SMA-10 Is a Non-Canonical Member of the TGF-β Sma/Mab Pathway and Immunity Regulator via the DAF-2 Insulin Receptor in Caenorhabditis elegans

 and
and
Abstract
:1. Introduction
2. Results
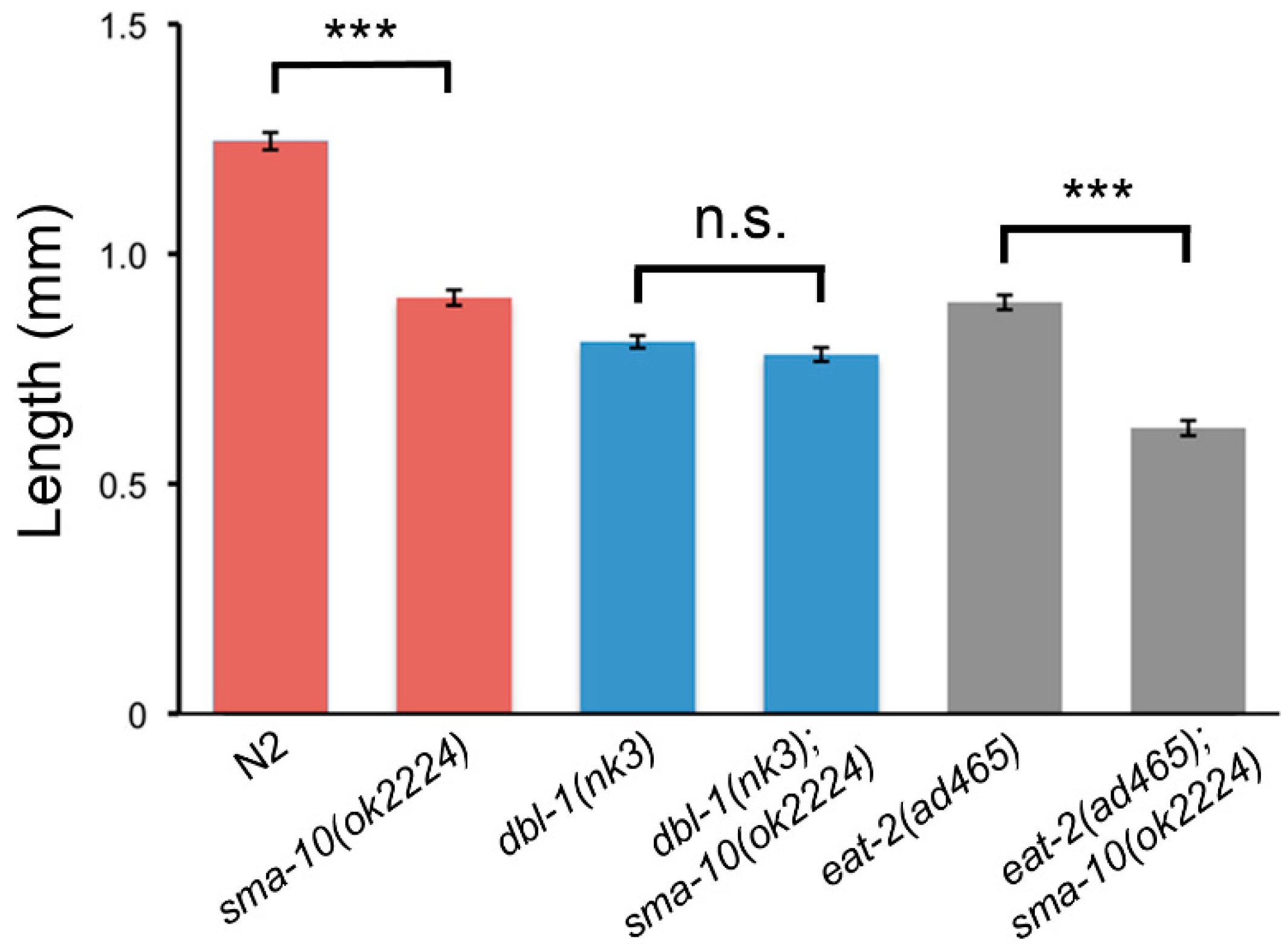
2.1. C. elegans SMA-10 Regulates Growth through Transforming Growth Factor β (TGF-β) Small/Male Abnormal (Sma/Mab) via Endoreduplication
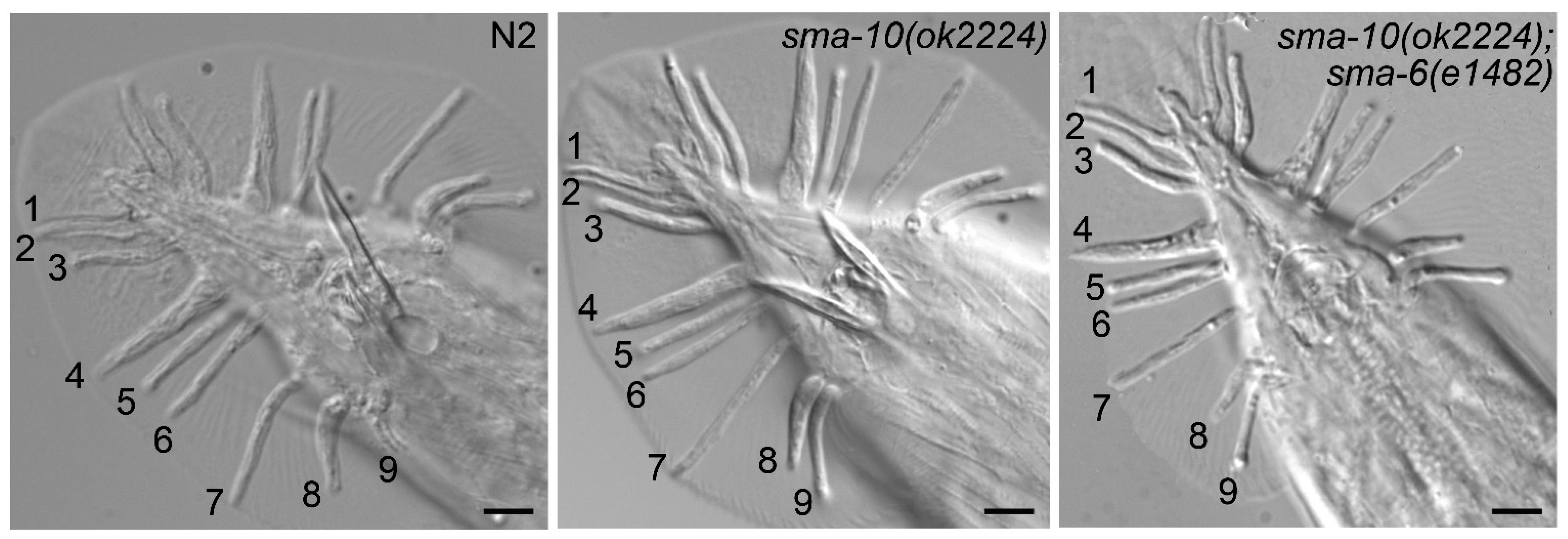
2.2. SMA-10 Is Dispensable for Male Tail Formation
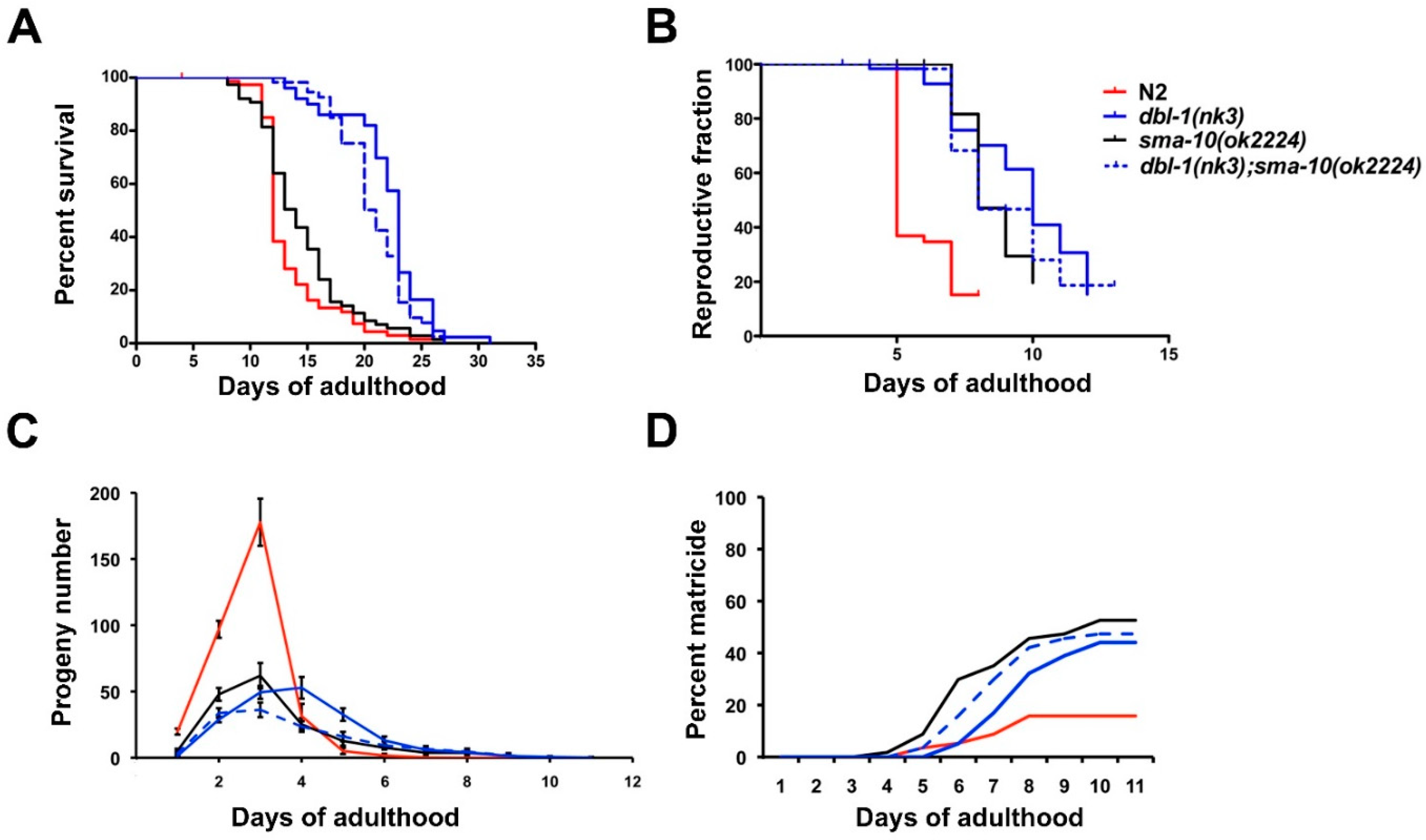
2.3. Longevity Is Not Affected by SMA-10 Expression
2.4. Reproductive Ageing Is Regulated by SMA-10
2.5. Matricide Rate Is Similar in sma-10 Defective Worms Than in dbl-1 Mutants
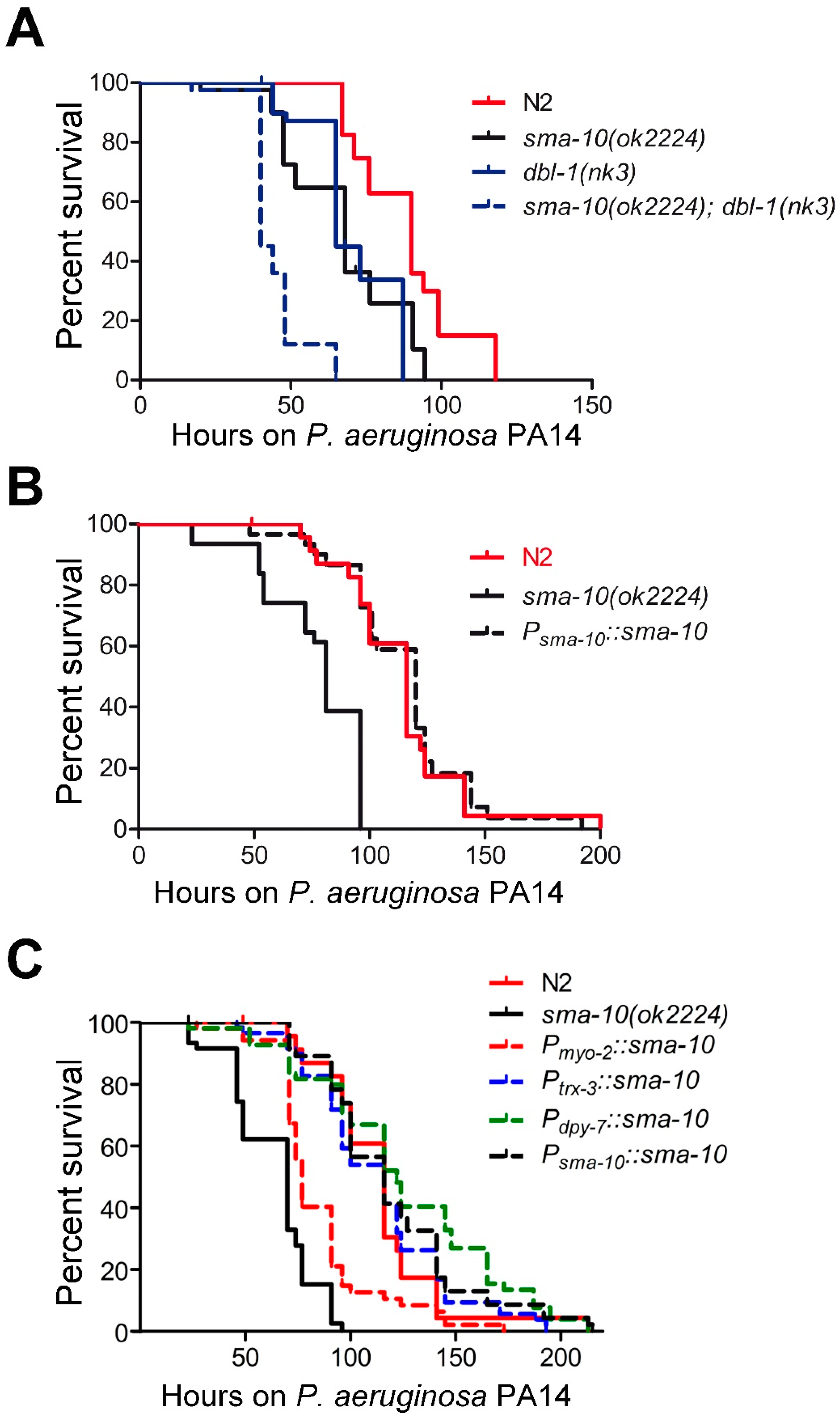
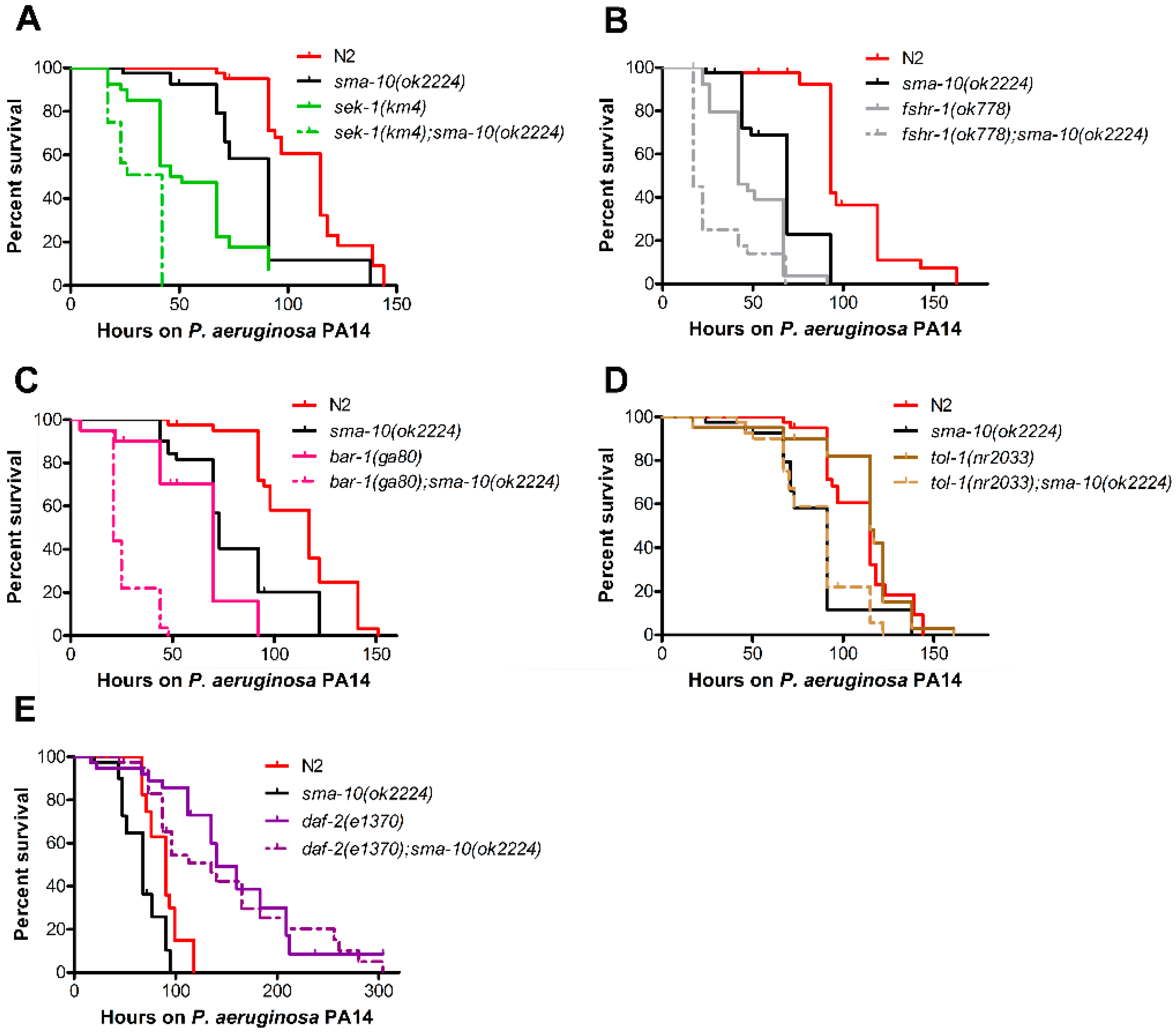
2.6. SMA-10 Plays a Role in Immunity Independently of TGF-β Sma/Mab Pathway
2.6.1. SMA-10 Functions in Hypodermis and Intestine to Regulate Immunity
2.6.2. Pathogenic Defence via SMA-10 Runs in Parallel to Various Immunity Pathways
2.6.3. SMA-10 Modulates Immunity via DAF-2 Insulin Receptor
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Generation of Transgenic Strains for Tissue Specific SMA-10 Expression
4.3. Morphometrics
4.4. Hypodermal Ploidy
4.5. Lifespan Analysis
4.6. Reproductive Span
4.7. Progeny Production
4.8. Matricide
4.9. Pathogenic Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TGF-β | Transforming growth factor beta |
| IIS | Insulin/insulin-like growth factor signalling |
| LRIG | Leucine-rich repeats and immunoglobulin-like domains |
| NGM | Nematode growth medium |
References
- Massagué, J. TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Gumienny, T.L.; Savage-Dunn, C. TGF-β signaling in C. elegans. WormBook 2013, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Savage-Dunn, C.; Padgett, R.W. The TGF-β Family in Caenorhabditis elegans. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, C.; Sen, D.; Shi, H.; Foehr, M.L.; Plavskin, Y.; Vatamaniuk, O.K.; Liu, J. The RGM protein DRAG-1 positively regulates a BMP-like signaling pathway in Caenorhabditis elegans. Development 2010, 137, 2375–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumienny, T.L.; Macneil, L.; Zimmerman, C.M.; Wang, H.; Chin, L.; Wrana, J.L.; Padgett, R.W. Caenorhabditis elegans SMA-10/LRIG is a conserved transmembrane protein that enhances bone morphogenetic protein signaling. PLoS Genet. 2010, 6, e1000963. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Shi, H.; Xiong, S.; Hu, F.; Xiong, W.-C.; Liu, J. The neogenin/DCC homolog UNC-40 promotes BMP signaling via the RGM protein DRAG-1 in C. elegans. Development 2013, 140, 4070–4080. [Google Scholar] [CrossRef] [Green Version]
- Savage-Dunn, C.; Maduzia, L.L.; Zimmerman, C.M.; Roberts, A.F.; Cohen, S.; Tokarz, R.; Padgett, R.W. Genetic screen for small body size mutants in C. elegans reveals many TGF-β pathway components. Genesis 2003, 35, 239–247. [Google Scholar] [CrossRef]
- Dolan, J.; Walshe, K.; Alsbury, S.; Hokamp, K.; O’Keeffe, S.; Okafuji, T.; Miller, S.F.C.; Tear, G.; Mitchell, K.J. The extracellular leucine-rich repeat superfamily; a comparative survey and analysis of evolutionary relationships and expression patterns. BMC Genomics 2007, 8, 320. [Google Scholar] [CrossRef] [Green Version]
- Simion, C.; Cedano-Prieto, M.E.; Sweeney, C. The LRIG family: Enigmatic regulators of growth factor receptor signaling. Endocr. Relat. Cancer 2014, 21, R431–R443. [Google Scholar] [CrossRef] [Green Version]
- Gleason, R.J.; Vora, M.; Li, Y.; Kane, N.S.; Liao, K.; Padgett, R.W. C. elegans SMA-10 regulates BMP receptor trafficking. PLoS ONE 2017, 12, e0180681. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.T.; Hu, P.J. Insulin/insulin-like growth factor signaling in C. elegans. WormBook 2013, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, T.A.; Haridas, H.; Duffy, J.B. Kekkon5 is an extracellular regulator of BMP signaling. Dev. Biol. 2009, 326, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raizen, D.M.; Lee, R.Y.; Avery, L. Interacting genes required for pharyngeal excitation by motor neuron MC in Caenorhabditis elegans. Genetics 1995, 141, 1365–1382. [Google Scholar]
- Lozano, E.; Sáez, A.G.; Flemming, A.J.; Cunha, A.; Leroi, A.M. Regulation of growth by ploidy in Caenorhabditis elegans. Curr. Biol. 2006, 16, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.; Maduzia, L.L.; Padgett, R.W. Specificity of TGF-β signaling is conferred by distinct type I receptors and their associated SMAD proteins in Caenorhabditis elegans. Development 1999, 126, 251–260. [Google Scholar]
- Luo, S.; Shaw, W.M.; Ashraf, J.; Murphy, C.T. TGF-β Sma/Mab signaling mutations uncouple reproductive aging from somatic aging. PLoS Genet. 2009, 5, e1000789. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Kleemann, G.A.; Ashraf, J.M.; Shaw, W.M.; Murphy, C.T. TGF-β and insulin signaling regulate reproductive aging via oocyte and germline quality maintenance. Cell 2010, 143, 299–312. [Google Scholar] [CrossRef] [Green Version]
- Mallo, G.V.; Kurz, C.L.; Couillault, C.; Pujol, N.; Granjeaud, S.; Kohara, Y.; Ewbank, J.J. Inducible antibacterial defense system in C. elegans. Curr. Biol. 2002, 12, 1209–1214. [Google Scholar] [CrossRef] [Green Version]
- Okkema, P.G.; Harrison, S.W.; Plunger, V.; Aryana, A.; Fire, A. Sequence requirements for myosin gene expression and regulation in Caenorhabditis elegans. Genetics 1993, 135, 385–404. [Google Scholar]
- Gilleard, J.S.; Barry, J.D.; Johnstone, I.L. cis regulatory requirements for hypodermal cell-specific expression of the Caenorhabditis elegans cuticle collagen gene dpy-7. Mol. Cell. Biol. 1997, 17, 2301–2311. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Hidalgo, M.; Kurz, C.L.; Pedrajas, J.R.; Naranjo-Galindo, F.J.; González-Barrios, M.; Cabello, J.; Sáez, A.G.; Lozano, E.; Button, E.L.; Veal, E.A.; et al. Functional characterization of thioredoxin 3 (TRX-3), a Caenorhabditis elegans intestine-specific thioredoxin. Free Radic. Biol. Med. 2014, 68, 205–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.-W.; et al. A Conserved p38 MAP Kinase Pathway in Caenorhabditis elegans Innate Immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irazoqui, J.E.; Ng, A.; Xavier, R.J.; Ausubel, F.M. Role for beta-catenin and HOX transcription factors in Caenorhabditis elegans and mammalian host epithelial-pathogen interactions. Proc. Natl. Acad. Sci. USA 2008, 105, 17469–17474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.R.; Kim, D.H.; Ausubel, F.M. The G protein-coupled receptor FSHR-1 is required for the Caenorhabditis elegans innate immune response. Proc. Natl. Acad. Sci. USA 2009, 106, 2782–2787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenor, J.L.; Aballay, A. A conserved Toll-like receptor is required for Caenorhabditis elegans innate immunity. EMBO Rep. 2008, 9, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Pujol, N.; Link, E.M.; Liu, L.X.; Kurz, C.L.; Alloing, G.; Tan, M.W.; Ray, K.P.; Solari, R.; Johnson, C.D.; Ewbank, J.J. A reverse genetic analysis of components of the Toll signaling pathway in Caenorhabditis elegans. Curr. Biol. 2001, 11, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Garsin, D.A.; Villanueva, J.M.; Begun, J.; Kim, D.H.; Sifri, C.D.; Calderwood, S.B.; Ruvkun, G.; Ausubel, F.M. Long-lived C. elegans daf-2 mutants are resistant to bacterial pathogens. Science 2003, 300, 1921. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Shi, H.; Szymczak, L.C.; Aydin, T.; Yun, S.; Constas, K.; Schaeffer, A.; Ranjan, S.; Kubba, S.; Alam, E.; et al. Promotion of bone morphogenetic protein signaling by tetraspanins and glycosphingolipids. PLoS Genet. 2015, 11, e1005221. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, Z.; Shi, H.; Liu, J. Two Paralogous Tetraspanins TSP-12 and TSP-14 Function with the ADAM10 Metalloprotease SUP-17 to Promote BMP Signaling in Caenorhabditis elegans. PLoS Genet. 2017, 13, e1006568. [Google Scholar] [CrossRef] [Green Version]
- DeGroot, M.S.; Shi, H.; Eastman, A.; McKillop, A.N.; Liu, J. The Caenorhabditis elegans SMOC-1 Protein Acts Cell Nonautonomously To Promote Bone Morphogenetic Protein Signaling. Genetics 2019, 211, 683–702. [Google Scholar] [CrossRef] [Green Version]
- Zugasti, O.; Ewbank, J.J. Neuroimmune regulation of antimicrobial peptide expression by a noncanonical TGF-β signaling pathway in Caenorhabditis elegans epidermis. Nat. Immunol. 2009, 10, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hunt-Newbury, R.; Viveiros, R.; Johnsen, R.; Mah, A.; Anastas, D.; Fang, L.; Halfnight, E.; Lee, D.; Lin, J.; Lorch, A.; et al. High-throughput in vivo analysis of gene expression in Caenorhabditis elegans. PLoS Biol. 2007, 5, e237. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.D.; Riddle, D.L.; Ruvkun, G. The C. elegans DAF-2 insulin-like receptor is abundantly expressed in the nervous system and regulated by nutritional status. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Kaletsky, R.; Yao, V.; Williams, A.; Runnels, A.M.; Tadych, A.; Zhou, S.; Troyanskaya, O.G.; Murphy, C.T. Transcriptome analysis of adult Caenorhabditis elegans cells reveals tissue-specific gene and isoform expression. PLoS Genet. 2018, 14, e1007559. [Google Scholar] [CrossRef] [Green Version]
- Rafidi, H.; Mercado, F.; Astudillo, M.; Fry, W.H.D.; Saldana, M.; Carraway, K.L.; Sweeney, C. Leucine-rich repeat and immunoglobulin domain-containing protein-1 (Lrig1) negative regulatory action toward ErbB receptor tyrosine kinases is opposed by leucine-rich repeat and immunoglobulin domain-containing protein 3 (Lrig3). J. Biol. Chem. 2013, 288, 21593–21605. [Google Scholar] [CrossRef] [Green Version]
- Shaw, W.M.; Luo, S.; Landis, J.; Ashraf, J.; Murphy, C.T. The C. elegans TGF-β Dauer pathway regulates longevity via insulin signaling. Curr. Biol. 2007, 17, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.D.; Yen, K.; Bansal, A.; Kwon, E.-S.; Padmanabhan, S.; Tissenbaum, H.A. PDP-1 links the TGF-β and IIS pathways to regulate longevity, development, and metabolism. PLoS Genet. 2011, 7, e1001377. [Google Scholar] [CrossRef] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mello, C.; Fire, A. DNA transformation. Methods Cell Biol. 1995, 48, 451–482. [Google Scholar]
- Zwaal, R.R.; Van Baelen, K.; Groenen, J.T.; van Geel, A.; Rottiers, V.; Kaletta, T.; Dode, L.; Raeymaekers, L.; Wuytack, F.; Bogaert, T. The sarco-endoplasmic reticulum Ca2+ ATPase is required for development and muscle function in Caenorhabditis elegans. J. Biol. Chem. 2001, 276, 43557–43563. [Google Scholar] [CrossRef] [Green Version]
- Sulston, J.; Hodgkin, J. Methods. In The Nematode Caenorhabditis elegans; Wood, W.B., Ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1988; pp. 587–606. [Google Scholar]
- Hedgecock, E.M.; White, J.G. Polyploid tissues in the nematode Caenorhabditis elegans. Dev. Biol. 1985, 107, 128–133. [Google Scholar] [CrossRef]
- Tain, L.S.; Lozano, E.; Sáez, A.G.; Leroi, A.M. Dietary regulation of hypodermal polyploidization in C. elegans. BMC Dev. Biol. 2008, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.W.; Mahajan-Miklos, S.; Ausubel, F.M. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.R.; Ausubel, F.M. Models of Caenorhabditis elegans infection by bacterial and fungal pathogens. Methods Mol. Biol. 2008, 415, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.A.; Kawli, T.; Tan, M.-W. Pseudomonas aeruginosa suppresses host immunity by activating the DAF-2 insulin-like signaling pathway in Caenorhabditis elegans. PLoS Pathog. 2008, 4, e1000175. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Hypodermal Ploidy | N | p Value |
|---|---|---|---|
| N2 | 10.86 ± 0.8 | 12 | |
| sma-10(ok2224) | 8.77 ± 0.7 | 14 | 0.0008 a |
| dbl-1(nk3) | 7.04 ± 0.3 | 17 | <0.0001 a, 0.0005 b |
| sma-10(ok2224);dbl-1(nk3) | 7.77 ± 0.6 | 12 | <0.0001 a, 0.067 b, 0.065 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lucas, M.P.; Jiménez, M.; Sánchez-Pavón, P.; Sáez, A.G.; Lozano, E. SMA-10 Is a Non-Canonical Member of the TGF-β Sma/Mab Pathway and Immunity Regulator via the DAF-2 Insulin Receptor in Caenorhabditis elegans. Int. J. Mol. Sci. 2021, 22, 638. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020638
de Lucas MP, Jiménez M, Sánchez-Pavón P, Sáez AG, Lozano E. SMA-10 Is a Non-Canonical Member of the TGF-β Sma/Mab Pathway and Immunity Regulator via the DAF-2 Insulin Receptor in Caenorhabditis elegans. International Journal of Molecular Sciences. 2021; 22(2):638. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020638
Chicago/Turabian Stylede Lucas, María Pilar, Marta Jiménez, Paloma Sánchez-Pavón, Alberto G. Sáez, and Encarnación Lozano. 2021. "SMA-10 Is a Non-Canonical Member of the TGF-β Sma/Mab Pathway and Immunity Regulator via the DAF-2 Insulin Receptor in Caenorhabditis elegans" International Journal of Molecular Sciences 22, no. 2: 638. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020638