The Protein Phosphatase PP2A Plays Multiple Roles in Plant Development by Regulation of Vesicle Traffic—Facts and Questions

Abstract
:1. Introduction
2. PP2A and Vesicle Traffic in Plant Cells
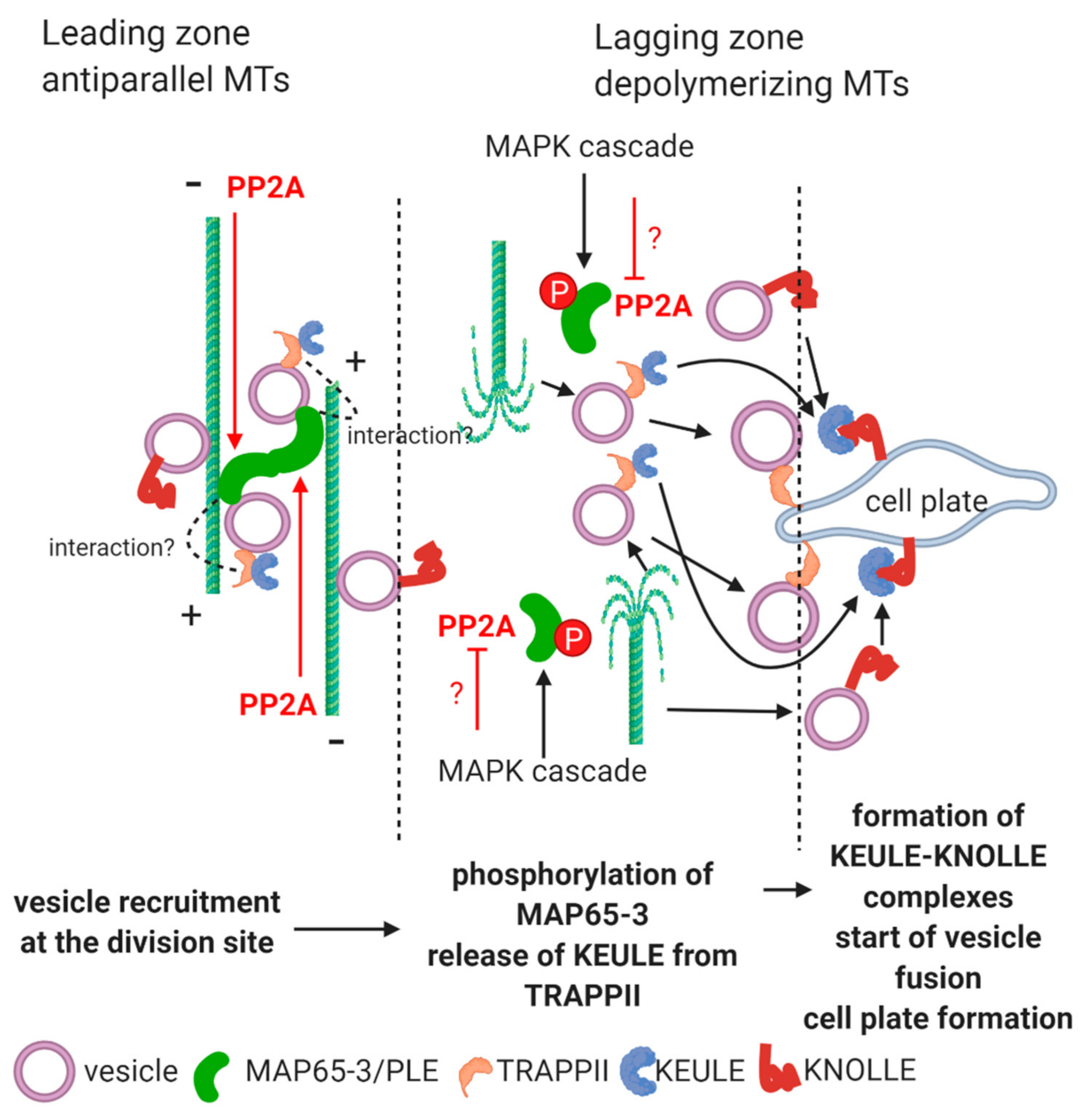
2.1. Phragmoplast, Vesicle Traffic and Cell Plate Assembly. Involvement of PP2A in a Developmental Context
2.1.1. Vesicle Delivery through MTs and Their Release at the Phragmoplast.
2.1.2. Vesicle Fusion during Cell Plate Formation
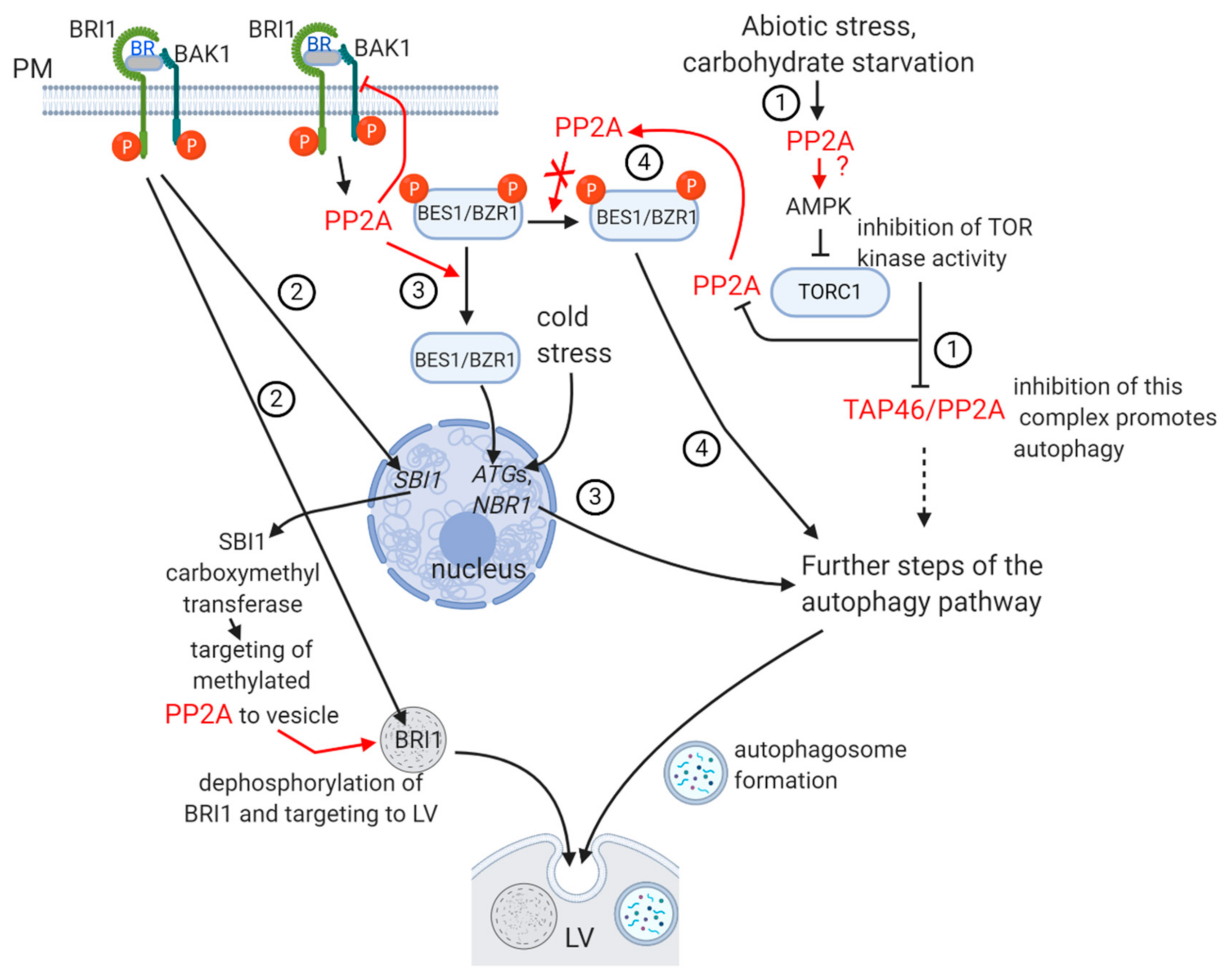
2.2. Pathways of Autophagy in Relation to PP2A; Developmental Aspects
Pathway 3 of Figure 2
- (i)
- What is the detailed mechanism of blocking of TORC-mediated signaling by PP2A (see Figure 2 as well)?
- (ii)
- Are biogenesis and fusion of autophagosomes to the lytic vacuole regulated by PP2A?
- (iii)
- Further evidence is needed to demonstrate that PP2A-mediated dephosphorylation of BZR, an important actor in BR signaling, is blocked during autophagy.
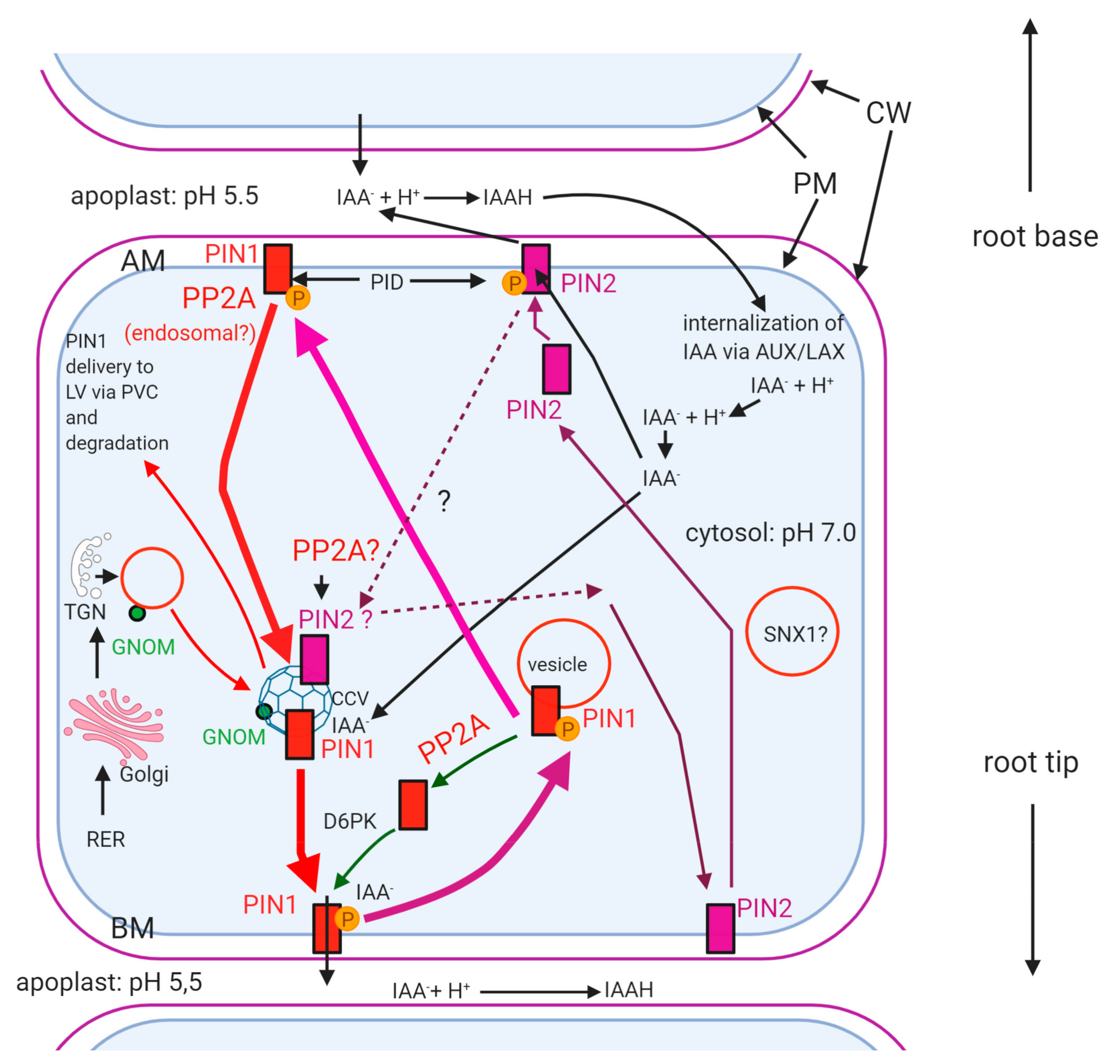
2.3. The Subcellular Fate of PINs in Relation to PP2A
- (i)
- Is internalization of IAA by AUX1/LAX regulated by reversible protein phosphorylation? This aspect is largely unknown.
- (ii)
- What mechanism directs differential membrane localization of PIN1 vs. PIN2 directed auxin efflux? How reversible PIN phosphorylation is involved in this (see Figure 3)? As we could see above, the reversible phosphorylation of PINs regulates their plasma membrane polar localization. Dephosphorylation will trigger internalization and then recycling, usually to the opposite cell side [90,91]. Phosphorylation of PIN1, 2 and 4 by PID results in their localization in the apical membrane of root cells, while dephosphorylation will redirect them to the basal plasma membrane ([92] and Figure 3). If PIN1 is maintained in a phosphorylated state at the apical membrane, auxin will be transported towards root base and the apical meristem of main roots will be depleted of auxin. At this site, it is phosphorylated by PID—the function of this is stabilization of PIN1 ([75]; Figure 3). What is the reason for the lack of auxin efflux by phospho-PIN1 at the apical membrane under normal conditions? PIN1 phosphorylation at the basal membrane is needed for proper auxin efflux, at the basal poles of plasma membranes in roots [78,91,92,93,94]. For PIN2, the mode and effects of reversible phosphorylation are contrasting to PIN1. How is phosphorylation directed differentially in different cell types and for different PINs? Normally, PIN2 should be localized apically in cell membranes, to direct upward auxin flux towards root base mainly in root epidermal cells. If PIN2 remains hyperphosphorylated, it will remain in the apical membranes and blocks its recycling [75]. As PIN1, PIN2 drives auxin efflux, when it is phosphorylated. At the basal (facing the root tip) pole of the cell, PIN1 is phosphorylated by brefeldin A (BFA) sensitive D6PK to drive auxin efflux and its local recycling and PP2A dephosphorylates it to drive its transcytosis into the apical (shootward) cell pole. In contrast to PIN1, PIN2 is not phosphorylated by D6PK at the basal pole of the cell, because its hydrophilic loop lacks the phosphorylation sites for D6PK [75]. Recently it has been shown that PID has a non-polar distribution, that is, it can be localized practically at any membrane sites [12]. Why then, its activity is “polarized” for PINs?
- (iii)
- Which A/B regulatory subunit(s) of PP2A are responsible for regulating phosphatase activity, correct targeting and substrate specificity to PIN? It was shown that the A scaffolding subunit of PP2A is not only stabilizing the holoenzyme, but it has other important regulatory functions [12]. RCN1 is an A (Aα) regulatory subunit of PP2A, important in polar auxin transport, thereby regulating mitotic activity in root apical meristem, root morphology, hypocotyl apical hook formation and axial organ elongation. PP2A/A mutations are altering root development and gravitropism by changing auxin transport, thus distribution [12,97]. In the rcn1 (roots curl in naphthylphtalamic acid) Arabidopsis mutant, PP2A activity was reduced, indicating this subunit is important for maintaining functionality of the holoenzyme. The loss-of-function of specific PP2A/A subunits results in the basal-to-apical shift of PIN1 membrane localization [68,70,93]. The A1 subunit is binding to phosphatidic acid derived from the activity of phospholipase D (PLD), thus it is crucial for the PIN1-related membrane localization of PP2A. Inhibition of PLD and/or mutated PP2AA1 results in the inhibition of PIN1 recycling leading to its mislocalization, i.e., accumulation on apical membranes [98]. However, it is still not clear why this mechanism is restricted to specific membrane areas? Is PLD localization polarized to drive polar localization of PIN1?
- (iv)
- Why only dephosphorylated PIN1 (and PIN2) is recycled via the GNOM pathway? There is evidence that phosphorylated PINs are not preferred by the GNOM-mediated vesicle traffic pathways. For example, Arabidopsis pp2aa1 plants are hypersensitive to BFA and GNOM-mediated transcytosis of PIN2 is impaired, probably because PIN2 remains phosphorylated due to the immobilizing of PP2AA1, thus preventing proper PP2A activity at the correct location. [77,84]. It was shown that GNOM decreases the abundance of PINOID, thus the level of phosphorylated PINs [84]. Is this inducing a conformation change of PINs that will facilitate GNOM-mediated vesicular transport? We are still far away from understanding the mechanism related to the above question.
- (v)
- Why only dephosphorylated excess PIN is poly-ubiquitinated and delivered to lytic vacuoles for degradation? PIN1 and PIN2 are targeted to the vacuole via other ARF-GEFs than GNOM [74]. How its PP2A-mediated dephosphorylation governs this pathway? What is the signaling mechanism involved?
3. Concluding Remarks
- How PP2A regulates vesicle fusion at the formation of cell plate? Is maintenance of phosphorylated state of MAP65 at the division plane caused by local PP2A inhibition? Does downregulation of PP2A at this level influence directly cell patterning in meristems?
- Is PP2A regulating directly the biogenesis of autophagosomes and their fusion to the lytic vacuole?
- How PP2A regulates differential vesicle traffic, membrane localization and functioning of different PINs? This research will contribute to a better understanding of auxin-regulated development of axial organs.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, R.D.; Walker, J.C. Plant Protein Phosphatases. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 101–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y. Serine/Threonine Phosphatases: Mechanism through Structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef] [Green Version]
- Virshup, D.M.; Shenolikar, S. From Promiscuity to Precision: Protein Phosphatases Get a Makeover. Mol. Cell 2009, 33, 537–545. [Google Scholar] [CrossRef]
- Uhrig, R.G.; Labandera, A.-M.; Moorhead, G.B. Arabidopsis PPP Family of Serine/Threonine Protein Phosphatases: Many Targets but Few Engines. Trends Plant Sci. 2013, 18, 505–513. [Google Scholar] [CrossRef]
- Farkas, I.; Dombrádi, V.; Miskei, M.; Szabados, L.; Koncz, C. Arabidopsis PPP Family of Serine/Threonine Phosphatases. Trends Plant Sci. 2007, 12, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Sigismund, S.; Confalonieri, S.; Ciliberto, A.; Polo, S.; Scita, G.; Di Fiore, P.P. Endocytosis and Signaling: Cell Logistics Shape the Eukaryotic Cell Plan. Physiol. Rev. 2012, 92, 273–366. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. (Eds.) Biochemistry and Molecular Biology of Plants, 2nd ed.; John Wiley & Sons: Chichester, UK, 2015; ISBN 978-0-470-71422-5. [Google Scholar]
- Baskin, T. The cytoskeleton. In Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; John Wiley & Sons: Chichester, UK, 2015; pp. 191–238. ISBN 978-0-470-71422-5. [Google Scholar]
- Smertenko, A.P.; Chang, H.-Y.; Sonobe, S.; Fenyk, S.I.; Weingartner, M.; Bögre, L.; Hussey, P.J. Control of the AtMAP65-1 Interaction with Microtubules through the Cell Cycle. J. Cell Sci. 2006, 119, 3227–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-G.; Zheng, A.; Jiang, L.; Rowse, M.; Stanevich, V.; Chen, H.; Li, Y.; Satyshur, K.A.; Johnson, B.; Gu, T.-J.; et al. Methylation-Regulated Decommissioning of Multimeric PP2A Complexes. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, H. Senescence and cell death. In Biochemistry and Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; John Wiley & Sons: Chichester, UK, 2015; pp. 191–238. ISBN 978-0-470-71422-5. [Google Scholar]
- Barbosa, I.C.R.; Hammes, U.Z.; Schwechheimer, C. Activation and Polarity Control of PIN-FORMED Auxin Transporters by Phosphorylation. Trends Plant Sci. 2018, 23, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Müller, S. Plant Cell Division—Defining and Finding the Sweet Spot for Cell Plate Insertion. Curr. Opin. Cell Biol. 2019, 60, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, R.; Steiner, A.; Assaad, F.F. Multisubunit Tethering Complexes in Higher Plants. Curr. Opin. Plant Biol. 2017, 40, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, C.; Azimzadeh, J.; Pastuglia, M.; Bellini, C.; Grandjean, O.; Bouchez, D. The Arabidopsis TONNEAU2 Gene Encodes a Putative Novel Protein Phosphatase 2A Regulatory Subunit Essential for the Control of the Cortical Cytoskeleton. Plant Cell 2002, 14, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-T.; Ahn, H.-K.; Pai, H.-S. The Subfamily II Catalytic Subunits of Protein Phosphatase 2A (PP2A) Are Involved in Cortical Microtubule Organization. Planta 2018, 248, 1551–1567. [Google Scholar] [CrossRef] [PubMed]
- Spinner, L.; Gadeyne, A.; Belcram, K.; Goussot, M.; Moison, M.; Duroc, Y.; Eeckhout, D.; Winne, N.D.; Schaefer, E.; Slijke, E.V.D.; et al. A Protein Phosphatase 2A Complex Spatially Controls Plant Cell Division. Nat. Commun. 2013, 4, 1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayaydin, F.; Vissi, E.; Mészáros, T.; Miskolczi, P.; Kovács, I.; Fehér, A.; Dombrádi, V.; Erdödi, F.; Gergely, P.; Dudits, D. Inhibition of Serine/Threonine-Specific Protein Phosphatases Causes Premature Activation of Cdc2MsF Kinase at G2/M Transition and Early Mitotic Microtubule Organisation in Alfalfa. Plant J. 2000, 23, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Beyer, D.; Tándor, I.; Kónya, Z.; Bátori, R.; Roszik, J.; Vereb, G.; Erdődi, F.; Vasas, G.; M-Hamvas, M.; Jambrovics, K.; et al. Microcystin-LR, a Protein Phosphatase Inhibitor, Induces Alterations in Mitotic Chromatin and Microtubule Organization Leading to the Formation of Micronuclei in Vicia Faba. Ann. Bot. 2012, 110, 797–808. [Google Scholar] [CrossRef] [Green Version]
- Máthé, C.; Beyer, D.; Erdődi, F.; Serfőző, Z.; Székvölgyi, L.; Vasas, G.; M-Hamvas, M.; Jámbrik, K.; Gonda, S.; Kiss, A. Microcystin-LR Induces Abnormal Root Development by Altering Microtubule Organization in Tissue-Cultured Common Reed (Phragmites Australis) Plantlets. Aquat. Toxicol. 2009, 92, 122–130. [Google Scholar] [CrossRef]
- She, Z.-Y.; Wei, Y.-L.; Lin, Y.; Li, Y.-L.; Lu, M.-H. Mechanisms of the Ase1/PRC1/MAP65 Family in Central Spindle Assembly. Biol. Rev. 2019, 94, 2033–2048. [Google Scholar] [CrossRef]
- Hamada, T. Microtubule Organization and Microtubule-Associated Proteins in Plant Cells. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 312, pp. 1–52. [Google Scholar]
- Vavrdová, T.; Šamaj, J.; Komis, G. Phosphorylation of Plant Microtubule-Associated Proteins During Cell Division. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Beck, M.; Komis, G.; Müller, J.; Menzel, D.; Šamaj, J. Arabidopsis Homologs of Nucleus- and Phragmoplast-Localized Kinase 2 and 3 and Mitogen-Activated Protein Kinase 4 Are Essential for Microtubule Organization. Plant Cell 2010, 22, 755–771. [Google Scholar] [CrossRef] [Green Version]
- Chang-Jie, J.; Sonobe, S. Identification and Preliminary Characterization of a 65 KDa Higher-Plant Microtubule-Associated Protein. J. Cell Sci. 1993, 105, 891–901. [Google Scholar] [PubMed]
- Hussey, P.J.; Hawkins, T.J.; Igarashi, H.; Kaloriti, D.; Smertenko, A. The Plant Cytoskeleton: Recent Advances in the Study of the Plant Microtubule-Associated Proteins MAP-65, MAP-190 and the Xenopus MAP215-like Protein, MOR1. Plant Mol. Biol. 2002, 50, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.-M.K.; Lee, Y.-R.J.; Kiyama, L.D.; Dinesh-Kumar, S.P.; Liu, B. Arabidopsis Microtubule-Associated Protein MAP65-3 Cross-Links Antiparallel Microtubules toward Their Plus Ends in the Phragmoplast via Its Distinct C-Terminal Microtubule Binding Domain. Plant Cell 2012, 24, 2071–2085. [Google Scholar] [CrossRef] [Green Version]
- Smertenko, A. Phragmoplast Expansion: The Four-Stroke Engine That Powers Plant Cytokinesis. Curr. Opin. Plant Biol. 2018, 46, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T. Microtubule-Associated Proteins in Higher Plants. J. Plant Res. 2007, 120, 79–98. [Google Scholar] [CrossRef]
- Sasabe, M.; Soyano, T.; Takahashi, Y.; Sonobe, S.; Igarashi, H.; Itoh, T.J.; Hidaka, M.; Machida, Y. Phosphorylation of NtMAP65-1 by a MAP Kinase down-Regulates Its Activity of Microtubule Bundling and Stimulates Progression of Cytokinesis of Tobacco Cells. Genes Dev. 2006, 20, 1004–1014. [Google Scholar] [CrossRef] [Green Version]
- Steiner, A.; Rybak, K.; Altmann, M.; McFarlane, H.E.; Klaeger, S.; Nguyen, N.; Facher, E.; Ivakov, A.; Wanner, G.; Kuster, B.; et al. Cell Cycle-Regulated PLEIADE/AtMAP65-3 Links Membrane and Microtubule Dynamics during Plant Cytokinesis. Plant J. 2016, 88, 531–541. [Google Scholar] [CrossRef] [Green Version]
- MacKintosh, C.; Diplexcito, J. CHAPTER 102—Naturally Occurring Inhibitors of Protein Serine/Threonine Phosphatases. In Handbook of Cell Signaling; Bradshaw, R.A., Dennis, E.A., Eds.; Academic Press: Burlington, NJ, USA, 2003; pp. 607–611. ISBN 978-0-12-124546-7. [Google Scholar]
- Soyano, T.; Nishihama, R.; Morikiyo, K.; Ishikawa, M.; Machida, Y. NQK1/NtMEK1 Is a MAPKK That Acts in the NPK1 MAPKKK-Mediated MAPK Cascade and Is Required for Plant Cytokinesis. Genes Dev. 2003, 17, 1055–1067. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Soyano, T.; Sasabe, M.; Machida, Y. A MAP Kinase Cascade That Controls Plant Cytokinesis. J. Biochem. 2004, 136, 127–132. [Google Scholar] [CrossRef]
- Boruc, J.; Weimer, A.K.; Stoppin-Mellet, V.; Mylle, E.; Kosetsu, K.; Cedeño, C.; Jaquinod, M.; Njo, M.; De Milde, L.; Tompa, P.; et al. Phosphorylation of MAP65-1 by Arabidopsis Aurora kinases is required for efficient cell cycle progression. Plant Physiol. 2017, 173, 582–599. [Google Scholar] [CrossRef] [Green Version]
- Petrovská, B.; Cenklová, V.; Pochylová, Ž.; Kourová, H.; Doskočilová, A.; Plíhal, O.; Binarová, L.; Binarová, P. Plant Aurora kinases play a role in maintenance of primary meristems and control of endoreduplication. New Phytol. 2012, 193, 590–604. [Google Scholar] [CrossRef]
- Müller, S.; Smertenko, A.; Wagner, V.; Heinrich, M.; Hussey, P.J.; Hauser, M.-T. The Plant Microtubule-Associated Protein AtMAP65-3/PLE Is Essential for Cytokinetic Phragmoplast Function. Curr. Biol. 2004, 14, 412–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasabe, M.; Machida, Y. Regulation of Organization and Function of Microtubules by the Mitogen-Activated Protein Kinase Cascade during Plant Cytokinesis. Cytoskeleton 2012, 69, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Sasabe, M.; Boudolf, V.; Veylder, L.D.; Inzé, D.; Genschik, P.; Machida, Y. Phosphorylation of a Mitotic Kinesin-like Protein and a MAPKKK by Cyclin-Dependent Kinases (CDKs) Is Involved in the Transition to Cytokinesis in Plants. Proc. Natl. Acad. Sci. USA 2011, 108, 17844–17849. [Google Scholar] [CrossRef] [Green Version]
- Carr, C.M.; Rizo, J. At the Junction of SNARE and SM Protein Function. Curr. Opin. Cell Biol. 2010, 22, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Südhof, T.C.; Rothman, J.E. Membrane Fusion: Grappling with SNARE and SM Proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, A.; Müller, L.; Rybak, K.; Vodermaier, V.; Facher, E.; Thellmann, M.; Ravikumar, R.; Wanner, G.; Hauser, M.-T.; Assaad, F.F. The Membrane-Associated Sec1/Munc18 KEULE Is Required for Phragmoplast Microtubule Reorganization During Cytokinesis in Arabidopsis. Mol. Plant 2016, 9, 528–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, S.; Jürgens, G. Plant Cytokinesis—No Ring, No Constriction but Centrifugal Construction of the Partitioning Membrane. Semin. Cell Dev. Biol. 2016, 53, 10–18. [Google Scholar] [CrossRef]
- Park, M.; Touihri, S.; Müller, I.; Mayer, U.; Jürgens, G. Sec1/Munc18 Protein Stabilizes Fusion-Competent Syntaxin for Membrane Fusion in Arabidopsis Cytokinesis. Dev. Cell 2012, 22, 989–1000. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Zhang, X.; Otegui, M.S. Plant Autophagy: New Flavors on the Menu. Curr. Opin. Plant Biol. 2018, 46, 113–121. [Google Scholar] [CrossRef]
- Dobrenel, T.; Caldana, C.; Hanson, J.; Robaglia, C.; Vincentz, M.; Veit, B.; Meyer, C. TOR Signaling and Nutrient Sensing. Annu. Rev. Plant Biol. 2016, 67, 261–285. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bassham, D.C. Autophagy: Pathways for Self-Eating in Plant Cells. Annu. Rev. Plant Biol. 2012, 63, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Nagy, M.; Kéki, S.; Rácz, D.; Mathur, J.; Vereb, G.; Garda, T.; M-Hamvas, M.; Chaumont, F.; Bóka, K.; Böddi, B.; et al. Novel Fluorochromes Label Tonoplast in Living Plant Cells and Reveal Changes in Vacuolar Organization after Treatment with Protein Phosphatase Inhibitors. Protoplasma 2018, 255, 829–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasaras, A.; Melzer, M.; Kunze, R. Arabidopsis Senescence-Associated Protein DMP1 Is Involved in Membrane Remodeling of the ER and Tonoplast. BMC Plant Biol. 2012, 12, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaillat, L.; Baars, T.L.; Mayer, A. Cell-Free Reconstitution of Vacuole Membrane Fragmentation Reveals Regulation of Vacuole Size and Number by TORC1. Mol. Biol. Cell 2012, 23, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.S.; Han, J.-A.; Lee, H.-S.; Lee, S.; Pai, H.-S. The PP2A Regulatory Subunit Tap46, a Component of the TOR Signaling Pathway, Modulates Growth and Metabolism in Plants. Plant Cell 2011, 23, 185–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacinto, E.; Hall, M.N. TOR Signalling in Bugs, Brain and Brawn. Nat. Rev. Mol. Cell Biol. 2003, 4, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wang, P.; Xiong, Y. Target of Rapamycin Signaling in Plant Stress Responses. Plant Physiol. 2020, 182, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- Viotti, C.; Krüger, F.; Krebs, M.; Neubert, C.; Fink, F.; Lupanga, U.; Scheuring, D.; Boutté, Y.; Frescatada-Rosa, M.; Wolfenstetter, S.; et al. The Endoplasmic Reticulum Is the Main Membrane Source for Biogenesis of the Lytic Vacuole in Arabidopsis. Plant Cell 2013, 25, 3434–3449. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J.; Emr, S.D. Autophagy as a Regulated Pathway of Cellular Degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Roustan, V.; Jain, A.; Teige, M.; Ebersberger, I.; Weckwerth, W. An Evolutionary Perspective of AMPK–TOR Signaling in the Three Domains of Life. J. Exp. Bot. 2016, 67, 3897–3907. [Google Scholar] [CrossRef] [PubMed]
- Ketelaar, T.; Voss, C.; Dimmock, S.A.; Thumm, M.; Hussey, P.J. Arabidopsis Homologues of the Autophagy Protein Atg8 Are a Novel Family of Microtubule Binding Proteins. FEBS Lett. 2004, 567, 302–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, T.M.; Brennan, B.; Yang, M.; Chen, J.; Zhang, M.; Li, Z.; Wang, X.; Bassham, D.C.; Walley, J.; Yin, Y. Selective Autophagy of BES1 Mediated by DSK2 Balances Plant Growth and Survival. Dev. Cell 2017, 41, 33–46.e7. [Google Scholar] [CrossRef] [Green Version]
- Suttangkakul, A.; Li, F.; Chung, T.; Vierstra, R.D. The ATG1/ATG13 Protein Kinase Complex Is Both a Regulator and a Target of Autophagic Recycling in Arabidopsis. Plant Cell 2011, 23, 3761–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, H. Senescence, Ageing and Death of the Whole Plant. New Phytol. 2013, 197, 696–711. [Google Scholar] [CrossRef]
- Kim, T.-W.; Wang, Z.-Y. Brassinosteroid Signal Transduction from Receptor Kinases to Transcription Factors. Annu. Rev. Plant Biol. 2010, 61, 681–704. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Wang, X.; Li, X.; Kamiya, Y.; Otegui, M.S.; Chory, J. Methylation of a Phosphatase Specifies Dephosphorylation and Degradation of Activated Brassinosteroid Receptors. Sci. Signal. 2011, 4, ra29. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.; Li, X.; Fang, P.; Xia, X.; Shi, K.; Zhou, Y.; Zhou, J.; Yu, J. Brassinosteroids Act as a Positive Regulator of NBR1-Dependent Selective Autophagy in Response to Chilling Stress in Tomato. J. Exp. Bot. 2020, 71, 1092–1106. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.-J.; Wang, K.-X.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q.; Zhou, J. BZR1 Mediates Brassinosteroid-Induced Autophagy and Nitrogen Starvation in Tomato. Plant Physiol. 2019, 179, 671–685. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, J.-Y.; Roh, J.; Marchive, C.; Kim, S.-K.; Meyer, C.; Sun, Y.; Wang, W.; Wang, Z.-Y. TOR Signaling Promotes Accumulation of BZR1 to Balance Growth with Carbon Availability in Arabidopsis. Curr. Biol. 2016, 26, 1854–1860. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.; Chen, J.; Yin, Y. Cross-Talk of Brassinosteroid Signaling in Controlling Growth and Stress Responses. Biochem. J. 2017, 474, 2641–2661. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Gupta, A.; Laxmi, A. Glucose Control of Root Growth Direction in Arabidopsis thaliana. J. Exp. Bot. 2014, 65, 2981–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLong, A.; Mockaitis, K.; Christensen, S. Protein phosphorylation in the delivery of and response to auxin signals. In Auxin Molecular Biology; Perrot-Rechenmann, C., Hagen, G., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 285–303. ISBN 978-94-010-0377-3. [Google Scholar]
- Adamowski, M.; Friml, J. PIN-Dependent Auxin Transport: Action, Regulation, and Evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Habets, M.E.J.; Offringa, R. PIN-Driven Polar Auxin Transport in Plant Developmental Plasticity: A Key Target for Environmental and Endogenous Signals. New Phytol. 2014, 203, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, X.; Li, B.; Chen, L.; Min, L.; Zhang, X. GhL1L1 Affects Cell Fate Specification by Regulating GhPIN1-Mediated Auxin Distribution. Plant Biotechnol. J. 2019, 17, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Michniewicz, M.; Brewer, P.B.; Friml, J. Polar Auxin Transport and Asymmetric Auxin Distribution. Arab. Book 2007, 5. [Google Scholar] [CrossRef] [Green Version]
- Omelyanchuk, N.A.; Kovrizhnykh, V.V.; Oshchepkova, E.A.; Pasternak, T.; Palme, K.; Mironova, V.V. A Detailed Expression Map of the PIN1 Auxin Transporter in Arabidopsis Thaliana Root. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Petrasek, J.; Friml, J. Auxin Transport Routes in Plant Development. Development 2009, 136, 2675–2688. [Google Scholar] [CrossRef] [Green Version]
- Zwiewka, M.; Bilanovičová, V.; Seifu, Y.W.; Nodzyński, T. The Nuts and Bolts of PIN Auxin Efflux Carriers. Front. Plant Sci. 2019, 10, 985. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Qin, Q.; Yan, J.; Niu, Y.; Huang, B.; Guan, L.; Li, Y.; Ren, D.; Li, J.; Hou, S. TYPE-ONE PROTEIN PHOSPHATASE4 Regulates Pavement Cell Interdigitation by Modulating PIN-FORMED1 Polarity and Trafficking in Arabidopsis. Plant Physiol. 2015, 167, 1058–1075. [Google Scholar] [CrossRef] [Green Version]
- Luschnig, C.; Vert, G. The Dynamics of Plant Plasma Membrane Proteins: PINs and Beyond. Development 2014, 141, 2924–2938. [Google Scholar] [CrossRef] [Green Version]
- Feraru, E.; Friml, J. PIN Polar Targeting. Plant Physiol. 2008, 147, 1553–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, K.; Goh, T.; Toyokura, K.; Kasahara, H.; Takebayashi, Y.; Mimura, T.; Kamiya, Y.; Fukaki, H. GNOM/FEWER ROOTS Is Required for the Establishment of an Auxin Response Maximum for Arabidopsis Lateral Root Initiation. Plant Cell Physiol. 2013, 54, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Nagawa, S.; Wang, G.; Yang, Z. Cell Polarity Signaling: Focus on Polar Auxin Transport. Mol. Plant 2008, 1, 899–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grones, P.; Abas, M.; Hajný, J.; Jones, A.; Waidmann, S.; Kleine-Vehn, J.; Friml, J. PID/WAG-Mediated Phosphorylation of the Arabidopsis PIN3 Auxin Transporter Mediates Polarity Switches during Gravitropism. Sci. Rep. 2018, 8, 10279. [Google Scholar] [CrossRef] [PubMed]
- Zažímalová, E.; Křeček, P.; Skůpa, P.; Hoyerová, K.; Petrášek, J. Polar Transport of the Plant Hormone Auxin—The Role of PIN-FORMED (PIN) Proteins. Cell. Mol. Life Sci. 2007, 64, 1621–1637. [Google Scholar] [CrossRef]
- Löfke, C.; Luschnig, C.; Kleine-Vehn, J. Posttranslational Modification and Trafficking of PIN Auxin Efflux Carriers. Mech. Dev. 2013, 130, 82–94. [Google Scholar] [CrossRef]
- Rahman, A.; Takahashi, M.; Shibasaki, K.; Wu, S.; Inaba, T.; Tsurumi, S.; Baskin, T.I. Gravitropism of Arabidopsis thaliana Roots Requires the Polarization of PIN2 toward the Root Tip in Meristematic Cortical Cells. Plant Cell 2010, 22, 1762–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlicht, M.; Strnad, M.; Scanlon, M.J.; Mancuso, S.; Hochholdinger, F.; Palme, K.; Volkmann, D.; Menzel, D.; Baluška, F. Auxin Immunolocalization Implicates Vesicular Neurotransmitter-Like Mode of Polar Auxin Transport in Root Apices. Plant Signal. Behav. 2006, 1, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, A.; Lee, S.-H.; Cho, H.-T. Functional Identification of the Phosphorylation Sites of Arabidopsis PIN-FORMED3 for Its Subcellular Localization and Biological Role. Plant J. 2012, 71, 810–823. [Google Scholar] [CrossRef]
- Ballesteros, I.; Domínguez, T.; Sauer, M.; Paredes, P.; Duprat, A.; Rojo, E.; Sanmartín, M.; Sánchez-Serrano, J.J. Specialized Functions of the PP2A Subfamily II Catalytic Subunits PP2A-C3 and PP2A-C4 in the Distribution of Auxin Fluxes and Development in Arabidopsis. Plant J. 2013, 73, 862–872. [Google Scholar] [CrossRef]
- Baba, A.I.; Andrási, N.; Valkai, I.; Gorcsa, T.; Koczka, L.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; Fehér, A.; Rigó, G.; et al. AtCRK5 Protein Kinase Exhibits a Regulatory Role in Hypocotyl Hook Development during Skotomorphogenesis. Int. J. Mol. Sci. 2019, 20, 3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigó, G.; Ayaydin, F.; Tietz, O.; Zsigmond, L.; Kovács, H.; Páy, A.; Salchert, K.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; et al. Inactivation of Plasma Membrane–Localized CDPK-RELATED KINASE5 Decelerates PIN2 Exocytosis and Root Gravitropic Response in Arabidopsis. Plant Cell 2013, 25, 1592–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleine-Vehn, J.; Huang, F.; Naramoto, S.; Zhang, J.; Michniewicz, M.; Offringa, R.; Friml, J. PIN Auxin Efflux Carrier Polarity Is Regulated by PINOID Kinase-Mediated Recruitment into GNOM-Independent Trafficking in Arabidopsis. Plant Cell 2009, 21, 3839–3849. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.-Y.; Xue, H.-W. Signals and Mechanisms Affecting Vesicular Trafficking during Root Growth. Curr. Opin. Plant Biol. 2011, 14, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Yang, X.; Michniewicz, M.; Weijers, D.; Quint, A.; Tietz, O.; Benjamins, R.; Ouwerkerk, P.B.F.; Ljung, K.; Sandberg, G.; et al. A PINOID-Dependent Binary Switch in Apical-Basal PIN Polar Targeting Directs Auxin Efflux. Science 2004, 306, 862–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michniewicz, M.; Zago, M.K.; Abas, L.; Weijers, D.; Schweighofer, A.; Meskiene, I.; Heisler, M.G.; Ohno, C.; Zhang, J.; Huang, F.; et al. Antagonistic Regulation of PIN Phosphorylation by PP2A and PINOID Directs Auxin Flux. Cell 2007, 130, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Zourelidou, M.; Absmanner, B.; Weller, B.; Barbosa, I.C.; Willige, B.C.; Fastner, A.; Streit, V.; Port, S.A.; Colcombet, J.; de la Fuente van Bentem, S.; et al. Auxin Efflux by PIN-FORMED Proteins Is Activated by Two Different Protein Kinases, D6 PROTEIN KINASE and PINOID. eLife 2014, 3, e02860. [Google Scholar] [CrossRef]
- Xi, W.; Gong, X.; Yang, Q.; Yu, H.; Liou, Y.-C. Pin1At Regulates PIN1 Polar Localization and Root Gravitropism. Nat. Commun. 2016, 7, 10430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, T. Cell Polarity and Tissue Patterning in Plants. Development 1991, 113, 83–93. [Google Scholar]
- Blakeslee, J.J.; Zhou, H.-W.; Heath, J.T.; Skottke, K.R.; Barrios, J.A.R.; Liu, S.-Y.; DeLong, A. Specificity of RCN1-Mediated Protein Phosphatase 2A Regulation in Meristem Organization and Stress Response in Roots. Plant Physiol. 2007, 146, 539–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.-B.; Chu, Y.-J.; Xue, H.-W. Phosphatidic Acid (PA) Binds PP2AA1 to Regulate PP2A Activity and PIN1 Polar Localization. Mol. Plant 2013, 6, 1692–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karampelias, M.; Neyt, P.; De Groeve, S.; Aesaert, S.; Coussens, G.; Rolčík, J.; Bruno, L.; De Winne, N.; Van Minnebruggen, A.; Van Montagu, M.; et al. ROTUNDA3 Function in Plant Development by Phosphatase 2A-Mediated Regulation of Auxin Transporter Recycling. Proc. Natl. Acad. Sci. USA 2016, 113, 2768–2773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillo, C.; Kataya, A.R.A.; Heidari, B.; Creighton, M.T.; Nemie-Feyissa, D.; Ginbot, Z.; Jonassen, E.M. Protein Phosphatases PP 2A, PP 4 and PP 6: Mediators and Regulators in Development and Responses to Environmental Cues. Plant Cell Environ. 2014, 37, 2631–2648. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Subunit Involved | Change of Protein Phosphatase Activity or Other PP2A Related Functions | Target Protein | Function | References |
|---|---|---|---|---|
| PP2A/C | activation | MAP65-1 MAP65-3? | Bundling of MTs, indirect regulation of vesicle traffic | [9,30] |
| PP2A/C | activation | AMPK, activation | Blocking of TORC pathway, triggering of autophagy | [56] |
| TAP46 subunit of PP2A | activation | proteins of the TORC pathway | Triggering of the TORC pathway and blocking of autophagy pathway | [4,51] |
| Methylation of PP2A/C by SBI1 | SBI1 targets PP2A to BRI1 containing endosomes | BRI1 | Dephosphorylation of excess BRI1 that will be targeted to lytic vacuoles | [4,62] |
| PP2A/B’, C | inhibition/deactivation | BZR1 | BZR1 remains phosphorylated, thus targeted to the autophagy pathway | [65,100] |
| PP2A/A, C3, C4, PP1 | activation | PINs, dephosphorylation | PIN recycling and proper membrane localization. Polar transport-tissue distribution of auxins | [69,70,76,87,98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Máthé, C.; M-Hamvas, M.; Freytag, C.; Garda, T. The Protein Phosphatase PP2A Plays Multiple Roles in Plant Development by Regulation of Vesicle Traffic—Facts and Questions. Int. J. Mol. Sci. 2021, 22, 975. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020975
Máthé C, M-Hamvas M, Freytag C, Garda T. The Protein Phosphatase PP2A Plays Multiple Roles in Plant Development by Regulation of Vesicle Traffic—Facts and Questions. International Journal of Molecular Sciences. 2021; 22(2):975. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020975
Chicago/Turabian StyleMáthé, Csaba, Márta M-Hamvas, Csongor Freytag, and Tamás Garda. 2021. "The Protein Phosphatase PP2A Plays Multiple Roles in Plant Development by Regulation of Vesicle Traffic—Facts and Questions" International Journal of Molecular Sciences 22, no. 2: 975. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22020975