
Properties of Transport Mediated by the Human Organic Cation Transporter 2 Studied in a Polarized Three-Dimensional Epithelial Cell Culture Model

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
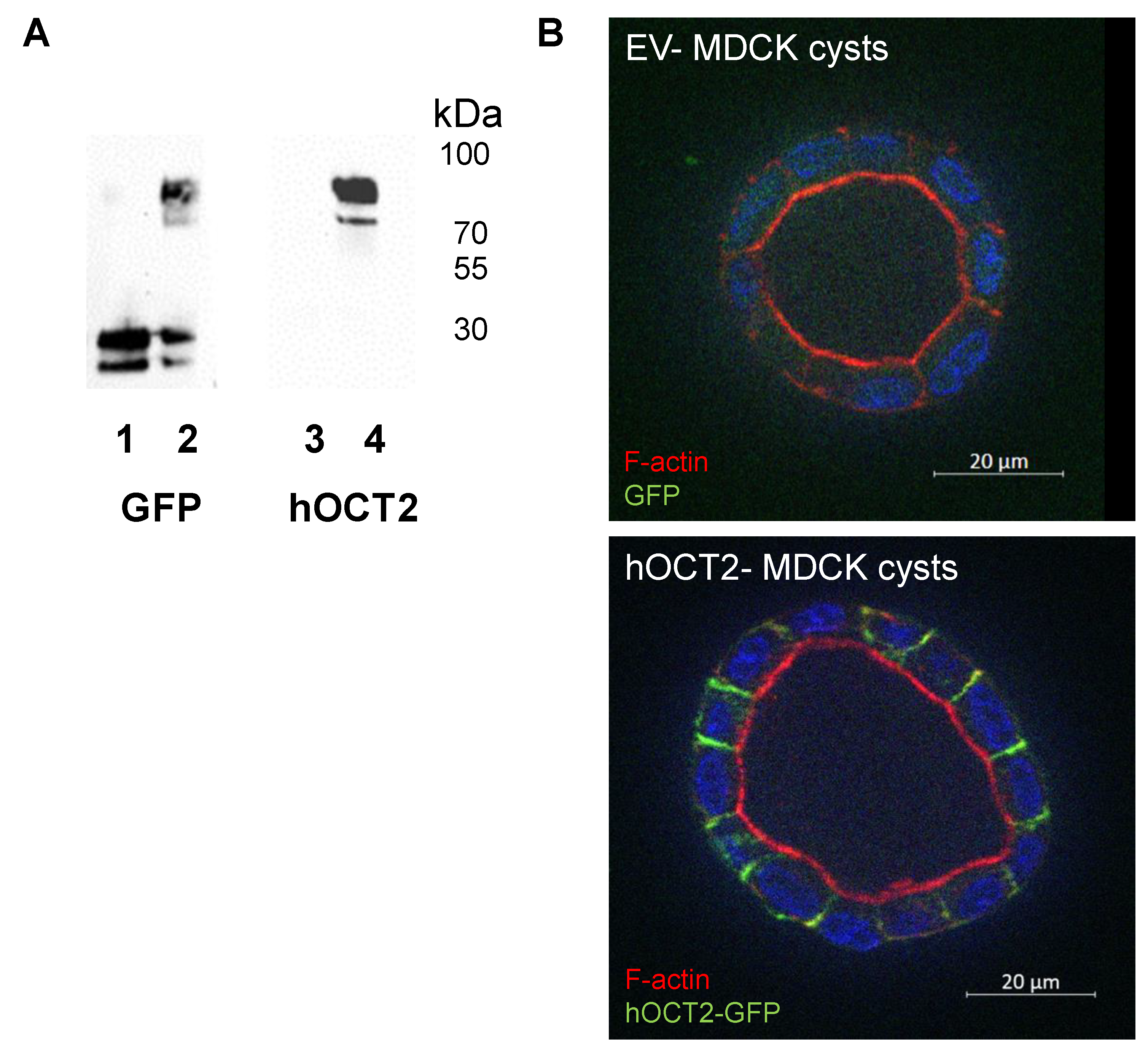
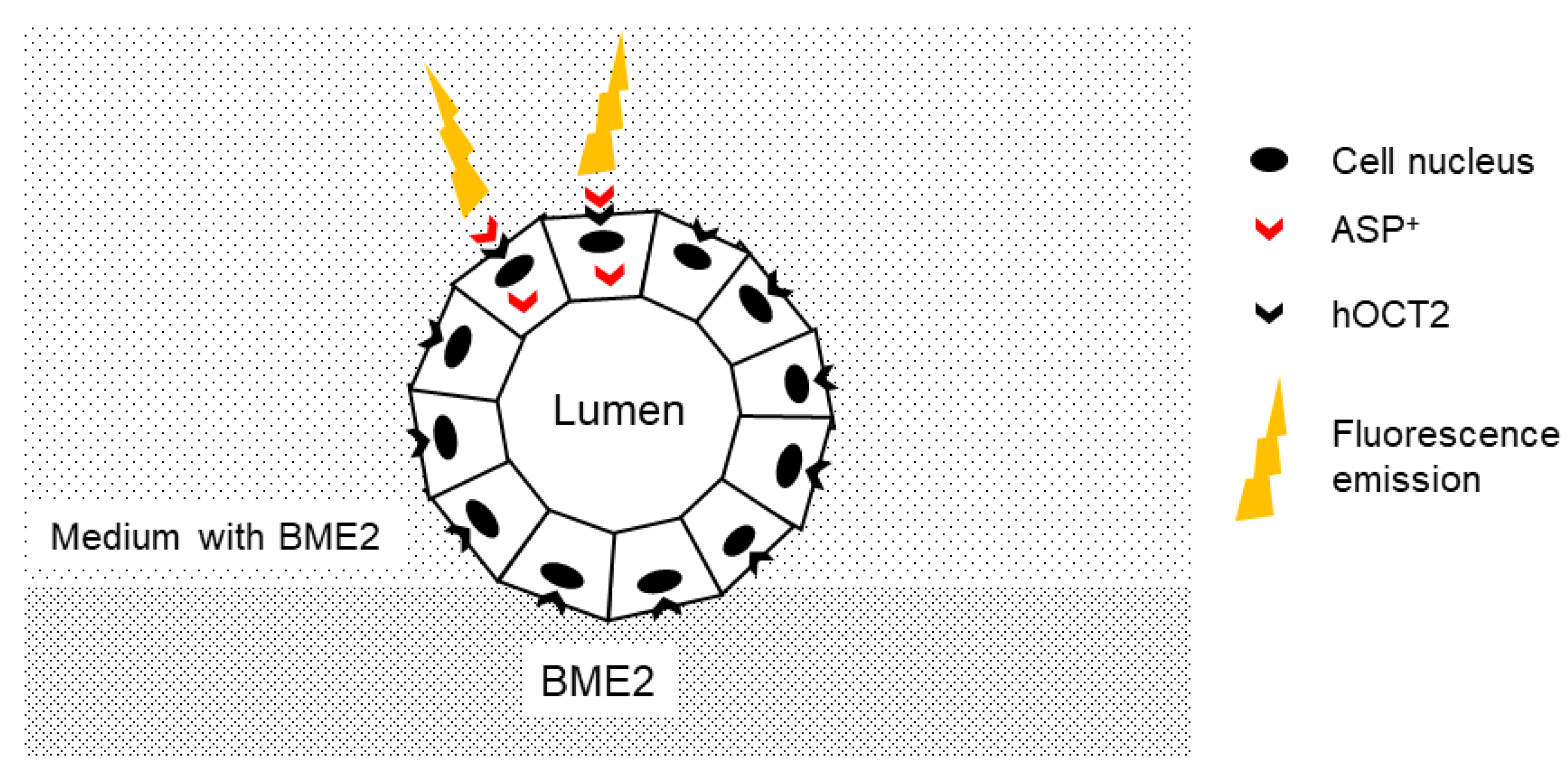
2.1. Characterization of MDCK Cysts
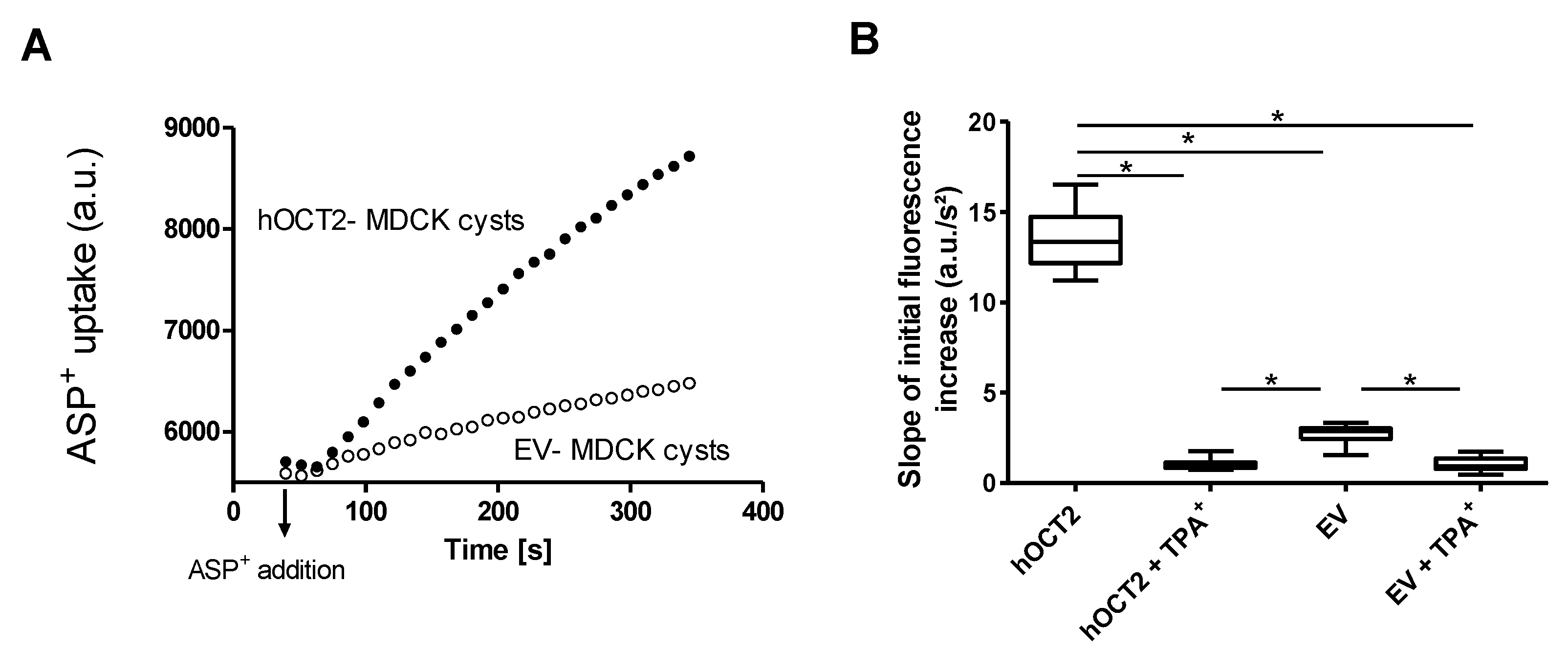
2.2. Uptake of the Fluorescent Organic Cation 4-(4-(Dimethylamino)styryl)-N-methylpyridinium (ASP+) in MDCK Cysts
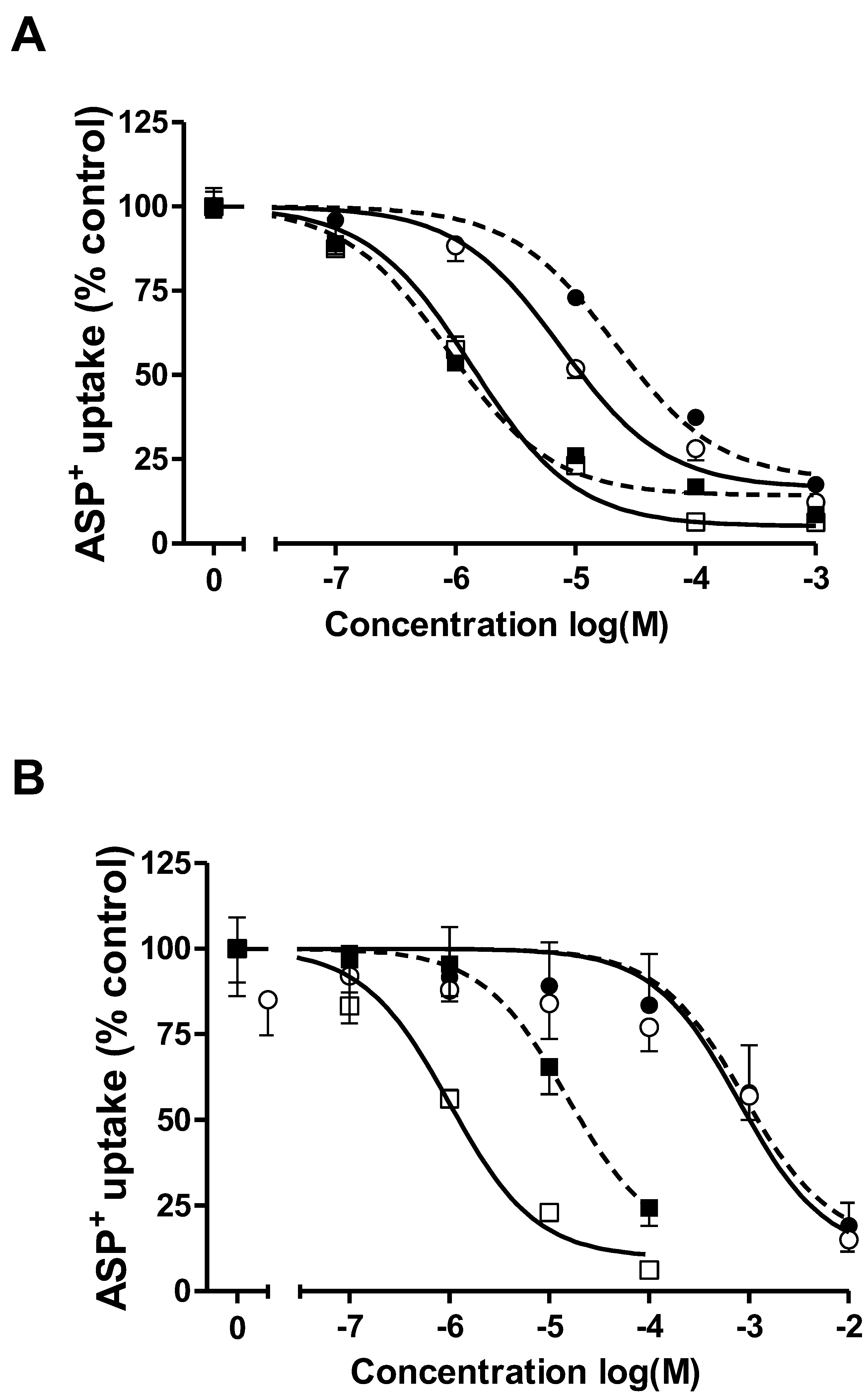
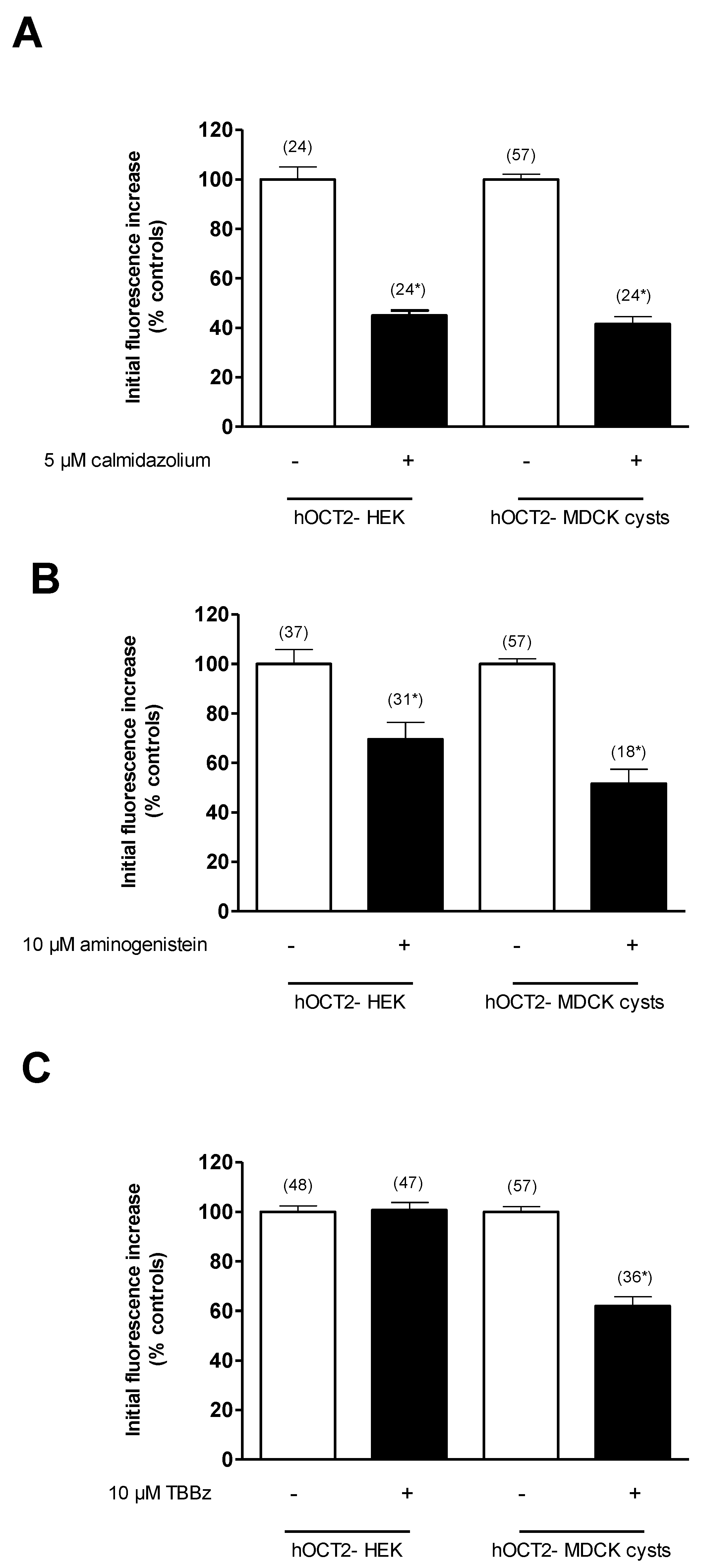
2.3. Comparison of ASP+ Transport Mediated by hOCT2 Stably Expressed in HEK293 (hOCT2-HEK) Cells or in MDCK Cysts (hOCT2- MDCK)
3. Discussion
4. Materials and Methods
4.1. Cloning of hOCT2-GFP into the Viral Transduction Vector
4.2. Cell Culture
4.3. Generation of MDCK Cell Lines Expressing hOCT2-GFP or GFP Alone
4.4. Western Blot Analysis
4.5. Immunofluorescence Analysis
4.6. Measurement of OCT Function
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Motohashi, H.; Sakurai, Y.; Saito, H.; Masuda, S.; Urakami, Y.; Goto, M.; Fukatsu, A.; Ogawa, O.; Inui, K.-I. Gene expression levels and immunolocalization of organic ion transporters in the human kidney. J. Am. Soc. Nephrol. 2002, 13, 866–874. [Google Scholar] [CrossRef]
- Oswald, S.; Muller, J.; Neugebauer, U.; Schroter, R.; Herrmann, E.; Pavenstadt, H.; Ciarimboli, G. Protein abundance of clinically relevant drug transporters in the human kidneys. Int. J. Mol. Sci. 2019, 20, 5303. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, H.; Nakao, Y.; Masuda, S.; Katsura, T.; Kamba, T.; Ogawa, O.; Inui, K.-I. Precise comparison of protein localization among OCT, OAT, and MATE in human kidney. J. Pharm. Sci. 2013, 102, 3302–3308. [Google Scholar] [CrossRef]
- Koepsell, H. Organic cation transporters in health and disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.H.; Dantzler, W.H. Molecular and cellular physiology of renal organic cation and anion transport. Physiol. Rev. 2004, 84, 987–1049. [Google Scholar] [CrossRef]
- Omote, H.; Hiasa, M.; Matsumoto, T.; Otsuka, M.; Moriyama, Y. The MATE proteins as fundamental transporters of metabolic and xenobiotic organic cations. Trends Pharmacol. Sci. 2006, 27, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Kestenbaum, B. Proximal tubular secretory clearance: A neglected partner of kidney function. Clin. J. Am. Soc. Nephrol. 2018, 13, 1291–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gründemann, D.; Koster, S.; Kiefer, N.; Breidert, T.; Engelhardt, M.; Spitzenberger, F.; Obermuller, N.; Schomig, E. Transport of monoamine transmitters by the organic cation transporter type 2, OCT2. J. Biol. Chem. 1998, 273, 30915–30920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baganz, N.L.; Horton, R.E.; Calderon, A.S.; Owens, W.A.; Munn, J.L.; Watts, L.T.; Koldzic-Zivanovic, N.; Jeske, N.A.; Koek, W.; Toney, G.M.; et al. Organic cation transporter 3: Keeping the brake on extracellular serotonin in serotonin-transporter-deficient mice. Proc. Natl. Acad. Sci. USA 2008, 105, 18976–18981. [Google Scholar] [CrossRef] [Green Version]
- Ciarimboli, G.; Lancaster, C.S.; Schlatter, E.; Franke, R.M.; Sprowl, J.A.; Pavenstädt, H.; Massmann, V.; Guckel, D.; Mathijssen, R.H.J.; Yang, W.; et al. Proximal tubular secretion of creatinine by organic cation transporter OCT2 in cancer patients. Clin. Cancer Res. 2012, 18, 1101–1108. [Google Scholar] [CrossRef] [Green Version]
- Urakami, Y.; Kimura, N.; Okuda, M.; Inui, K.-I. Creatinine transport by basolateral organic cation transporter hOCT2 in the human kidney. Pharm. Res. 2004, 21, 976–981. [Google Scholar] [CrossRef]
- Sala-Rabanal, M.; Li, D.C.; Dake, G.R.; Kurata, H.T.; Inyushin, M.; Skatchkov, S.N.; Nichols, C.G. Polyamine transport by the polyspecific organic cation transporters OCT1, OCT2, and OCT3. Mol. Pharm. 2013, 10, 1450–1458. [Google Scholar] [CrossRef] [Green Version]
- Song, I.S.; Shin, H.J.; Shin, J.G. Genetic variants of organic cation transporter 2 (OCT2) significantly reduce metformin uptake in oocytes. Xenobiotica 2008, 38, 1252–1262. [Google Scholar] [CrossRef]
- Kimura, N.; Okuda, M.; Inui, K.-I. Metformin transport by renal basolateral organic cation transporter hOCT2. Pharm. Res. 2005, 22, 255–259. [Google Scholar] [CrossRef]
- Hasannejad, H.; Takeda, M.; Narikawa, S.; Huang, X.L.; Enomoto, A.; Taki, K.; Niwa, T.; Jung, S.H.; Onozato, M.L.; Tojo, A.; et al. Human organic cation transporter 3 mediates the transport of antiarrhythmic drugs. Eur. J. Pharmacol. 2004, 499, 45–51. [Google Scholar] [CrossRef]
- Dudley, A.J.; Bleasby, K.; Brown, C.D. The organic cation transporter OCT2 mediates the uptake of beta-adrenoceptor antagonists across the apical membrane of renal LLC-PK(1) cell monolayers. Br. J. Pharmacol. 2000, 131, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarimboli, G.; Schroter, R.; Neugebauer, U.; Vollenbroker, B.; Gabriels, G.; Brzica, H.; Sabolic, I.; Pietig, G.; Pavemstadt, H.; Schlatter, E.; et al. Kidney transplantation down-regulates expression of organic cation transporters, which translocate beta-blockers and fluoroquinolones. Mol. Pharm. 2013, 10, 2370–2380. [Google Scholar] [CrossRef]
- Bachmakov, I.; Glaeser, H.; Endress, B.; Morl, F.; Konig, J.; Fromm, M.F. Interaction of beta-blockers with the renal uptake transporter OCT2. Diabetes Obes. Metab. 2009, 11, 1080–1083. [Google Scholar] [CrossRef] [PubMed]
- Urakami, Y.; Akazawa, M.; Saito, H.; Okuda, M.; Inui, K.-I. cDNA cloning, functional characterization, and tissue distribution of an alternatively spliced variant of organic cation transporter hOCT2 predominantly expressed in the human kidney. J. Am. Soc. Nephrol. 2002, 13, 1703–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenisch, B.; Drescher, E.; Thiemer, L.; Xin, H.; Giros, B.; Gautron, S.; Bonisch, H. Interaction of antidepressant and antipsychotic drugs with the human organic cation transporters hOCT1, hOCT2 and hOCT3. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 1017–1023. [Google Scholar] [CrossRef]
- Gorboulev, V.; Ulzheimer, J.C.; Akhoundova, A.; Ulzheimer-Teuber, I.; Karbach, U.; Quester, S.; Baumann, C.; Lang, F.; Busch, A.E.; Koepsell, H. Cloning and characterization of two human polyspecific organic cation transporters. DNA Cell Biol. 1997, 16, 871–881. [Google Scholar] [CrossRef]
- Busch, A.E.; Quester, S.; Ulzheimer, J.C.; Waldegger, S.; Gorboulev, V.; Arndt, P.; Lang, F.; Koepsell, H. Electrogenic properties and substrate specificity of the polyspecific rat cation transporter rOCT1. J. Biol. Chem. 1996, 271, 32599–32604. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Dresser, M.J.; Gray, A.T.; Yost, S.C.; Terashita, S.; Giacomini, K.M. Cloning and functional expression of a human liver organic cation transporter. Mol. Pharmacol. 1997, 51, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Elia, N.; Lippincott-Schwartz, J. Culturing MDCK cells in three dimensions for analyzing intracellular dynamics. Curr. Protoc. Cell Biol. 2009, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Yoshihama, Y.; Sasaki, K.; Horikoshi, Y.; Suzuki, A.; Ohtsuka, T.; Hakuno, F.; Takahashi, S.-I.; Ohno, S.; Chida, K. KIBRA suppresses apical exocytosis through inhibition of aPKC kinase activity in epithelial cells. Curr. Biol. 2011, 21, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, T.; Riethmuller, C.; Gekle, M.; Schwerdt, G.; Oberleithner, H. Nephrotoxicity of platinum complexes is related to basolateral organic cation transport. Kidney Int. 2004, 66, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gorset, W.; Dresser, M.J.; Giacomini, K.M. The interaction of n-tetraalkylammonium compounds with a human organic cation transporter, hOCT1. J. Pharmacol. Exp. Ther. 1999, 288, 1192–1198. [Google Scholar]
- Dresser, M.J.; Xiao, G.; Leabman, M.K.; Gray, A.T.; Giacomini, K.M. Interactions of n-tetraalkylammonium compounds and biguanides with a human renal organic cation transporter (hOCT2). Pharm. Res. 2002, 19, 1244–1247. [Google Scholar] [CrossRef]
- Kimura, N.; Masuda, S.; Katsura, T.; Inui, K.-I. Transport of guanidine compounds by human organic cation transporters, hOCT1 and hOCT2. Biochem. Pharmacol. 2009, 77, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wang, L.; Grundemann, D.; Sweet, D.H. Interaction of Ethambutol with human organic cation transporters of the SLC22 family indicates potential for drug-drug interactions during antituberculosis therapy. Antimicrob. Agents Chemother. 2013, 57, 5053–5059. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Sun, X.; Chen, J.; Lu, Y.; Zhang, Y.; Wang, C.; Li, J.; Zhang, Q.; Zhao, D.; Chen, X. Investigation of the role of organic cation transporter 2 (OCT2) in the renal transport of guanfacine, a selective alpha-adrenoreceptor agonist. Xenobiotica 2014, 45, 88–94. [Google Scholar] [CrossRef]
- Pietig, G.; Mehrens, T.; Hirsch, J.R.; Cetinkaya, I.; Piechota, H.; Schlatter, E. Properties and regulation of organic cation transport in freshly isolated human proximal tubules. J. Biol. Chem. 2001, 276, 33741–33746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukes, J.D.; Whitley, P.; Chalmers, A.D. The MDCK variety pack: Choosing the right strain. BMC Cell Biol. 2011, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, L.E.; Zegers, M.M.; Mostov, K.E. Opinion: Building epithelial architecture: Insights from three-dimensional culture models. Nat. Rev. Mol. Cell Biol. 2002, 3, 531–537. [Google Scholar] [CrossRef]
- Sato, T.; Masuda, S.; Yonezawa, A.; Tanihara, Y.; Katsura, T.; Inui, K.-I. Transcellular transport of organic cations in double-transfected MDCK cells expressing human organic cation transporters hOCT1/hMATE1 and hOCT2/hMATE1. Biochem. Pharmacol. 2008, 76, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Konig, J.; Zolk, O.; Singer, K.; Hoffmann, C.; Fromm, M.F. Double-transfected MDCK cells expressing human OCT1/MATE1 or OCT2/MATE1: Determinants of uptake and transcellular translocation of organic cations. Br. J. Pharmacol. 2011, 163, 546–555. [Google Scholar] [CrossRef] [Green Version]
- Kantauskaite, M.; Hucke, A.; Reike, M.; Ahmed, E.S.; Xiao, C.; Barz, V.; Ciarimboli, G. Rapid regulation of human multidrug and extrusion transporters hMATE1 and hMATE2K. Int. J. Mol. Sci. 2020, 21, 5157. [Google Scholar] [CrossRef]
- Ciarimboli, G.; Koepsell, H.; Iordanova, M.; Gorboulev, V.; Dürner, B.; Lang, D.; Edemir, B.; Schroter, R.; Van le Truc, E. Individual PKC-phosphorylation sites in organic cation transporter 1 determine substrate selectivity and transport regulation. J. Am. Soc. Nephrol. 2005, 16, 1562–1570. [Google Scholar] [CrossRef] [Green Version]
- Popp, C.; Gorboulev, V.; Muller, T.D.; Gorbunov, D.; Shatskaya, N.; Koepsell, H. Amino acids critical for substrate affinity of rat organic cation transporter 1 line the substrate binding region in a model derived from the tertiary structure of lactose permease. Mol. Pharmacol. 2005, 67, 1600–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koepsell, H. Multiple binding sites in organic cation transporters require sophisticated procedures to identify interactions of novel drugs. Biol. Chem. 2019, 400, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S.H. Molecular and cellular physiology of organic cation transporter 2. Am. J. Physiol. Renal Physiol. 2019, 317, F1669–F1679. [Google Scholar] [CrossRef] [PubMed]
- Bijlmakers, M.J.; Isobe-Nakamura, M.; Ruddock, L.J.; Marsh, M. Intrinsic signals in the unique domain target p56(lck) to the plasma membrane independently of CD4. J. Cell Biol. 1997, 137, 1029–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.M.; Roderick, H.L.; McDonald, F.; Bootman, M.D. Interaction between store-operated and arachidonate-activated calcium entry. Cell Calcium 2007, 41, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Galan, C.; Dionisio, N.; Smani, T.; Salido, G.M.; Rosado, J.A. The cytoskeleton plays a modulatory role in the association between STIM1 and the Ca2+ channel subunits Orai1 and TRPC1. Biochem. Pharmacol. 2011, 82, 400–410. [Google Scholar] [CrossRef] [Green Version]
- Jan, C.R.; Tseng, C.J. Calmidazolium-induced rises in cytosolic calcium concentrations in Madin Darby canine kidney cells. Toxicol. Appl. Pharmacol. 2000, 162, 142–150. [Google Scholar] [CrossRef]
- Deshiere, A.; Theis-Febvre, N.; Martel, V.; Cochet, C.; Filhol, O. Protein kinase CK2 and cell polarity. Mol. Cell Biochem. 2008, 316, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.K.; Reichold, M.; Edemir, B.; Ciarimboli, G.; Warth, R.; Koepsell, H.; Thevenod, F. Organic cation transporters OCT1, 2, and 3 mediate high-affinity transport of the mutagenic vital dye ethidium in the kidney proximal tubule. Am. J. Physiol. Renal Physiol. 2009, 296, F1504–F1513. [Google Scholar] [CrossRef] [Green Version]
- Brast, S.; Grabner, A.; Sucic, S.; Sitte, H.H.; Hermann, E.; Pavenstadt, H.; Schlatter, E.; Ciarimboli, G. The cysteines of the extracellular loop are crucial for trafficking of human organic cation transporter 2 to the plasma membrane and are involved in oligomerization. FASEB J. 2012, 26, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Schulze, U.; Brast, S.; Grabner, A.; Albiker, C.; Snieder, B.; Holle, S.; Schlatter, E.; Schroter, R.; Pavenstadt, H.; Herrmann, E.; et al. Tetraspanin CD63 controls basolateral sorting of organic cation transporter 2 in renal proximal tubules. FASEB J. 2017, 31, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- Schulze, U.; Vollenbroker, B.; Braun, D.A.; Le, T.V.; Granado, D.; Kremerskothen, J.; Franzel, B.; Klosowski, R.; Barth, J.; Fufezan, C.; et al. The Vac14-interaction network is linked to regulators of the endolysosomal and autophagic pathway. Mol. Cell. Proteom. 2014, 13, 1397–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wennmann, D.O.; Vollenbroker, B.; Eckart, A.K.; Bonse, J.; Erdmann, F.; Wolters, D.A.; Schenk, L.K.; Schulze, U.; Kremerskothen, J.; Weide, T.; et al. The Hippo pathway is controlled by Angiotensin II signaling and its reactivation induces apoptosis in podocytes. Cell Death Dis. 2014, 5, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djuric, I.; Siebrasse, J.P.; Schulze, U.; Granado, D.; Schluter, M.A.; Kubitscheck, U.; Pavenstadt, H.; Weide, T. The C-terminal domain controls the mobility of Crumbs 3 isoforms. Biochim. Biophys. Acta 2016, 1863, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Biermann, J.; Lang, D.; Gorboulev, V.; Koepsell, H.; Sindic, A.; Schröter, R.; Zvirbliene, A.; Pavenstadt, H.; Schlatter, E.; Ciarimboli, G. Characterization of regulatory mechanisms and states of human organic cation transporter 2. Am. J. Physiol. Cell. Physiol. 2006, 290, C1521–C1531. [Google Scholar] [CrossRef] [PubMed]
- Ciarimboli, G.; Schlatter, E. Organic cation transport measurements using fluorescence techniques. In Neurotransmitter Transporters: Investigative Methods; Bönisch, H., Sitte, H.H., Eds.; Springer Science and Business Media: New York, NY, USA, 2016; pp. 173–187. [Google Scholar]
- Wilde, S.; Schlatter, E.; Koepsell, H.; Edemir, B.; Reuter, S.; Pavenstädt, H.; Neugebauer, U.; Schroter, R.; Brast, S.; Ciarimboli, G. Calmodulin-associated post-translational regulation of rat organic cation transporter 2 in the kidney is gender dependent. Cell Mol. Life Sci. 2009, 66, 1729–1740. [Google Scholar] [CrossRef]
- Frenzel, D.; Koppen, C.; Bauer, O.B.; Karst, U.; Schroter, R.; Tzvetkov, M.V.; Ciarimboli, G. Effects of single nucleotide polymorphism Ala270Ser (rs316019) on the function and regulation of hOCT2. Biomolecules 2019, 9, 578. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substances | Km ± SEM or IC50 (logIC50 ± SEM) in µM and Number of Replicates (n) Measured in at Least 3 Independent Experiments | |
|---|---|---|
| hOCT2-HEK | hOCT2-MDCK Cysts | |
| ASP+ | 3.1 ± 0.2 n = 14–15 | 3.6 ± 1.2 n = 9 |
| TEA+ | 7.6 (−5.12 ± 0.08) n = 24 | 21.3 * (−4.67 ± 0.11) n = 6–12 |
| TPA+ | 1.4 (−5.85 ± 0.11) n = 12–30 | 0.9 (−6.04 ± 0.05) n = 18–21 |
| Metformin | 804 (−3.10 ± 0.11) n = 8–48 | 884 (−3.05 ± 0.09) n = 11–24 |
| Baricitinib | 1.1 (−6.00 ± 0.06) n = 12–24 | 15.4 * (−4.81 ± 0.06) n = 18–24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koepp, T.N.; Tokaj, A.; Nedvetsky, P.I.; Conchon Costa, A.C.; Snieder, B.; Schröter, R.; Ciarimboli, G. Properties of Transport Mediated by the Human Organic Cation Transporter 2 Studied in a Polarized Three-Dimensional Epithelial Cell Culture Model. Int. J. Mol. Sci. 2021, 22, 9658. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179658
Koepp TN, Tokaj A, Nedvetsky PI, Conchon Costa AC, Snieder B, Schröter R, Ciarimboli G. Properties of Transport Mediated by the Human Organic Cation Transporter 2 Studied in a Polarized Three-Dimensional Epithelial Cell Culture Model. International Journal of Molecular Sciences. 2021; 22(17):9658. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179658
Chicago/Turabian StyleKoepp, Tim N., Alexander Tokaj, Pavel I. Nedvetsky, Ana Carolina Conchon Costa, Beatrice Snieder, Rita Schröter, and Giuliano Ciarimboli. 2021. "Properties of Transport Mediated by the Human Organic Cation Transporter 2 Studied in a Polarized Three-Dimensional Epithelial Cell Culture Model" International Journal of Molecular Sciences 22, no. 17: 9658. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179658