
The Mandible Ameliorates Facial Allograft Rejection and Is Associated with the Development of Regulatory T Cells and Mixed Chimerism

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
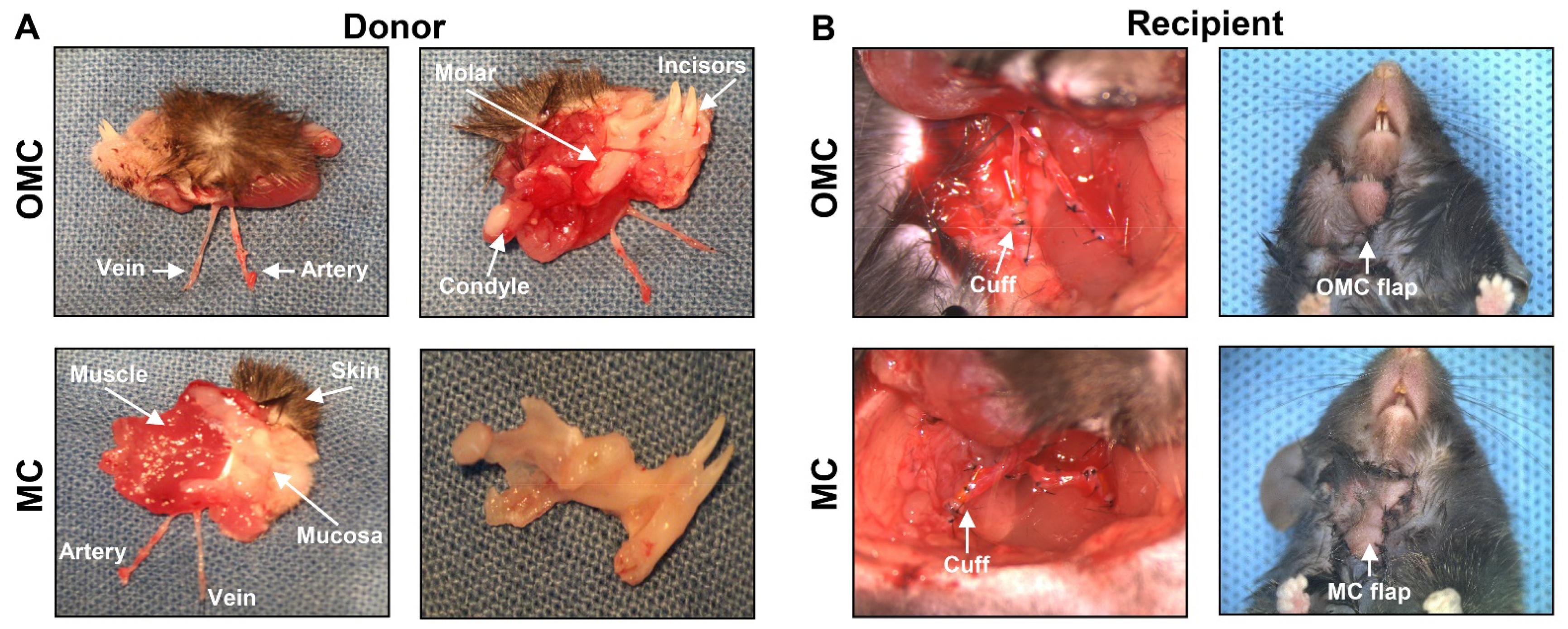
2.1. Surgery in the Facial Osteomyocutaneous and Myocutaneous Grafts
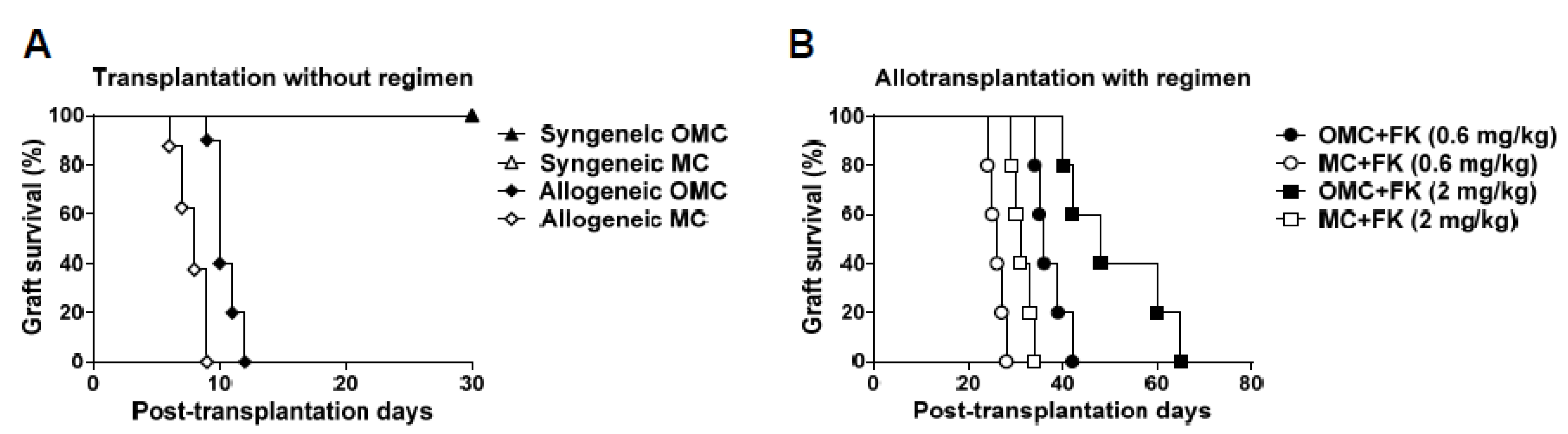
2.2. The Mandible Assists Facial Allograft Survival
2.3. Facial Allograft with the Mandible Exhibits Decreased Lymphocytic Infiltration
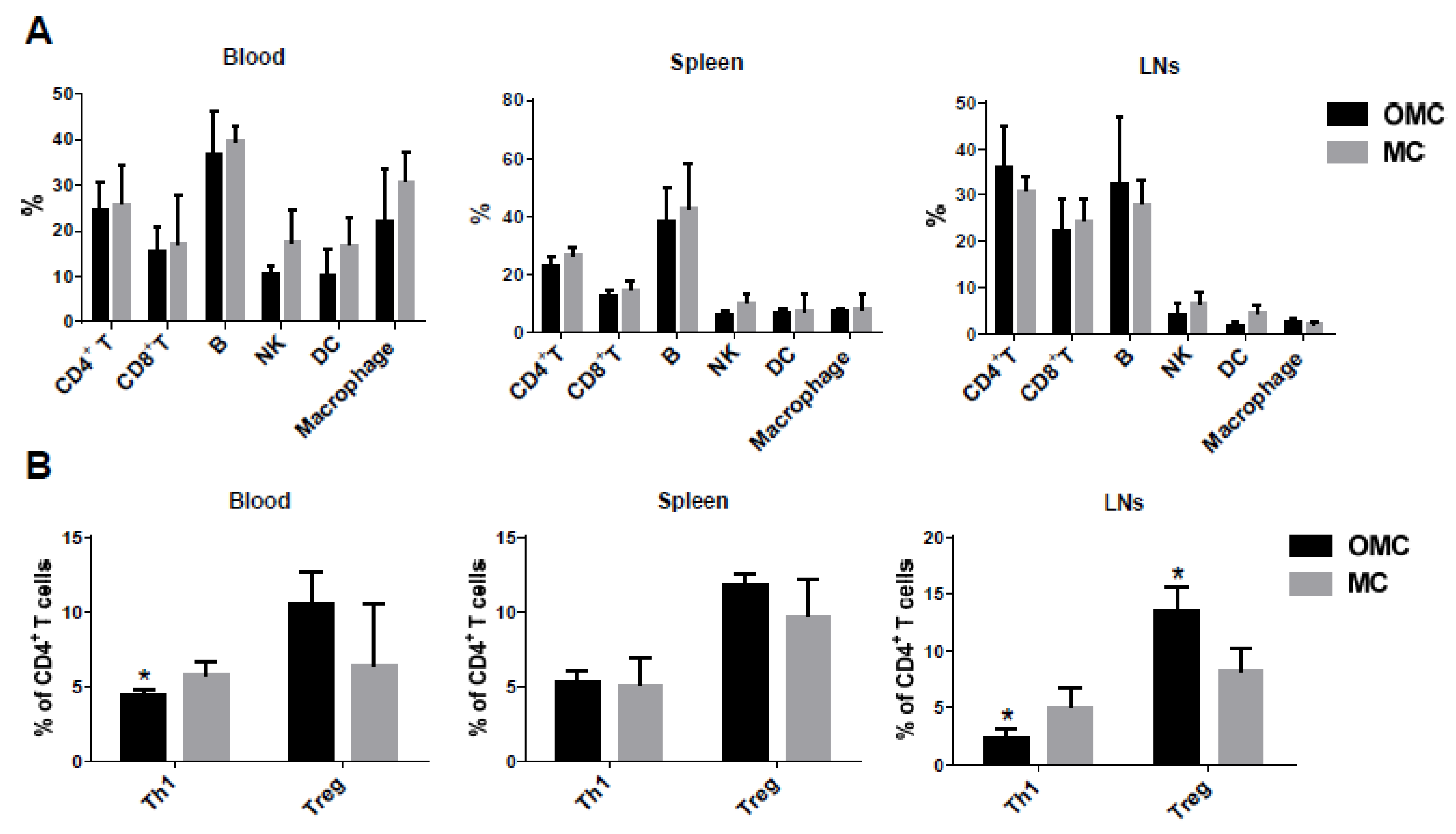
2.4. The Mandible Facilitates the Induction of Anti-Inflammatory Responses
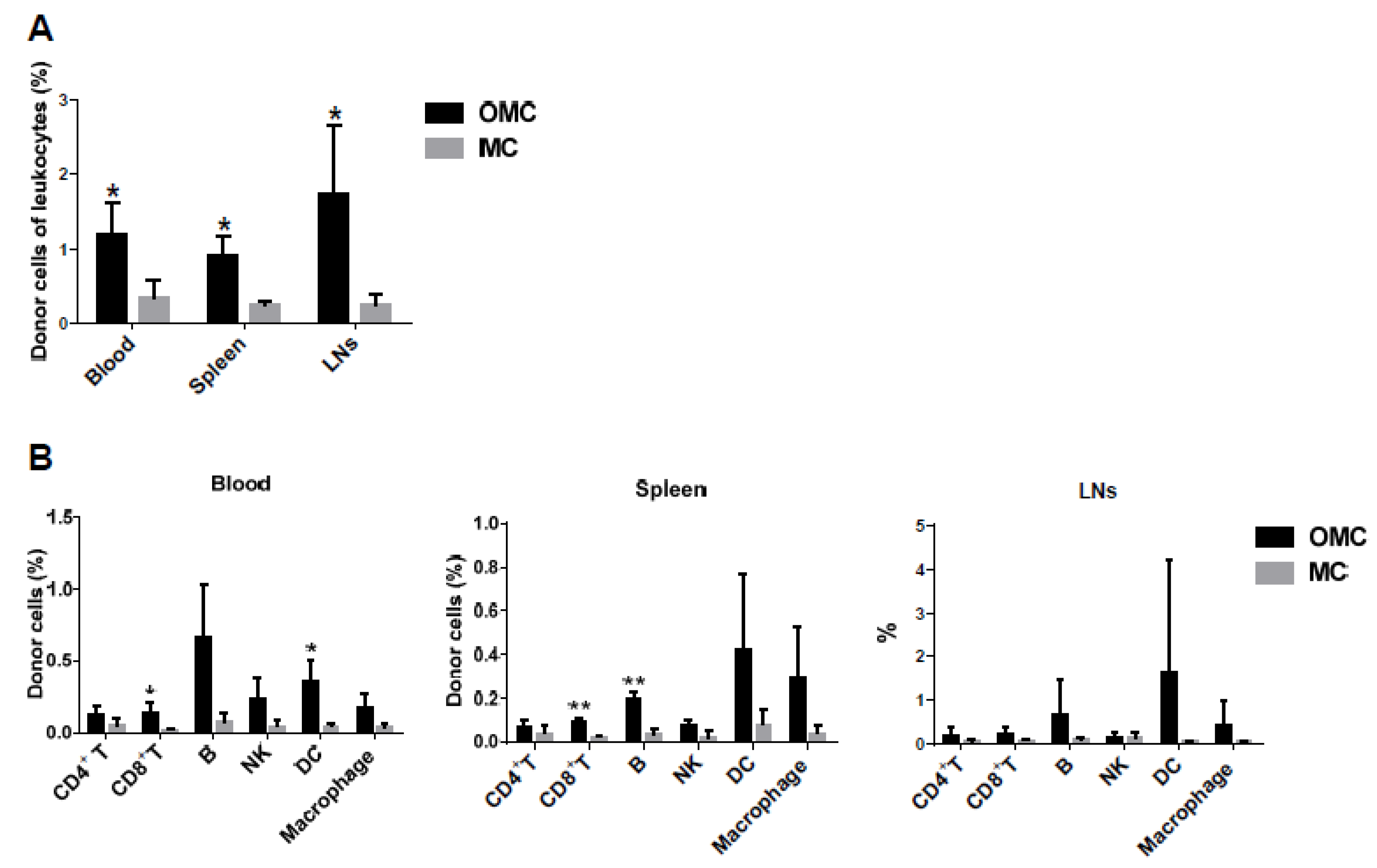
2.5. The Mandible Is Associated with the Induction of Mixed Chimerism
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Heterotopic Face Transplantation
4.2.1. Donor Surgery
4.2.2. Recipient Surgery
4.3. Postoperative Care
4.4. Histological Evaluation
4.5. Flow Cytometry Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Minervini, G.; Lucchese, A.; Perillo, L.; Serpico, R.; Minervini, G. Unilateral superior condylar neck fracture with dislocation in a child treated with an acrylic splint in the upper arch for functional repositioning of the mandible. Cranio 2017, 35, 337–341. [Google Scholar] [CrossRef]
- Antonelli, A.; Bennardo, F.; Brancaccio, Y.; Barone, S.; Femiano, F.; Nucci, L.; Minervini, G.; Fortunato, L.; Attanasio, F.; Giudice, A. Correction: Antonelli, A.; et al. Can bone compaction improve primary implant stability? An in vitro comparative study with osseodensification technique. Appl. Sci. 2020, 10, 8623. Appl. Sci. 2021, 11, 3427. [Google Scholar] [CrossRef]
- Makary, C.; Menhall, A.; Zammarie, C.; Lombardi, T.; Lee, S.Y.; Stacchi, C.; Park, K.B. Primary stability optimization by using fixtures with different thread depth according to bone density: A clinical prospective study on early loaded implants. Materials 2019, 12, 2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minervini, G.; Romano, A.; Petruzzi, M.; Maio, C.; Serpico, R.; Stasio, D.D.; Lucchese, A. Oral-facial-digital syndrome (OFD): 31-year follow-up management and monitoring. J. Biol. Regul. Homeost. Agents 2018, 32, 127–130. [Google Scholar] [PubMed]
- Boening, K.; Wieckiewicz, M.; Paradowska-Stolarz, A.; Wiland, P.; Shiau, Y.Y. Temporomandibular disorders and oral parafunctions: Mechanism, diagnostics, and therapy. Biomed. Res. Int. 2015, 2015, 354759. [Google Scholar] [CrossRef] [PubMed]
- Miernik, M.; Wieckiewicz, M.; Paradowska, A.; Wieckiewicz, W. Massage therapy in myofascial TMD pain management. Adv. Clin. Exp. Med. 2012, 21, 681–685. [Google Scholar] [PubMed]
- Moccia, S.; Nucci, L.; Spagnuolo, C.; d’Apuzzo, F.; Piancino, M.G.; Minervini, G. Polyphenols as potential agents in the management of temporomandibular disorders. Appl. Sci. 2020, 10, 5305. [Google Scholar] [CrossRef]
- d’Apuzzo, F.; Minervini, G.; Grassia, V.; Rotolo, R.P.; Perillo, L.; Nucci, L. Mandibular coronoid process hypertrophy: Diagnosis and 20-year follow-up with CBCT, MRI and EMG evaluations. Appl. Sci. 2021, 11, 4504. [Google Scholar] [CrossRef]
- Landin, L.; Bonastre, J.; Casado-Sanchez, C.; Diez, J.; Ninkovic, M.; Lanzetta, M.; Bene, M.D.; Schneeberger, S.; Hautz, T.; Lovic, A.; et al. Outcomes with respect to disabilities of the upper limb after hand allograft transplantation: A systematic review. Transpl. Int. 2012, 25, 424–432. [Google Scholar] [CrossRef]
- Lee, W.P. Hand transplantation: Evolution of a personal outlook. J. Hand. Surg. Am. 2017, 42, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Eun, S.C.; Kwon, S.T. Hand transplantation: Current status and immunologic obstacles. Exp. Clin. Transplant. 2019, 17, 97–104. [Google Scholar] [CrossRef]
- Shores, J.T.; Malek, V.; Lee, W.P.A.; Brandacher, G. Outcomes after hand and upper extremity transplantation. J. Mater. Sci. Mater. Med. 2017, 28, 72. [Google Scholar] [CrossRef]
- Wilks, D.J.; Clark, B.; Kay, S.P.J. The histocompatibility and immunogenetics of hand transplantation. Int. J. Immunogenet. 2020, 47, 24–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosin, M.; Rodriguez, E.D. The face transplantation update: 2016. Plast. Reconstr. Surg. 2016, 137, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Smeets, R.; Rendenbach, C.; Birkelbach, M.; Al-Dam, A.; Gröbe, A.; Hanken, H.; Heiland, M. Face transplantation: On the verge of becoming clinical routine? Biomed. Res. Int. 2014, 2014, 907272. [Google Scholar] [CrossRef] [PubMed]
- Kueckelhaus, M.; Lehnhardt, M.; Fischer, S.; Eriksson, E.; Pomahac, B.; Hirsch, T. Progress in face transplantation. Handchir. Mikrochir. Plast. Chir. 2014, 46, 206–213. [Google Scholar]
- Siemionow, M.; Gharb, B.B.; Rampazzo, A. Pathways of sensory recovery after face transplantation. Plast. Reconstr. Surg. 2011, 127, 1875–1889. [Google Scholar] [CrossRef] [PubMed]
- Kollar, B.; Pomahac, B. Facial restoration by transplantation. Surgeon 2018, 16, 245–249. [Google Scholar] [CrossRef]
- Aycart, M.A.; Kiwanuka, H.; Krezdorn, N.; Alhefzi, M.; Bueno, E.M.; Pomahac, B.; Oser, M.L. Quality of life after face transplantation: Outcomes, assessment tools, and future directions. Plast. Reconstr. Surg. 2017, 139, 194–203. [Google Scholar] [CrossRef]
- Siemionow, M.; Gharb, B.B.; Rampazzo, A. Successes and lessons learned after more than a decade of upper extremity and face transplantation. Curr. Opin. Organ Transplant. 2013, 18, 633–639. [Google Scholar] [CrossRef]
- Breidenbach, W.C.; Meister, E.A.; Becker, G.W.; Turker, T.; Gorantla, V.S.; Hassan, K.; Kaplan, B. A statistical comparative assessment of face and hand transplantation outcomes to determine whether either meets the standard of care threshold. Plast. Reconstr. Surg. 2016, 137, 214e–222e. [Google Scholar] [CrossRef]
- Siemionow, M. The miracle of face transplantation after 10 years. Br. Med. Bull. 2016, 120, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rifkin, W.J.; Manjunath, A.K.; Kantar, R.S.; Jacoby, A.; Kimberly, L.L.; Gelb, B.E.; Diaz-Siso, J.R.; Rodriguez, E.D. A comparison of immunosuppression regimens in hand, face, and kidney transplantation. J. Surg. Res. 2021, 258, 17–22. [Google Scholar] [CrossRef]
- Mills, E.C.; Alfonso, A.R.; Wolfe, E.M.; Park, J.J.; Sweeney, G.N.; Hoffman, A.F.; Felsenheld, J.H.; Sosin, M.; Ramly, E.P.; Rodriguez, E.D. Public Perceptions of cross-sex vascularized composite allotransplantation. Ann. Plast. Surg. 2020, 85, 685–690. [Google Scholar] [CrossRef]
- Diep, G.K.; Ramly, E.P.; Alfonso, A.R.; Berman, Z.P.; Rodriguez, E.D. Enhancing face transplant outcomes: Fundamental principles of facial allograft revision. Plast. Reconstr. Surg. Glob. Open 2020, 8, e2949. [Google Scholar] [PubMed]
- Wang, A.Y.L.; Loh, C.Y.Y. Reviewing immunosuppressive regimens in animal models for vascularized composite allotransplantation. Plast. Aesthet. Res. 2018, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Lao, W.W.; Wang, Y.L.; Ramirez, A.E.; Cheng, H.Y.; Wei, F.C. A new rat model for orthotopic abdominal wall allotransplantation. Plast. Reconstr. Surg. Glob. Open 2014, 2, e136. [Google Scholar] [CrossRef]
- Lee, W.P.; Yaremchuk, M.J.; Pan, Y.C.; Randolph, M.A.; Tan, C.M.; Weiland, A.J. Relative antigenicity of components of a vascularized limb allograft. Plast. Reconstr. Surg. 1991, 87, 401–411. [Google Scholar] [CrossRef]
- Murray, J.E. Organ transplantation (skin, kidney, heart) and the plastic surgeon. Plast. Reconstr. Surg. 1971, 47, 425–431. [Google Scholar] [CrossRef]
- Fryer, M.; Grahammer, J.; Khalifian, S.; Furtmüller, G.J.; Lee, W.P.A.; Raimondi, G.; Brandacher, G. Exploring cell-based tolerance strategies for hand and face transplantation. Expert. Rev. Clin. Immunol. 2015, 11, 1189–1204. [Google Scholar] [CrossRef]
- Leonard, D.A.; Gordon, C.R.; Sachs, D.H.; Cetrulo, C.L., Jr. Immunobiology of face transplantation. J. Craniofac. Surg. 2012, 23, 268–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, E.; Wang, A.Y.L.; Loh, C.Y.Y.; Chuang, S.H.; Lee, C.M.; ALDeek, N.; Lin, C.H.; Wei, F.C. A new face subunit transplant model in mice, containing skin, mandible, and oral mucosa for future face vascularized composite allotransplantation studies. Plast. Reconstr. Surg. 2019, 144, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Matsuzaka, T.; Uetani, M.; Hayashi, K.; Tsuji, Y.; Nakamura, T. Normal age-related conversion of bone marrow in the mandible: MR imaging findings. AJR Am. J. Roentgenol. 1995, 165, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, T.; Minami, M.; Ozawa, K.; Akimoto, Y.; Okada, H.; Yamamoto, H.; Suzuki, H.; Sasaki, Y. Magnetic resonance appearance of bone marrow in the mandible at different ages. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 1996, 82, 229–233. [Google Scholar] [CrossRef]
- Yamaza, T.; Ren, G.; Akiyama, K.; Chen, C.; Shi, Y.; Shi, S. Mouse mandible contains distinctive mesenchymal stem cells. J. Dent. Res. 2011, 90, 317–324. [Google Scholar] [CrossRef]
- Aghaloo, T.L.; Chaichanasakul, T.; Bezouglaia, O.; Kang, B.; Franco, R.; Dry, S.M.; Atti, E.; Tetradis, S. Osteogenic potential of mandibular vs. long-bone marrow stromal cells. J. Dent. Res. 2010, 89, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Rojas, M. Anti-inflammatory effects of mesenchymal stem cells: Novel concept for future therapies. Expert. Opin. Biol. Ther. 2008, 8, 569–581. [Google Scholar] [CrossRef]
- Pers, Y.M.; Ruiz, M.; Noël, D.; Jorgensen, C. Mesenchymal stem cells for the management of inflammation in osteoarthritis: State of the art and perspectives. Osteoarthr. Cartil. 2015, 23, 2027–2035. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by mesenchymal stem cells (MSCs): Mechanisms of action of living, apoptotic, and dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [Green Version]
- Kinsella, F.A.M.; Zuo, J.; Inman, C.F.; Pearce, H.; Maggs, L.; Eldershaw, S.E.; Chan, Y.L.T.; Nunnick, J.; Nagra, S.; Griffiths, M.; et al. Mixed chimerism established by hematopoietic stem cell transplantation is maintained by host and donor T regulatory cells. Blood Adv. 2019, 3, 734–743. [Google Scholar] [CrossRef]
- Fouzia, N.A.; Edison, E.S.; Lakshmi, K.M.; Korula, A.; Velayudhan, S.R.; Balasubramanian, P.; Abraham, A.; Viswabandya, A.; George, B.; Mathews, V.; et al. Long-term outcome of mixed chimerism after stem cell transplantation for thalassemia major conditioned with busulfan and cyclophosphamide. Bone Marrow Transplant. 2018, 53, 169–174. [Google Scholar] [CrossRef]
- Pathak, S.; Meyer, E.H. Tregs and mixed chimerism as approaches for tolerance induction in islet transplantation. Front. Immunol. 2020, 11, 612737. [Google Scholar] [CrossRef] [PubMed]
- Lejman, M.; Zaucha-Prażmo, A.; Zawitkowska, J.; Mroczkowska, A.; Grabowski, D.; Kowalczyk, J.R.; Drabko, K. Impact of early chimerism status on clinical outcome in children with acute lymphoblastic leukaemia after haematopoietic stem cell transplantation. BMC Cancer 2019, 19, 1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnani, A.; Pondarré, C.; Bouazza, N.; Magalon, J.; Miccio, A.; Six, E.; Roudaut, C.; Arnaud, C.; Kamdem, A.; Touzot, F.; et al. Extensive multilineage analysis in patients with mixed chimerism after allogeneic transplantation for sickle cell disease: Insight into hematopoiesis and engraftment thresholds for gene therapy. Haematologica 2020, 105, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Kako, S.; Yamazaki, H.; Ohashi, K.; Ozawa, Y.; Ota, S.; Kanda, Y.; Maeda, T.; Kato, J.; Ishiyama, K.; Matsuoka, K.I.; et al. Mixed chimerism and secondary graft failure in allogeneic hematopoietic stem cell transplantation for aplastic anemia. Biol. Blood Marrow Transplant. 2020, 26, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Sachs, D.H. Mixed chimerism as an approach to transplantation tolerance. Clin. Immunol. 2000, 95, S63–S68. [Google Scholar] [CrossRef]
- Pilat, N.; Wekerle, T. Transplantation tolerance through mixed chimerism. Nat. Rev. Nephrol. 2010, 6, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Sachs, D.H.; Sykes, M.; Kawai, T.; Cosimi, A.B. Immuno-intervention for the induction of transplantation tolerance through mixed chimerism. Semin. Immunol. 2011, 23, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Bedke, J.; Kiss, E.; Behnes, C.L.; Popovic, Z.V.; Heuser, M.; Stojanovic, T.; Sijmonsma, T.; Huber, P.; Domhan, S.; Muschal, S.; et al. Anti-inflammatory effects of alphav integrin antagonism in acute kidney allograft rejection. Am. J. Pathol. 2007, 171, 1127–1139. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.A. Anti-inflammatory agents: Present and future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef] [Green Version]
- Salari, V.; Mengoni, F.; Gallo, F.D.; Bertini, G.; Fabene, P.F. The Anti-inflammatory properties of mesenchymal stem cells in epilepsy: Possible treatments and future perspectives. Int. J. Mol. Sci. 2020, 21, 9683. [Google Scholar] [CrossRef]
- Oh, S.; Jang, A.Y.; Chae, S.; Choi, S.; Moon, J.; Kim, M.; Spiekerkoetter, E.; Zamanian, R.T.; Yang, P.C.; Hwang, D.; et al. Comparative analysis on the anti-inflammatory/immune effect of mesenchymal stem cell therapy for the treatment of pulmonary arterial hypertension. Sci. Rep. 2021, 11, 2012. [Google Scholar] [CrossRef]
- Noronha, N.D.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petryk, N.; Shevchenko, O. Mesenchymal stem cells anti-inflammatory activity in rats: Proinflammatory cytokines. J. Inflamm. Res. 2020, 13, 293–301. [Google Scholar] [CrossRef]
- Suzuki, K.; Chosa, N.; Sawada, S.; Takizawa, N.; Yaegashi, T.; Ishisaki, A. Enhancement of anti-Inflammatory and osteogenic abilities of mesenchymal stem cells via cell-to-cell adhesion to periodontal ligament-derived fibroblasts. Stem Cells Int. 2017, 2017, 3296498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, S.E.; Luger, D.; Lipinski, M.J. Paracrine-mediated systemic anti-inflammatory activity of intravenously administered mesenchymal stem cells: A transformative strategy for cardiac stem cell therapeutics. Circ. Res. 2017, 121, 1044–1046. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, M.; Zhao, R.; Zhao, C.; Liu, Y.; Zou, H.; Chen, L.; Guan, Y.; Zhang, Y.A. Intravenously delivered allogeneic mesenchymal stem cells bidirectionally regulate inflammation and induce neurotrophic effects in distal middle cerebral artery occlusion rats within the first 7 days after stroke. Cell Physiol. Biochem. 2018, 46, 1951–1970. [Google Scholar] [CrossRef]
- Wang, J.; Ren, H.; Yuan, X.; Ma, H.; Shi, X.; Ding, Y. Interleukin-10 secreted by mesenchymal stem cells attenuates acute liver failure through inhibiting pyroptosis. Hepatol. Res. 2018, 48, E194–E202. [Google Scholar] [CrossRef] [Green Version]
- Sucher, R.; Lin, C.H.; Zanoun, R.; Atsina, K.K.; Weinstock, M.; Pulikkotil, B.; Schneeberger, S.; Zheng, X.X.; Pratschke, J.; Lee, W.P.A.; et al. Mouse hind limb transplantation: A new composite tissue allotransplantation model using nonsuture supermicrosurgery. Transplantation 2010, 90, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Sucher, R.; Shih, Y.C.; Ng, T.W.; Mao, Q.; Lin, C.H.; Lee, W.P.A.; Brandacher, G. The neck as a preferred recipient site for vascularized composite allotransplantation in the mouse. Plast. Reconstr. Surg. 2014, 133, 133e–141e. [Google Scholar] [CrossRef]
- Wu, C.; Xue, Y.; Wang, P.; Lin, L.; Liu, Q.; Li, N.; Xu, J.; Cao, X. IFN-gamma primes macrophage activation by increasing phosphatase and tensin homolog via downregulation of miR-3473b. J. Immunol. 2014, 193, 3036–3044. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Fan, H.; Jiang, S. CD4(+) T-cell subsets in transplantation. Immunol. Rev. 2013, 252, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Brustle, K.; Jungraithmayr, W. T helper cell subsets in experimental lung allograft rejection. J. Surg. Res. 2019, 233, 74–81. [Google Scholar] [CrossRef]
- Akintoye, S.O.; Lam, T.; Shi, S.; Brahim, J.; Collins, M.T.; Robey, P.G. Skeletal site-specific characterization of orofacial and iliac crest human bone marrow stromal cells in same individuals. Bone 2006, 38, 758–768. [Google Scholar] [CrossRef]
- Damek-Poprawa, M.; Stefanik, D.; Levin, L.M.; Akintoye, S.O. Human bone marrow stromal cells display variable anatomic site-dependent response and recovery from irradiation. Arch. Oral Biol. 2010, 55, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Chen, M.; Yang, G.; Xiang, L.; He, L.; Hei, T.K.; Chotkowski, G.; Tarnow, D.P.; Finkel, M.; Ding, L.; et al. Hematopoietic stem cells in neural-crest derived bone marrow. Sci. Rep. 2016, 6, 36411. [Google Scholar] [CrossRef] [Green Version]
- Marini, M.; Bertolai, R.; Ambrosini, S.; Sarchielli, E.; Vannelli, G.B.; Sgambati, E. Differential expression of vascular endothelial growth factor in human fetal skeletal site-specific tissues: Mandible versus femur. Acta. Histochem. 2015, 117, 228–234. [Google Scholar] [CrossRef]
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2008, 2, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Anggelia, M.R.; Cheng, H.Y.; Wang, A.Y.L.; Chuang, W.Y.; Lin, C.H.; Lee, W.P.A.; Wei, F.C.; Brandacher, G. The intragraft vascularized bone marrow component plays a critical role in tolerance induction after reconstructive transplantation. Cell Mol. Immunol. 2021, 18, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Anggelia, M.R.; Cheng, H.Y.; Chuang, W.Y.; Hsieh, Y.H.; Wang, A.Y.L.; Lin, C.H.; Wei, F.C.; Brandacher, G.; Lin, C.H. Unraveling the crucial roles of FoxP3+ regulatory T cells in vascularized composite allograft tolerance induction and maintenance. Transplantation 2021, 105, 1238–1249. [Google Scholar] [CrossRef]
- Kulahci, Y.; Klimczak, A.; Madajka, M.; Altuntas, S.; Siemionow, M. Long-term survival of composite hemiface/mandible/tongue allografts correlates with multilineage chimerism development in the lymphoid and myeloid compartments of recipients. Transplantation 2010, 90, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.; Lee, S.J.; Lee, J.H.; Kim, K.H.; Suh, D.S.; Kwon, B.S.; Choi, K.U. Stromal tumor-infiltrating lymphocytes evaluated on H&E-stained slides are an independent prognostic factor in epithelial ovarian cancer and ovarian serous carcinoma. Oncol. Lett. 2019, 17, 4557–4565. [Google Scholar]
- Kleczek, P.; Jaworek-Korjakowska, J.; Gorgon, M. A novel method for tissue segmentation in high-resolution H&E-stained histopathological whole-slide images. Comput. Med. Imaging Graph. 2020, 79, 101686. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Paz, D.; Aviña, A.E.; Cardona, E.; Lee, C.-M.; Lin, C.-H.; Lin, C.-H.; Wei, F.-C.; Wang, A.Y.L. The Mandible Ameliorates Facial Allograft Rejection and Is Associated with the Development of Regulatory T Cells and Mixed Chimerism. Int. J. Mol. Sci. 2021, 22, 11104. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011104
De Paz D, Aviña AE, Cardona E, Lee C-M, Lin C-H, Lin C-H, Wei F-C, Wang AYL. The Mandible Ameliorates Facial Allograft Rejection and Is Associated with the Development of Regulatory T Cells and Mixed Chimerism. International Journal of Molecular Sciences. 2021; 22(20):11104. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011104
Chicago/Turabian StyleDe Paz, Dante, Ana Elena Aviña, Esteban Cardona, Chin-Ming Lee, Chia-Hsien Lin, Cheng-Hung Lin, Fu-Chan Wei, and Aline Yen Ling Wang. 2021. "The Mandible Ameliorates Facial Allograft Rejection and Is Associated with the Development of Regulatory T Cells and Mixed Chimerism" International Journal of Molecular Sciences 22, no. 20: 11104. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011104