
KIT Expression Is Regulated by DNA Methylation in Uveal Melanoma Tumors

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Clinicopathologic Characteristics of the Patients
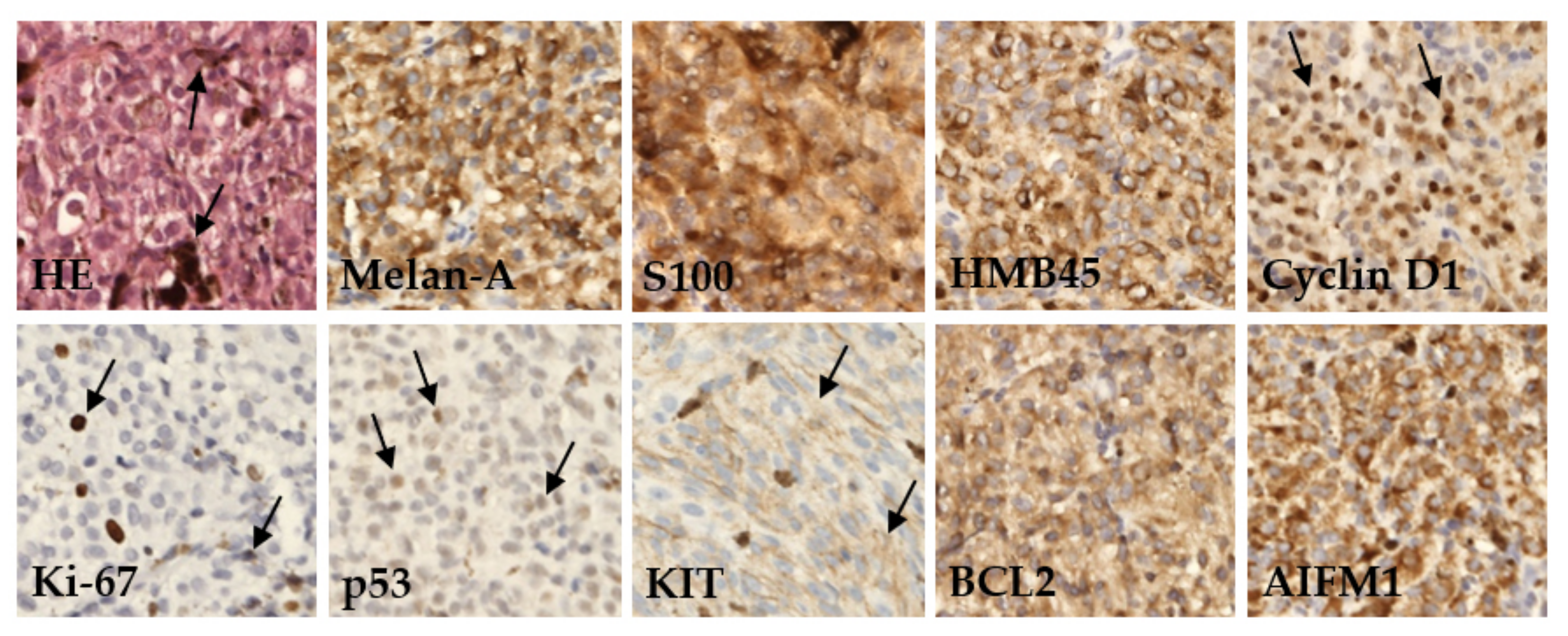
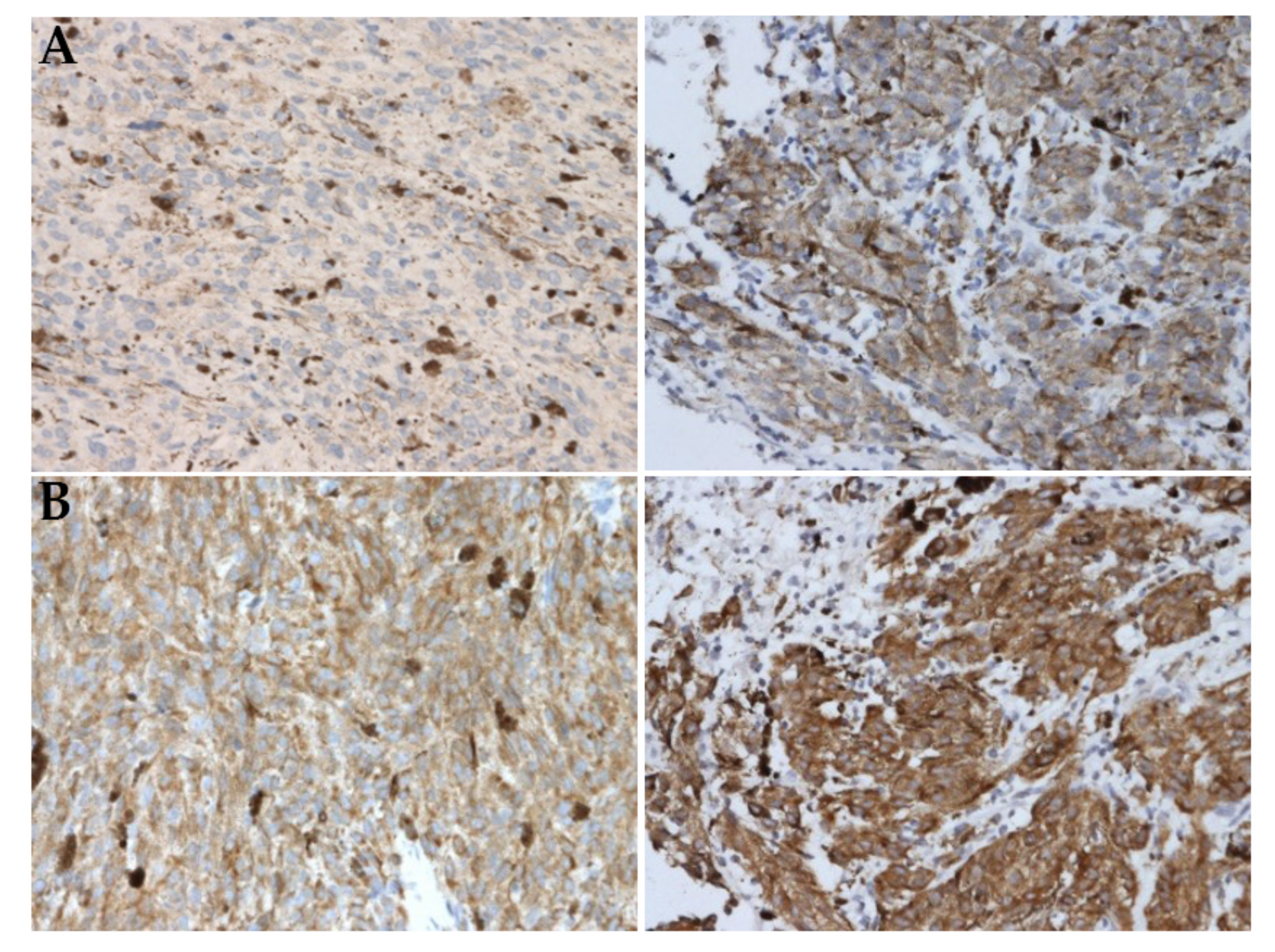
2.2. Immunohistochemistry
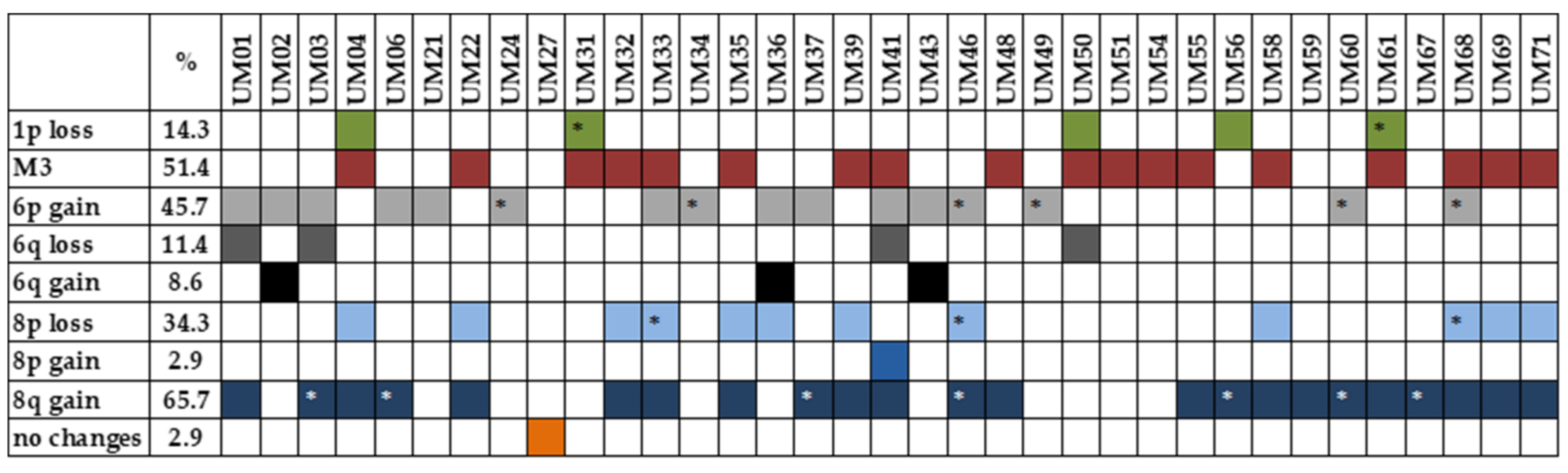
2.3. MLPA Analysis
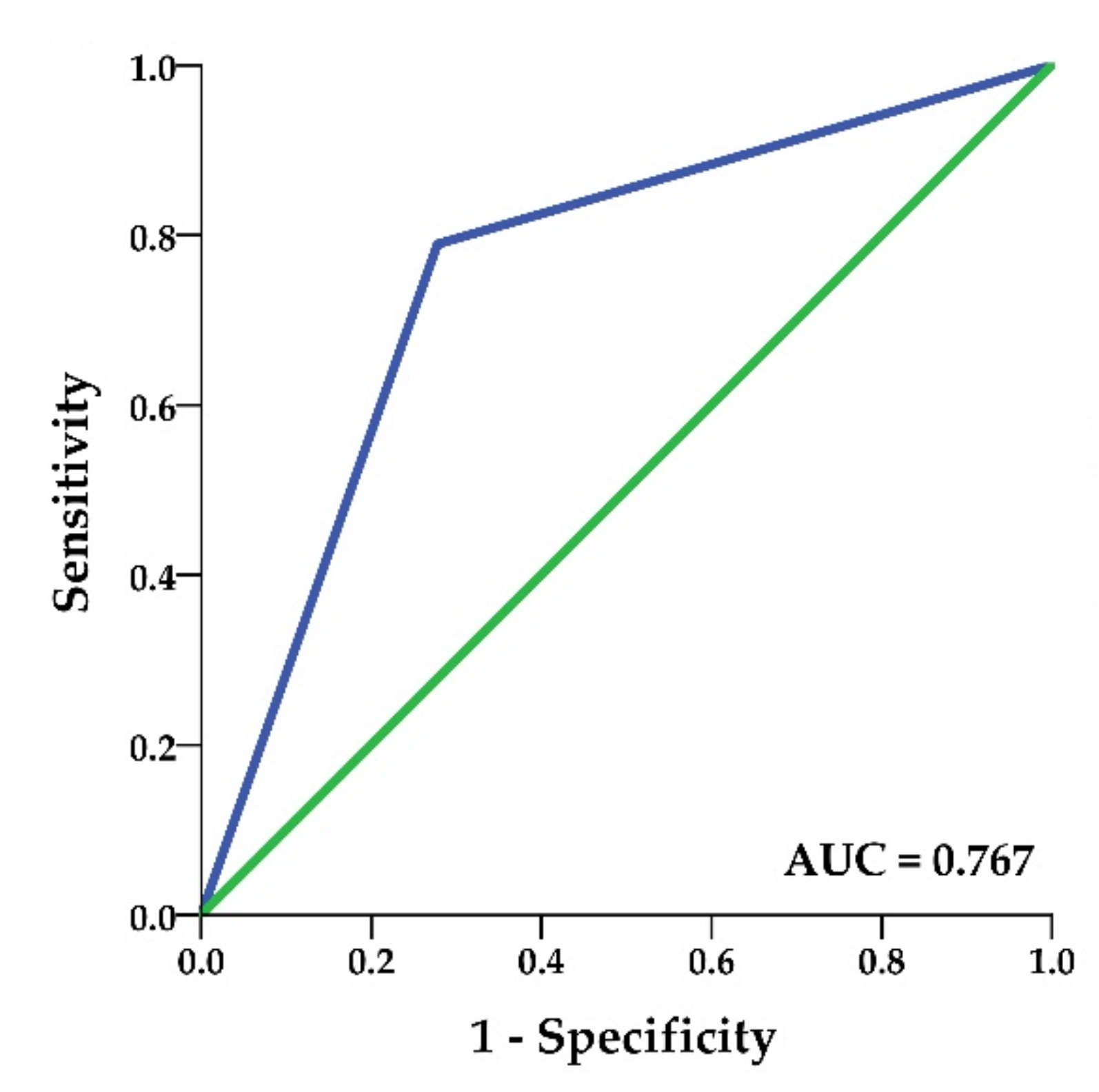
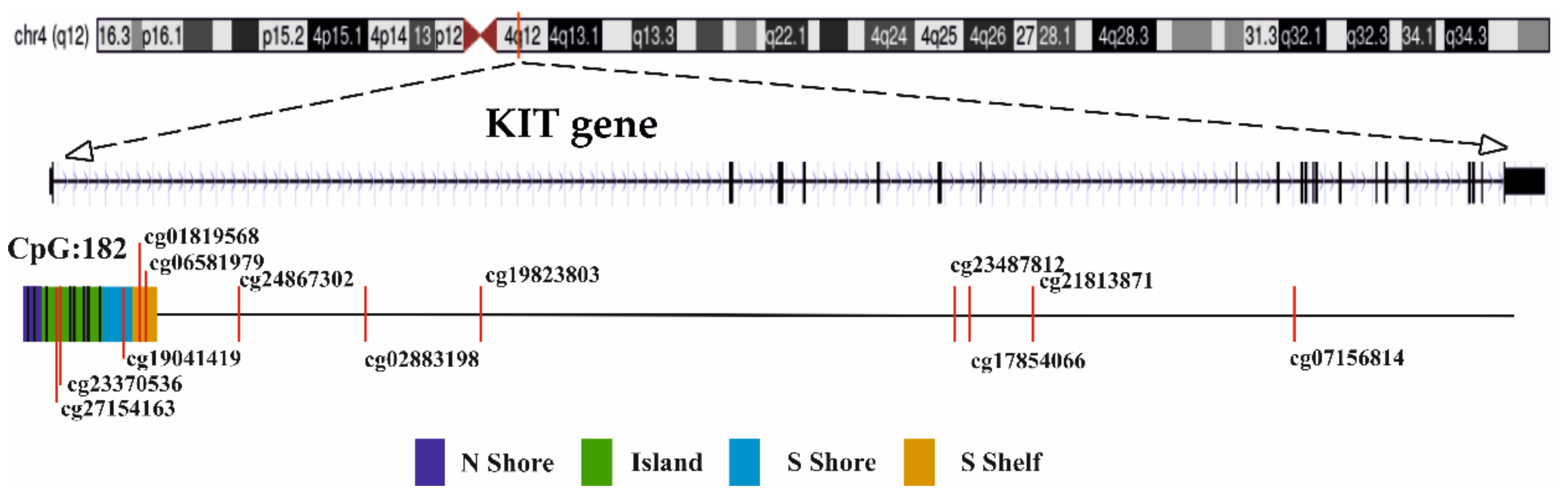
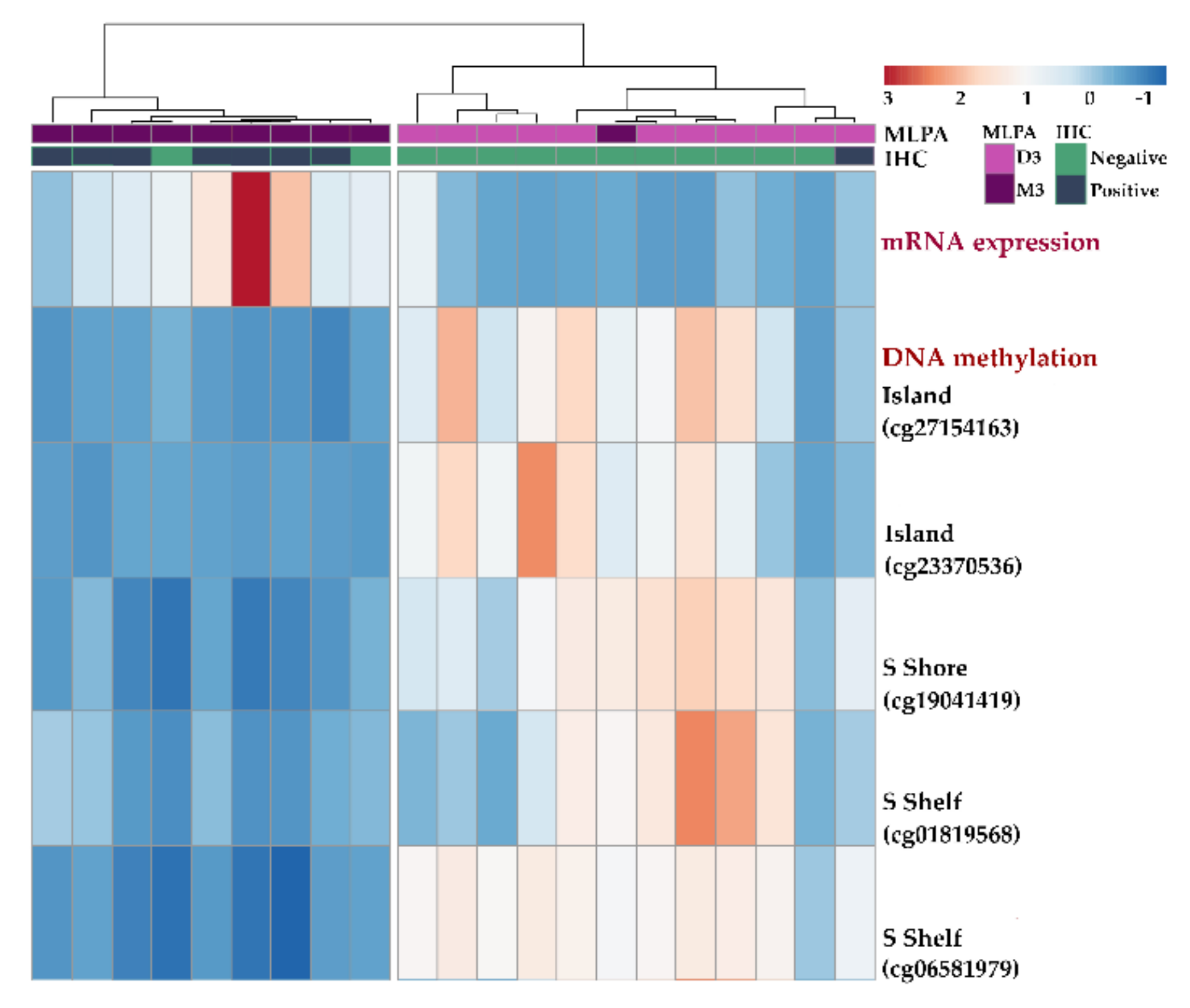
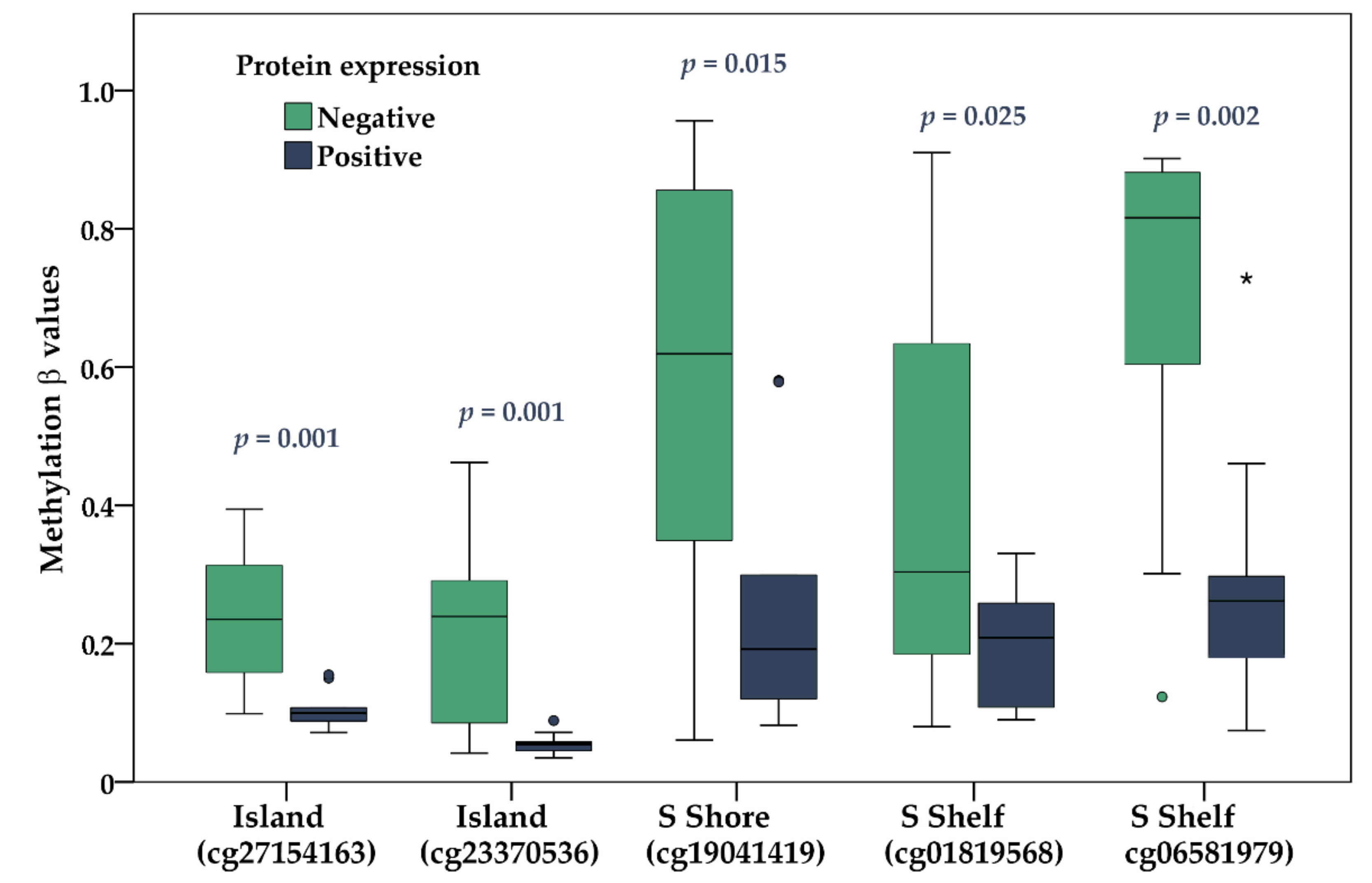
2.4. Gene Expression and DNA Methylation of KIT Gene
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Immunohistochemistry
4.3. MLPA Analysis
4.4. Microarray Analysis
4.5. Methylation Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AIFM1 | Apoptosis-inducing factor 1: mitochondrial |
| AUC | Area under curve |
| BAP1 | BRCA1 associated protein 1 |
| BCL2 | Apoptosis regulator Bcl-2 |
| CI | Confidence interval |
| CM | Cutaneous melanoma |
| CpG | Cytosines followed by guanine residues |
| CTCF | Transcription repressor |
| Cyclin D1 | Cyclin-D1-binding protein 1 |
| D3 | Disomy 3 |
| EIF1AX | Eukaryotic translation initiation factor 1A X-linked |
| FC | Fold change analysis |
| FFPE | Formalin-fixed, paraffin-embedded tumors |
| HE | Hematoxylin and eosin |
| HMB45 | Human melanoma black |
| IHC | Immunohistochemistry |
| IQR | Interquartile range |
| KIT | Mast/stem cell growth factor receptor Kit, alternative name CD117 |
| Ki-67 | Proliferation marker protein Ki-67 |
| Melan A | Melanoma antigen recognized by T-cells 1 |
| MITF | Melanocyte-inducing transcription factor |
| MLPA | Multiplex ligation-dependent probe amplification |
| M3 | Monosomy of chromosome 3 |
| OR | Odds ratio |
| p53 | Cellular tumor antigen p53 |
| r | Correlation coefficient |
| ROC | Receiver operating characteristic |
| SCF | Stem cell factor |
| SF3B1 | Splicing factor 3b subunit 1 |
| SRS | Stereotactic radiosurgery |
| S100 | S100 calcium binding protein |
| TNM | Classification of malignant tumors, T—site and size of primary tumor, N—regional lymph nodes involvement, M—presence of distant metastasis |
| QS | Quick score |
| UM | Uveal melanoma |
| 1p | Short arm of chromosome 1 |
| 6p | Short arm of chromosome 6 |
| 6q | Long arm of chromosome 6 |
| 8p | Short arm of chromosome 8 |
| 8q | Long arm of chromosome 8 |
References
- Singh, A.D.; Turell, M.E.; Topham, A.K. Uveal melanoma: Trends in incidence, treatment, and survival. Ophthalmology 2011, 118, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, C.C.; Wu, X.C.; Jemal, A.; Martin, H.J.; Roche, L.M.; Chen, V.W. Incidence of noncutaneous melanomas in the U.S. Cancer 2005, 103, 1000–1007. [Google Scholar] [CrossRef]
- Eskelin, S.; Pyrhönen, S.; Summanen, P.; Hahka-Kemppinen, M.; Kivelä, T. Tumor doubling times in metastatic malignant melanoma of the uvea: Tumor progression before and after treatment. Ophthalmology 2000, 107, 1443–1449. [Google Scholar] [CrossRef]
- Diener-West, M.; Reynolds, S.M.; Agugliaro, D.J.; Caldwell, R.; Cumming, K.; Earle, J.D.; Hawkins, B.S.; Hayman, J.A.; Jaiyesimi, I.; Jampol, L.M.; et al. Development of metastatic disease after enrollment in the COMS trials for treatment of choroidal melanoma: Collaborative Ocular Melanoma Study Group Report No. 26. Arch. Ophthalmol. 2005, 123, 1639–1643. [Google Scholar] [PubMed]
- Rantala, E.S.; Hernberg, M.; Kivelä, T.T. Overall survival after treatment for metastatic uveal melanoma: A systematic review and meta-analysis. Melanoma Res. 2019, 29, 561–568. [Google Scholar] [CrossRef]
- Damato, B.; Coupland, S.E. Translating Uveal Melanoma Cytogenetics into Clinical Care. Arch. Ophthalmol. 2009, 127, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Onken, M.D.; Worley, L.A.; Ehlers, J.P.; Harbour, J.W. Gene expression profiling in uveal melanoma reveals two molecular classes and predicts metastatic death. Cancer Res. 2004, 64, 7205–7209. [Google Scholar] [CrossRef] [Green Version]
- Johansson, P.A.; Brooks, K.; Newell, F.; Palmer, J.M.; Wilmott, J.S.; Pritchard, A.L.; Broit, N.; Wood, S.; Carlino, M.S.; Leonard, C.; et al. Whole genome landscapes of uveal melanoma show an ultraviolet radiation signature in iris tumours. Nat. Commun. 2020, 11, 2408. [Google Scholar] [CrossRef]
- Prescher, G.; Bornfeld, N.; Becher, R. Nonrandom chromosomal abnormalities in primary uveal melanoma. J. Natl. Cancer Inst. 1990, 82, 1765–1769. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, J.P.; Worley, L.; Onken, M.D.; Harbour, J.W. Integrative genomic analysis of aneuploidy in uveal melanoma. Clin. Cancer Res. 2008, 14, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Ewens, K.G.; Kanetsky, P.A.; Richards-Yutz, J.; Al-Dahmash, S.; De Luca, M.C.; Bianciotto, C.G.; Shields, C.L.; Ganguly, A. Genomic Profile of 320 Uveal Melanoma Cases: Chromosome 8p-Loss and Metastatic Outcome. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5721–5729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versluis, M.; De Lange, M.J.; Van Pelt, S.I.; Ruivenkamp, C.A.; Kroes, W.G.; Cao, J.; Jager, M.J.; Luyten, G.P.; Van der Velden, P.A. Digital PCR validates 8q dosage as prognostic tool in uveal melanoma. PLoS ONE 2015, 10, e0116371. [Google Scholar] [CrossRef] [Green Version]
- Amaro, A.; Gangemi, R.; Piaggio, F.; Angelini, G.; Barisione, G.; Ferrini, S.; Pfeffer, U. The biology of uveal melanoma. Cancer Metastasis Rev. 2017, 36, 109–140. [Google Scholar] [CrossRef]
- Onken, M.D.; Worley, L.A.; Char, D.H.; Augsburger, J.J.; Correa, Z.M.; Nudleman, E.; Aaberg, T.M., Jr.; Altaweel, M.M.; Bardenstein, D.S.; Finger, P.T.; et al. Collaborative Ocular Oncology Group report number 1: Prospective validation of a multi-gene prognostic assay in uveal melanoma. Ophthalmology 2012, 119, 1596–1603. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.G.; Shih, J.; Yau, C.; Gibb, E.A.; Oba, J.; Mungall, K.L.; Hess, J.M.; Uzunangelov, V.; Walter, V.; Danilova, L.; et al. Integrative Analysis Identifies Four Molecular and Clinical Subsets in Uveal Melanoma. Cancer Cell 2017, 32, 204–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jager, M.J.; Shields, C.L.; Cebulla, C.M.; Abdel-Rahman, M.H.; Grossniklaus, H.E.; Stern, M.H.; Carvajal, R.D.; Belfort, R.N.; Jia, R.; Shields, J.A.; et al. Uveal melanoma. Nat. Rev. Dis. Primers 2020, 6, 020–0158. [Google Scholar] [CrossRef] [PubMed]
- Smit, K.N.; Jager, M.J.; De Klein, A.; Kiliç, E. Uveal melanoma: Towards a molecular understanding. Prog. Retin. Eye Res. 2020, 75, 26. [Google Scholar] [CrossRef]
- Schuster, R.; Bechrakis, N.E.; Stroux, A.; Busse, A.; Schmittel, A.; Scheibenbogen, C.; Thiel, E.; Foerster, M.H.; Keilholz, U. Circulating tumor cells as prognostic factor for distant metastases and survival in patients with primary uveal melanoma. Clin. Cancer Res. 2007, 13, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, S.; Burrows, R.C.; Kalina, R.E.; George, D.; Boehm, M.; Bothwell, M.A.; Schmidt, R. Immunophenotypic differences between uveal and cutaneous melanomas. Arch. Ophthalmol. 2002, 120, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Coupland, S.E.; Anastassiou, G.; Stang, A.; Schilling, H.; Anagnostopoulos, I.; Bornfeld, N.; Stein, H. The prognostic value of cyclin D1, p53, and MDM2 protein expression in uveal melanoma. J. Pathol. 2000, 191, 120–126. [Google Scholar] [CrossRef]
- Lukenda, A.; Dotlic, S.; Vukojevic, N.; Saric, B.; Vranic, S.; Zarkovic, K. Expression and prognostic value of putative cancer stem cell markers CD117 and CD15 in choroidal and ciliary body melanoma. J. Clin. Pathol. 2016, 69, 234–239. [Google Scholar] [CrossRef] [Green Version]
- All-Ericsson, C.; Girnita, L.; Müller-Brunotte, A.; Brodin, B.; Seregard, S.; Ostman, A.; Larsson, O. c-Kit-dependent growth of uveal melanoma cells: A potential therapeutic target? Investig. Ophthalmol. Vis. Sci. 2004, 45, 2075–2082. [Google Scholar] [CrossRef] [PubMed]
- Soltysova, A.; Sedlackova, T.; Dvorska, D.; Jasek, K.; Chokhachi Baradaran, P.; Horvathova Kajabova, V.; Demkova, L.; Buocikova, V.; Kurucova, T.; Lyskova, D.; et al. Monosomy 3 Influences Epithelial-Mesenchymal Transition Gene Expression in Uveal Melanoma Patients; Consequences for Liquid Biopsy. Int. J. Mol. Sci. 2020, 21, 9651. [Google Scholar] [CrossRef]
- Hussein, M.R. Analysis of Bcl-2 protein expression in choroidal melanomas. J. Clin. Pathol. 2005, 58, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Krasnik, V.; Furdova, A.; Svetlosakova, Z.; Kobzova, D.; Gergisakova, H.; Feketeova, L.; Svetlosak, M.; Barta, A.; Babal, P. Prognostic value of apoptosis inducing factor in uveal melanoma. Neoplasma 2017, 64, 262–268. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef] [PubMed]
- Romero-Garcia, S.; Prado-Garcia, H.; Carlos-Reyes, A. Role of DNA Methylation in the Resistance to Therapy in Solid Tumors. Front. Oncol. 2020, 10, 1152. [Google Scholar] [CrossRef]
- Kim, J.; Bretz, C.L.; Lee, S. Epigenetic instability of imprinted genes in human cancers. Nucleic Acids Res. 2015, 43, 10689–10699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locke, W.J.; Guanzon, D.; Ma, C.; Liew, Y.J.; Duesing, K.R.; Fung, K.Y.C.; Ross, J.P. DNA Methylation Cancer Biomarkers: Translation to the Clinic. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- White, V.A.; Chambers, J.D.; Courtright, P.D.; Chang, W.Y.; Horsman, D.E. Correlation of cytogenetic abnormalities with the outcome of patients with uveal melanoma. Cancer 1998, 83, 354–359. [Google Scholar] [CrossRef]
- Shields, C.L.; Say, E.A.T.; Hasanreisoglu, M.; Saktanasate, J.; Lawson, B.M.; Landy, J.E.; Badami, A.U.; Sivalingam, M.D.; Hauschild, A.J.; House, R.J.; et al. Personalized Prognosis of Uveal Melanoma Based on Cytogenetic Profile in 1059 Patients over an 8-Year Period: The 2017 Harry S. Gradle Lecture. Ophthalmology 2017, 124, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Prescher, G.; Bornfeld, N.; Hirche, H.; Horsthemke, B.; Jöckel, K.H.; Becher, R. Prognostic implications of monosomy 3 in uveal melanoma. Lancet 1996, 347, 1222–1225. [Google Scholar]
- Damato, B.; Dopierala, J.A.; Coupland, S.E. Genotypic profiling of 452 choroidal melanomas with multiplex ligation-dependent probe amplification. Clin. Cancer Res. 2010, 16, 6083–6092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasota, J.; Miettinen, M. Clinical significance of oncogenic KIT and PDGFRA mutations in gastrointestinal stromal tumours. Histopathology 2008, 53, 245–266. [Google Scholar] [CrossRef]
- Williams, D.E.; Eisenman, J.; Baird, A.; Rauch, C.; Van Ness, K.; March, C.J.; Park, L.S.; Martin, U.; Mochizuki, D.Y.; Boswell, H.S.; et al. Identification of a ligand for the c-kit proto-oncogene. Cell 1990, 63, 167–174. [Google Scholar] [CrossRef]
- Dahl, C.; Abildgaard, C.; Riber-Hansen, R.; Steiniche, T.; Lade-Keller, J.; Guldberg, P. KIT is a frequent target for epigenetic silencing in cutaneous melanoma. J. Investig. Dermatol. 2015, 135, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Mouriaux, F.; Kherrouche, Z.; Maurage, C.A.; Demailly, F.X.; Labalette, P.; Saule, S. Expression of the c-kit receptor in choroidal melanomas. Melanoma Res. 2003, 13, 161–166. [Google Scholar] [CrossRef]
- Lüke, J.; Wegner, J.; Wegner, R.; Nassar, K.; Tatar, O.; Rohrbach, J.M.; Hilgers, R.D.; Lüke, M.; Grisanti, S. Expression of c-Kit and its ligand SCF in primary uveal melanoma. Eur. J. Ophthalmol. 2011, 21, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.R.; Odashiro, A.N.; Marshall, J.C.; Correa, Z.M.; Belfort, R., Jr.; Burnier, M.N., Jr. The role of c-kit and imatinib mesylate in uveal melanoma. J. Carcinog. 2005, 4, 19. [Google Scholar] [CrossRef]
- De Wit, N.J.; Van Muijen, G.N.; Ruiter, D.J. Immunohistochemistry in melanocytic proliferative lesions. Histopathology 2004, 44, 517–541. [Google Scholar] [CrossRef]
- Velez, G.; Nguyen, H.V.; Chemudupati, T.; Ludwig, C.A.; Toral, M.; Reddy, S.; Mruthyunjaya, P.; Mahajan, V.B. Liquid biopsy proteomics of uveal melanoma reveals biomarkers associated with metastatic risk. Mol. Cancer 2021, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.S.; Shi, L.; Foster, B.M.; Mobley, M.E.; Elliott, P.L.; Song, C.J.; Watabe, K.; Langefeld, C.D.; Kerr, B.A. CD117/c-kit defines a prostate CSC-like subpopulation driving progression and TKI resistance. Sci. Rep. 2021, 11, 1465. [Google Scholar] [CrossRef] [PubMed]
- Foster, B.; Zaidi, D.; Young, T.; Mobley, M.; Kerr, B. CD117/c-kit in cancer stem cell-mediated progression and therapeutic resistance. Biomedicines 2018, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Debiec-Rychter, M.; Dumez, H.; Judson, I.; Wasag, B.; Verweij, J.; Brown, M.; Dimitrijevic, S.; Sciot, R.; Stul, M.; Vranck, H.; et al. Use of c-KIT/PDGFRA mutational analysis to predict the clinical response to imatinib in patients with advanced gastrointestinal stromal tumours entered on phase I and II studies of the EORTC Soft Tissue and Bone Sarcoma Group. Eur. J. Cancer 2004, 40, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, U.B.; Kauczok-Vetter, C.S.; Houben, R.; Becker, J.C. Overexpression of the KIT/SCF in uveal melanoma does not translate into clinical efficacy of imatinib mesylate. Clin. Cancer Res. 2009, 15, 324–329. [Google Scholar] [CrossRef] [Green Version]
- Wallander, M.L.; Layfield, L.J.; Emerson, L.L.; Mamalis, N.; Davis, D.; Tripp, S.R.; Holden, J.A. KIT mutations in ocular melanoma: Frequency and anatomic distribution. Mod. Pathol. 2011, 24, 1031–1035. [Google Scholar] [CrossRef]
- Janostiak, R.; Vyas, M.; Cicek, A.F.; Wajapeyee, N.; Harigopal, M. Loss of c-KIT expression in breast cancer correlates with malignant transformation of breast epithelium and is mediated by KIT gene promoter DNA hypermethylation. Exp. Mol. Pathol. 2018, 105, 41–49. [Google Scholar] [CrossRef]
- Chang, S.-W.; Chao, W.-R.; Ruan, A.; Wang, P.-H.; Lin, J.-C.; Han, C.-P. A promising hypothesis of c-KIT methylation/ expression paradox in c-KIT (+) squamous cell carcinoma of uterine cervix ----- CTCF transcriptional repressor regulates c-KIT proto-oncogene expression. Diagn. Pathol. 2015, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Martino, D.; Saffery, R. Characteristics of DNA methylation and gene expression in regulatory features on the Infinium 450k Beadchip. bioRxiv 2015, 032862. [Google Scholar] [CrossRef] [Green Version]
- Siemens, H.; Jackstadt, R.; Kaller, M.; Hermeking, H. Repression of c-Kit by p53 is mediated by miR-34 and is associated with reduced chemoresistance, migration and stemness. Oncotarget 2013, 4, 1399–1415. [Google Scholar] [CrossRef] [Green Version]
- Hemming, M.L.; Lawlor, M.A.; Andersen, J.L.; Hagan, T.; Chipashvili, O.; Scott, T.G.; Raut, C.P.; Sicinska, E.; Armstrong, S.A.; Demetri, G.D.; et al. Enhancer Domains in Gastrointestinal Stromal Tumor Regulate KIT Expression and Are Targetable by BET Bromodomain Inhibition. Cancer Res. 2019, 79, 994–1009. [Google Scholar] [CrossRef] [Green Version]
- Chokhachi Baradaran, P.; Kozovska, Z.; Furdova, A.; Smolkova, B. Targeting Epigenetic Modifications in Uveal Melanoma. Int. J. Mol. Sci. 2020, 21, 5314. [Google Scholar] [CrossRef]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th edition of the AJCC cancer staging manual and the future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef]
- Viray, H.; Bradley, W.R.; Schalper, K.A.; Rimm, D.L.; Rothberg, B.E.G. Marginal and joint distributions of S100, HMB-45, and Melan-A across a large series of cutaneous melanomas. Arch. Pathol. Lab. Med. 2013, 137, 1063–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, N.A.; Yussif, S.M. Ki-67 as a prognostic marker according to breast cancer molecular subtype. Cancer Biol. Med. 2016, 13, 496–504. [Google Scholar]
- Chana, J.S.; Wilson, G.D.; Cree, I.A.; Alexander, R.A.; Myatt, N.; Neale, M.; Foss, A.J.; Hungerford, J.L. c-myc, p53, and Bcl-2 expression and clinical outcome in uveal melanoma. Br. J. Ophthalmol. 1999, 83, 110–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gass, J.D. Comparison of uveal melanoma growth rates with mitotic index and mortality. Arch. Ophthalmol. 1985, 103, 924–931. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathologic Characteristics | All n (%) | Primary UM n (%) | Metastatic UM n (%) | p-Value |
|---|---|---|---|---|
| Gender | ||||
| Male | 26 (51.0) | 23 (56.1) | 3 (30.0) | 0.139 |
| Female | 25 (49.0) | 18 (43.9) | 7 (70.0) | |
| Eye | ||||
| Right | 29 (56.9) | 25 (61.0) | 4 (40.0) | 0.230 |
| Left | 22 (43.1) | 16 (39.0) | 6 (60.0) | |
| Median age (range) | 65.0 years (32–87) | 66.5 years (35–80) | 63.0 years (32–87) | 0.658 |
| Median tumor volume (range) | 1.3 cm3 (0.3–2.6) | 1.3 cm3 (0.3–2.6) | 1.45 cm3 (0.3–2.6) | 0.694 |
| <1.55 cm3 | 34 (66.7) | 27 (65.9) | 7 (70.0) | 0.803 |
| ≥1.55 cm3 | 17 (33.3) | 14 (34.1) | 3 (30.0) | |
| Diagnosis | ||||
| C69.3 | 40 (78.4) | 34 (82.9) | 6 (60.0) | 0.114 |
| C69.4 | 11 (21.6) | 7 (17.1) | 4 (40.0) | |
| Cell type * | ||||
| Spindle | 26 (51.0) | 22 (53.7) | 4 (40.0) | 0.612 |
| Epitheloid and Mixed | 24 (47.0) | 18 (43.9) | 6 (60.0) | |
| Therapy | ||||
| Enucleation | 36 (70.6) | 27 (65.9) | 9 (90.0) | 0.133 |
| Enucleation after SRS 1 | 15 (29.4) | 14 (34.1) | 1 (10.0) | |
| Vascular invasion | ||||
| Present | 9 (17.6) | 6 (14.6) | 3 (30.0) | 0.253 |
| Absent | 42 (82.4) | 35 (85.4) | 7 (70.0) | |
| Lymphogenic invasion | ||||
| Present | 15 (29.4) | 10 (24.4) | 5 (50.0) | 0.111 |
| Absent | 36 (70.6) | 31 (75.6) | 5 (50.0) | |
| Perineural invasion | ||||
| Present | 12 (23.5) | 9 (22.0) | 3 (30.0) | 0.591 |
| Absent | 39 (76.5) | 32 (78.0) | 7 (70.0) | |
| Extrabulbar overgrowth | ||||
| Present | 8 (15.7) | 4 (9.8) | 4 (40.0) | 0.018 |
| Absent | 43 (84.3) | 37 (90.2) | 6 (60.0) | |
| TNM staging | ||||
| I–IIB | 23 (45.1) | 20 (48.8) | 3 (30.0) | 0.285 |
| IIIA–C | 28 (54.9) | 21 (51.2) | 7 (70.0) |
| IHC Markers | All n (%) | Primary UM n (%) | Metastatic UM n (%) | p-Value |
|---|---|---|---|---|
| Melan-A | ||||
| Positive, ≥25% | 48 (96.0) | 38 (95.0) | 10 (100.0) | 0.470 |
| Negative | 2 (4.0) | 2 (5.0) | 0 (0.0) | |
| S100 | ||||
| Positive, ≥25% | 46 (92.0) | 37 (92.5) | 9 (90.0) | 0.794 |
| Negative | 4 (8.0) | 3 (7.5) | 1 (10.0) | |
| HMB45 | ||||
| Positive, ≥25% | 47 (94.0) | 38 (95.0) | 9 (90.0) | 0.552 |
| Negative | 3 (6.0) | 2 (5.0) | 1 (10.0) | |
| Cyclin D1 | ||||
| Positive, >15% | 42 (84.0) | 34 (85.0) | 8 (80.0) | 0.700 |
| Negative | 8 (16.0) | 6 (15.0) | 2 (20.0) | |
| Ki-67 | ||||
| Positive, >15% | 13 (26.0) | 9 (22.5) | 4 (40.0) | 0.259 |
| Negative | 37 (74.0) | 31 (77.5) | 6 (60.0) | |
| p53 | ||||
| Positive, >15% | 15 (30.0) | 12 (30.0) | 3 (30.0) | 1.000 |
| Negative | 35 (70.0) | 28 (70.0) | 7 (70.0) | |
| KIT | ||||
| Positive, >10% | 20 (41.7) | 15 (39.5) | 5 (50) | 0.548 |
| Negative | 28 (58.3) | 23 (60.5) | 5 (50) | |
| BCL2 | ||||
| Positive, ≥ ++ | 32 (68.1) | 25 (67.6) | 7 (70.0) | 0.884 |
| Negative | 15 (31.9) | 12 (32.4) | 3 (30.0) | |
| AIFM1 | ||||
| Positive, QS 1 > 4 | 43 (91.5) | 33 (89.2) | 10 (100.0) | 0.277 |
| Negative | 4 (8.5) | 4 (10.8) | 0 (0.0) |
| Clinicopathologic Characteristics | Disomy 3 n (%) | Monosomy 3 n (%) | p-Value |
|---|---|---|---|
| Gender 1 | |||
| Male | 11 (64.7) | 10 (55.6) | 0.581 |
| Female | 6 (35.3) | 8 (44.4) | |
| Eye | |||
| Right | 11 (64.7) | 8 (44.4) | 0.229 |
| Left | 6 (35.3) | 10 (55.6) | |
| Median age (range) | 69 years (35–87) | 61 (32–81) | 0.189 |
| Median tumor volume (range) | 1.4 cm3 (0.3–2.6) | 1.5 cm3 (0.4–2.4) | 0.681 |
| <1.55 cm3 | 9 (52.9) | 10 (55.6) | 0.877 |
| ≥1.55 cm3 | 8 (47.1) | 8 (44.4) | |
| Diagnosis | |||
| C69.3 | 17 (100.0) | 11 (61.1) | 0.004 |
| C69.4 | 0 (0.0) | 7 (38.9) | |
| Cell type | |||
| Spindle | 12 (70.6) | 6 (33.3) | 0.025 |
| Epitheloid and Mixed | 5 (29.4) | 12 (66.7) | |
| Vascular invasion | |||
| Present | 4 (23.5) | 2 (11.1) | 0.330 |
| Absent | 13 (76.5) | 16 (88.9) | |
| Lymphogenic invasion | |||
| Present | 4 (23.5) | 6 (33.3) | 0.521 |
| Absent | 13 (76.5) | 12 (66.7) | |
| Perineural invasion | |||
| Present | 5 (29.4) | 5 (27.8) | 0.915 |
| Absent | 12 (70.6) | 13 (72.2) | |
| Extrabulbar overgrowth | |||
| Present | 1 (5.6) | 5 (25.0) | 0.101 |
| Absent | 17 (94.4) | 15 (75.0) | |
| TNM staging | |||
| I–IIB | 9 (52.9) | 2 (11.1) | 0.008 |
| IIIA–C | 8 (47.1) | 16 (88.9) | |
| Metastasis | |||
| Present | 3 (17.6) | 6 (33.3) | 0.289 |
| Absent | 14 (82.4) | 12 (66.7) |
| IHC Markers 1 | Disomy 3 n (%) | Monosomy 3 n (%) | p-Value |
|---|---|---|---|
| Melan-A | |||
| Positive, ≥25% | 16 (94.1) | 17 (94.4) | 0.967 |
| Negative | 1 (5.9) | 1 (5.6) | |
| S100 | |||
| Positive, ≥25% | 16 (94.1) | 15 (83.3) | 0.316 |
| Negative | 1 (5.9) | 3 (16.7) | |
| HMB45 | |||
| Positive, ≥25% | 15 (88.2) | 17 (94.4) | 0.512 |
| Negative | 2 (11.8) | 1 (5.6) | |
| Cyclin D1 | |||
| Positive, >15% | 15 (88.2) | 16 (88.9) | 0.952 |
| Negative | 2 (11.8) | 2 (11.1) | |
| Ki-67 | |||
| Positive, >15% | 3 (17.6) | 8 (44.4) | 0.088 |
| Negative | 14 (82.4) | 10 (55.6) | |
| p53 | |||
| Positive, >15% | 5 (29.4) | 7 (38.9) | 0.555 |
| Negative | 12 (70.6) | 11 (61.1) | |
| KIT | |||
| Positive, >10% | 3 (18.8) | 13 (72.2) | 0.002 |
| Negative | 13 (81.2) | 5 (27.8) | |
| BCL2 | |||
| Positive, ≥ ++ | 8 (50.0) | 15 (88.2) | 0.017 |
| Negative | 8 (50.0) | 2 (11.8) | |
| AIFM1 | |||
| Positive, QS 2 > 4 | 14 (93.3) | 17 (100.0) | 0.279 |
| Negative | 1 (6.7) | 0 (0.0) |
| Variable | OR | 95% CI | p-Value |
|---|---|---|---|
| KIT positivity | 17.41 | 1.53–198.69 | 0.021 |
| Epithelioid and Mixed cell type | 18.04 | 1.34–243.48 | 0.029 |
| T stage IIIA–C | 11.28 | 0.68–188.46 | 0.092 |
| Constant | 0.010 |
| Probe | UCSC Refgene Group | Feature | M3 vs. D3 p-Value £ | IHC * p-Value | Correlation Coefficient # r (p-Value) |
|---|---|---|---|---|---|
| cg16928454 | TSS1500 | N Shore | NS | NS | NS |
| cg06483432 | TSS1500 | N Shore | NS | NS | NS |
| cg26635759 | TSS1500 | Island | NS | NS | NS |
| cg27154163 | TSS1500 | Island | 5.146 × 10−4 | 0.001 | −0.658 (0.001) |
| cg23370536 | TSS1500 | Island | 6.015 × 10−4 | 0.001 | −0.662 (0.001) |
| cg11935854 | 5’UTR; 1st Exon | Island | NS | NS | NS |
| cg10087973 | 1st Exon | Island | NS | NS | NS |
| cg18836493 | Body | Island | NS | NS | NS |
| cg17891820 | Body | Island | NS | NS | NS |
| cg05786661 | Body | Island | NS | NS | NS |
| cg19041419 | Body | S Shore | 3.866 × 10−4 | 0.015 | −0.816 (<0.001) |
| cg01819568 | Body | S Shelf | 0.019 | 0.025 | −0.689 (0.001) |
| cg06581979 | Body | S Shelf | 1.072 × 10−6 | 0.002 | −0.809 (<0.000) |
| cg24867302 | Body | 0.015 | 0.011 | −0.643 (0.002) | |
| cg02883198 | Body | 0.005 | 0.028 | −0.591 (0.005) | |
| cg19823803 | Body | 0.006 | 0.044 | −0.425 (0.055) | |
| cg23487812 | Body | 1.685 × 10−4 | 0.002 | −0.616 (0.003) | |
| cg17854066 | Body | 2.281 × 10−6 | 0.001 | −0.681 (0.001) | |
| cg21813871 | Body | 2.908 × 10−5 | 0.001 | −0.727 (<0.000) | |
| cg07156814 | Body | 0.046 | 0.034 | −0.532 (0.013) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horvathova Kajabova, V.; Soltysova, A.; Demkova, L.; Plesnikova, P.; Lyskova, D.; Furdova, A.; Babal, P.; Smolkova, B. KIT Expression Is Regulated by DNA Methylation in Uveal Melanoma Tumors. Int. J. Mol. Sci. 2021, 22, 10748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910748
Horvathova Kajabova V, Soltysova A, Demkova L, Plesnikova P, Lyskova D, Furdova A, Babal P, Smolkova B. KIT Expression Is Regulated by DNA Methylation in Uveal Melanoma Tumors. International Journal of Molecular Sciences. 2021; 22(19):10748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910748
Chicago/Turabian StyleHorvathova Kajabova, Viera, Andrea Soltysova, Lucia Demkova, Paulina Plesnikova, Darina Lyskova, Alena Furdova, Pavel Babal, and Bozena Smolkova. 2021. "KIT Expression Is Regulated by DNA Methylation in Uveal Melanoma Tumors" International Journal of Molecular Sciences 22, no. 19: 10748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221910748