
Phenotypic Characterization of Immortalized Chondrocytes from a Desbuquois Dysplasia Type 1 Mouse Model: A Tool for Studying Defects in Glycosaminoglycan Biosynthesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
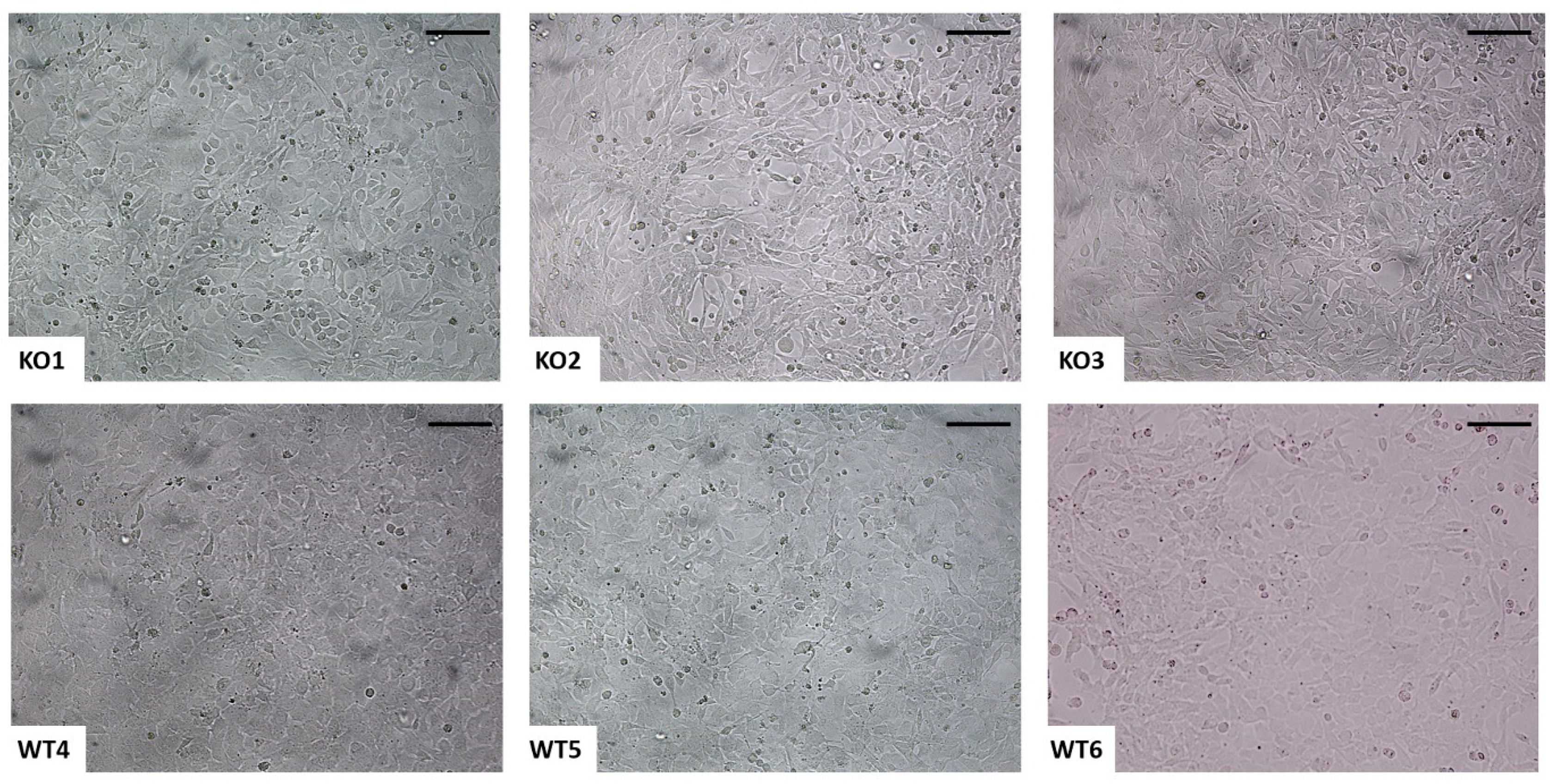
2.1. Immortalization of Murine Chondrocytes from Femoral Head Cartilage
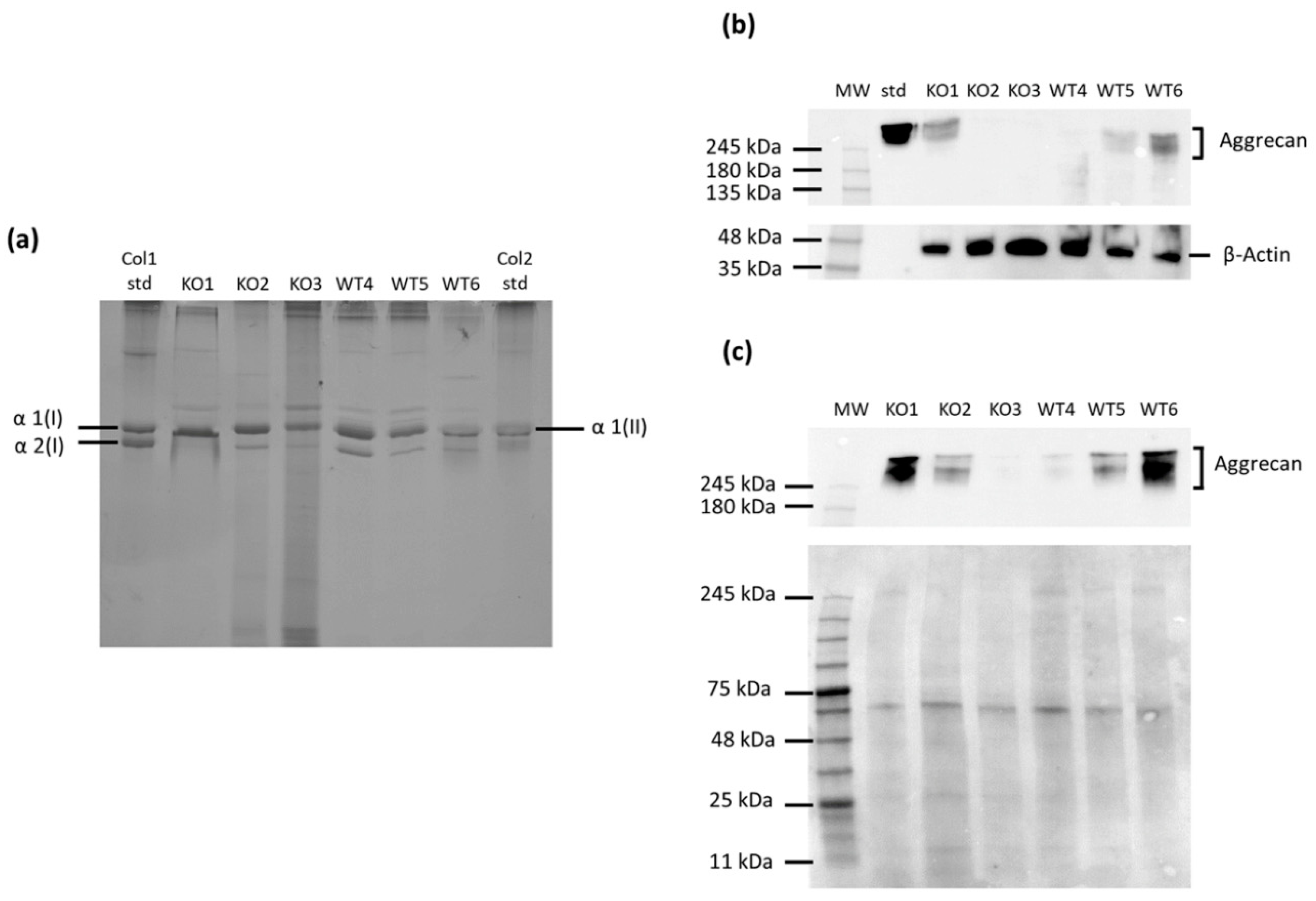
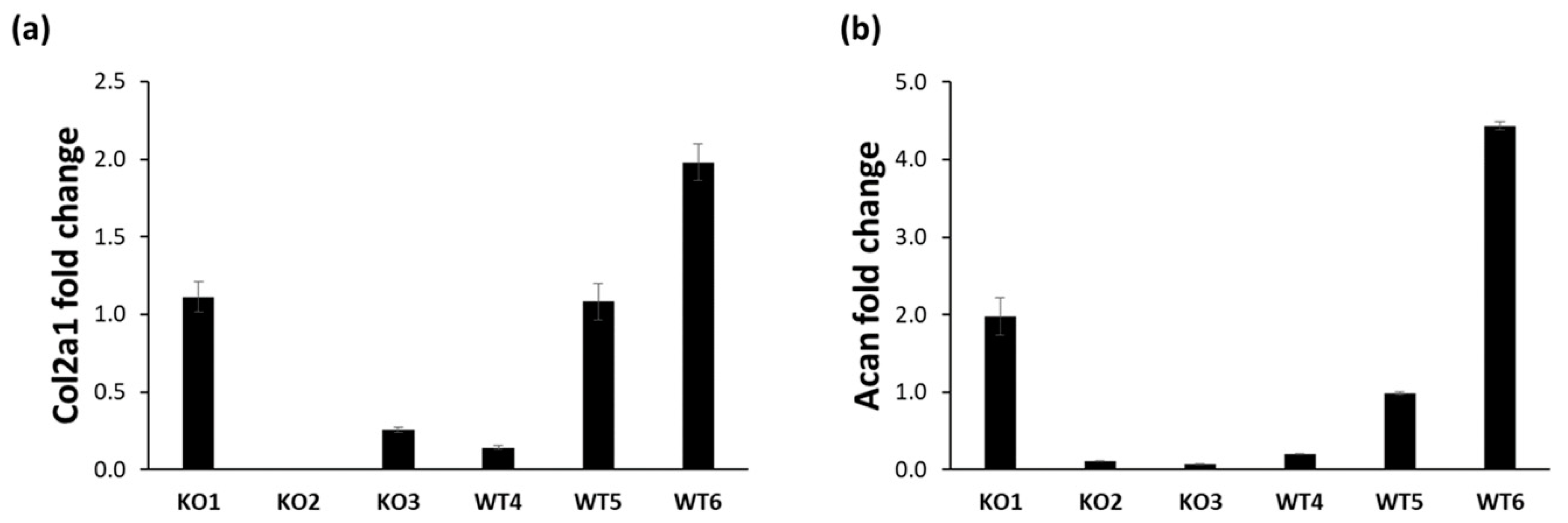
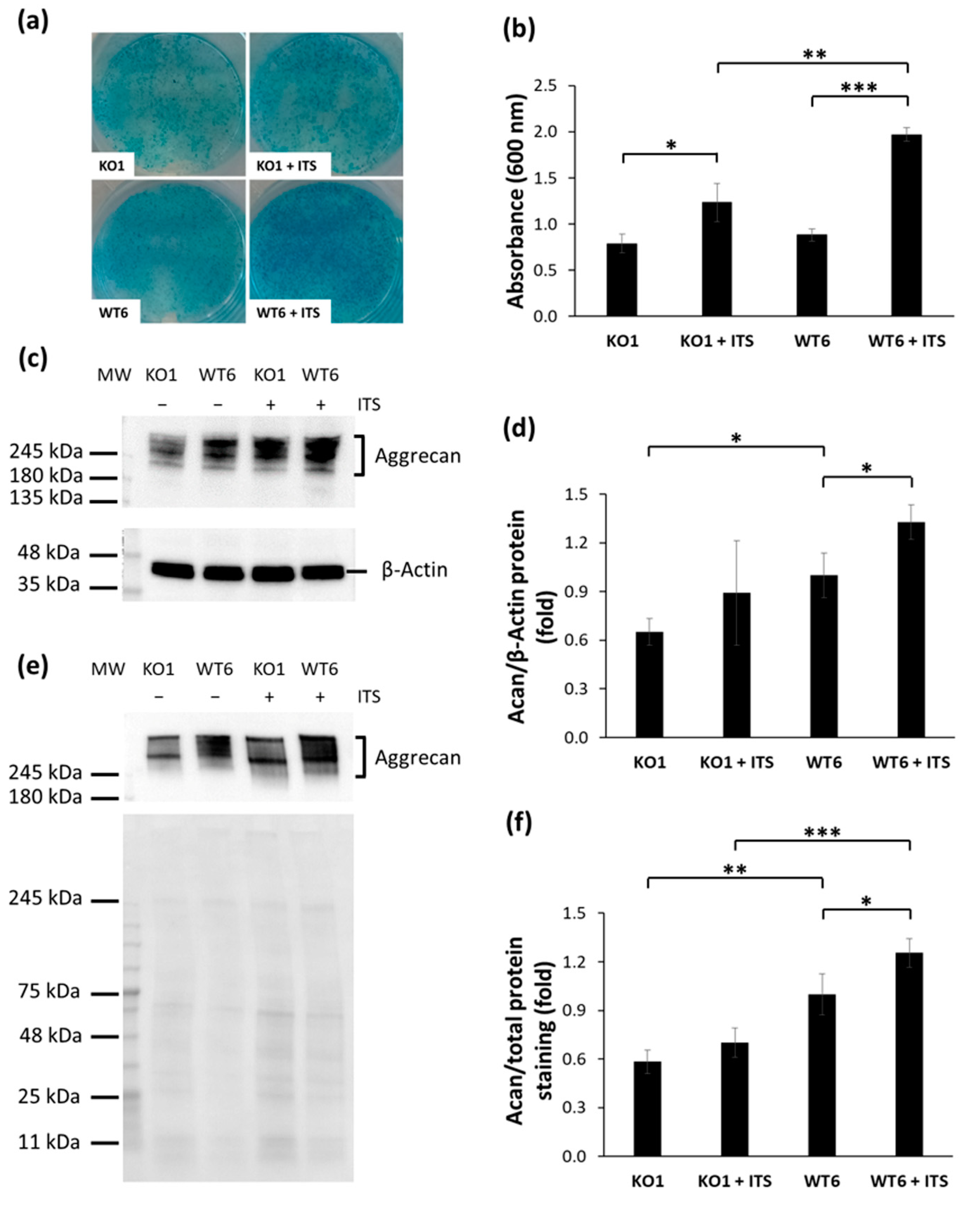
2.2. Analysis of the Cartilage Phenotype in Immortalized Chondrocytes
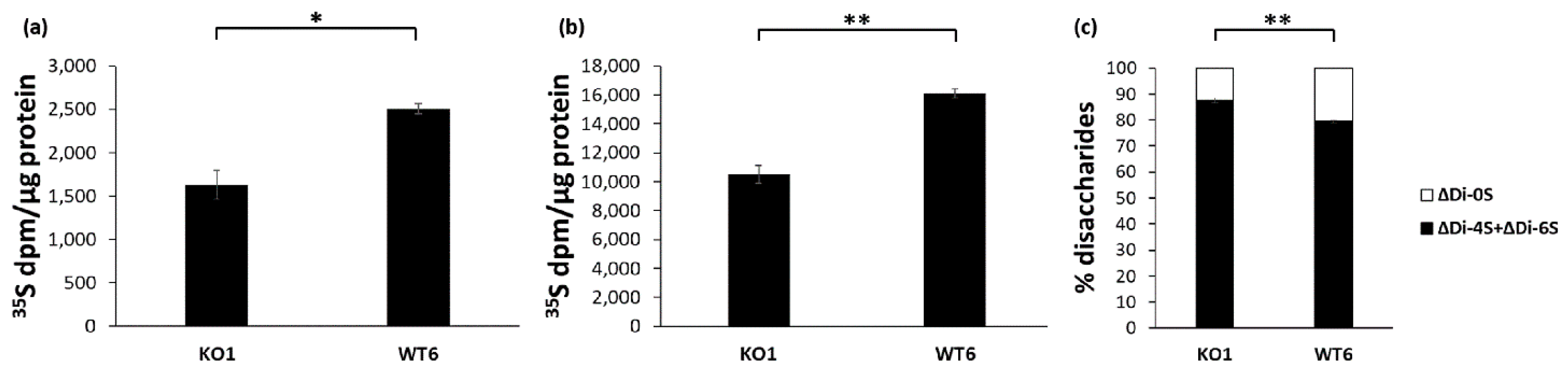
2.3. Glycosaminoglycan Synthesis and Sulfation in Immortalized Cant1 Knock-Out and Wild-Type Chondrocytes
2.4. Characterization of the Extracellular Matrix Produced by Immortalized Cells
3. Discussion
4. Materials and Methods
4.1. Animal Model and Care
4.2. Chondrocyte Isolation and Immortalization
4.3. Collagen Analysis
4.4. Real-Time PCR
4.5. Analysis of Proteoglycan Synthesis by Metabolic Labeling
4.6. Proteoglycan Sulfation Analysis
4.7. Western Blot Analysis
4.8. Alcian Blue Staining
4.9. Immunofluorescence Studies
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Acan | aggrecan |
| BCA | bicinchoninic acid |
| BMP | bone morphogenetic protein |
| BSA | bovine serum albumin |
| CANT1 | calcium activated nucleotidase 1 |
| Col2a1 | α1 chain of type II collagen |
| COMP | cartilage oligomeric matrix protein |
| DBQD1 | Desbuquois dysplasia type 1 |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ECM | extracellular matrix |
| EDTA | ethylenediaminetetraacetic acid |
| ER | endoplasmic reticulum |
| FBS | fetal bovine serum |
| GAG | glycosaminoglycan |
| Gapdh | glyceraldehyde 3-phosphate dehydrogenase |
| HPLC | high performance liquid chromatography |
| IL-1 | interleukin-1 |
| ITS | insulin-transferrin-selenium |
| MEM | minimal essential medium |
| NEM | N-ethylmaleimide |
| PBS | phosphate buffered saline |
| PCR | polymerase chain reaction |
| PG | proteoglycan |
| PMSF | phenylmethylsulfonyl fluoride |
| PVDF | polyvinylidene difluoride |
| SDS | sodium dodecyl sulfate |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TBS | Tris buffered saline |
| TEM | transmission electron microscopy |
| ΔDi-0S | 3-O-β(D-gluc-4-eneuronosyl)-N-acetylgalactosamine |
| ΔDi-4S and ΔDi-6S | derivatives of ΔDi-0S with a sulfate at the 4 or 6 position of hexosamine moiety respectively |
References
- Archer, C.W.; Francis-West, P. The chondrocyte. Int. J. Biochem. Cell Biol. 2003, 35, 401–404. [Google Scholar] [CrossRef]
- Kiani, C.; Chen, L.; Wu, Y.J.; Yee, A.J.; Yang, B.B. Structure and function of aggrecan. Cell Res. 2002, 12, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozhemyakina, E.; Lassar, A.B.; Zelzer, E. A pathway to bone: Signaling molecules and transcription factors involved in chondrocyte development and maturation. Development 2015, 142, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, Y.; Grodzinsky, A.J. Cartilage diseases. Matrix Biol. 2018, 71, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Mortier, G.R.; Cohn, D.H.; Cormier-Daire, V.; Hall, C.; Krakow, D.; Mundlos, S.; Nishimura, G.; Robertson, S.; Sangiorgi, L.; Savarirayan, R.; et al. Nosology and classification of genetic skeletal disorders: 2019 revision. Am. J. Med. Genet. A 2019, 179, 2393–2419. [Google Scholar] [CrossRef] [PubMed]
- Briggs, M.D.; Bell, P.A.; Pirog, K.A. The utility of mouse models to provide information regarding the pathomolecular mechanisms in human genetic skeletal diseases: The emerging role of endoplasmic reticulum stress (Review). Int. J. Mol. Med. 2015, 35, 1483–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, C.I.; Soler, C. Animal Models of Osteoarthritis in Small Mammals. Vet. Clin. N. Am. Exot. Anim. Pract. 2019, 22, 211–221. [Google Scholar] [CrossRef]
- Hambright, W.S.; Niedernhofer, L.J.; Huard, J.; Robbins, P.D. Murine models of accelerated aging and musculoskeletal disease. Bone 2019, 125, 122–127. [Google Scholar] [CrossRef]
- Zaucke, F.; Dinser, R.; Maurer, P.; Paulsson, M. Cartilage oligomeric matrix protein (COMP) and collagen IX are sensitive markers for the differentiation state of articular primary chondrocytes. Biochem. J. 2001, 358, 17–24. [Google Scholar] [CrossRef]
- Goldring, M.B.; Sandell, L.J.; Stephenson, M.L.; Krane, S.M. Immune interferon suppresses levels of procollagen mRNA and type II collagen synthesis in cultured human articular and costal chondrocytes. J. Biol. Chem. 1986, 261, 9049–9055. [Google Scholar] [CrossRef]
- Watt, F.M. Effect of seeding density on stability of the differentiated phenotype of pig articular chondrocytes in culture. J. Cell Sci. 1988, 89 Pt 3, 373–378. [Google Scholar] [CrossRef]
- Meretoja, V.V.; Dahlin, R.L.; Kasper, F.K.; Mikos, A.G. Enhanced chondrogenesis in co-cultures with articular chondrocytes and mesenchymal stem cells. Biomaterials 2012, 33, 6362–6369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, S.S.; Masuda, K.; Häuselmann, H.J.; Aydelotte, M.B.; Thonar, E.J. Aggrecan synthesized by mature bovine chondrocytes suspended in alginate. Identification of two distinct metabolic matrix pools. J. Biol. Chem. 1994, 269, 33021–33027. [Google Scholar] [CrossRef]
- Benya, P.D.; Shaffer, J.D. Dedifferentiated chondrocytes reexpress the differentiated collagen phenotype when cultured in agarose gels. Cell 1982, 30, 215–224. [Google Scholar] [CrossRef]
- Grigolo, B.; Lisignoli, G.; Piacentini, A.; Fiorini, M.; Gobbi, P.; Mazzotti, G.; Duca, M.; Pavesio, A.; Facchini, A. Evidence for redifferentiation of human chondrocytes grown on a hyaluronan-based biomaterial (HYAff 11): Molecular, immunohistochemical and ultrastructural analysis. Biomaterials 2002, 23, 1187–1195. [Google Scholar] [CrossRef]
- Jin, G.Z.; Kim, H.W. Efficacy of collagen and alginate hydrogels for the prevention of rat chondrocyte dedifferentiation. J. Tissue Eng. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mallein-Gerin, F.; Olsen, B.R. Expression of simian virus 40 large T (tumor) oncogene in mouse chondrocytes induces cell proliferation without loss of the differentiated phenotype. Proc. Natl. Acad. Sci. USA 1993, 90, 3289–3293. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B.; Birkhead, J.R.; Suen, L.F.; Yamin, R.; Mizuno, S.; Glowacki, J.; Arbiser, J.L.; Apperley, J.F. Interleukin-1 beta-modulated gene expression in immortalized human chondrocytes. J. Clin. Investig. 1994, 94, 2307–2316. [Google Scholar] [CrossRef]
- Mataga, N.; Tamura, M.; Yanai, N.; Shinomura, T.; Kimata, K.; Obinata, M.; Noda, M. Establishment of a novel chondrocyte-like cell line derived from transgenic mice harboring the temperature-sensitive simian virus 40 large T-antigen gene. J. Bone Miner. Res. 1996, 11, 1646–1654. [Google Scholar] [CrossRef]
- Takazawa, Y.; Nifuji, A.; Mataga, N.; Yamauchi, Y.; Kurosawa, H.; Noda, M. Articular cartilage cells immortalized by a temperature sensitive mutant of SV40 large T antigen survive and form cartilage tissue in articular cartilage environment. J. Cell Biochem. 1999, 75, 338–345. [Google Scholar] [CrossRef]
- Kokenyesi, R.; Tan, L.; Robbins, J.R.; Goldring, M.B. Proteoglycan production by immortalized human chondrocyte cell lines cultured under conditions that promote expression of the differentiated phenotype. Arch. Biochem. Biophys. 2000, 383, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.R.; Thomas, B.; Tan, L.; Choy, B.; Arbiser, J.L.; Berenbaum, F.; Goldring, M.B. Immortalized human adult articular chondrocytes maintain cartilage-specific phenotype and responses to interleukin-1beta. Arthritis Rheum 2000, 43, 2189–2201. [Google Scholar] [CrossRef]
- Grigolo, B.; Roseti, L.; Neri, S.; Gobbi, P.; Jensen, P.; Major, E.O.; Facchini, A. Human articular chondrocytes immortalized by HPV-16 E6 and E7 genes: Maintenance of differentiated phenotype under defined culture conditions. Osteoarthr. Cartil. 2002, 10, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.H.; Lai, W.F.; Deng, W.P.; Yang, W.K.; Lo, W.C.; Wu, C.C.; Yang, D.M.; Lai, M.T.; Lin, C.T.; Lin, T.W.; et al. Tissue engineered cartilage using human articular chondrocytes immortalized by HPV-16 E6 and E7 genes. J. Biomed. Mater. Res. Part A 2006, 76, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, M.K.; Chockalingam, P.S.; Bhat, R.A.; Sheldon, R.; Keohan, C.; Blanchet, T.; Glasson, S.; Morris, E.A. Immortalized cell lines from mouse xiphisternum preserve chondrocyte phenotype. J. Cell Physiol. 2006, 209, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, M.K.; Chockalingam, P.S.; Bhat, R.A.; Sheldon, R.; Keohan, C.; Blanchet, T.; Glasson, S.; Morris, E.A. Immortalized mouse articular cartilage cell lines retain chondrocyte phenotype and respond to both anabolic factor BMP-2 and pro-inflammatory factor IL-1. J. Cell Physiol. 2008, 215, 68–76. [Google Scholar] [CrossRef]
- Hoffman, B.E.; Newman-Tarr, T.M.; Gibbard, A.; Wang, S.; Hanning, C.; Pratta, M.A.; Boyle, R.J.; Kumar, S.; Majumdar, M.K. Development and characterization of a human articular cartilage-derived chondrocyte cell line that retains chondrocyte phenotype. J. Cell Physiol. 2010, 222, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Hodax, J.K.; Quintos, J.B.; Gruppuso, P.A.; Chen, Q.; Desai, S.; Jayasuriya, C.T. Aggrecan is required for chondrocyte differentiation in ATDC5 chondroprogenitor cells. PLoS ONE 2019, 14, e0218399. [Google Scholar] [CrossRef]
- Cinque, L.; De Leonibus, C.; Iavazzo, M.; Krahmer, N.; Intartaglia, D.; Salierno, F.G.; De Cegli, R.; Di Malta, C.; Svelto, M.; Lanzara, C.; et al. MiT/TFE factors control ER-phagy via transcriptional regulation of FAM134B. EMBO J. 2020, 39, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Paganini, C.; Costantini, R.; Superti-Furga, A.; Rossi, A. Bone and connective tissue disorders caused by defects in glycosaminoglycan biosynthesis: A panoramic view. FEBS J. 2019, 286, 3008–3032. [Google Scholar] [CrossRef] [Green Version]
- Paganini, C.; Gramegna Tota, C.; Superti-Furga, A.; Rossi, A. Skeletal Dysplasias Caused by Sulfation Defects. Int. J. Mol. Sci. 2020, 21, 2710. [Google Scholar] [CrossRef] [Green Version]
- Atsumi, T.; Miwa, Y.; Kimata, K.; Ikawa, Y. A chondrogenic cell line derived from a differentiating culture of AT805 teratocarcinoma cells. Cell Differ. Dev. 1990, 30, 109–116. [Google Scholar] [CrossRef]
- Paganini, C.; Monti, L.; Costantini, R.; Besio, R.; Lecci, S.; Biggiogera, M.; Tian, K.; Schwartz, J.M.; Huber, C.; Cormier-Daire, V.; et al. Calcium activated nucleotidase 1 (CANT1) is critical for glycosaminoglycan biosynthesis in cartilage and endochondral ossification. Matrix Biol. 2019, 81, 70–90. [Google Scholar] [CrossRef]
- Huber, C.; Oules, B.; Bertoli, M.; Chami, M.; Fradin, M.; Alanay, Y.; Al-Gazali, L.I.; Ausems, M.G.; Bitoun, P.; Cavalcanti, D.P.; et al. Identification of CANT1 mutations in Desbuquois dysplasia. Am. J. Hum. Genet. 2009, 85, 706–710. [Google Scholar] [CrossRef] [Green Version]
- Nizon, M.; Huber, C.; De Leonardis, F.; Merrina, R.; Forlino, A.; Fradin, M.; Tuysuz, B.; Abu-Libdeh, B.Y.; Alanay, Y.; Albrecht, B.; et al. Further delineation of CANT1 phenotypic spectrum and demonstration of its role in proteoglycan synthesis. Hum. Mutat. 2012, 33, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Leijten, J.C.; Wu, L.; Kip, M.; van Blitterswijk, C.A.; Post, J.N.; Karperien, M. Gene expression profiling of dedifferentiated human articular chondrocytes in monolayer culture. Osteoarthr. Cartil. 2013, 21, 599–603. [Google Scholar] [CrossRef] [Green Version]
- Mullan, L.A.; Mularczyk, E.J.; Kung, L.H.; Forouhan, M.; Wragg, J.M.; Goodacre, R.; Bateman, J.F.; Swanton, E.; Briggs, M.D.; Boot-Handford, R.P. Increased intracellular proteolysis reduces disease severity in an ER stress-associated dwarfism. J. Clin. Investig. 2017, 127, 3861–3865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steimberg, N.; Viengchareun, S.; Biehlmann, F.; Guénal, I.; Mignotte, B.; Adolphe, M.; Thenet, S. SV40 large T antigen expression driven by col2a1 regulatory sequences immortalizes articular chondrocytes but does not allow stabilization of type II collagen expression. Exp. Cell Res. 1999, 249, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, M.; Pan, H.O.; Kinoshita, A.; Tajima, K.; Takano, Y. Establishment from a human chondrosarcoma of a new immortal cell line with high tumorigenicity in vivo, which is able to form proteoglycan-rich cartilage-like nodules and to respond to insulin in vitro. Int. J. Cancer 1991, 48, 717–725. [Google Scholar] [CrossRef]
- Böhme, K.; Conscience-Egli, M.; Tschan, T.; Winterhalter, K.H.; Bruckner, P. Induction of proliferation or hypertrophy of chondrocytes in serum-free culture: The role of insulin-like growth factor-I, insulin, or thyroxine. J. Cell Biol. 1992, 116, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepko, C.L.; Roberts, B.E.; Mulligan, R.C. Construction and applications of a highly transmissible murine retrovirus shuttle vector. Cell 1984, 37, 1053–1062. [Google Scholar] [CrossRef]
- Yagüe, E.; Arance, A.; Kubitza, L.; O’Hare, M.; Jat, P.; Ogilvie, C.M.; Hart, I.R.; Higgins, C.F.; Raguz, S. Ability to acquire drug resistance arises early during the tumorigenesis process. Cancer Res. 2007, 67, 1130–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganini, C.; Costantini, R.; Rossi, A. Analysis of Proteoglycan Synthesis and Secretion in Cell Culture Systems. Methods Mol. Biol. 2019, 1952, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Monti, L.; Paganini, C.; Lecci, S.; De Leonardis, F.; Hay, E.; Cohen-Solal, M.; Villani, S.; Superti-Furga, A.; Tenni, R.; Forlino, A.; et al. N-acetylcysteine treatment ameliorates the skeletal phenotype of a mouse model of diastrophic dysplasia. Hum. Mol. Genet. 2015, 24, 5570–5580. [Google Scholar] [CrossRef] [Green Version]
- Paganini, C.; Gramegna Tota, C.; Monti, L.; Monti, I.; Maurizi, A.; Capulli, M.; Bourmaud, M.; Teti, A.; Cohen-Solal, M.; Villani, S.; et al. Improvement of the skeletal phenotype in a mouse model of diastrophic dysplasia after postnatal treatment with N-acetylcysteine. Biochem. Pharmacol. 2021, 185, 114452. [Google Scholar] [CrossRef] [PubMed]
- Hedman, K.; Kurkinen, M.; Alitalo, K.; Vaheri, A.; Johansson, S.; Höök, M. Isolation of the pericellular matrix of human fibroblast cultures. J. Cell Biol. 1979, 81, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gramegna Tota, C.; Valenti, B.; Forlino, A.; Rossi, A.; Paganini, C. Phenotypic Characterization of Immortalized Chondrocytes from a Desbuquois Dysplasia Type 1 Mouse Model: A Tool for Studying Defects in Glycosaminoglycan Biosynthesis. Int. J. Mol. Sci. 2021, 22, 9304. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179304
Gramegna Tota C, Valenti B, Forlino A, Rossi A, Paganini C. Phenotypic Characterization of Immortalized Chondrocytes from a Desbuquois Dysplasia Type 1 Mouse Model: A Tool for Studying Defects in Glycosaminoglycan Biosynthesis. International Journal of Molecular Sciences. 2021; 22(17):9304. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179304
Chicago/Turabian StyleGramegna Tota, Chiara, Beatrice Valenti, Antonella Forlino, Antonio Rossi, and Chiara Paganini. 2021. "Phenotypic Characterization of Immortalized Chondrocytes from a Desbuquois Dysplasia Type 1 Mouse Model: A Tool for Studying Defects in Glycosaminoglycan Biosynthesis" International Journal of Molecular Sciences 22, no. 17: 9304. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179304