Biomineralization of Collagen-Based Materials for Hard Tissue Repair



1
Department of Chemical and Biomolecular Engineering, Ohio University, Athens, OH 45701, USA
2
Department of Mechanical Engineering, Ohio University, Athens, OH 45701, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(2), 944; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020944
Submission received: 25 December 2020
/
Revised: 15 January 2021
/
Accepted: 16 January 2021
/
Published: 19 January 2021
(This article belongs to the Special Issue Novel Extracellular and Intracellular Signalling Mechanisms in Bone)
Abstract
:Hydroxyapatite (HA) reinforced collagen fibrils serve as the basic building blocks of natural bone and dentin. Mineralization of collagen fibrils play an essential role in ensuring the structural and mechanical functionalities of hard tissues such as bone and dentin. Biomineralization of collagen can be divided into intrafibrillar and extrafibrillar mineralization in terms of HA distribution relative to collagen fibrils. Intrafibrillar mineralization is termed when HA minerals are incorporated within the gap zone of collagen fibrils, while extrafibrillar mineralization refers to the minerals that are formed on the surface of collagen fibrils. However, the mechanisms resulting in these two types of mineralization still remain debatable. In this review, the evolution of both classical and non-classical biomineralization theories is summarized. Different intrafibrillar mineralization mechanisms, including polymer induced liquid precursor (PILP), capillary action, electrostatic attraction, size exclusion, Gibbs-Donnan equilibrium, and interfacial energy guided theories, are discussed. Exemplary strategies to induce biomimetic intrafibrillar mineralization using non-collagenous proteins (NCPs), polymer analogs, small molecules, and fluidic shear stress are discussed, and recent applications of mineralized collagen fibers for bone regeneration and dentin repair are included. Finally, conclusions are drawn on these proposed mechanisms, and the future trend of collagen-based materials for bone regeneration and tooth repair is speculated.
1. Introduction
Natural bone is an organic and inorganic composite constituting of mineralized collagen fibrils, which serves as the second level of its seven hierarchical levels. The inner dentin-pulp layer of human tooth has a similar mineralization profile to native bone [1]. The main inorganic component in these tissues is nanosized hydroxyapatite (HA; Ca10(PO4)6(OH)2). Its crystal structure ascribes to the hexagonal crystal system, which enables it with structural and compositional flexibility and stability. It allows for an abundant variety of ion substitutions involving divalent or trivalent cations for the Ca2+ sites while anions for the phosphate and hydroxyl sites. For examples, carbonate-substituted (carbonated) hydroxyapatite forms the basic mineral component of hard tissues in vertebrates [2]; fluoride-substituted hydroxyapatite (fluorapatite) is found in the teeth of sharks and other fishes [3]. It is a common strategy to use transition metals, such as Mn, Fe, Co and Ni, to substitute Ca(I) (surrounded by phosphate), and Ca(II) (surrounded by hydroxyl) in the HA lattice to grant materials with magnetic properties to be used in many biomedical applications, such as drug delivery, diagnostic testing, and magnetic resonance imaging [4,5,6].
The main organic component in bone and dentin is type I collagen, which has a complex structure including four levels of hierarchy. The primary level is amino acid triplets consisting mostly of proline (Pro), hydroxyproline (Hyp), and glycyl (Gly), following the pattern of Gly-Pro-X or Gly-X-Hyp, where X can be any of other amino acid species [7]. The amino acid triplets stack repeatedly to form its secondary level of structure [8]. The tertiary level is the well-known triple helix consisting of three interconnected α-chains [9]. The last hierarchy is in supermolecular level that forms collagen fibrils or fibers [10]. These collagen fibrils then continue to self-assemble both linearly and laterally into a hydrogel network under suitable physicochemical conditions [11]. As a naturally extracted polymer from extracellular matrix (ECM), collagen-based materials demonstrate excellent biocompatibility, which supports cellular adhesion, proliferation, migration and differentiation. As a result, they have been widely used in many biomedical applications, such as drug delivery, tissue engineering, wound healing, and cosmetic surgery [8,12,13].
Bone tissue engineering aims to repair or regenerate native bones in defective areas that are beyond their self-healing capabilities with the aid of grafting materials, typically scaffolds, progenitor cells and/or growth factors. Thus, selection of scaffolds becomes the key to the success of bone tissue engineering [14]. First, the material should be biocompatible and not induce any adverse effects or toxicity to the host. Second, it should be biodegradable to avoid secondary surgery after healing. Third, it should have interconnected pores to support cell adhesion, vascularization and transportation of nutrients and metabolic wastes. Finally, it should possess suitable mechanical properties to against various stresses and maintain structural integrity [8,15]. Collagen-hydroxyapatite-based (Col-HA-based) biomimetic scaffolds that structurally and compositionally resemble the natural bone have drawn extensive attention from researchers because of their excellent biocompatibility, biodegradability, low antigenicity, and compositional and structural flexibility. In contrast to bone, dental hard tissues, such as the outmost enamel and inner dentin-pulp complex, cannot self-heal if damaged. The cause of dental caries is a dynamic process of imbalance between demineralization and remineralization [16]. The caries is typically filled with artificial materials, such as amalgam, ceramics and resin composites. However, these materials could also bring about side effects, including trauma to sound dental tissue, dentin hypersensitivity, and interface microleakage between the filling materials and dental tissue [17]. Thus, biomimetic Col-HA-based materials provide a promising alternative for accelerating remineralization of dentin for tooth defect repair at an early stage of pathological conditions [18]. Overall, biomimetic mineralization of collagen becomes an essential strategy for developing bioinspired materials for both orthopedic and dental applications.
Existing reviews about collagen-based materials have been focused on (1) the collagen sources, such as bovine tendons, rat tails, and fish, (2) the physical forms and morphologies, such as hydrogels, fibers, microspheres, and scaffolds or (3) their applications, such as tissue engineering, wound healing, and cosmetic surgery. Thus, there is a need to develop a comprehensive review on these mechanisms, especially to include the most recent progress of biomineralization mechanisms. In this review, both intrafibrillar and extrafibrillar mineralization mechanisms are discussed and strategies for biomimetic mineralization of collagen-based materials are reviewed, and their applications in bone tissue engineering and tooth remineralization are covered. The future trend of mineralized collagen-based material in tissue engineering and dentin repair is speculated.
2. Biomineralization of Collagen
Ordered biomineralization holds the basis of structural organization and functionality for multiple organisms. In comparison, pathological mineralization leads to the formation of undesired biominerals, such as kidney stones and dental calculus [19]. It has been recently reported that deficiency of pyrophosphate, a known endogenous biomineralization inhibitor, could result in abnormal calcification process such as ectopic vascular calcification in the form of hydroxyapatite crystals and thus increase the risk of cardiovascular disease [20,21,22]. Developing an in-depth understanding of biomineralization mechanism(s) is therefore imperative for promoting the desired mineralization while minimizing the undesired ones. It will also shed light on synthetic chemistry to develop bioinspired materials via biomimetic mineralization. As shown in Figure 1, the biomineralization of type I collagen, in terms of the distribution patterns of hydroxyapatite in collagen, can be distinguished into intrafibrillar mineralization, where HA exhibits within the gap zone of collagen fibrils, and extrafibrillar mineralization when HA deposits on the surface of collagen fibrils. However, the mechanisms driving these processes remain unclear and debatable [19,23].
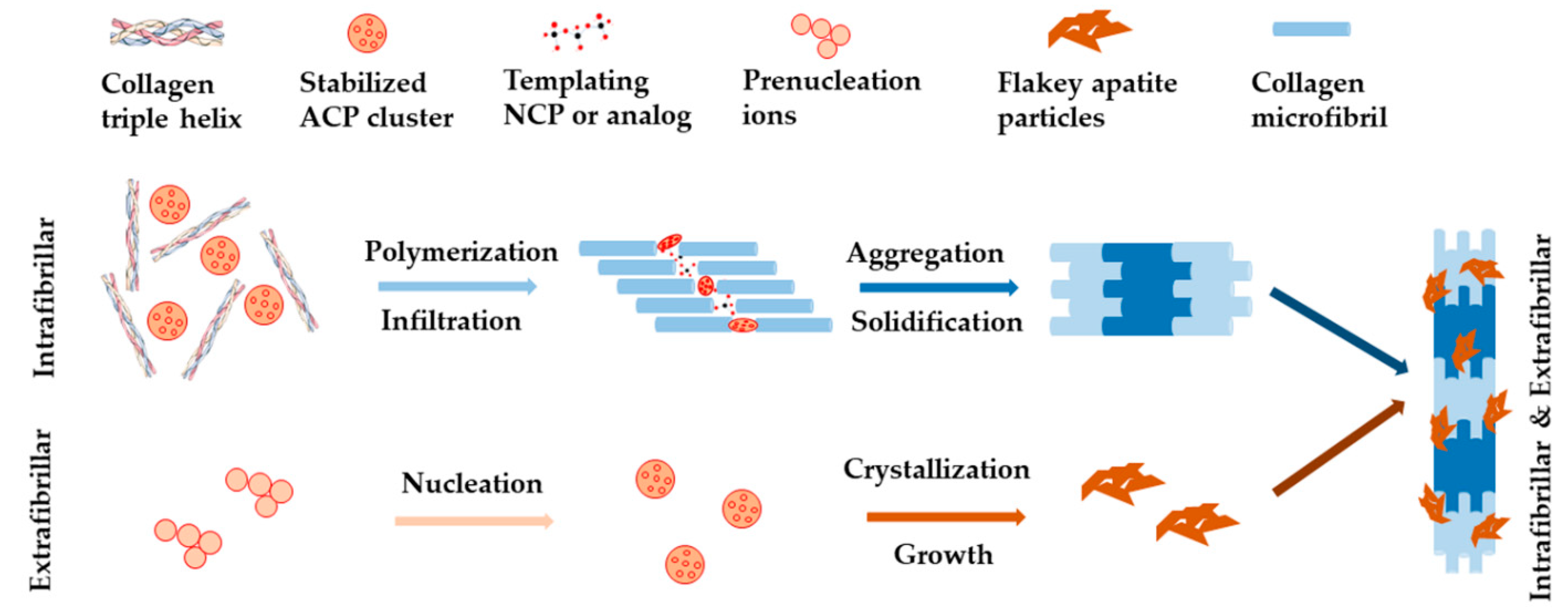
The classical view of biomineralization relies on crystallization pathway, which describes the movement of ions from their original source into targeted tissue forming mineralized bone and dentin. In this process, cells (or bacteria) are believed to play essential roles, by temporarily concentrating inorganic ions in intracellular membrane-bound vesicles in the form of a highly disordered solid phase. The solid phase is then destabilized and crystallized into minerals when it is transported into the final mineralization position [25]. Challenging this, a newer standpoint of biomineralization came from the non-classical theory, which was built upon the discovery of prenucleation clusters in the process of calcium carbonate-based biomineralization [26]. For example, it was proposed that the regulation of calcium phosphate (CaP) phases exhibited two major processes, namely the sequestration of calcium and phosphorous ions into fluidic amorphous calcium phosphate (ACP) precursors, and the templating of these clusters into ordered forms [27,28]. Both processes were believed to be well regulated by extracellular non-collagenous proteins (NCPs) and other factors such as biomolecules [29,30]. As shown in Figure 2, with these regulations, HA nanocrystals deposited within the gap zone of collagen fibrils, forming intrafibrillar mineralization, at an early stage of the biomineralization process. Then they precipitated on the surface of collagen fibrils, forming extrafibrillar mineralization, at a later stage of biomineralization. These processes led to collagen fibrils with both intrafibrillar and extrafibrillar mineralization [24,31,32]. The mineralized fibrils further bundled into highly ordered and stacked structure, forming either compact or spongy bone [33].
The role that collagen plays during biomineralization process has also been advanced with the development of better understanding of the system. Collagen was long believed to only serve as a structural matrix and considered to be inactive in biomineralization. At the end of last decade, collagen was discovered to play an active role in the apatite mineralization process. This was supported by the discovery of a positively charged region existing in collagen fibrils at the interface of the gap and overlap zones [19,31]. By systematically analyzing the primary amino acid sequence of human type I collagen, it was revealed that collagen is a biomacromolecule enriched with multiple charged amino acids [34]. Therefore, the existence of positively charged domains in collagen fibrils provides binding sites for templating NCPs, which further contributes to the regulation of mineral deposition in an ordered manner. In a recent endeavor of explaining the role of collagen molecules in mineral nucleation, a simulation study using all-atom Hamiltonian replica exchange molecular dynamics has been conducted [35] The study showed that the charged amino acid side chains in collagen molecules are oriented toward the fibril gap zones and played a significant role in templating the nucleation of ACP phase. Thus, at the atomic level, calcium phosphate was verified to nucleate primarily in these regions. Despite substantial merits and significance of these findings and proposed theories, most of these studies were conducted using simulations or simplified in vitro models, where more complicated in vivo biological environment was not taken into consideration.
3. Intrafibrillar Mineralization Mechanisms
In general, extrafibrillar mineralization of collagen takes place in the form of mineral deposition on the surface of collagen fibrils in the absence of nucleation inhibitors [36]. Based on the classical theory, apatite nucleates at suitable physicochemical conditions according to the following reactions [37]:
when the pH of solution is brought up to an appropriate range with the decomposition of bicarbonate, HA starts to nucleate, and particles slowly deposit on the surface of collagen [38]. While based on the non-classical theory, prenucleation clusters and spherical ACP are formed as intermediate products before apatite crystals deposit and grow on collagen fibrils to form extrafibrillar mineralization [39]. Thus, the extrafibrillar mineralization of collagen is a relatively clear process and has been well studied [40,41]. In recent years, extensive studies have been focused particularly on the exploration of the mechanisms of intrafibrillar mineralization of collagen, which were mostly built upon the classical and the non-classical theory.
3.1. Capillary Action
Polymer induced liquid precursor (PILP) is considered to be one of the most important transient states before mineral infiltration into collagen fibrils and crystallization during intrafibrillar mineralization. The term “PILP” was first introduced by Gower et al. [42,43] along with the discovery of liquid-liquid phase separation of amorphous calcium carbonate (ACC) in the presence of polyanionic polyaspartic acid (PASP) under optical microscopy. They proposed that during the PILP process, the charged polymer acted as a processing-directing agent, with the aid of which the conventional solution crystallization process was converted into an amorphous liquid precursor-like process [44]. Using the formation of calcium carbonate and CaP based on PILP as examples, the dynamic compositional evolution and interactions between the polymer and the amorphous precursor have been revealed recently with the aid of advanced analytical techniques, such as cryogenic transmission electron microscopy (cryo-TEM), in situ atomic force microscopy (AFM), nuclear magnetic resonance (NMR), and modeling simulations [45,46,47,48]. A comprehensive understanding of PILP theory along with its applications for bone regeneration has been reported recently [49]. The basis for the PILP theory is the liquid-liquid phase separation as liquid or liquid-like component can take flexible shape prior to solidification, which largely facilitates the infiltration of minerals into collagen fibrils to form intrafibrillar mineralization.
Upon the proposal of PILP, Gower et al. [50,51] opined subsequently that a liquid phase mineral precursor could be drawn into the gap spaces of collagen fibrils by capillary action in the PILP process before it was solidified and crystallized into minerals, forming intrafibrillar mineralized collagen fibrils that were embedded with nano crystallites. The assumption was initially made based on their observations from capillary forces acting on the phase boundaries between the liquid precursor phase and collagen fibrils in a calcium carbonate system [50,52]. With the evolution of techniques and knowledge, they subsequently confirmed their theory by duplicating similar observations via a combination analyses of transmission electron microscopy (TEM), scanning electron microscopy (SEM), confocal microscopy, and X-ray diffraction (XRD) in a CaP system that is more closely mimicking the nanostructure of natural bone [29,44,53]. Briefly, they found that PASP (processing-directing agent) incorporated with calcium ions could only penetrate into turkey tendon for around 100 μm in comparison to PASP incorporated with CaP fluidic precursor that had a penetration depth over 500 μm. With a smaller Stokes radius, PASP + Ca would be expected to penetrate deeper into the dense-packed collagen scaffolds of turkey tendon than PASP + CaP if they migrated through ion diffusion [29]. Therefore, they suggested that a capillary action might have been involved in intrafibrillar mineralization as capillary forces are expected to provide rapid and long-range transportation of a fluidic precursor compared to ion diffusion. In this process, the fluidic nature of the ACP precursors allowed them to be drawn into the nano gaps and grooves of collagen fibrils via capillary infiltration [53,54]. However, whether capillary action is involved in intrafibrillar mineralization due to PILP is still debatable as this has not yet been directly observed through experiments. Exclusion of ion diffusion does not guarantee that capillary action plays a role in the intrafibrillar mineralization process. Nevertheless, capillary action has been regarded as one of the major mechanisms to date to induce intrafibrillar mineralization of collagen, although other mechanisms may have also been involved [1,24,28,55,56,57].
3.2. Electrostatic Attraction
The phenomenon of electrostatic attraction in collagen mineralization has been recognized as early as last century [58]. Not until a decade ago that the role of collagen in bone apatite mineralization was convincingly revealed [31], had electrostatic attraction been fully accepted as one of the major mechanisms in inducing intrafibrillar mineralization of collagen fibrils. With a combination of cryo-TEM, cryogenic electron tomography and molecular modeling, Nudelman el at. [31] found that the positive net charge close to the C-terminal end of collagen molecules promotes the infiltration of ACP into collagen fibrils. Moreover, they revealed that the clusters of charged amino acids in the gap and overlap zones formed nucleation sites for apatite crystallization converted from ACP, giving a parallel array of oriented apatite crystals to collagen fibrils. At the same time, they identified the negative net charge of PASP stabilized ACP precursors. Based on these findings, they opined that, in vivo, negatively charged NCPs might not only stabilize the amorphous phase by inhibiting apatite nucleation, but also play an active role in aiding the formation of negatively charged complexes of mineral precursors that allowed the minerals to enter the collagen through electrostatic attraction [31,59]. The predominant role of collagen in apatite nucleation, growth, structure and orientation during bone mineralization was thoroughly investigated in a subsequent study reported by Wang et al. [60]. They discovered that the collagen matrix controls the size and three-dimensional distribution of apatite, further supporting the mechanisms proposed by Nudelman et al. Thereafter, electrostatic attraction has become a popular mechanism and has been accepted in many studies [61,62,63]. In some of our work, we also believe that electrostatic attraction is one of the mechanisms in inducing intrafibrillar mineralization while polyanionic analogs of NCPs are used to stabilize the amorphous precursors of minerals, though other mechanisms such as capillary action and size exclusion theories may have also played a role [24,28,64,65]. Nevertheless, the mechanism alters while polycationic analogs are involved in the reaction [66].
In recent years, there are more and more experimental observations suggesting that the collagen intrafibrillar mineralization process cannot be simply explained by electrostatic attraction. For example, it is hard to explain why site specificity exists in different parts of turkey leg tendons as some parts are heavily mineralized while some are never mineralized [67], giving that positively charged zones are distributed periodically on the surface of the entire collagen molecules. It is also incapable of explaining why electrostatic interaction occurs only at a-band of collagen molecules while both the a-band and c-band exhibit positively charged regions [68]. It has been found in a few studies that cationic polyelectrolyte, such as polyallyamine hydrochloride (PAH), could also stabilize amorphous precursors of mineral crystals and induce intrafibrillar mineralization thereafter [66,69,70]. These cationic polymers are unlikely to bind to the positive net charge of collagen fibrils through electrostatic attraction and facilitate the infiltration of amorphous precursors. A recent work conducted by Song et al. [71] indicated that the zeta potential of collagen bound with high-molecular weight PAA (450 kDa) was –17.17 ± 1.98 eV, while that of CaP precursors was –0.88 ± 0.07 eV. Apparently, attachment of these two negatively charged species cannot be simply explained by electrostatic attractions according to the basis of Coulombic interpretations.
3.3. Size Exclusion
The size exclusion characteristics in collagen mineralization was observed in 2007 by Price et al. [72]. It was derived from the theory that the collagen fibrils were formed first and then water compartment within the fibrils was replaced with minerals during mineralization. It was proposed that collagen not only provided the position for mineral deposition and growth, but also acted as a gatekeeper to screen different molecules and determine whether they could diffuse into the inner spaces of collagen fibrils. A gel filtration-like column chromatography was used in the study to investigate the size exclusion characteristic of type I collagen extracted from bovine Achilles tendon. The results demonstrated that molecules smaller than 6 kDa could diffuse into all of the water compartment within the collagen fibril, whereas molecules larger than 40 kDa were excluded from this liquid. This means protein molecules smaller than 6 kDa, such as osteocalcin (OCN; 5.8 kDa) and bone Gla protein (BGP; 5.7 kDa), can interact directly with minerals and grow within collagen fibrils, while those larger than 40 kDa, such as fetuin (48 kDa) and albumin (66 kDa), cannot [32,73]. Importantly, Price et al. [73] suggested that although large molecules could not directly penetrate into collagen fibrils, some large proteins could still facilitate collagen intrafibrillar mineralization via other mechanisms. For example, osteopontin (OPN; 60–65 kDa) and fetuin, can direct intrafibrillar mineralization by inhibiting apatite nucleation and growth everywhere but within collagen fibrils [55,74,75,76]. While some other large molecules, such as bone sialoprotein (BSP; 60–80 kDa), can generate apatite crystals outside collagen fibrils, and these small crystals then diffuse into the gap zone of collagen and grow to replace water compartment inside collagen fibrils, thereby forming intrafibrillar mineralized collagen [75,77]. This theory was termed “mineralization by inhibitor exclusion”.
In a recent work, Niu et al. [70] examined this theory in PAH-directed collagen intrafibrillar mineralization where electrostatic attraction theory was not applicable as discussed in the last section. They found that although amine-containing PAH (15 kDa) could only access to 50.5% of the intrafibrillar water, it was still able to act as a nucleation inhibitor and produce excellent intrafibrillar mineralization. Interestingly, when another amine-containing small molecule, spermine (202 Da), which can access 98.64% of intrafibrillar water, was added to PAH-containing mineralization solution, the rate of collagen intrafibrillar mineralization was significantly reduced. Hence, the rate or degree of intrafibrillar mineralization induced by NCPs is more likely dependent on the nature of NCPs, such as their ability to inhibit nucleation of CaP, rather than their size or accessibility to water compartment within collagen fibrils. This was also evidenced in one of our recent works, where carboxyl-rich brush-like polymers were used to induce collagen intrafibrillar mineralization [64]. Two large-molecular, brush-like polymers with enriched carboxyl branches, carboxylated polyethylene glycol terpolymer (PEG-COOH; 42 kDa) and polyethylene glycol/polyacrylic acid copolymer (PEG-PAA; 44 kDa), were synthesized in house. Compared to linear PAA (2 kDa), which theoretically has full access to the intrafibrillar water, both brush-like polymers have demonstrated better capability to induce intrafibrillar mineralization, which might be attributed to the high availability of carboxylic groups of these polymers [61,78,79]. These results partially revealed the limitation of size exclusion theory in intrafibrillar mineralization but also endorsed the selective permeability of collagen.
3.4. Gibbs-Donnan Equilibrium
After verifying the selective permeability of collagen in PAH-directed mineralization, Niu et al. [70] compared the effect of polycationic and polyanionic electrolyte in directing intrafibrillar mineralization. Based on experimental observations and molecular dynamic simulations, they proposed that in polyelectrolyte-directed (both polycation- and polyanion-directed) collagen mineralization systems, Gibbs-Donnan equilibrium had to be reached between the outside (extrafibrillar) and inside (intrafibrillar) spaces of collagen fibrils via simultaneously balancing electrostatic neutrality and osmotic equilibrium. Since no difference was observed while either polycationic or polyanionic electrolyte was used to induce collagen mineralization, they concluded that electrostatic attraction was not the only mechanism for inducing intrafibrillar mineralization of collagen suggesting that additional driving forces might have controlled the movement of prenucleation clusters (classical theory) or ACP (non-classical theory) toward collagen fibrils. Osmosis describes how water moves from a low concentration to a higher concentration region across a semipermeable membrane [80]. While collagen is evidenced to have selective permeability, the osmolality of PAH in mineralization solution (ACP solution) and water was compared. It was found that the osmolality of PAH-ACP is about 20-fold as high as that of PAH in water at the same concentration. Hence, due to concentration differences of osmotically-active species and salt ions (Na+, Cl–) within and outside of collagen fibrils, it is reasonable to assume that the high oncotic pressure between the intrafibrillar and extrafibrillar spaces in the presence of a selectively permeable membrane (collagen) could provide a long-ranged interaction for ACP to infiltrate into fibrillar collagen. In one of their subsequent work, Ma et al. [47] further highlighted experimentally the importance of establishing Gibbs-Donnan equilibrium in forming intrafibrillar mineralization of collagen in a PASP and PAH dual-analog-directed mineralization system.
Overall, the establishment of Gibbs-Donnan equilibrium suggested that both short-range electrostatic forces driven by charge distribution and long-range osmosis driven by osmotic pressure between the extrafibrillar and intrafibrillar water compartments are responsible for collagen mineralization. The work provided significant insight of the effect of driving forces on ACP infiltration and for the first time disclosed the crucial role of osmotic forces in collagen mineralization [75,81,82]. It opened a new pathway in explaining collagen mineralization mechanisms. Despite of the great breakthrough, only partial in vitro models and molecular simulations were employed in these studies, so there is a need to develop direct experiments to support these theories. Moreover, none of these studies have ruled out the role of capillary infiltration in intrafibrillar mineralization, which is believed to provide rapid and long-range transportation of either prenucleation clusters or fluidic amorphous precursors [50,83,84,85]. Still, the question raised in Section 3.2 regarding the site specificity of mineralization observed in turkey tendon is not answered by the Gibbs-Donnan equilibrium theory.
3.5. Interfacial Energy Guided Mineralization
Recently, Kim el at. [23] proposed an intrafibrillar mineralization mechanism directed by nucleation energy barrier. By using in situ small-angle and wide-angle X-ray scattering (SAXS and WAXS) observations and classical nucleation theory, they found that PASP could increase interfacial energy between CaP nuclei and mineralization fluid. In contrast, the confined gap region within collagen fibrils could decrease the energy barrier by reducing the reactive surface area of CaP nuclei and corresponding surface energy penalty. Therefore, it was believed that the confined gap geometry guided the two-dimensional morphology and structure of apatite and changes of nucleation pathway by reducing the total energy barrier for intrafibrillar mineralization. Their work for the first time calculated the free energy change per molecule (∆G) during apatite nucleation and the interfacial energy (α) between CaP nuclei and mineralization solution, for intrafibrillar and extrafibrillar mineralization, respectively, at different nucleation sites. Another work regarding interfacial energy guided collagen mineralization was reported by Shao et al. [86] in a citrate-aided intrafibrillar mineralization system. It was found that the adsorbed citrate molecules on collagen fibrils significantly reduced the interfacial energy between the collagen matrix and the ACP precursor via interface wetting effect, and therefore facilitating intrafibrillar mineralization of apatite at an early stage of biomineralization. Similarly, polydopamine (PDA) was utilized in their subsequent work by Qu et al. [87] to promote dentin remineralization with decreased heterogeneous nucleation energy barrier by reducing interfacial energy of collagen/ACP.
Overall, these studies further advanced our understanding on the interactions between collagen fibrils and minerals during intrafibrillar mineralization, and quantitatively emphasized the importance of reducing interfacial energy of collagen/ACP to promote heterogeneous nucleation, which could potentially enhance mineral formation within collagen fibrils. However, the proposed mechanism still has some limitations. For instance, the molecular weight of PASP that Kim et al. used was about 5 kDa, which is supposed to be completely permeable to intrafibrillar water compartment [72]. Thus, the interfacial energies at both the extrafibrillar and intrafibrillar spaces would be affected by PASP, but it was assumed that the addition of PASP only kinetically inhibited the formation of extrafibrillar mineralization. Moreover, the calculation of heterogenous nucleation barrier relied largely on the classical nucleation theory, which is still debatable from the point of view of non-classical theory [88,89].
4. Biomimetic Intrafibrillar Mineralization Strategies for Hard Tissue Repair
The high strength of mussel shells and seashells reflects how nature applies nanotechnology to strengthen load-bearing hard tissues, and biomineralization of collagen is an excellent example [27,90]. Conversely, self-assembly of collagen-based materials through biomimetic mineralization is a nanotechnology inspired by nature. It provides an effective approach to develop advanced biomaterials in tissue engineering with abilities to not only control the functions of osteoblasts and bone marrow mesenchymal cells (BMSCs), but also guide and promote new bone regeneration in defined shapes [91]. Compared to pure collagen and extrafibrillar mineralized collagen, collagen-based tissue engineering scaffolds composed of intrafibrillar mineralized fibrils demonstrated solid advantages, such as enhanced mechanical property, increased in vitro osteogenesis and in vivo bone healing [92]. Intrafibrillar mineralized collagen was also thought to promote bone regeneration via activation of the Wnt signaling pathway [93]. The acceleration of dentin remineralization through various biomimetic strategies have also been reported recently to be an effective approach for tooth defect repair [94,95,96,97]. In this section, typical strategies for biomimetic synthesis of intrafibrillar mineralized collagen-based materials are included.
4.1. NCPs
Besides HA and collagen, there are various kinds of NCPs and extracellular molecules existing in natural bone. Hence, it is reasonable to believe that biomineralization is regulated by these components. As discussed above, collagen has been suggested to be responsible for the deposition of apatite minerals even at the absence of other factors, such as NCPs [23]. However, based on most published studies, NCPs are believed to mediate vertebrate mineralization associated with collagen by influencing the formation and maintenance of the mineralized matrix [98]. OCN is the most abundant NCP in bone influencing matrix mineralization and global metabolism that exclusively produced by osteoblasts [99]. Simon el at. [100] first evidenced the importance of this protein using high resolution TEM and Fourier analysis during collagen triple helix fibril mineralization. It was found that OCN was directly bonded with octacalcium phosphate and was often seen spreading between and over collagen fibrils like pearl necklace strings or as single spherical particles. This study revealed experimentally that OCN is attached to the collagen structure and interacts with the Ca-sites on the (100) plane dominated HA platelets. Wang et al. [75] found that in the absence of OCN, rapid and random extrafibrillar mineralization of flakey CaP particles was observed mainly on the surfaces of collagen fibrils through TEM. In contrast, hydrated, spherical nanoclusters of CaP were observed on the surface of collagen fibrils when OCN was added. These nanoclusters then infiltrated into the fibrils, forming intrafibrillarly mineralized collagen with HA nanocrystals aligned within the collagen fibrils. Recently, OCN has been found to be key to the alignment of apatite crystallites during mineralization [101]. OPN is another important NCP that mediates various biological events involving the immune and the vascular systems [102]. It was used to induce collagen intrafibrillar mineralization by serving as a process-directing agent [62]. In this study, OPN was also found to promote the interaction of mouse marrow-derived osteoclasts with re-mineralized bone, contributing to both biomineralization and bone remodeling. Such prepared intrafibrillarly mineralized materials therefore have demonstrated great potential in bone tissue engineering. Meanwhile, França et al. [55] engineered intrafibrillarly mineralized scaffolds through an OPN-guided PILP process to reproduce the nanostructure of bone. The obtained scaffolds had a similar mineral/matrix ration to native bone and its compressive modulus was increased up to 15-fold compared to pure collagen without mineralization. Although other natural NCPs, such as fetuin-A, BSP, detin matrix protein 1 (DMP1) and dentin sialophosphoprotein (DSPP), were revealed to mediate intrafibrillar mineralization [88,103,104,105], limited studies have been conducted to directly use these proteins to synthesize intrafibrillarly mineralized materials for orthopedic and dental applications due to their low availability and high cost.
4.2. Polymer Analogs
To overcome the limitations of natural NCPs, synthetic polymer analogs of NCPs are usually used to replicate their functions and induce collagen intrafibrillar mineralization. The use of polymers in collagen mineralization is generally considered to be achieved through a PILP process while the intrafibrillar mineralization process can be explained in one or more different mechanisms as discussed in the above section. Table 1 summarizes some recent examples of commonly used polyanionic and polycationic polymers in directing collagen intrafibrillar mineralization and their major contributions in biomineralization, bone regeneration and tooth repair.
4.3. Small Molecules and Fluid Shear Stress
Some small molecules, such as citrate and fluoride, have been discovered to affect intrafibrillar mineralization. Citrate is widely found in biological hard tissues such as bone and is believed to be an osteopromotive factor in mediating citrate metabolism and its downstream effects on the osteogenic differentiation of BMSCs [118]. It has also been reported that the citrate content in bone is reduced in osteoporotic and aged animals [119]. As summarized in Section 3.5, the influence of citrate on biomineralization has been well studied [86]. Citrate molecules could significantly reduce the interfacial energy between the collagen matrix and the ACP precursors, thereby facilitating intrafibrillar mineralization. Experimental results indicated that only collagen fibrils containing similar level of bound citrate to natural bone (~8.2%) could reach full mineralization comparable to bone [86]. Recently, citrate has been incorporated into collagen fibrils during their fibrillogenesis to promote intrafibrillar mineralization [63]. It was found that citrate-functionalized collagen fibrils underwent extensive intrafibrillar mineralization within 12 h in m-SBF. Another small molecule, fluoride, also showed to influence the mineralization of collagen. Different concentrations of NaF were added to the mineralization solution. As the fluoride concentration increased, the crystals became larger and more rod-like, with an increasing tendency to form fluorapatite on the fibril surfaces rather than the interior, therefore forming extra- fibrillar mineralization dominated collagen fibrils [56]. However, as fluorapatite has a lower solubility and higher hardness than HA, and thus is less likely to be dissolved in aqueous or mild acidic environment, it has been widely used in enamel and dentin remineralization to reduce the chance of cavity formation [120,121]. Although enamel does not contain collagen like bone and dentin, the addition of fluoride in collagen mineralization overall showed bright future for dental repair.
Additionally, Niu and Du et al. [122,123] shared their recent findings on collagen intrafibrillar mineralization induced by fluid shear stress (FSS). FSS is considered as a predominant stimulus to bone cells and matrix, but its role in biomineralization has not been well investigated. They discovered that the application of a small amount of FSS (less than 2 Pa, especially within 1.5 Pa) has a positive influence on mineralization by enhancing collagen self-assembly and accelerating ACP formation and transition. The capability of FSS in inducing intrafibrillar mineralization is comparable to that of commonly used NCPs analogs, PAA. Moreover, under the action of templating analogs (such as TPP), periodic FSS can also promote the formation of highly oriented hierarchical intrafibrillarly mineralized collagen, giving D-banding patterns. Their work opened a new avenue in investigation of biomineralization mechanisms by introducing vascular flow in in vitro models. It also provided a novel biomimetic strategy to induce intrafibrillar mineralization without the addition of NCPs and biomolecules.
5. Conclusions and Perspectives
Natural bone and dentin are organic-inorganic composites mainly consisting of collagen and HA that are arranged in multilevel hierarchical structures from atomic- to macro-scale. Their excellent mechanical property and favorable biocompatibility are largely determined by the biomineralization of collagen fibrils. To have a better understanding on collagen biomineralization and provide insight on the design of mineralized collagen-based materials, this review summarized current collagen mineralization mechanisms and recent biomimetic strategies for bone regeneration and tooth repair. Overall, these proposed mechanisms are either derived from the classical ion-cluster-based or non-classical amorphous precursor-based theories. A complementary proposal to both the classical and non-classical theories could be a future trend in establishing new mechanisms or perfecting existing ones. In addition, in vitro models involving more complicated conditions, such as fluid flow and cell loading, that better mimic the in vivo environment could provide clearer and more realistic cognition on biomineralization. The improvement of bioinspired collagen-based materials could start from the development of a simple, reliable, reproducible, and costless collagen extraction process. Despite extensive efforts, to date, only simplified biomimetic structures have been achieved. Therefore, biomaterials better mimicking the complex hierarchical natural hard tissues may be achieved using newly developed techniques such as 3D printing in combination with biomimetic strategies. Appropriate incorporation of essential elements, drugs, or biomolecules into well-designed 3D collagen-based structure during biomimetic mineralization could also be a direction to promote their performance for bone regeneration and tooth repair.
Author Contributions
Conceptualization, L.Y. and M.W.; investigation, L.Y.; writing—original draft preparation, L.Y.; writing—review and editing, M.W.; funding acquisition, M.W. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
Wei has an ownership interest in OrteoPoniX LLC, which develops and commercializes biomaterials for orthopedic applications.
Abbreviations
| HA | Hydroxyapatite |
| Pro | Proline |
| Hyp | Hydroxyproline |
| Gly | Glycyl |
| ECM | Extracellular matrix |
| Col-HA | Collagen-hydroxyapatite |
| CaP | Calcium phosphate |
| NCPs | Non-collagenous proteins |
| ACP | Amorphous calcium phosphate |
| PILP | Polymer induced liquid precursor |
| ACC | Amorphous calcium carbonate |
| PASP | Polyaspartic acid |
| cyro-TEM | Cryogenic transmission electron microscopy |
| AFM | Atomic force microscopy |
| NMR | Nuclear magnetic resonance |
| PAA | Polyacrylic acid |
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
| XRD | X-ray diffraction |
| PAH | Polyallyamine hydrochloride |
| OCN | Osteocalcin |
| BGP | Bone Gla protein |
| OPN | Osteopontin |
| BSP | Bone sialoprotein |
| PEG-COOH | Carboxylated polyethylene glycol terpolymer |
| PEG-PAA | Polyethylene glycol/polyacrylic acid copolymer |
| SAXS | Small-angle X-ray scattering |
| WAXS | Wide-angle X-ray scattering |
| PDA | Polydopamine |
| TPP | Sodium tripolyphosphate |
| CMC | Carboxymethyl chitosan |
| m-SBF | Modified-simulated body fluid |
| Pchi | Phosphorylated chitosan |
| PAMAM | Polyamidoamine dendrimer |
| CS | Chondroitin sulfate |
| 3D | Three-dimensional |
| FSS | Fluid shear stress |
References
- Thrivikraman, G.; Athirasala, A.; Gordon, R.; Zhang, L.; Bergan, R.; Keene, D.R.; Jones, J.M.; Xie, H.; Chen, Z.; Tao, J. Rapid fabrication of vascularized and innervated cell-laden bone models with biomimetic intrafibrillar collagen mineralization. Nat. Commun. 2019, 10, 3520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R.; Brown, W. Biological Mineralization and Demineralization; Springer: Berlin/Heidelberg, Germany, 1982. [Google Scholar]
- Chang, H.-H.; Chien, M.-J.; Kao, C.-C.; Chao, Y.-J.; Yu, P.-T.; Chang, C.-Y.; Huang, S.-J.; Lee, Y.-L.; Chan, J.C. Structural characterization of fluoride species in shark teeth. Chem. Commun. 2017, 53, 3838–3841. [Google Scholar] [CrossRef] [PubMed]
- Zilm, M.E.; Yu, L.; Hines, W.A.; Wei, M. Magnetic properties and cytocompatibility of transition-metal-incorporated hydroxyapatite. Mater. Sci. Eng. C 2018, 87, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-J.; Dong, L.; Lu, Y.; Zhang, L.-C.; An, D.; Gao, H.-L.; Yang, D.-M.; Hu, W.; Sui, C.; Xu, W.-P. Magnetic hydroxyapatite nanoworms for magnetic resonance diagnosis of acute hepatic injury. Nanoscale 2016, 8, 1684–1690. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Manivasagan, P.; Bharathiraja, S.; Moorthy, M.S.; Kim, H.H.; Seo, H.; Lee, K.D.; Oh, J. Magnetic hydroxyapatite: A promising multifunctional platform for nanomedicine application. Int. J. Nanomed. 2017, 12, 8389. [Google Scholar] [CrossRef] [Green Version]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.; Zhang, D.; Macedo, M.H.; Cui, W.; Sarmento, B.; Shen, G. Advanced Collagen-Based Biomaterials for Regenerative Biomedicine. Adv. Funct. Mater. 2019, 29, 1804943. [Google Scholar] [CrossRef]
- Ottani, V.; Martini, D.; Franchi, M.; Ruggeri, A.; Raspanti, M. Hierarchical structures in fibrillar collagens. Micron 2002, 33, 587–596. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Gentile, P.; Chiono, V.; Ciardelli, G. Collagen for bone tissue regeneration. Acta Biomater. 2012, 8, 3191–3200. [Google Scholar] [CrossRef] [PubMed]
- O’leary, L.E.; Fallas, J.A.; Bakota, E.L.; Kang, M.K.; Hartgerink, J.D. Multi-hierarchical self-assembly of a collagen mimetic peptide from triple helix to nanofibre and hydrogel. Nat. Chem. 2011, 3, 821. [Google Scholar] [CrossRef]
- Lutolf, M.P.; Gilbert, P.M.; Blau, H.M. Designing materials to direct stem-cell fate. Nature 2009, 462, 433–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, T.K.; Han, S.-H.; Han, J. Protective effects of biodegradable collagen implants on thinned sclera after strabismus surgery: A paired-eye study. J. Am. Assoc. Pediatric Ophthalmol. Strabismus 2017, 21, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, A.; Liverani, L.; Rainer, A.; Salvatore, G.; Trombetta, M.; Denaro, V. Electrospun scaffolds for bone tissue engineering. Musculoskelet. Surg. 2011, 95, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, M.; Kim, G.H. 3D-printed biomimetic scaffold simulating microfibril muscle structure. Adv. Funct. Mater. 2018, 28, 1800405. [Google Scholar] [CrossRef]
- Featherstone, J.D. Dental caries: A dynamic disease process. Aust. Dent. J. 2008, 53, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Li, Q.-L.; Cao, Y.; Fang, H.; Xia, R.; Zhang, Z.-H. In vivo remineralization of dentin using an agarose hydrogel biomimetic mineralization system. Sci. Rep. 2017, 7, 41955. [Google Scholar] [CrossRef] [Green Version]
- Habibah, T.U.; Salisbury, H.G. StatPearls [Internet]; StatPearls Publishing: Petersburg, FL, USA, 2019. [Google Scholar]
- Cölfen, H. A crystal-clear view. Nat. Mater. 2010, 9, 960–961. [Google Scholar] [CrossRef]
- Azpiazu, D.; Gonzalo, S.; González-Parra, E.; Egido, J.; Villa-Bellosta, R. Role of pyrophosphate in vascular calcification in chronic kidney disease. Nefrologia 2018, 38, 250–257. [Google Scholar] [CrossRef]
- Bhadada, S.K.; Rao, S.D. Role of phosphate in biomineralization. Calcif. Tissue Int. 2020, 108, 32–40. [Google Scholar] [CrossRef]
- Ziegler, S.G.; Ferreira, C.R.; MacFarlane, E.G.; Riddle, R.C.; Tomlinson, R.E.; Chew, E.Y.; Martin, L.; Ma, C.-T.; Sergienko, E.; Pinkerton, A.B. Ectopic calcification in pseudoxanthoma elasticum responds to inhibition of tissue-nonspecific alkaline phosphatase. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Lee, B.; Thomopoulos, S.; Jun, Y.-S. The role of confined collagen geometry in decreasing nucleation energy barriers to intrafibrillar mineralization. Nat. Commun. 2018, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zilm, M.; Wei, M. Fabrication of intrafibrillar and extrafibrillar mineralized collagen/apatite scaffolds with a hierarchical structure. J. Biomed. Mater. Res. Part A 2016, 104, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S.; Addadi, L. Crystallization pathways in biomineralization. Annu. Rev. Mater. Res. 2011, 41, 21–40. [Google Scholar] [CrossRef]
- Gebauer, D.; Völkel, A.; Cölfen, H. Stable prenucleation calcium carbonate clusters. Science 2008, 322, 1819–1822. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, N.; Qi, Y.p.; Dai, L.; Bryan, T.E.; Mao, J.; Pashley, D.H.; Tay, F.R. Intrafibrillar collagen mineralization produced by biomimetic hierarchical nanoapatite assembly. Adv. Mater. 2011, 23, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Zhang, L.; Wei, M. Development of biomimetic scaffolds with both intrafibrillar and extrafibrillar mineralization. ACS Biomater. Sci. Eng. 2015, 1, 669–676. [Google Scholar] [CrossRef]
- Jee, S.-S.; Thula, T.T.; Gower, L.B. Development of bone-like composites via the polymer-induced liquid-precursor (PILP) process. Part 1: Influence of polymer molecular weight. Acta Biomater. 2010, 6, 3676–3686. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Gu, L.-S.; Bryan, T.E.; Kim, J.R.; Chen, L.; Liu, Y.; Yoon, J.C.; Breschi, L.; Pashley, D.H.; Tay, F.R. Mineralisation of reconstituted collagen using polyvinylphosphonic acid/polyacrylic acid templating matrix protein analogues in the presence of calcium, phosphate and hydroxyl ions. Biomaterials 2010, 31, 6618–6627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nudelman, F.; Pieterse, K.; George, A.; Bomans, P.H.; Friedrich, H.; Brylka, L.J.; Hilbers, P.A.; de With, G.; Sommerdijk, N.A. The role of collagen in bone apatite formation in the presence of hydroxyapatite nucleation inhibitors. Nat. Mater. 2010, 9, 1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Jacquet, R.; Lowder, E.; Landis, W.J. Refinement of collagen–mineral interaction: A possible role for osteocalcin in apatite crystal nucleation, growth and development. Bone 2015, 71, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Thula, T.T.; Rodriguez, D.E.; Lee, M.H.; Pendi, L.; Podschun, J.; Gower, L.B. In vitro mineralization of dense collagen substrates: A biomimetic approach toward the development of bone-graft materials. Acta Biomater. 2011, 7, 3158–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, F.H.; Landis, W.J. Deposition of apatite in mineralizing vertebrate extracellular matrices: A model of possible nucleation sites on type I collagen. Connect. Tissue Res. 2011, 52, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yang, Y.; Zhao, W.; Wang, Z.; Landis, W.J.; Cui, Q.; Sahai, N. Molecular mechanisms for intrafibrillar collagen mineralization in skeletal tissues. Biomaterials 2015, 39, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, B.; Thomopoulos, S.; Jun, Y.-S. In situ evaluation of calcium phosphate nucleation kinetics and pathways during intra-and extrafibrillar mineralization of collagen matrices. Cryst. Growth Des. 2016, 16, 5359–5366. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.; Xia, Z.; Knecht, D.A.; Wei, M. Synthesis of dense collagen/apatite composites using a biomimetic method. J. Am. Ceram. Soc. 2008, 91, 3211–3215. [Google Scholar] [CrossRef]
- Hu, C.; Yu, L.; Wei, M. Sectioning studies of biomimetic collagen-hydroxyapatite coatings on Ti-6Al-4V substrates using focused ion beam. Appl. Surf. Sci. 2018, 444, 590–597. [Google Scholar] [CrossRef]
- Gebauer, D.; Cölfen, H. Prenucleation clusters and non-classical nucleation. Nano Today 2011, 6, 564–584. [Google Scholar] [CrossRef] [Green Version]
- Habraken, W.J.; Tao, J.; Brylka, L.J.; Friedrich, H.; Bertinetti, L.; Schenk, A.S.; Verch, A.; Dmitrovic, V.; Bomans, P.H.; Frederik, P.M. Ion-association complexes unite classical and non-classical theories for the biomimetic nucleation of calcium phosphate. Nat. Commun. 2013, 4, 1507. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, A.S.; Beniash, E. Bioinspired synthesis of mineralized collagen fibrils. Cryst. Growth Des. 2008, 8, 3084–3090. [Google Scholar] [CrossRef] [Green Version]
- Gower, L.; Tirrell, D. Calcium carbonate films and helices grown in solutions of poly (aspartate). J. Cryst. Growth 1998, 191, 153–160. [Google Scholar] [CrossRef]
- Gower, L.B.; Odom, D.J. Deposition of calcium carbonate films by a polymer-induced liquid-precursor (PILP) process. J. Cryst. Growth 2000, 210, 719–734. [Google Scholar] [CrossRef]
- Olszta, M.J.; Cheng, X.; Jee, S.S.; Kumar, R.; Kim, Y.-Y.; Kaufman, M.J.; Douglas, E.P.; Gower, L.B. Bone structure and formation: A new perspective. Mater. Sci. Eng. R Rep. 2007, 58, 77–116. [Google Scholar] [CrossRef]
- Xu, Y.; Tijssen, K.C.; Bomans, P.H.; Akiva, A.; Friedrich, H.; Kentgens, A.P.; Sommerdijk, N.A. Microscopic structure of the polymer-induced liquid precursor for calcium carbonate. Nat. Commun. 2018, 9, 2582. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.L.; Caballero, L.; Melo, F.; Cölfen, H. Gel-like calcium carbonate precursors observed by in situ AFM. Langmuir 2017, 33, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-X.; Hoff, S.E.; Huang, X.-Q.; Liu, J.; Wan, Q.-Q.; Song, Q.; Gu, J.-T.; Heinz, H.; Tay, F.R.; Niu, L.-N. Involvement of prenucleation clusters in calcium phosphate mineralization of collagen. Acta Biomater. 2020, 120, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Garcia, N.A.; Malini, R.I.; Freeman, C.L.; Demichelis, R.; Raiteri, P.; Sommerdijk, N.A.; Harding, J.H.; Gale, J.D. Simulation of calcium phosphate prenucleation clusters in aqueous solution: Association beyond ion pairing. Cryst. Growth Des. 2019, 19, 6422–6430. [Google Scholar] [CrossRef] [PubMed]
- Sugawara-Narutaki, A.; Nakamura, J.; Ohtsuki, C. Bioceramics; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Olszta, M.; Douglas, E.; Gower, L. Scanning electron microscopic analysis of the mineralization of type I collagen via a polymer-induced liquid-precursor (PILP) process. Calcif. Tissue Int. 2003, 72, 583–591. [Google Scholar] [CrossRef]
- Olszta, M.J.; Douglas, E.P.; Gower, L.B. Intrafibrillar mineralization of collagen using a liquid-phase mineral precursor. MRS Online Proc. Libr. Arch. 2003, 774. [Google Scholar] [CrossRef]
- Olszta, M.J.; Odom, D.J.; Douglas, E.P.; Gower, L.B. A new paradigm for biomineral formation: Mineralization via an amorphous liquid-phase precursor. Connect. Tissue Res. 2003, 44, 326–334. [Google Scholar] [CrossRef]
- Jee, S.S.; Culver, L.; Li, Y.; Douglas, E.P.; Gower, L.B. Biomimetic mineralization of collagen via an enzyme-aided PILP process. J. Cryst. Growth 2010, 312, 1249–1256. [Google Scholar] [CrossRef]
- Jee, S.S.; Kasinath, R.K.; DiMasi, E.; Kim, Y.-Y.; Gower, L. Oriented hydroxyapatite in turkey tendon mineralized via the polymer-induced liquid-precursor (PILP) process. CrystEngComm 2011, 13, 2077–2083. [Google Scholar] [CrossRef]
- França, C.M.; Thrivikraman, G.; Athirasala, A.; Tahayeri, A.; Gower, L.B.; Bertassoni, L.E. The influence of osteopontin-guided collagen intrafibrillar mineralization on pericyte differentiation and vascularization of engineered bone scaffolds. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 1522–1532. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.; Cremer, M.A.; Dolling, E.S.; Nurrohman, H.; Habelitz, S.; Marshall, G.W.; Gower, L.B. Influence of fluoride on the mineralization of collagen via the polymer-induced liquid-precursor (PILP) process. Dent. Mater. 2018, 34, 1378–1390. [Google Scholar] [CrossRef]
- James, B.D.; Guerin, P.; Iverson, Z.; Allen, J.B. Mineralized DNA-collagen complex-based biomaterials for bone tissue engineering. Int. J. Biol. Macromol. 2020, 161, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Li, S.T.; Katz, E.P. An electrostatic model for collagen fibrils. The interaction of reconstituted collagen with Ca++, Na+, and Cl−. Biopolym. Orig. Res. Biomol. 1976, 15, 1439–1460. [Google Scholar] [CrossRef] [PubMed]
- Nudelman, F.; Lausch, A.J.; Sommerdijk, N.A.; Sone, E.D. In vitro models of collagen biomineralization. J. Struct. Biol. 2013, 183, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Azaïs, T.; Robin, M.; Vallée, A.; Catania, C.; Legriel, P.; Pehau-Arnaudet, G.; Babonneau, F.; Giraud-Guille, M.-M.; Nassif, N. The predominant role of collagen in the nucleation, growth, structure and orientation of bone apatite. Nat. Mater. 2012, 11, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Gu, L.; Huang, Z.; Sun, Q.; Chen, H.; Ling, J.; Mai, S. Intrafibrillar mineralization of polyacrylic acid-bound collagen fibrils using a two-dimensional collagen model and Portland cement-based resins. Eur. J. Oral Sci. 2017, 125, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, D.E.; Thula-Mata, T.; Toro, E.J.; Yeh, Y.-W.; Holt, C.; Holliday, L.S.; Gower, L.B. Multifunctional role of osteopontin in directing intrafibrillar mineralization of collagen and activation of osteoclasts. Acta Biomater. 2014, 10, 494–507. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Griffanti, G.; Tamimi, F.; McKee, M.D.; Nazhat, S.N. Multiscale structural evolution of citrate-triggered intrafibrillar and interfibrillar mineralization in dense collagen gels. J. Struct. Biol. 2020, 212, 107592. [Google Scholar] [CrossRef]
- Yu, L.; Martin, I.J.; Kasi, R.M.; Wei, M. Enhanced intrafibrillar mineralization of collagen fibrils induced by brushlike polymers. ACS Appl. Mater. Interfaces 2018, 10, 28440–28449. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Rowe, D.W.; Perera, I.P.; Zhang, J.; Suib, S.L.; Xin, X.; Wei, M. Intrafibrillar Mineralized Collagen-Hydroxyapatite-Based Scaffolds for Bone Regeneration. ACS Appl. Mater. Interfaces 2020, 12, 18235–18249. [Google Scholar] [CrossRef]
- Hu, C.; Yu, L.; Wei, M. Biomimetic intrafibrillar silicification of collagen fibrils through a one-step collagen self-assembly/silicification approach. RSC Adv. 2017, 7, 34624–34632. [Google Scholar] [CrossRef] [Green Version]
- Kerns, J.G.; Buckley, K.; Churchwell, J.; Parker, A.W.; Matousek, P.; Goodship, A.E. Is the collagen primed for mineralization in specific regions of the turkey tendon? An investigation of the protein–mineral interface using Raman spectroscopy. Anal. Chem. 2016, 88, 1559–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, K.; Niu, L.N.; Ma, C.F.; Huang, X.Q.; Pei, D.D.; Luo, T.; Huang, Q.; Chen, J.H.; Tay, F.R. Complementarity and uncertainty in intrafibrillar mineralization of collagen. Adv. Funct. Mater. 2016, 26, 6858–6875. [Google Scholar] [CrossRef]
- Li, Y.; Rodriguez-Cabello, J.C.; Aparicio, C. Intrafibrillar mineralization of self-assembled elastin-like recombinamer fibrils. ACS Appl. Mater. Interfaces 2017, 9, 5838–5846. [Google Scholar] [CrossRef]
- Niu, L.-N.; Jee, S.E.; Jiao, K.; Tonggu, L.; Li, M.; Wang, L.; Yang, Y.-D.; Bian, J.-H.; Breschi, L.; Jang, S.S. Collagen intrafibrillar mineralization as a result of the balance between osmotic equilibrium and electroneutrality. Nat. Mater. 2017, 16, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Jiao, K.; Tonggu, L.; Wang, L.; Zhang, S.; Yang, Y.; Zhang, L.; Bian, J.; Hao, D.; Wang, C. Contribution of biomimetic collagen-ligand interaction to intrafibrillar mineralization. Sci. Adv. 2019, 5, eaav9075. [Google Scholar] [CrossRef] [Green Version]
- Toroian, D.; Lim, J.E.; Price, P.A. The size exclusion characteristics of type I collagen implications for the role of noncollagenous bone constituents in mineralization. J. Biol. Chem. 2007, 282, 22437–22447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, P.A.; Toroian, D.; Lim, J.E. Mineralization by inhibitor exclusion the calcification of collagen with fetuin. J. Biol. Chem. 2009, 284, 17092–17101. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Gill, G.; Kaur, H.; Amhmed, M.; Jakhu, H. Role of osteopontin in bone remodeling and orthodontic tooth movement: A review. Prog. Orthod. 2018, 19, 18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ustriyana, P.; Chen, K.; Zhao, W.; Xu, Z.; Sahai, N. Towards the Understanding of Small Protein-Mediated Collagen Intrafibrillar Mineralization. ACS Biomater. Sci. Eng. 2020. [Google Scholar] [CrossRef]
- Cai, M.M.; Smith, E.R.; Holt, S.G. The role of fetuin-A in mineral trafficking and deposition. BoneKEy Rep. 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ping, H.; Xie, H.; Su, B.-L.; Cheng, Y.-B.; Wang, W.; Wang, H.; Wang, Y.; Zhang, J.; Zhang, F.; Fu, Z. Organized intrafibrillar mineralization, directed by a rationally designed multi-functional protein. J. Mater. Chem. B 2015, 3, 4496–4502. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wu, H.; Luo, X.-J. Biomineralization Precursor Carrier System Based on Carboxyl-Functionalized Large Pore Mesoporous Silica Nanoparticles. Curr. Med. Sci. 2020, 40, 155–167. [Google Scholar] [CrossRef]
- Du, T.; Niu, X.; Hou, S.; Li, Z.; Li, P.; Fan, Y. Apatite minerals derived from collagen phosphorylation modification induce the hierarchical intrafibrillar mineralization of collagen fibers. J. Biomed. Mater. Res. Part A 2019, 107, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Marbach, S.; Bocquet, L. Osmosis, from molecular insights to large-scale applications. Chem. Soc. Rev. 2019, 48, 3102–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, B.D.; Sone, E.D. The effect of polyaspartate chain length on mediating biomimetic remineralization of collagenous tissues. J. R. Soc. Interface 2018, 15, 20180269. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Tang, T.; Macías-Sánchez, E.; Sviben, S.; Landis, W.J.; Bertinetti, L.; Fratzl, P. Three-dimensional structural interrelations between cells, extracellular matrix, and mineral in normally mineralizing avian leg tendon. Proc. Natl. Acad. Sci. USA 2020. [Google Scholar] [CrossRef]
- Hor, J.L.; Wang, H.; Fakhraai, Z.; Lee, D. Effect of Physical Nanoconfinement on the Viscosity of Unentangled Polymers during Capillary Rise Infiltration. Macromolecules 2018, 51, 5069–5078. [Google Scholar] [CrossRef]
- Vinay, T.V.; Banuprasad, T.N.; George, S.D.; Varghese, S.; Varanakkottu, S.N. Additive-Free Tunable Transport and Assembly of Floating Objects at Water-Air Interface Using Bubble-Mediated Capillary Forces. Adv. Mater. Interfaces 2017, 4, 1601231. [Google Scholar] [CrossRef]
- Feng, J.; Song, Q.; Zhang, B.; Wu, Y.; Wang, T.; Jiang, L. Large-Scale, Long-Range-Ordered Patterning of Nanocrystals via Capillary-Bridge Manipulation. Adv. Mater. 2017, 29, 1703143. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Zhao, R.; Jiang, S.; Yao, S.; Wu, Z.; Jin, B.; Yang, Y.; Pan, H.; Tang, R. Citrate improves collagen mineralization via interface wetting: A physicochemical understanding of biomineralization control. Adv. Mater. 2018, 30, 1704876. [Google Scholar] [CrossRef]
- Qu, Y.; Gu, T.; Du, Q.; Shao, C.; Wang, J.; Jin, B.; Kong, W.; Sun, J.; Chen, C.; Pan, H. Polydopamine Promotes Dentin Remineralization via Interfacial Control. ACS Biomater. Sci. Eng. 2020, 6, 3327–3334. [Google Scholar] [CrossRef]
- He, L.; Hao, Y.; Zhen, L.; Liu, H.; Shao, M.; Xu, X.; Liang, K.; Gao, Y.; Yuan, H.; Li, J. Biomineralization of dentin. J. Struct. Biol. 2019, 207, 115–122. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J.; Andrei, C.M.; Soleymani, L.; Grandfield, K. Biomineralization of calcium phosphate revealed by in situ liquid-phase electron microscopy. Commun. Chem. 2018, 1, 80. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Rehman, F.U.; Zhao, C.; Liu, B.; He, N. Recent advances in nano scaffolds for bone repair. Bone Res. 2016, 4, 16050. [Google Scholar] [CrossRef]
- Zou, L.; Zhang, Y.; Liu, X.; Chen, J.; Zhang, Q. Biomimetic mineralization on natural and synthetic polymers to prepare hybrid scaffolds for bone tissue engineering. Colloids Surf. B Biointerfaces 2019, 178, 222–229. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, D.; Kou, X.X.; Wang, X.D.; Tay, F.R.; Sha, Y.L.; Gan, Y.H.; Zhou, Y.H. Hierarchical intrafibrillar nanocarbonated apatite assembly improves the nanomechanics and cytocompatibility of mineralized collagen. Adv. Funct. Mater. 2013, 23, 1404–1411. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Z.; Zhang, C.; Liu, J.; Bai, Y.; Li, S.; Zhang, C. Biomimetic intrafibrillar mineralized collagen promotes bone regeneration via activation of the Wnt signaling pathway. Int. J. Nanomed. 2018, 13, 7503. [Google Scholar] [CrossRef] [Green Version]
- Chien, Y.-C.; Tao, J.; Saeki, K.; Chin, A.F.; Lau, J.L.; Chen, C.-L.; Zuckermann, R.N.; Marshall, S.J.; Marshall, G.W.; De Yoreo, J.J. Using biomimetic polymers in place of noncollagenous proteins to achieve functional remineralization of dentin tissues. ACS Biomater. Sci. Eng. 2017, 3, 3469–3479. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Jin, R.; Li, X.; Fang, X.; Yuan, D.; Chen, Z.; Yao, S.; Tang, R.; Chen, Z. Biomimetic remineralization of artificial caries dentin lesion using Ca/P-PILP. Dent. Mater. 2020, 36, 1397–1406. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Jiang, T.; Wang, Y. Biomimetic regulation of dentine remineralization by amino acid in vitro. Dent. Mater. 2019, 35, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Martins, L.F.; Sousa, J.P.d.; Alves, L.A.; Davies, R.P.W.; Puppin-Rontanti, R.M. Biomimetic mineralizing agents recover the micro tensile bond strength of demineralized dentin. Materials 2018, 11, 1733. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.Y.; Mei, M.L.; Li, Q.-L.; Lo, E.C.M.; Chu, C.H. Methods for biomimetic remineralization of human dentine: A systematic review. Int. J. Mol. Sci. 2015, 16, 4615–4627. [Google Scholar] [CrossRef]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2016, 82, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, P.; Grüner, D.; Worch, H.; Pompe, W.; Lichte, H.; El Khassawna, T.; Heiss, C.; Wenisch, S.; Kniep, R. First evidence of octacalcium phosphate@ osteocalcin nanocomplex as skeletal bone component directing collagen triple–helix nanofibril mineralization. Sci. Rep. 2018, 8, 13696. [Google Scholar] [CrossRef]
- Moriishi, T.; Ozasa, R.; Ishimoto, T.; Nakano, T.; Hasegawa, T.; Miyazaki, T.; Liu, W.; Fukuyama, R.; Wang, Y.; Komori, H. Osteocalcin is necessary for the alignment of apatite crystallites, but not glucose metabolism, testosterone synthesis, or muscle mass. PLoS Genet. 2020, 16, e1008586. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, Q.; Alam, A.; Cui, J.; Suen, K.C.; Soo, A.P.; Eguchi, S.; Gu, J.; Ma, D. The role of osteopontin in the progression of solid organ tumour. Cell Death Dis. 2018, 9, 356. [Google Scholar] [CrossRef]
- Zuo, Q.; Yao, J.; Lu, S.; Du, Z.; Li, S.; Lin, F.; Shi, W.; Zhang, Y.; Xiao, Y. The role of organic phosphate in the spatial control of periodontium complex bio-mineralization: An in vitro study. J. Mater. Chem. B 2019, 7, 5956–5965. [Google Scholar] [CrossRef]
- Ling, Z.; He, Y.; Huang, H.; Xie, X.; Li, Q.-l.; Cao, C.Y. Effects of oligopeptide simulating DMP-1/mineral trioxide aggregate/agarose hydrogel biomimetic mineralisation model for the treatment of dentine hypersensitivity. J. Mater. Chem. B 2019, 7, 5825–5833. [Google Scholar] [CrossRef] [PubMed]
- Brylka, L.; Jahnen-Dechent, W. The role of fetuin-A in physiological and pathological mineralization. Calcified Tissue Int. 2013, 93, 355–364. [Google Scholar] [CrossRef]
- Qi, Y.; Ye, Z.; Fok, A.; Holmes, B.N.; Espanol, M.; Ginebra, M.-P.; Aparicio, C. Effects of molecular weight and concentration of poly (acrylic acid) on biomimetic mineralization of collagen. ACS Biomater. Sci. Eng. 2018, 4, 2758–2766. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Van Manh, N.; Wang, H.; Zhong, X.; Zhang, X.; Li, C. Synergistic intrafibrillar/extrafibrillar mineralization of collagen scaffolds based on a biomimetic strategy to promote the regeneration of bone defects. Int. J. Nanomed. 2016, 11, 2053. [Google Scholar]
- Liu, H.; Lin, M.; Liu, X.; Zhang, Y.; Luo, Y.; Pang, Y.; Chen, H.; Zhu, D.; Zhong, X.; Ma, S. Doping bioactive elements into a collagen scaffold based on synchronous self-assembly/mineralization for bone tissue engineering. Bioact. Mater. 2020, 5, 844–858. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Mao, C.; Gu, T.; Pan, H.; Shao, C.; Sun, J.; Chen, C.; Tang, R.; Gu, X. Phosphorylated chitosan to promote biomimetic mineralization of type I collagen as a strategy for dentin repair and bone tissue engineering. New J. Chem. 2019, 43, 2002–2010. [Google Scholar] [CrossRef]
- Lin, X.; Xie, F.; Ma, X.; Hao, Y.; Qin, H.; Long, J. Fabrication and characterization of dendrimer-functionalized nano-hydroxyapatite and its application in dentin tubule occlusion. J. Biomater. Sci. Polym. Ed. 2017, 28, 846–863. [Google Scholar] [CrossRef] [PubMed]
- Bapat, R.A.; Dharmadhikari, S.; Chaubal, T.V.; Amin, M.C.I.M.; Bapat, P.; Gorain, B.; Choudhury, H.; Vincent, C.; Kesharwani, P. The potential of dendrimer in delivery of therapeutics for dentistry. Heliyon 2019, 5, e02544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Shao, C.; Mu, Z.; Mao, C.; Sun, J.; Chen, C.; Tang, R.; Gu, X. Promotion effect of immobilized chondroitin sulfate on intrafibrillar mineralization of collagen. Carbohydr. Polym. 2020, 229, 115547. [Google Scholar] [CrossRef]
- Su, W.; Ma, L.; Ran, Y.; Ma, X.; Yi, Z.; Chen, G.; Chen, X.; Li, X. Alginate-Assisted Mineralization of Collagen by Collagen Reconstitution and Calcium Phosphate Formation. ACS Biomater. Sci. Eng. 2020, 6, 3275–3286. [Google Scholar] [CrossRef]
- Sun, J.-L.; Jiao, K.; Niu, L.-N.; Jiao, Y.; Song, Q.; Shen, L.-J.; Tay, F.R.; Chen, J.-H. Intrafibrillar silicified collagen scaffold modulates monocyte to promote cell homing, angiogenesis and bone regeneration. Biomaterials 2017, 113, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Huang, Z.; Liu, N.; Tang, X.; Dusabe, E.; Zhou, B.; Mao, J. The Role of Genetically Engineered Peptides in Inducing Intrafibrillar Mineralization Using Calcium Phosphate Precursors. Sci. Adv. Mater. 2016, 8, 2204–2215. [Google Scholar] [CrossRef]
- Gungormus, M.; Tulumbaci, F. Peptide-assisted pre-bonding remineralization of dentin to improve bonding. J. Mech. Behav. Biomed. Mater. 2020, 113, 104119. [Google Scholar] [CrossRef]
- Mukherjee, K.; Visakan, G.; Phark, J.-H.; Moradian-Oldak, J. Enhancing Collagen Mineralization with Amelogenin Peptide: Toward the Restoration of Dentin. ACS Biomater. Sci. Eng. 2020, 6, 2251–2262. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Tian, X.; Kim, J.P.; Xie, D.; Ao, X.; Shan, D.; Lin, Q.; Hudock, M.R.; Bai, X.; Yang, J. Citrate-based materials fuel human stem cells by metabonegenic regulation. Proc. Natl. Acad. Sci. USA 2018, 115, E11741–E11750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wang, Y.; Dai, H.; Tian, X.; Cui, Z.-K.; Chen, Z.; Hu, L.; Song, Q.; Liu, A.; Zhang, Z. Bone and plasma citrate is reduced in osteoporosis. Bone 2018, 114, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Olson, T.Y.; Orme, C.A.; Han, T.Y.-J.; Worsley, M.A.; Rose, K.A.; Satcher, J.H.; Kuntz, J.D. Shape control synthesis of fluorapatite structures based on supersaturation: Prismatic nanowires, ellipsoids, star, and aggregate formation. CrystEngComm 2012, 14, 6384–6389. [Google Scholar] [CrossRef]
- Pajor, K.; Pajchel, L.; Kolmas, J. Hydroxyapatite and fluorapatite in conservative dentistry and oral implantology—A review. Materials 2019, 12, 2683. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Fan, R.; Guo, X.; Du, T.; Yang, Z.; Feng, Q.; Fan, Y. Shear-mediated orientational mineralization of bone apatite on collagen fibrils. J. Mater. Chem. B 2017, 5, 9141–9147. [Google Scholar] [CrossRef]
- Du, T.; Niu, X.; Hou, S.; Xu, M.; Li, Z.; Li, P.; Fan, Y. Highly aligned hierarchical intrafibrillar mineralization of collagen induced by periodic fluid shear stress. J. Mater. Chem. B 2020, 8, 2562–2572. [Google Scholar] [CrossRef]
Figure 1.
Typical transmission electron microscopy (TEM) images of (a) highly oriented intrafibrillar (showing clear D-banding) and (b) extrafibrillar mineralized collagen fibrils. Images are modified from [24] with permission. Copyright @ John Wiley & Sons, Inc., Hoboken, NJ, USA.
Figure 1.
Typical transmission electron microscopy (TEM) images of (a) highly oriented intrafibrillar (showing clear D-banding) and (b) extrafibrillar mineralized collagen fibrils. Images are modified from [24] with permission. Copyright @ John Wiley & Sons, Inc., Hoboken, NJ, USA.

Figure 2.
Schematic illustration of non-collagenous proteins (NCPs)-regulated collagen biomineralization with both intrafibrillar and extrafibrillar mineralization according to the non-classical theory.
Figure 2.
Schematic illustration of non-collagenous proteins (NCPs)-regulated collagen biomineralization with both intrafibrillar and extrafibrillar mineralization according to the non-classical theory.


{kind=link}
{kind=link}
Table 1.
Recent examples of synthetic polymer analogs used to mediate biomimetic intrafibrillar mineralization of collagen.
Table 1.
Recent examples of synthetic polymer analogs used to mediate biomimetic intrafibrillar mineralization of collagen.
| Polymers | Major Findings | Ref. |
|---|---|---|
| PAA | Systematically investigated the effect of molecular weight and concentration of PAA on the rate and pattern of collagen intrafibrillar mineralization. Decreasing PAA molecular weight (2 kDa) and increasing PAA concentration (50 mg/L) resulted in increased stability of ACP solution and therefore enhanced intrafibrillar mineralization. | [106] |
| PAA or carboxyl-rich brush-like polymers; TPP | Two brush-like polymers derived from PEG were synthesized and modified with enriched carboxylic groups. Sodium tripolyphosphate (TPP) was used as a templating analog. The mineralization degree of collagen fibrils induced by brush-like polymers was higher than those induced by linear PAA. Therefore, the formation of intrafibrillarly mineralized apatite might be partially determined by the availability of carboxylic groups in the nucleation inhibitors. | [64] |
| PAA; TPP | Intrafibrillarly mineralized collagen-based scaffolds were engineered into both cellular and lamellar structures. Fe and Mn were incorporated separately or jointly into the lamellar scaffold. Lamellar scaffolds were much better in supporting in vitro osteogenic differentiation and in vivo bone regeneration than cellular scaffolds. These promotion effects were further enhanced by the addition of both Fe and Mn ions. | [65] |
| PASP | PASP chain length contributed to the effectiveness of mediating intrafibrillar mineralization. The process appeared to be associated with the inhibition of apatite crystallization by PASP through slowing the growth of ACP stabilizing this phase. | [81] |
| CMC | Collagen scaffolds with both intrafibrillar and extrafibrillar mineralization were obtained using carboxymethyl chitosan (CMC) to stabilize ACP. Such prepared scaffolds exhibited increased modulus, in vitro cell proliferation and differentiation, and in vivo new bone regeneration compared to unmineralized collagen scaffolds as well as those fabricated using traditional modified-simulated body fluid (m-SBF) solution without CMC. | [107] |
| CMC | Sr- and Ag-doped intrafibrillarly mineralized collagen scaffolds were developed. Ag-doped scaffolds showed enhanced antibacterial effect on S. aureus while Sr-doped scaffolds illustrated enhanced new bone regeneration. | [108] |
| Pchi | Phosphorylated chitosan (Pchi) was used to promote collagen intrafibrillar mineralization through a biomimetic approach. It was found that Pchi significantly shortened the self-assembly process by accelerating the rate of crystallization due to the excellent ion chelating properties of chitosan derivatives and their ability to induce high-degree conglutination. | [109] |
| PAMAM | Carboxyl-terminated hyperbranched polyamidoamine dendrimer (PAMAM) was used to induce biomimetic remineralization on dentine organic matrix. Such prepared material showed great potential to be used as a new therapeutic material for the treatment of dentin hypersensitivity. | [110,111] |
| PDA | Dentine was successfully repaired using polydopamine (PDA), which promoted intrafibrillar mineralization by decreasing the interfacial energy between collagen matrix and ACP. The re-mineralized dentin exhibited comparable mechanical properties to natural dentin. | [87] |
| CS | The immobilized chondroitin sulfate (CS) on collagen fibrils accelerated CaP nucleation and significantly promoted collagen intrafibrillar mineralization by providing specific sites for CaP nucleation within the collagen fibrils. Remarkably accelerated remineralization of CS immobilized demineralized dentin was achieved. | [112] |
| Alginate | As an anionic polyelectrolyte with Ca-capturing capacity, alginate was used to successfully mediate intrafibrillar mineralization of collagen. The alginate-assisted mineralization of collagen resulted in an exquisite three-dimensional (3D) mineralized architecture with enhanced mechanical properties as well as excellent proliferation, adhesion, and differentiation of rat BMSCs. | [113] |
| PAH; TPP | Intrafibrillarly silicified collagen scaffolds were prepared in the presence of PAH as a directing agent. The silicified scaffolds supported in vitro cell proliferation, in situ bone regeneration and angiogenesis via monocyte immunomodulation. | [66,114] |
| Peptide | Various kinds of mineral-promoting peptides with different amino acid sequences were designed to mimic the functions of NCPs to enhance intrafibrillar mineralization of collagen. The obtained materials displayed: (1) stronger influence on biomineralization than traditional used PAA; (2) excellent ability for rapid remineralization of dentin; or (3) apparent improvement in restoring incipient enamel decay and mineralization defects localized in peripheral dentin. | [115,116,117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, L.; Wei, M. Biomineralization of Collagen-Based Materials for Hard Tissue Repair. Int. J. Mol. Sci. 2021, 22, 944. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020944
AMA Style
Yu L, Wei M. Biomineralization of Collagen-Based Materials for Hard Tissue Repair. International Journal of Molecular Sciences. 2021; 22(2):944. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020944
Chicago/Turabian StyleYu, Le, and Mei Wei. 2021. "Biomineralization of Collagen-Based Materials for Hard Tissue Repair" International Journal of Molecular Sciences 22, no. 2: 944. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020944
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.