
Regulation of O-Linked N-Acetyl Glucosamine Transferase (OGT) through E6 Stimulation of the Ubiquitin Ligase Activity of E6AP

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
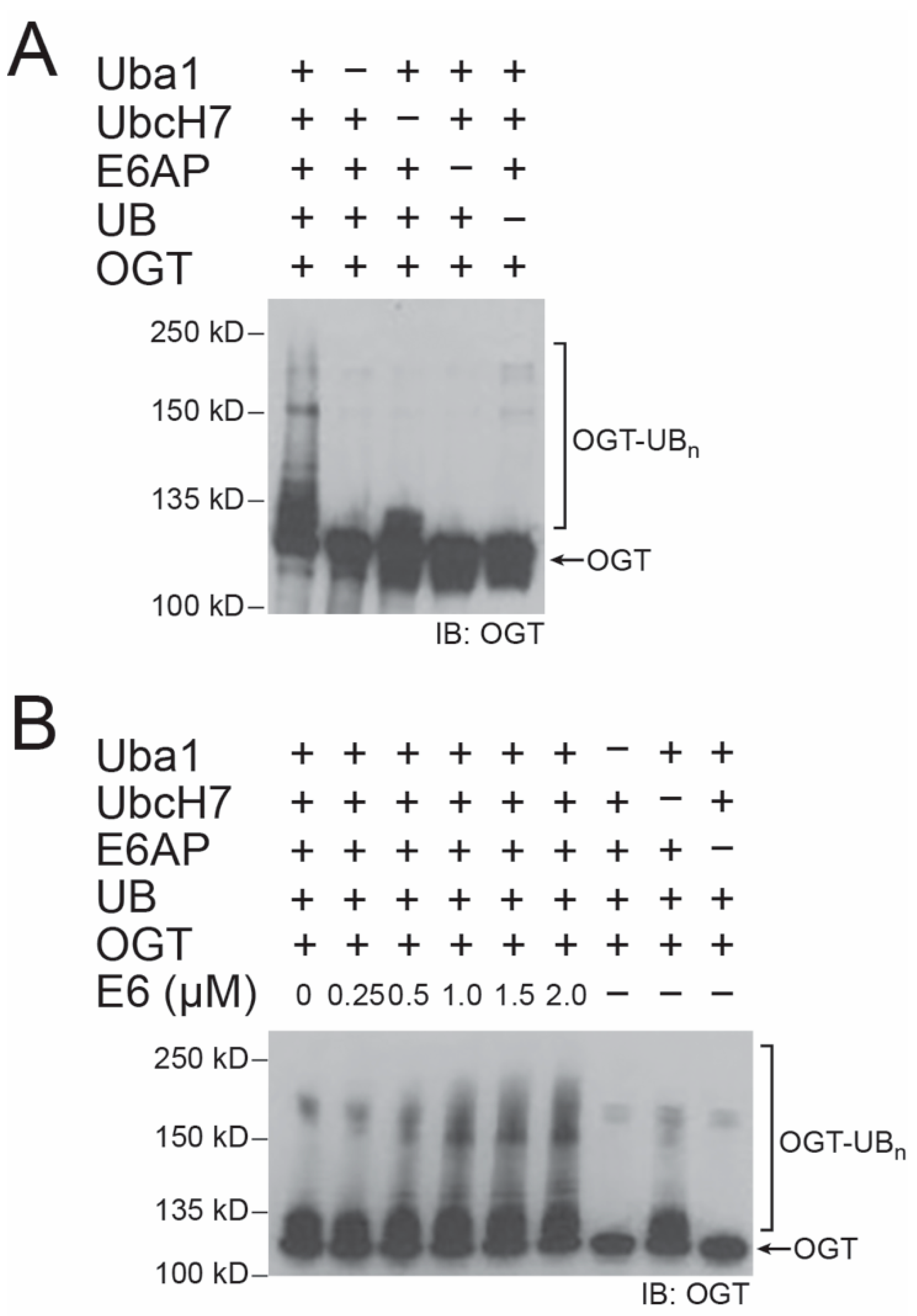
2.1. E6 Stimulation of E6AP in Catalyzing OGT Ubiquitination In Vitro
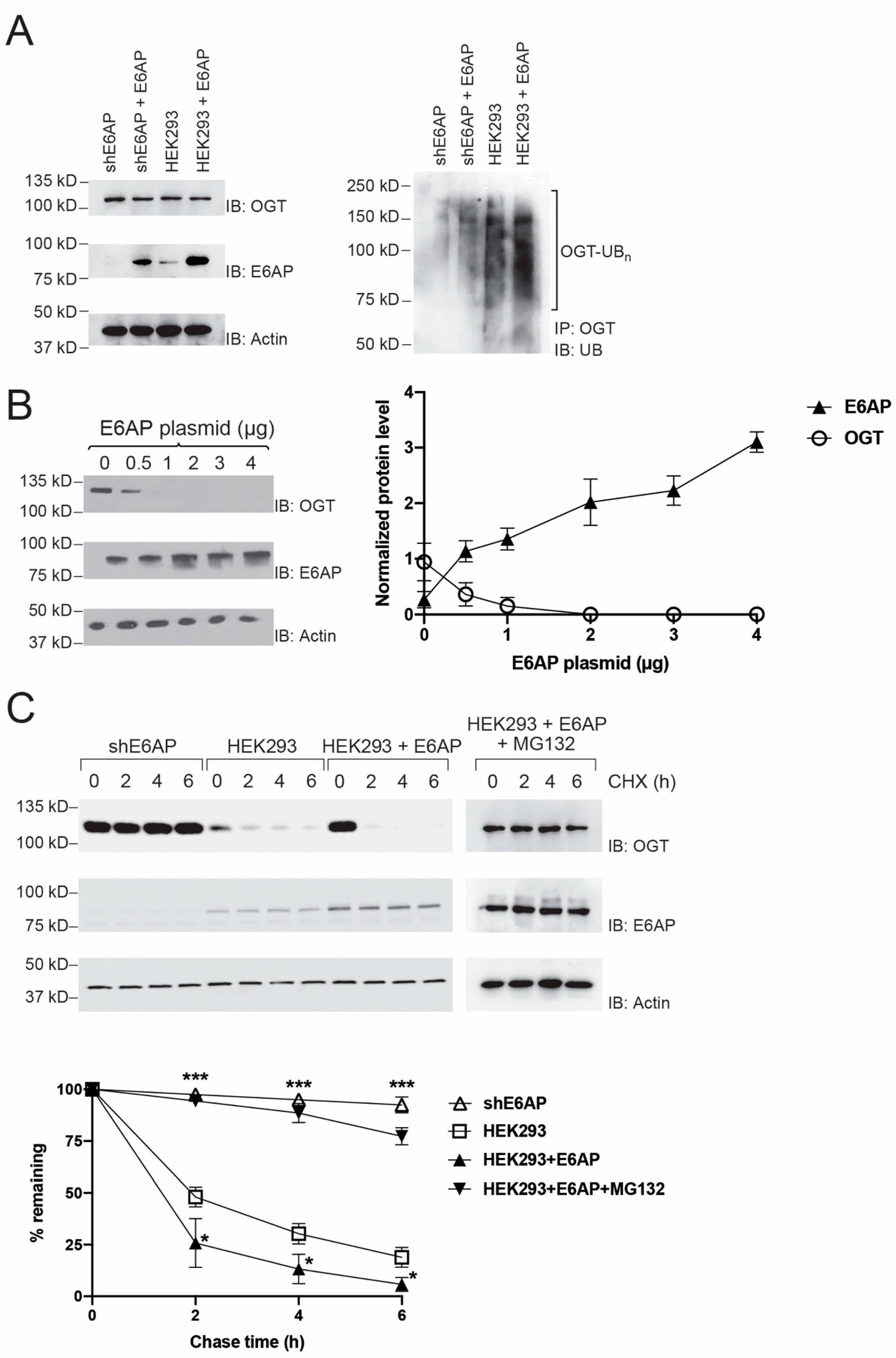
2.2. Regulation of OGT Stability in HEK293 Cells by E6AP
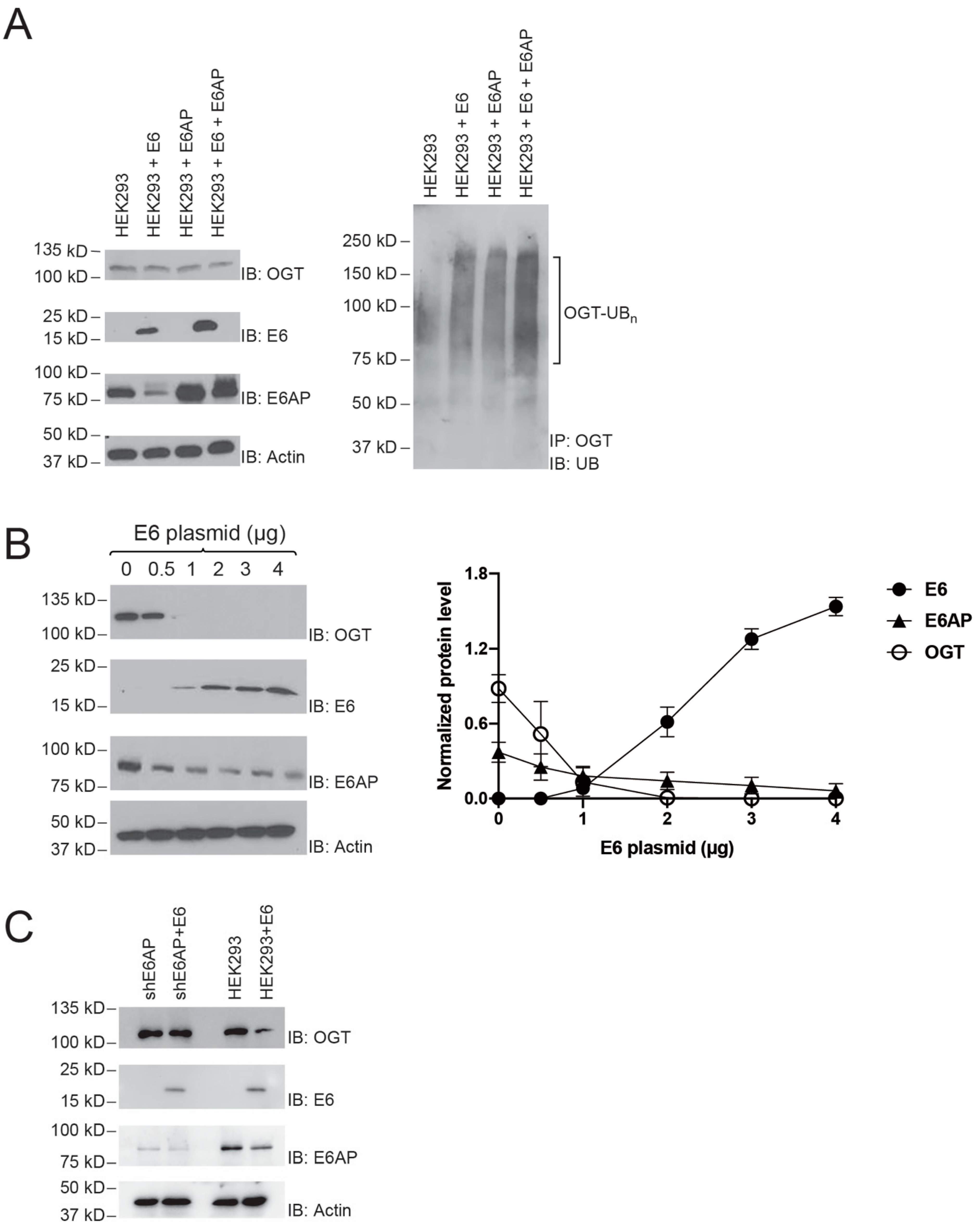
2.3. E6 stimulation of OGT Ubiquitination by E6AP
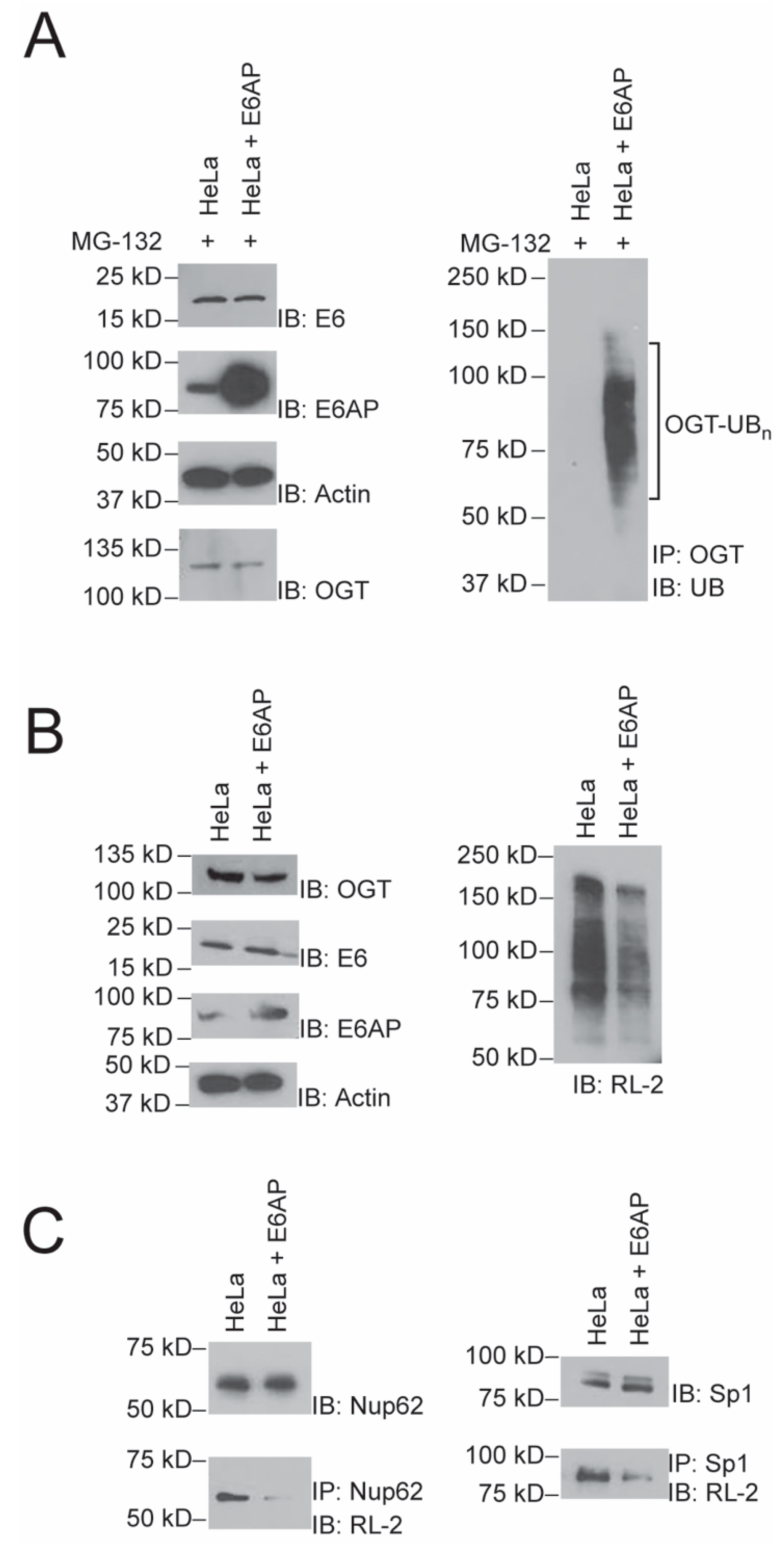
2.4. Effect of E6AP Activity on Protein O-GlcNAylation in HeLa Cells
3. Discussion
4. Materials and Methods
4.1. Plasmids
4.2. Antibodies
4.3. Expression and Purification of Recombinant Proteins
4.4. In Vitro Ubiquitination of OGT
4.5. Identification of the Ubiquitination Sites of OGT
4.6. Cell Culture and Transfection
4.7. Coimmunoprecipitation to Confirm OGT Ubiquitination in HEK 293 Cells
4.8. OGT Degradation Assays
4.9. Coimmunoprecipitation to Confirm E6AP Regulation of Protein O-GlcNAcylation
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CHX | cycloheximide |
| HPV | human papillomavirus |
| OGT | O-linked N-acetyl glucosamine transferase |
| O-GlcNAcylation | O-GlcNAc modification |
| OUT | orthogonal ubiquitin transfer |
| RL-2 | anti-O-GlcNAc antibody |
| UB | ubiquitin |
| UPS | ubiquitin–proteasome system |
References
- Hart, G.W.; Slawson, C.; Ramirez-Correa, G.; Lagerlof, O. Cross talk between O-GlcNAcylation and phosphorylation: Roles in signaling, transcription, and chronic disease. Ann. Rev. Biochem. 2011, 80, 825–858. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Qian, K. Protein O-GlcNAcylation: Emerging mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2017, 18, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Hanover, J.A.; Krause, M.W.; Love, D.C. Bittersweet memories: Linking metabolism to epigenetics through O-GlcNAcylation. Nat. Rev. Mol. Cell Biol. 2012, 13, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.B.; Nie, Y.; Yang, X. Regulation of protein degradation by O-GlcNAcylation: Crosstalk with ubiquitination. Mol. Cell Proteom. 2013, 12, 3489–3497. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kim, H.S.; Kim, N.H.; Ji, S.; Cha, S.Y.; Kang, J.G.; Ota, I.; Shimada, K.; Konishi, N.; Nam, H.W.; et al. Snail1 is stabilized by O-GlcNAc modification in hyperglycaemic condition. Embo J. 2010, 29, 3787–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.H.; Kim, J.E.; Nam, H.W.; Ju, J.W.; Kim, H.S.; Kim, Y.S.; Cho, J.W. Modification of p53 with O-linked N-acetylglucosamine regulates p53 activity and stability. Nat. Cell Biol. 2006, 8, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Zaro, B.W.; Yang, Y.Y.; Hang, H.C.; Pratt, M.R. Chemical reporters for fluorescent detection and identification of O-GlcNAc-modified proteins reveal glycosylation of the ubiquitin ligase NEDD4-1. Proc. Natl. Acad. Sci. USA 2011, 108, 8146–8151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, K.; Bai, B.; Ta, Y.; Zhang, T.; Xiao, Z.; Wang, P.G.; Zhang, L. O-GlcNAc regulates NEDD4-1 stability via caspase-mediated pathway. Biochem. Biophys. Res. Commun. 2016, 47, 539–544. [Google Scholar] [CrossRef]
- Zhang, F.; Su, K.; Yang, X.; Bowe, D.B.; Paterson, A.J.; Kudlow, J.E. O-GlcNAc modification is an endogenous inhibitor of the proteasome. Cell 2003, 115, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yin, X.; Yang, H.; Xu, Y. Histone demethylase LSD2 acts as an E3 ubiquitin ligase and inhibits cancer cell growth through promoting proteasomal degradation of OGT. Mol. Cell 2015, 58, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.G.; Kim, H.B.; Yoon, J.Y.; Kweon, T.H.; Park, Y.S.; Kang, J.; Jung, J.; Son, S.; Yi, E.C.; Lee, T.H.; et al. Mutual regulation between OGT and XIAP to control colon cancer cell growth and invasion. Cell Death Dis. 2020, 11, 815. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M. Mechanisms underlying ubiquitination. Ann. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Ann. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef]
- Xu, P.; Duong, D.M.; Seyfried, N.T.; Cheng, D.; Xie, Y.; Robert, J.; Rush, J.; Hochstrasser, M.; Finley, D.; Peng, J. Quantitative proteomics reveals the function of unconventional ubiquitin chains in proteasomal degradation. Cell 2009, 137, 133–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrends, C.; Harper, J.W. Constructing and decoding unconventional ubiquitin chains. Nat. Struct. Mol. Biol. 2011, 18, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Tsai, Y.C.; Jin, B.; Wang, B.; Wang, Y.; Zhou, H.; Carpenter, T.; Weissman, A.M.; Yin, J. Protein Engineering in the Ubiquitin System: Tools for Discovery and Beyond. Pharmacol. Rev. 2020, 72, 380–413. [Google Scholar] [CrossRef]
- Zhao, B.; Bhuripanyo, K.; Zhang, K.; Kiyokawa, H.; Schindelin, H.; Yin, J. Orthogonal Ubiquitin Transfer through Engineered E1-E2 Cascades for Protein Ubiquitination. Chem. Biol. 2012, 19, 1265–1277. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fang, S.; Chen, G.; Ganti, R.; Chernova, T.A.; Zhou, L.; Duong, D.; Kiyokawa, H.; Li, M.; Zhao, B.; et al. Regulation of the endocytosis and prion-chaperoning machineries by yeast E3 ubiquitin ligase Rsp5 as revealed by orthogonal ubiquitin transfer. Cell Chem. Biol. 2021, 28, 1283–1297. [Google Scholar] [CrossRef]
- Bhuripanyo, K.; Wang, Y.; Liu, X.; Zhou, L.; Liu, R.; Duong, D.; Zhao, B.; Bi, Y.; Zhou, H.; Chen, G.; et al. Identifying the substrate proteins of U-box E3s E4B and CHIP by orthogonal ubiquitin transfer. Sci. Adv. 2018, 4, e1701393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, X.; Zhou, L.; Duong, D.; Bhuripanyo, K.; Zhao, B.; Zhou, H.; Liu, R.; Bi, Y.; Kiyokawa, H.; et al. Identifying the ubiquitination targets of E6AP by orthogonal ubiquitin transfer. Nat. Commun. 2017, 8, 2232. [Google Scholar] [CrossRef] [Green Version]
- Pol, S.B.V.; Klingelhutz, A.J. Papillomavirus E6 oncoproteins. Virology 2013, 445, 115–137. [Google Scholar]
- Mortensen, F.; Schneider, D.; Barbic, T.; Sladewska-Marquardt, A.; Kühnle, S.; Marx, A.; Scheffner, M. Role of ubiquitin and the HPV E6 oncoprotein in E6AP-mediated ubiquitination. Proc. Natl. Acad. Sci. USA 2015, 112, 9872–9877. [Google Scholar] [CrossRef] [Green Version]
- Hornbeck, P.V.; Kornhauser, J.M.; Latham, V.; Murray, B.; Nandhikonda, V.; Nord, A.; Skrzypek, E.; Wheeler, T.; Zhang, B.; Gnad, F. 15 years of PhosphoSitePlus®: Integrating post-translationally modified sites, disease variants and isoforms. Nucleic Acids Res. 2019, 47, D433–D441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.; Tomaic, V.; Pim, D.; Myers, M.P.; Tommasino, M.; Banks, L. Interactions between E6AP and E6 proteins from alpha and beta HPV types. Virology 2013, 435, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Kao, W.H.; Beaudenon, S.L.; Talis, A.L.; Huibregtse, J.M.; Howley, P.M. Human papillomavirus type 16 E6 induces self-ubiquitination of the E6AP ubiquitin-protein ligase. J. Virol. 2000, 74, 6408–6417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seedorf, K.; Oltersdorf, T.; Krammer, G.; Rowekamp, W. Identification of early proteins of the human papilloma viruses type 16 (HPV 16) and type 18 (HPV 18) in cervical carcinoma cells. Embo J. 1987, 6, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Ruba, A.; Yang, W. O-GlcNAc-ylation in the nuclear pore complex. Cell Mol. Bioeng. 2016, 9, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Lubas, W.; Smith, M.; Starr, C.; Hanover, J. Analysis of nuclear pore protein p62 glycosylation. Biochemistry 1995, 34, 1686–1694. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi-Hata, C.; Ogawa, Y.; Oka, M.; Yoneda, Y. Quantitative regulation of nuclear pore complex proteins by O-GlcNAcylation. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 2682–2689. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Su, K.; Roos, M.D.; Chang, Q.; Paterson, A.J.; Kudlow, J.E. O-linkage of N-acetylglucosamine to Sp1 activation domain inhibits its transcriptional capability. Proc. Natl. Acad. Sci. USA 2001, 98, 6611–6616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issad, T.; Kuo, M. O-GlcNAc modification of transcription factors, glucose sensing and glucotoxicity. Trends Endocrinol. Metab. 2008, 19, 380–389. [Google Scholar] [CrossRef]
- Butkinaree, C.; Park, K.; Hart, G.W. O-linked β-N-acetylglucosamine (O-GlcNAc): Extensive crosstalk with phosphorylation to regulate signaling and transcription in response to nutrients and stress. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullen, J.W.; Balsbaugh, J.L.; Chanda, D.; Shabanowitz, J.; Hunt, D.F.; Neumann, D.; Hart, G.W. Cross-talk between two essential nutrient-sensitive enzymes: O-GlcNAc transferase (OGT) and AMP-activated protein kinase (AMPK). J. Biol. Chem. 2014, 289, 10592–10606. [Google Scholar] [CrossRef] [Green Version]
- Dias, W.B.; Hart, G.W. O-GlcNAc modification in diabetes and Alzheimer’s disease. Mol. Biosyst. 2007, 3, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Slawson, C.; Hart, G.W. O-GlcNAc signalling: Implications for cancer cell biology. Nat. Rev. Cancer 2011, 11, 678–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, Z.; Yu, S.; Xu, J. Proteasomal degradation of O-GlcNAc transferase elevates hypoxia-induced vascular endothelial inflammatory responsedagger. Cardiovasc. Res. 2014, 103, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Snead, C.M.; Catravas, J.D. Hsp90 regulates O-linked beta-N-acetylglucosamine transferase: A novel mechanism of modulation of protein O-linked beta-N-acetylglucosamine modification in endothelial cells. Am. J. Physiol. Cell Physiol. 2012, 302, C1786–C1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giles, A.C.; Desbois, M.; Opperman, K.J.; Tavora, R.; Maroni, M.J.; Grill, B. A complex containing the O-GlcNAc transferase OGT-1 and the ubiquitin ligase EEL-1 regulates GABA neuron function. J. Biol. Chem. 2019, 294, 6843–6856. [Google Scholar] [CrossRef]
- Zeng, Q.; Zhao, R.-X.; Chen, J.; Li, Y.; Li, X.-D.; Liu, X.-L.; Zhang, W.-M.; Quan, C.-S.; Wang, Y.-S.; Zhai, Y.-X.; et al. O-linked GlcNAcylation elevated by HPV E6 mediates viral oncogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, 9333–9338. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Kim, Y.S.; Kim, H.; Kang, M.Y.; Park, J.; Lee, D.H.; Roh, G.S.; Kim, H.J.; Kang, S.S.; Cho, G.J.; et al. O-linked N-acetylglucosamine transferase promotes cervical cancer tumorigenesis through human papillomaviruses E6 and E7 oncogenes. Oncotarget 2016, 7, 44596–44607. [Google Scholar] [CrossRef] [Green Version]
- Jochmann, R.; Thurau, M.; Jung, S.; Hofmann, C.; Naschberger, E.; Kremmer, E.; Harrer, T.; Miller, M.; Schaft, N.; Sturzl, M. O-linked N-acetylglucosaminylation of Sp1 inhibits the human immunodeficiency virus type 1 promoter. J. Virol. 2009, 83, 3704–3718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.C.; Tsai, W.H.; Wang, P.W.; Wu, I.L.; Lin, S.Y.; Chen, Y.L.; Chen, J.Y.; Lin, S.F. Suppressive regulation of KSHV RTA with O-GlcNAcylation. J. Biomed. Sci. 2012, 19, 12. [Google Scholar] [CrossRef] [Green Version]
- Jochmann, R.; Pfannstiel, J.; Chudasama, P.; Kuhn, E.; Konrad, A.; Sturzl, M. O-GlcNAc transferase inhibits KSHV propagation and modifies replication relevant viral proteins as detected by systematic O-GlcNAcylation analysis. Glycobiology 2013, 23, 1114–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lin, Y.; Liu, S.; Zhu, Y.; Lu, K.; Broering, R.; Lu, M. O-GlcNAcylation modulates HBV replication through regulating cellular autophagy at multiple levels. FASEB J. 2020, 34, 14473–14489. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Gao, Q.; Yang, Y.; Xia, J.; Zhang, W.; Chen, Y.; Zhou, Z.; Chang, L.; Hu, Y.; Zhou, H.; et al. Hexosamine biosynthetic pathway promotes the antiviral activity of SAMHD1 by enhancing O-GlcNAc transferase-mediated protein O-GlcNAcylation. Theranostics 2021, 11, 805–823. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.; Pol, S.V.; Banerjee, N.S.; Dutta, A.B.; Chow, L.T.; Dutta, A. Destabilization of TIP60 by human papillomavirus E6 results in attenuation of TIP60-dependent transcriptional regulation and apoptotic pathway. Mol. Cell 2010, 38, 700–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbaiah, V.; Zhang, Y.; Rajagopalan, D.; Abdullah, L.; Yeo-Teh, N.; Tomaić, V.; Banks, L.; Myers, M.; Chow, E.; Jha, S. E3 ligase EDD1/UBR5 is utilized by the HPV E6 oncogene to destabilize tumor suppressor TIP60. Oncogene 2016, 35, 2062–2074. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, K.; Liu, R.; Jia, C.; Wang, Y.; Jeong, G.H.; Zhou, L.; Hu, R.; Kiyokawa, H.; Yin, J.; Zhao, B. Regulation of O-Linked N-Acetyl Glucosamine Transferase (OGT) through E6 Stimulation of the Ubiquitin Ligase Activity of E6AP. Int. J. Mol. Sci. 2021, 22, 10286. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910286
Peng K, Liu R, Jia C, Wang Y, Jeong GH, Zhou L, Hu R, Kiyokawa H, Yin J, Zhao B. Regulation of O-Linked N-Acetyl Glucosamine Transferase (OGT) through E6 Stimulation of the Ubiquitin Ligase Activity of E6AP. International Journal of Molecular Sciences. 2021; 22(19):10286. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910286
Chicago/Turabian StylePeng, Kangli, Ruochuan Liu, Caiwei Jia, Yiyang Wang, Geon H. Jeong, Li Zhou, Ronggui Hu, Hiroaki Kiyokawa, Jun Yin, and Bo Zhao. 2021. "Regulation of O-Linked N-Acetyl Glucosamine Transferase (OGT) through E6 Stimulation of the Ubiquitin Ligase Activity of E6AP" International Journal of Molecular Sciences 22, no. 19: 10286. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910286