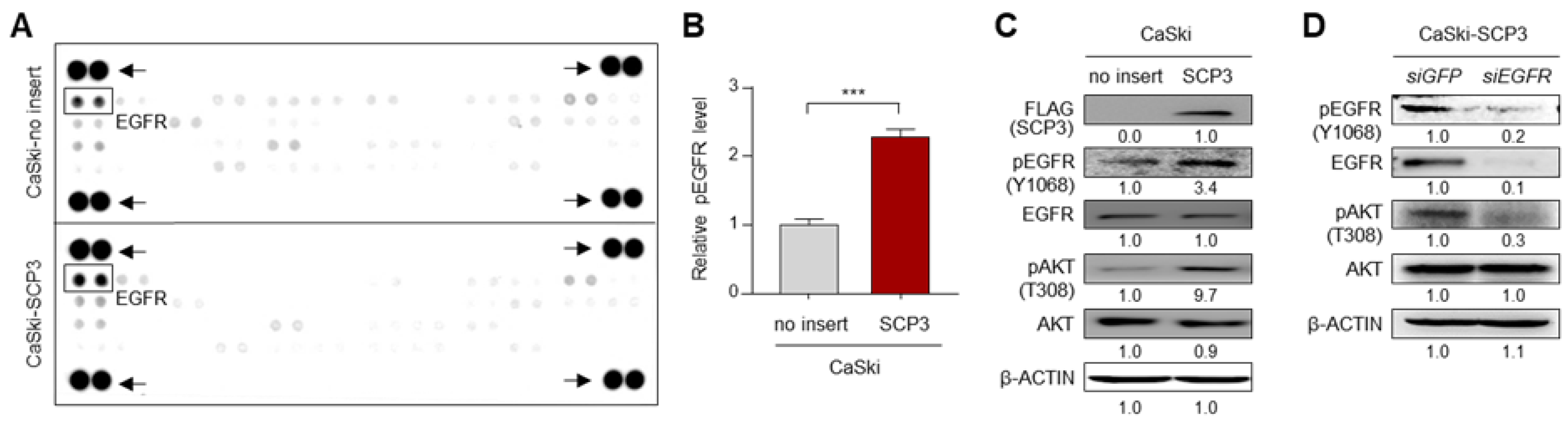
2.1. SCP3 Activates AKT via EGFR Signaling
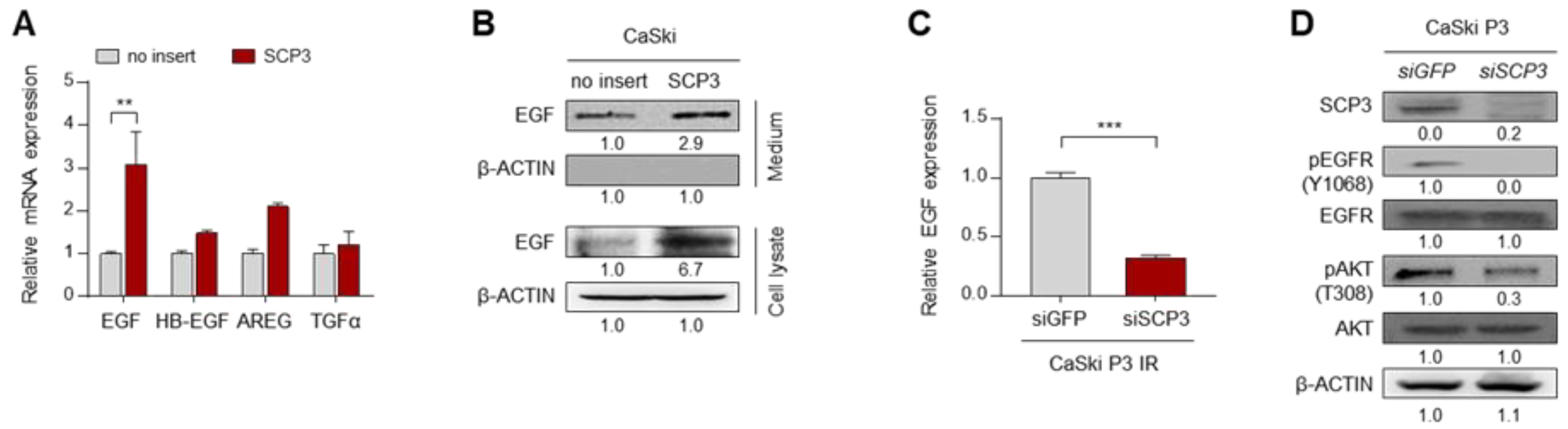
2.2. SCP3 Triggers EGFR Signaling through Transcriptional Activation of EGF
2.3. Neutralization of Secreted EGF by Its Specific Monoclonal Antibody Reverses SCP3-Mediated Aggressive Phenotypes
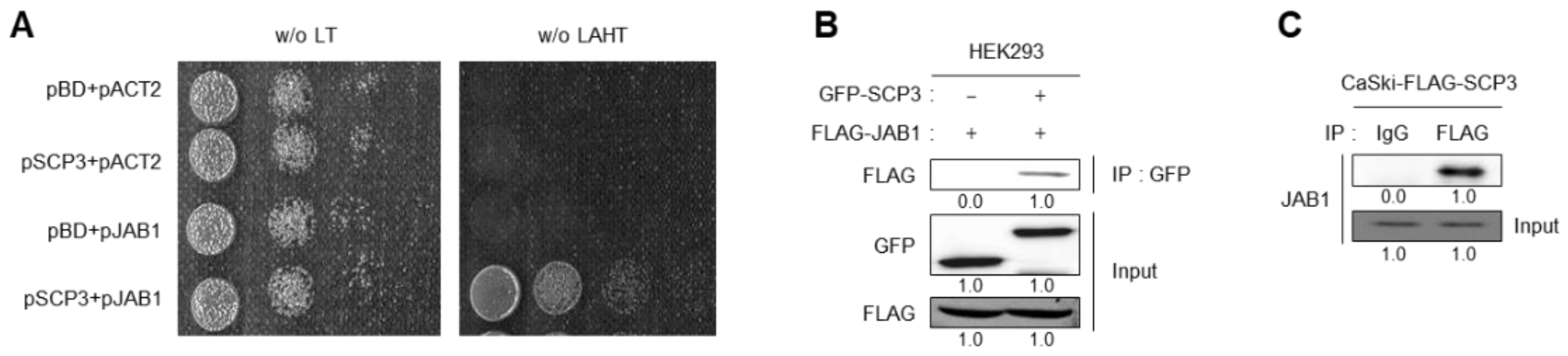
2.4. SCP3 Physically Interacts with JAB1
2.5. SCP3 Induces EGF Transcription through Physical Interaction with JAB1
3. Discussion
4. Materials and Methods
4.1. Cell Lines Reagents
4.2. DNA Constructs
4.3. Site-Directed Mutagenesis
4.4. siRNA Constructs
4.5. Antibody Arrays of Phospho-RTK
4.6. Real-Time Quantitative RT-PCR
4.7. Tumor Sphere-Forming Assay
4.8. Granzyme B-Mediated Apoptosis Assay
4.9. Trypan Blue Exclusion Assay
4.10. Yeast Two-Hybrid (Y2H) Assay
4.11. Immunoprecipitation
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Di Carlo, A.D.; Travia, G.; De Felici, M. The meiotic specific synaptonemal complex protein SCP3 is expressed by female and male primordial germ cells of the mouse embryo. Int. J. Dev. Biol. 2000, 44, 241–244. [Google Scholar] [PubMed]
- Chung, J.Y.; Kitano, H.; Takikita, M.; Cho, H.; Noh, K.H.; Kim, T.W.; Ylaya, K.; Hanaoka, J.; Fukuoka, J.; Hewitt, S.M. Synaptonemal complex protein 3 as a novel prognostic marker in early stage non-small cell lung cancer. Hum. Pathol. 2013, 44, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Noh, K.H.; Chung, J.Y.; Takikita, M.; Chung, E.J.; Kim, B.W.; Hewitt, S.M.; Kim, T.W.; Kim, J.H. Synaptonemal complex protein 3 is a prognostic marker in cervical cancer. PLoS ONE 2014, 9, e98712. [Google Scholar] [CrossRef]
- Mobasheri, M.B.; Shirkoohi, R.; Modarressi, M.H. Synaptonemal Complex Protein 3 Transcript Analysis in Breast Cancer. Iran. J. Public Health 2016, 45, 1618–1624. [Google Scholar]
- Kang, T.H.; Noh, K.H.; Kim, J.H.; Bae, H.C.; Lin, K.Y.; Monie, A.; Pai, S.I.; Hung, C.F.; Wu, T.C.; Kim, T.W. Ectopic expression of X-linked lymphocyte-regulated protein pM1 renders tumor cells resistant to antitumor immunity. Cancer Res. 2010, 70, 3062–3070. [Google Scholar] [CrossRef] [Green Version]
- Kitano, H.; Chung, J.Y.; Noh, K.H.; Lee, Y.H.; Kim, T.W.; Lee, S.H.; Eo, S.H.; Cho, H.J.; Choi, C.H.; Inoue, S.; et al. Synaptonemal complex protein 3 is associated with lymphangiogenesis in non-small cell lung cancer patients with lymph node metastasis. J. Transl. Med. 2017, 15, 138. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.J.; Cho, H.; Kim, S.; Noh, K.H.; Song, K.H.; Lee, H.J.; Woo, S.R.; Kim, S.; Choi, C.H.; Chung, J.Y.; et al. Targeting Cyclin D-CDK4/6 Sensitizes Immune-Refractory Cancer by Blocking the SCP3-NANOG Axis. Cancer Res. 2018, 78, 2638–2653. [Google Scholar] [CrossRef] [Green Version]
- Chauchereau, A.; Georgiakaki, M.; Perrin-Wolff, M.; Milgrom, E.; Loosfelt, H. JAB1 interacts with both the progesterone receptor and SRC-1. J. Biol. Chem. 2000, 275, 8540–8548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeger, M.; Kraft, R.; Ferrell, K.; Bech-Otschir, D.; Dumdey, R.; Schade, R.; Gordon, C.; Naumann, M.; Dubiel, W. A novel protein complex involved in signal transduction possessing similarities to 26S proteasome subunits. FASEB J. 1998, 12, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Tsuge, T.; Serino, G.; Dohmae, N.; Takio, K.; Matsui, M.; Deng, X.W. The COP9 complex is conserved between plants and mammals and is related to the 26S proteasome regulatory complex. Curr. Biol. 1998, 8, 919–922. [Google Scholar] [CrossRef] [Green Version]
- Wei, N.; Deng, X.W. Making sense of the COP9 signalosome: A regulatory protein complex conserved from Arabidopsis to human. Trends Genet. 1999, 15, 98–103. [Google Scholar] [CrossRef]
- Lee, M.H.; Zhao, R.; Phan, L.; Yeung, S.C. Roles of COP9 signalosome in cancer. Cell Cycle 2011, 10, 3057–3066. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Claret, F.X.; Zhou, F.; Pan, Y. Jab1/COPS5 as a Novel Biomarker for Diagnosis, Prognosis, Therapy Prediction and Therapeutic Tools for Human Cancer. Front. Pharmacol. 2018, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Shackleford, T.J.; Claret, F.X. JAB1/CSN5: A new player in cell cycle control and cancer. Cell Div. 2010, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Cho, H.; Hong, S.O.; Oh, S.J.; Lee, H.J.; Cho, E.; Woo, S.R.; Song, J.S.; Chung, J.Y.; Son, S.W.; et al. LC3B upregulation by NANOG promotes immune resistance and stem-like property through hyperactivation of EGFR signaling in immune-refractory tumor cells. Autophagy 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Scaltriti, M.; Baselga, J. The epidermal growth factor receptor pathway: A model for targeted therapy. Clin. Cancer Res. 2006, 12, 5268–5272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.H.; Bae, H.C.; Noh, K.H.; Song, K.H.; Ye, S.K.; Mao, C.P.; Lee, K.M.; Wu, T.C.; Kim, T.W. Gain of HIF-1alpha under normoxia in cancer mediates immune adaptation through the AKT/ERK and VEGFA axes. Clin. Cancer Res. 2015, 21, 1438–1446. [Google Scholar] [CrossRef] [Green Version]
- West, N.R.; Farnell, B.; Murray, J.I.; Hof, F.; Watson, P.H.; Boulanger, M.J. Structural and functional characterization of a triple mutant form of S100A7 defective for Jab1 binding. Protein Sci. 2009, 18, 2615–2623. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y. Cancer immunotherapy: Harnessing the immune system to battle cancer. J. Clin. Investig. 2015, 125, 3335–3337. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, R.D.; Old, L.J.; Smyth, M.J. Cancer immunoediting: Integrating immunity’s roles in cancer suppression and promotion. Science 2011, 331, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajewski, T.F.; Meng, Y.; Blank, C.; Brown, I.; Kacha, A.; Kline, J.; Harlin, H. Immune resistance orchestrated by the tumor microenvironment. Immunol. Rev. 2006, 213, 131–145. [Google Scholar] [CrossRef]
- Song, K.H.; Oh, S.J.; Kim, S.; Cho, H.; Lee, H.J.; Song, J.S.; Chung, J.Y.; Cho, E.; Lee, J.; Jeon, S.; et al. HSP90A inhibition promotes anti-tumor immunity by reversing multi-modal resistance and stem-like property of immune-refractory tumors. Nat. Commun. 2020, 11, 562. [Google Scholar] [CrossRef]
- Oh, S.J.; Lee, J.; Kim, Y.; Song, K.H.; Cho, E.; Kim, M.; Jung, H.; Kim, T.W. Far Beyond Cancer Immunotherapy: Reversion of Multi-Malignant Phenotypes of Immunotherapeutic-Resistant Cancer by Targeting the NANOG Signaling Axis. Immune Netw. 2020, 20, e7. [Google Scholar] [CrossRef]
- Noh, K.H.; Kim, B.W.; Song, K.H.; Cho, H.; Lee, Y.H.; Kim, J.H.; Chung, J.Y.; Kim, J.H.; Hewitt, S.M.; Seong, S.Y.; et al. Nanog signaling in cancer promotes stem-like phenotype and immune evasion. J. Clin. Investig. 2012, 122, 4077–4093. [Google Scholar] [CrossRef] [Green Version]
- Song, K.H.; Choi, C.H.; Lee, H.J.; Oh, S.J.; Woo, S.R.; Hong, S.O.; Noh, K.H.; Cho, H.; Chung, E.J.; Kim, J.H.; et al. HDAC1 Upregulation by NANOG Promotes Multidrug Resistance and a Stem-like Phenotype in Immune Edited Tumor Cells. Cancer Res. 2017, 77, 5039–5053. [Google Scholar] [CrossRef] [Green Version]
- Song, K.H.; Kim, J.H.; Lee, Y.H.; Bae, H.C.; Lee, H.J.; Woo, S.R.; Oh, S.J.; Lee, K.M.; Yee, C.; Kim, B.W.; et al. Mitochondrial reprogramming via ATP5H loss promotes multimodal cancer therapy resistance. J. Clin. Investig. 2018, 128, 4098–4114. [Google Scholar] [CrossRef]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. Small molecule inhibitors targeting the EGFR/ErbB family of protein-tyrosine kinases in human cancers. Pharmacol. Res. 2019, 139, 395–411. [Google Scholar] [CrossRef]
- Rho, S.B.; Lee, K.H.; Kim, J.W.; Shiba, K.; Jo, Y.J.; Kim, S. Interaction between human tRNA synthetases involves repeated sequence elements. Proc. Natl. Acad. Sci. USA 1996, 93, 10128–10133. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, S.J.; Noh, K.H.; Song, K.-H.; Kim, T.W. Interaction between SCP3 and JAB1 Confers Cancer Therapeutic Resistance and Stem-like Properties through EGF Expression. Int. J. Mol. Sci. 2021, 22, 8839. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168839
Oh SJ, Noh KH, Song K-H, Kim TW. Interaction between SCP3 and JAB1 Confers Cancer Therapeutic Resistance and Stem-like Properties through EGF Expression. International Journal of Molecular Sciences. 2021; 22(16):8839. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168839
Chicago/Turabian StyleOh, Se Jin, Kyung Hee Noh, Kwon-Ho Song, and Tae Woo Kim. 2021. "Interaction between SCP3 and JAB1 Confers Cancer Therapeutic Resistance and Stem-like Properties through EGF Expression" International Journal of Molecular Sciences 22, no. 16: 8839. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168839