
Photophysical Properties of BADAN Revealed in the Study of GGBP Structural Transitions

 ,
,  , , , , and
, , , , and 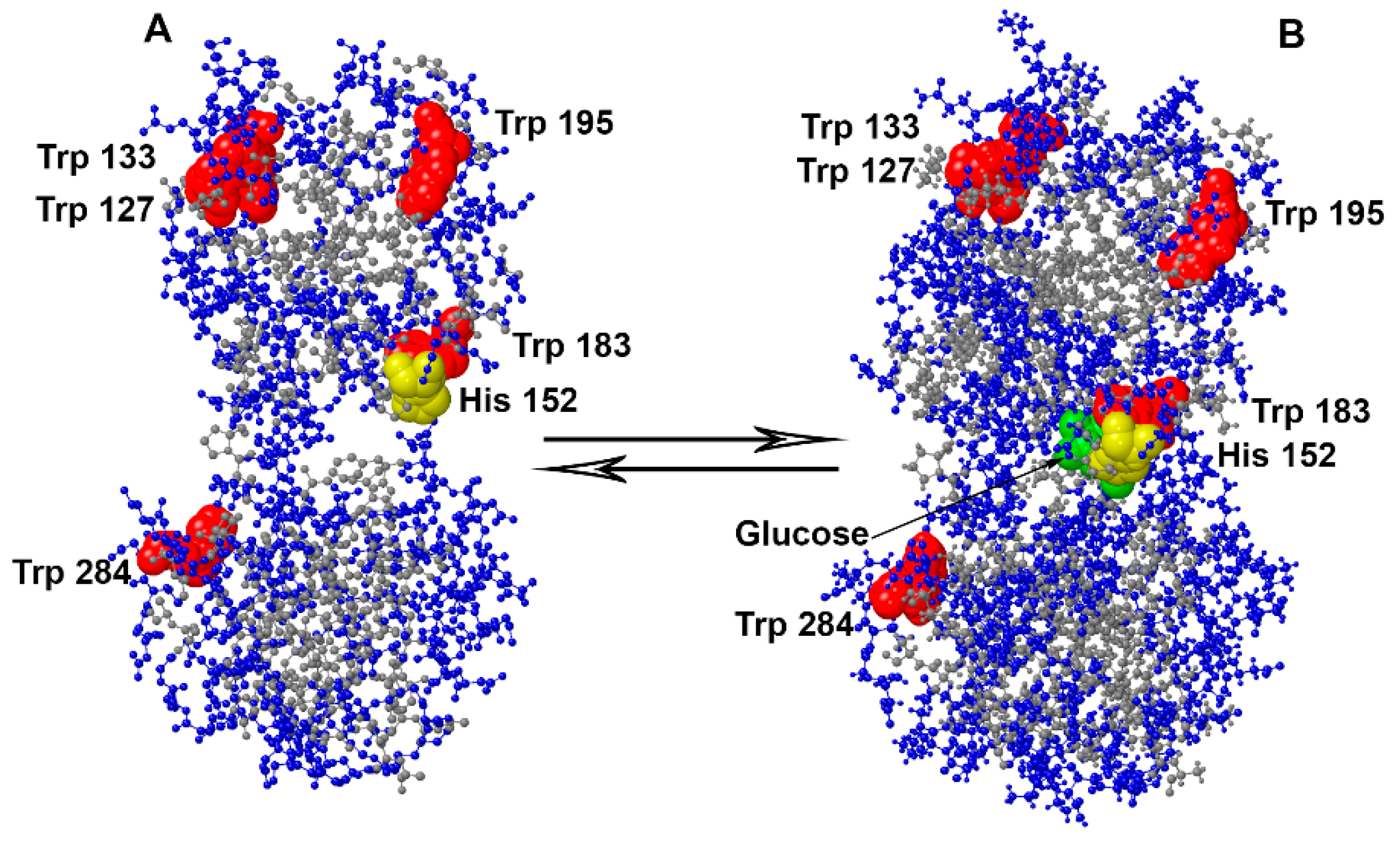{kind=link}
{kind=link}
{kind=link}
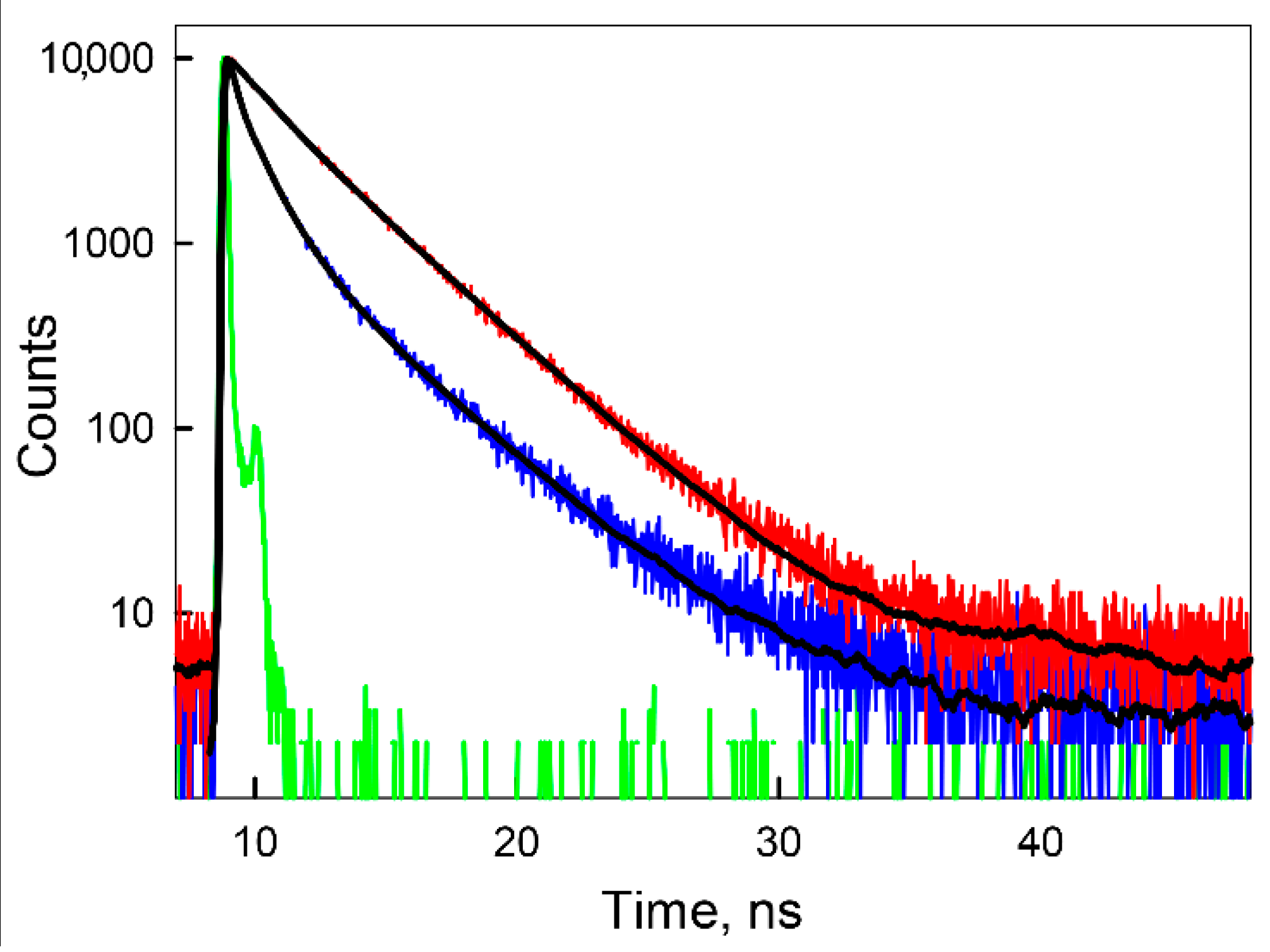
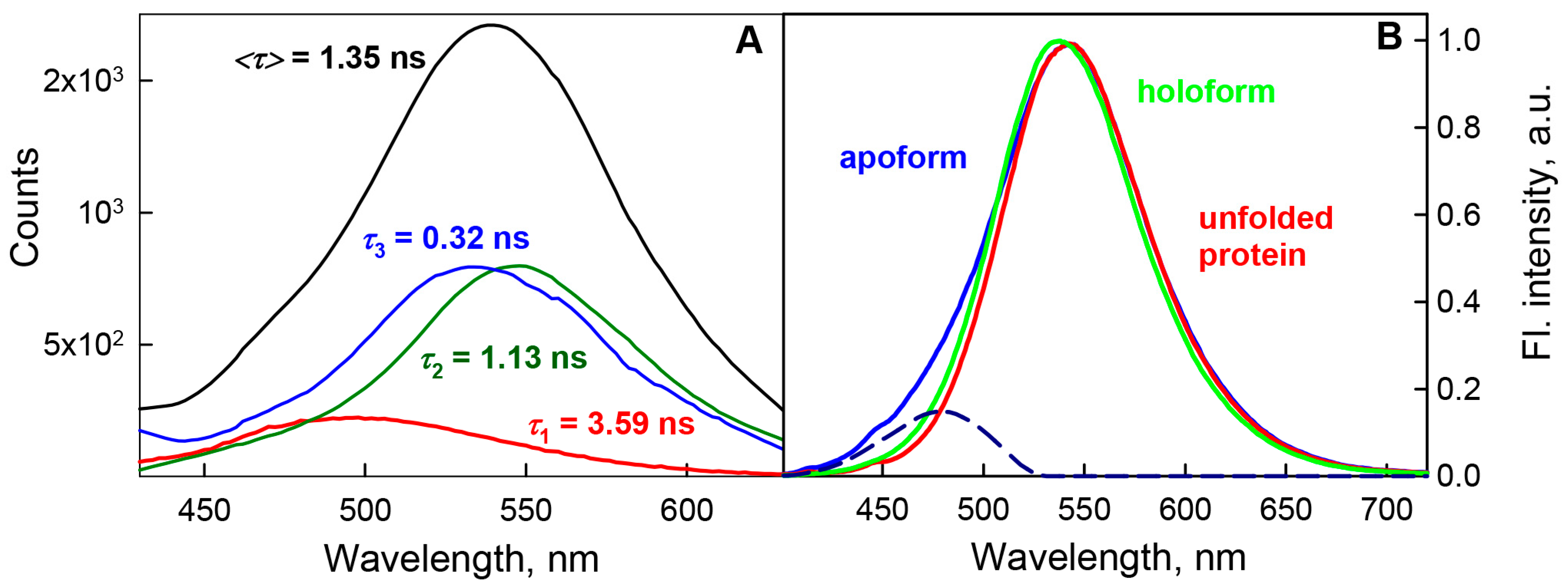{kind=link}
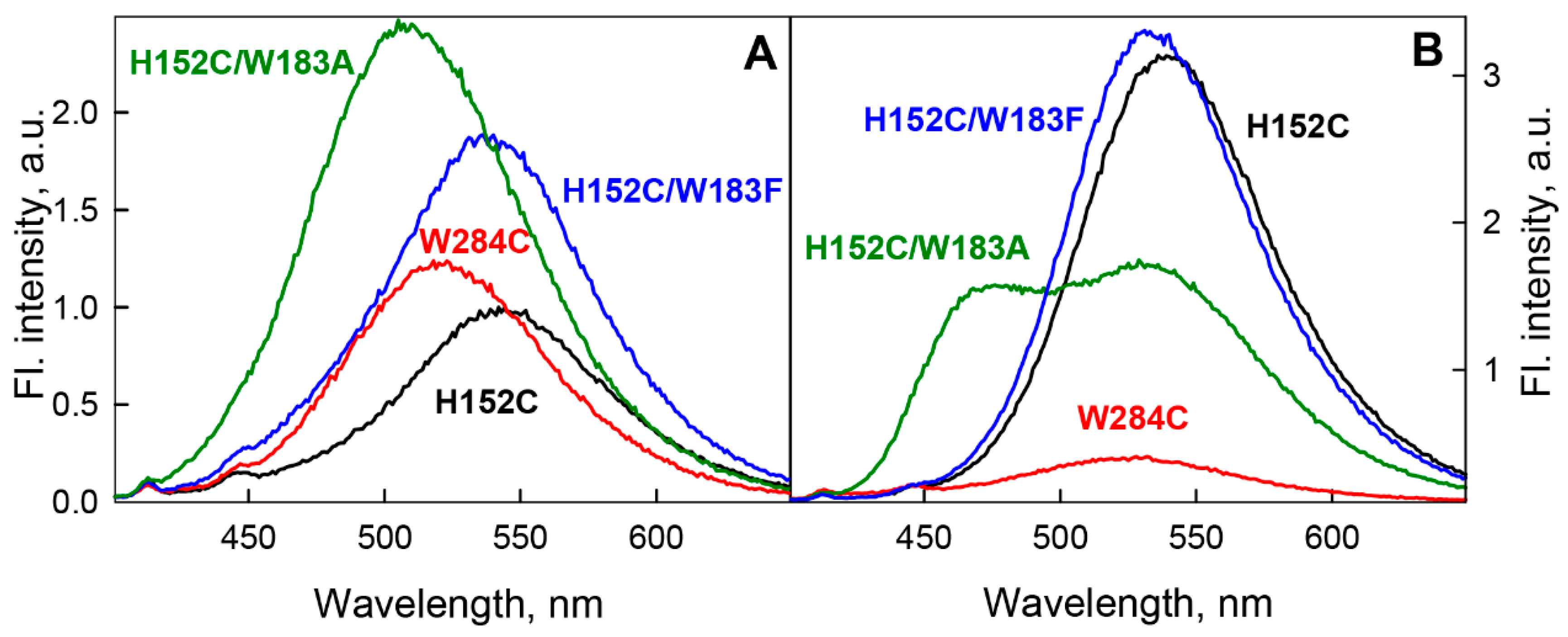{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Spectral Characteristics of BADAN Associated with Various Structural States of GGBP Variants
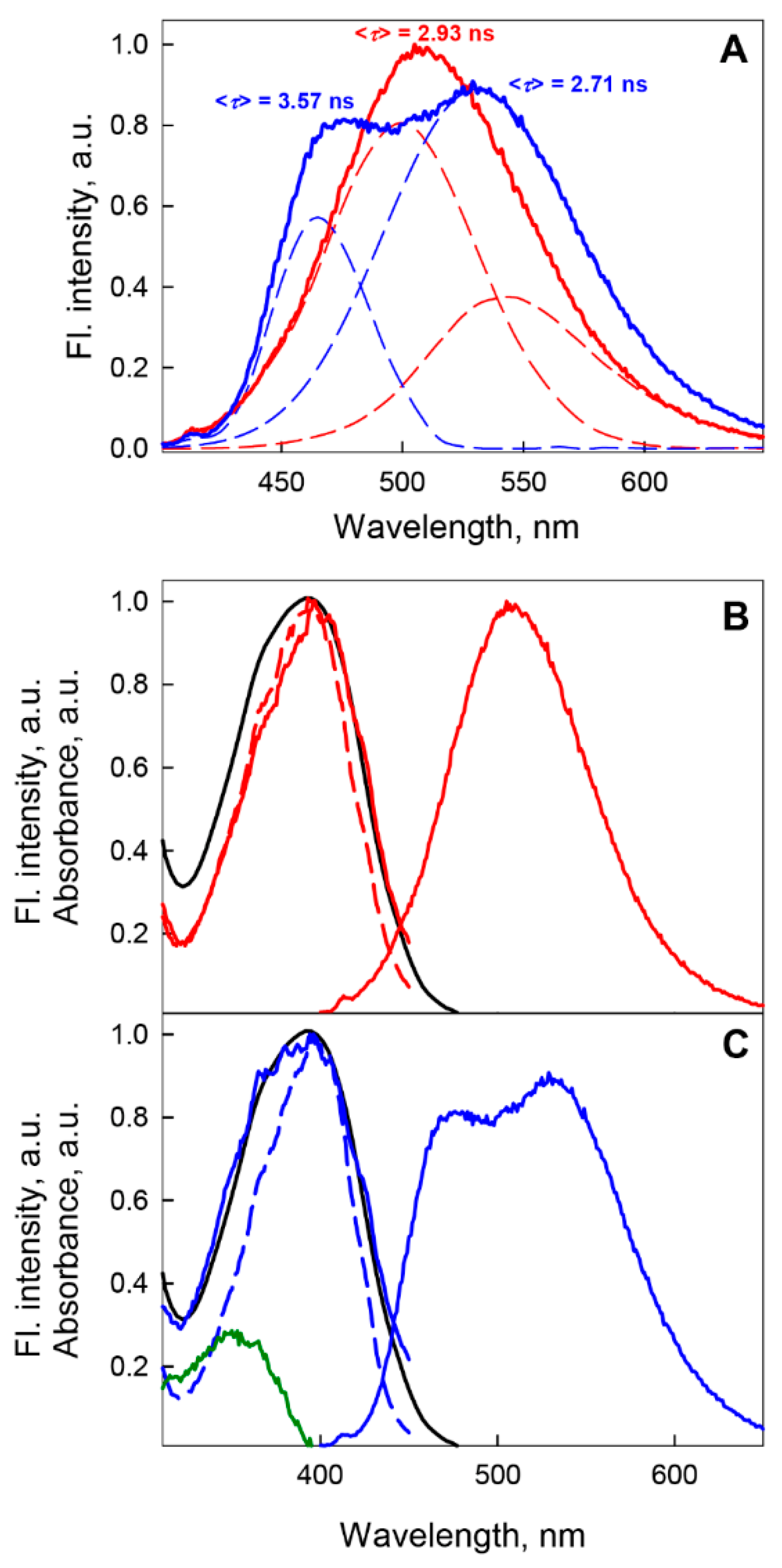
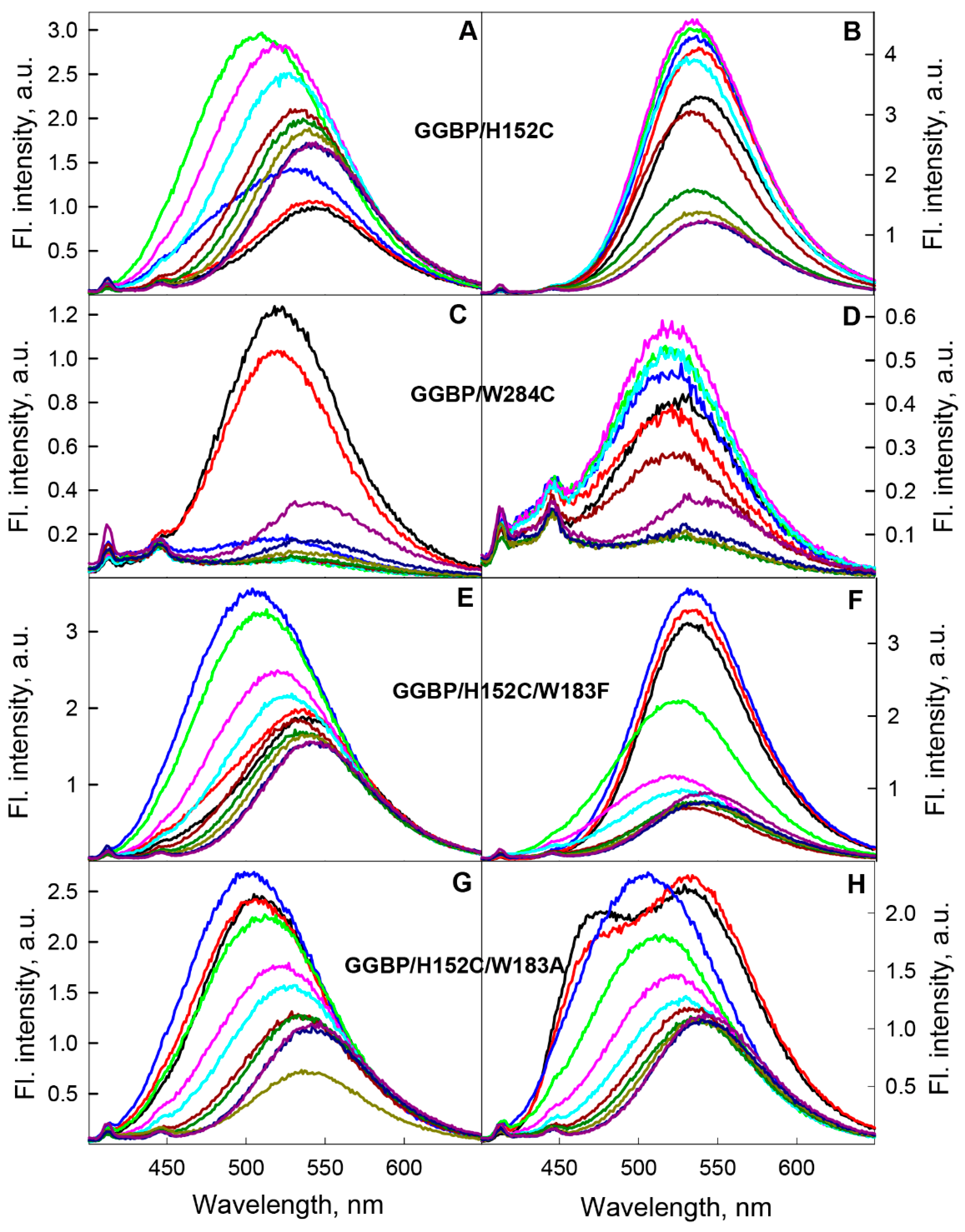
2.1.1. GGBP/H152C Apo- and Holoforms
2.1.2. GGBP/W284C Apo- and Holoforms
2.1.3. GGBP/H152C/W183F and GGBP/H152C/W183A Apo- and Holoforms
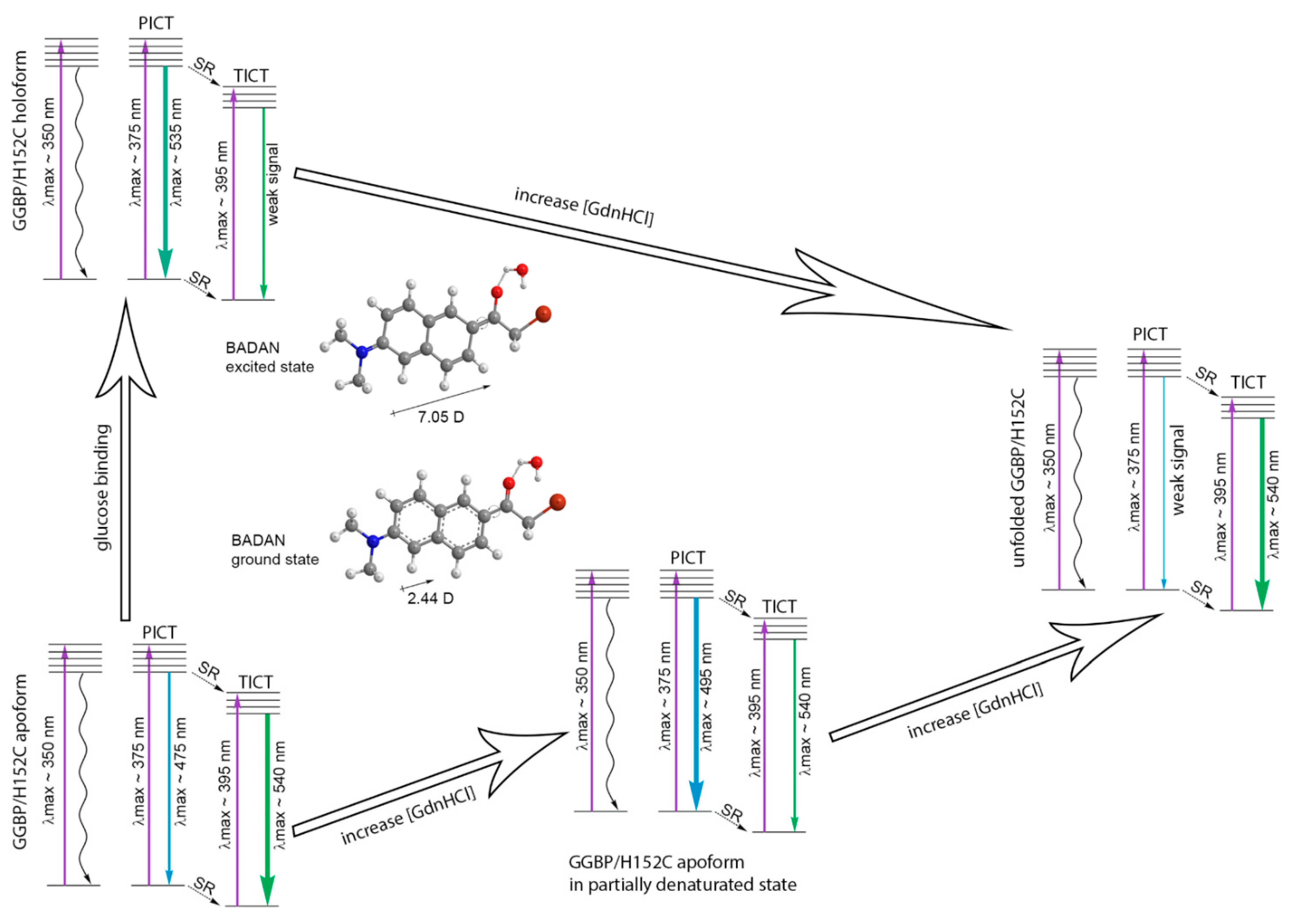
2.2. C. Hanging of BADAN Fluorescent Characteristics with Unfolding of GGBP Mutants
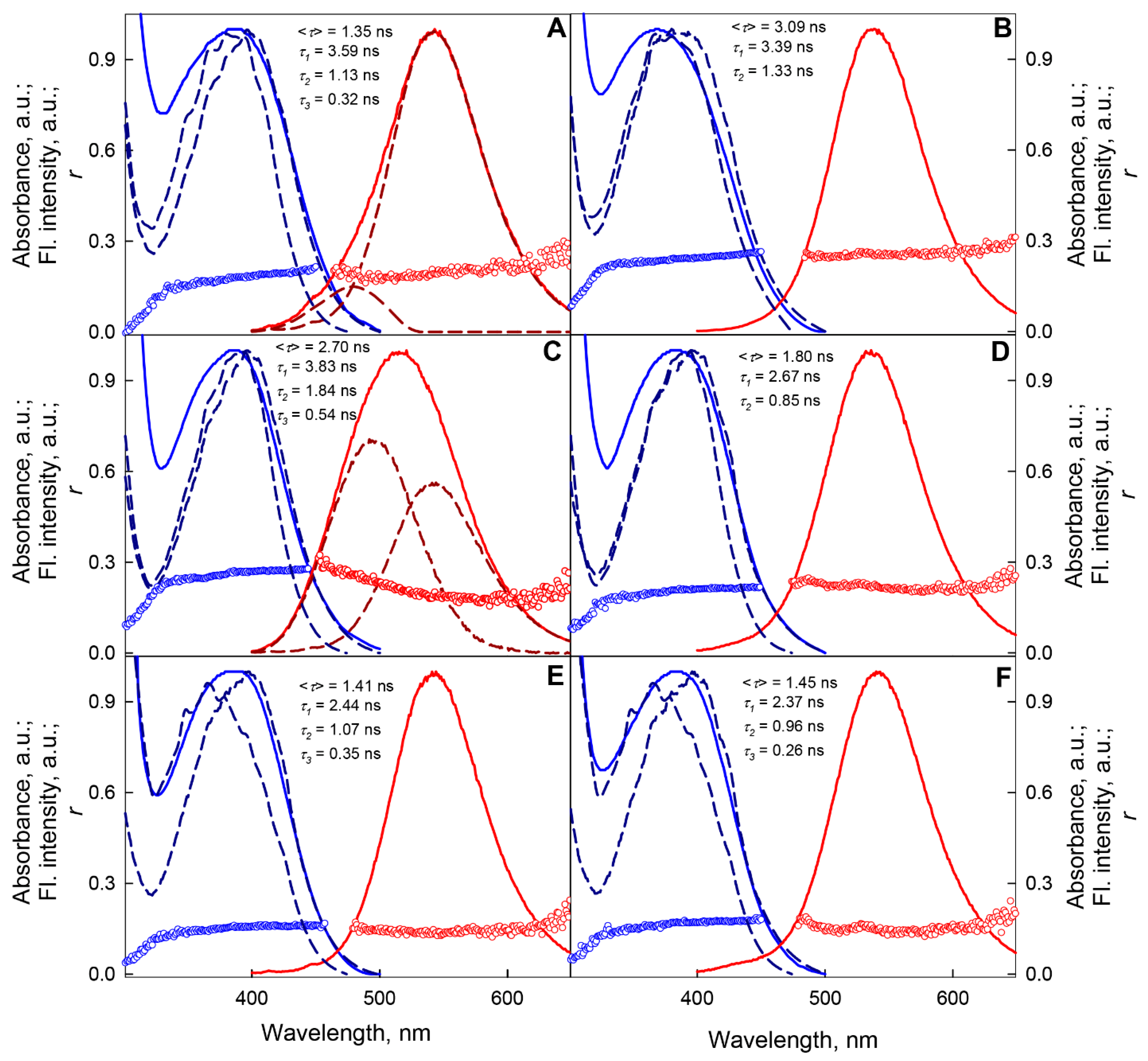
2.2.1. Unfolding of GGBP/H152C in Ligand-Free and Ligand-Bound States
2.2.2. Unfolding of GGBP/H152C in Ligand-Free and Ligand-Bound States
2.2.3. Unfolding of GGBP/W284C in Ligand-Free and Ligand-Bound States
2.2.4. Unfolding of GGBP/H152C/W183F and GGBP/H152C/W183A Mutant Forms in Ligand-Free and Ligand-Bound States
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Steady-State Fluorescence Spectroscopy
4.2.2. Time-Resolved Fluorescence Spectroscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Adhikary, R.; Barnes, C.A.; Petrich, J.W. Solvation dynamics of the fluorescent probe PRODAN in heterogeneous environments: Contributions from the locally excited and charge-transferred states. J. Phys. Chem. B 2009, 113, 11999–12004. [Google Scholar] [CrossRef]
- Alty, I.G.; Cheek, D.W.; Chen, T.; Smith, D.B.; Walhout, E.Q.; Abelt, C.J. Intramolecular Hydrogen-Bonding Effects on the Fluorescence of PRODAN Derivatives. J. Phys. Chem. A 2016, 120, 3518–3523. [Google Scholar] [CrossRef]
- Artukhov, V.Y.; Zharkova, O.M.; Morozova, J.P. Features of absorption and fluorescence spectra of prodan. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2007, 68, 36–42. [Google Scholar] [CrossRef]
- Artyukhov, V.Y.; Zharkova, O.M.; Morozova, Y.P. Complexing and photoprocesses in a PRODAN molecule. Russ. Phys. J. 2004, 47, 1172–1177. [Google Scholar] [CrossRef]
- Bakalova, S.M.; Kaneti, J. Solvent induced shifts of electronic spectra IV. Computational study on PRODAN fluorescence and implications to the excited state structure. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2009, 72, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Barucha-Kraszewska, J.; Kraszewski, S.; Jurkiewicz, P.; Ramseyer, C.; Hof, M. Numerical studies of the membrane fluorescent dyes dynamics in ground and excited states. Biochim. Biophys. Acta 2010, 1798, 1724–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, B.N.; Abelt, C.J. Synthesis and photophysical properties of models for twisted PRODAN and dimethylaminonaphthonitrile. J. Phys. Chem. A 2005, 109, 1295–1298. [Google Scholar] [CrossRef] [PubMed]
- Ilich, P.; Prendergast, F.G. Singlet adiabatic states of solvated PRODAN: A semiempirical molecular orbital study. J. Phys. Chem. 1989, 93, 4441–4447. [Google Scholar] [CrossRef]
- Lobo, B.C.; Abelt, C.J. Does PRODAN Possess a Planar or Twisted Charge-Transfer Excited State? Photophysical Properties of Two PRODAN Derivatives. J. Phys. Chem. A 2003, 107, 10938–10943. [Google Scholar] [CrossRef]
- Nikitina, Y.Y.; Iqbal, E.S.; Yoon, H.J.; Abelt, C.J. Preferential solvation in carbonyl-twisted PRODAN derivatives. J. Phys. Chem. A 2013, 117, 9189–9195. [Google Scholar] [CrossRef] [PubMed]
- Parusel, A.B.J.; Schneider, F.W.; Köhler, G. An ab initio study on excited and ground state properties of the organic fluorescence probe PRODAN. J. Mol. Struct. 1997, 398, 341–346. [Google Scholar] [CrossRef]
- Pederzoli, M.; Sobek, L.; Brabec, J.; Kowalski, K.; Cwiklik, L.; Pittner, J. Fluorescence of PRODAN in water: A computational QM/MM MD study. Chem. Phys. Lett. 2014, 597, 57–62. [Google Scholar] [CrossRef]
- Rowe, B.A.; Roach, C.A.; Lin, J.; Asiago, V.; Dmitrenko, O.; Neal, S.L. Spectral heterogeneity of PRODAN fluorescence in isotropic solvents revealed by multivariate photokinetic analysis. J. Phys. Chem. A 2008, 112, 13402–13412. [Google Scholar] [CrossRef]
- Samanta, A.; Fessenden, R.W. Excited State Dipole Moment of PRODAN as Determined from Transient Dielectric Loss Measurements. J. Phys. Chem. A 2000, 104, 8972–8975. [Google Scholar] [CrossRef]
- Titova, T.Y.; Morozova, Y.P.; Korolev, B.V. Photophysics of a Fluorescent Probe of the Laurdan Molecule in Homogeneous and Binary Solvents. Russ. Phys. J. 2016, 59, 672–678. [Google Scholar] [CrossRef]
- Tomin, V.I.; Heldt, J. The Red-Edge Effects in Laurdan Solutions. Z. Nat. A 2003, 58, 109. [Google Scholar] [CrossRef]
- Zharkova, O.M.; Rakhimov, S.I.; Morozova, Y.P. Quantum-chemical investigation of the spectral properties of fluorescent probes based on naphthalene derivatives (prodan, promen). Russ. Phys. J. 2013, 56, 411–419. [Google Scholar] [CrossRef]
- Titova, T.Y.; Morozova, Y.P.; Zharkova, O.M.; Artyukhov, V.Y.; Korolev, B.V. Quantum-chemical investigations of spectroscopic properties of a fluorescence probe. Russ. Phys. J. 2012, 55, 369–377. [Google Scholar] [CrossRef]
- Nemkovich, N.A.; Detert, H.; Roeder, N. Electrooptical Absorption Measurements (EOAM) Testify Existence of two Conformers of Prodan and Laurdan with Different Dipole Moments in Equilibrium Ground and Franck-Condon Excited State. J. Fluoresc. 2016, 26, 1563–1572. [Google Scholar] [CrossRef]
- Koehorst, R.B.; Laptenok, S.; van Oort, B.; van Hoek, A.; Spruijt, R.B.; van Stokkum, I.H.; van Amerongen, H.; Hemminga, M.A. Profiling of dynamics in protein-lipid-water systems: A time-resolved fluorescence study of a model membrane protein with the label BADAN at specific membrane depths. Eur. Biophys. J. 2010, 39, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Koehorst, R.B.; Spruijt, R.B.; Hemminga, M.A. Site-directed fluorescence labeling of a membrane protein with BADAN: Probing protein topology and local environment. Biophys. J. 2008, 94, 3945–3955. [Google Scholar] [CrossRef] [Green Version]
- Kawski, A.; Kuklinski, B.; Bojarski, P. Ground and excited state dipole moments of BADAN and ACRYLODAN determined from solvatochromic shifts of absorption and fluorescence spectra. Z. Nat. A 2001, 56, 407–411. [Google Scholar] [CrossRef]
- Kawski, A.; Kuklinski, B.; Bojarski, P. Thermochromic absorption, fluorescence band shifts and dipole moments of BADAN and ACRYLODAN. Z. Nat. A 2002, 57, 716–722. [Google Scholar] [CrossRef]
- Owenius, R.; Osterlund, M.; Lindgren, M.; Svensson, M.; Olsen, O.H.; Persson, E.; Freskgard, P.O.; Carlsson, U. Properties of spin and fluorescent labels at a receptor-ligand interface. Biophys. J. 1999, 77, 2237–2250. [Google Scholar] [CrossRef] [Green Version]
- Khan, F.; Gnudi, L.; Pickup, J.C. Fluorescence-based sensing of glucose using engineered glucose/galactose-binding protein: A comparison of fluorescence resonance energy transfer and environmentally sensitive dye labelling strategies. Biochem. Biophys. Res. Commun. 2008, 365, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Saxl, T.E.; Pickup, J.C. Fluorescence intensity- and lifetime-based glucose sensing using an engineered high-Kd mutant of glucose/galactose-binding protein. Anal. Biochem. 2010, 399, 39–43. [Google Scholar] [CrossRef]
- Pickup, J.C.; Khan, F.; Zhi, Z.L.; Coulter, J.; Birch, D.J. Fluorescence intensity- and lifetime-based glucose sensing using glucose/galactose-binding protein. J. Diabetes Sci. Technol. 2013, 7, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxl, T.; Khan, F.; Ferla, M.; Birch, D.; Pickup, J. A fluorescence lifetime-based fibre-optic glucose sensor using glucose/galactose-binding protein. Analyst 2011, 136, 968–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxl, T.; Khan, F.; Matthews, D.R.; Zhi, Z.L.; Rolinski, O.; Ameer-Beg, S.; Pickup, J. Fluorescence lifetime spectroscopy and imaging of nano-engineered glucose sensor microcapsules based on glucose/galactose-binding protein. Biosens. Bioelectron. 2009, 24, 3229–3234. [Google Scholar] [CrossRef]
- Fonin, A.V.; Povarova, O.I.; Staiano, M.; D’Auria, S.; Turoverov, K.K.; Kuznetsova, I.M. The trehalose/maltose-binding protein as the sensitive element of a glucose biosensor. Opt. Mater. 2014, 36, 1676–1679. [Google Scholar] [CrossRef]
- Vyas, N.K.; Vyas, M.N.; Quiocho, F.A. Sugar and signal-transducer binding sites of the Escherichia coli galactose chemoreceptor protein. Science 1988, 242, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Aqvist, J.; Mowbray, S.L. Sugar recognition by a glucose/galactose receptor. Evaluation of binding energetics from molecular dynamics simulations. J. Biol. Chem. 1995, 270, 9978–9981. [Google Scholar]
- Borrok, M.J.; Kiessling, L.L.; Forest, K.T. Conformational changes of glucose/galactose-binding protein illuminated by open, unliganded, and ultra-high-resolution ligand-bound structures. Protein Sci. 2007, 16, 1032–1041. [Google Scholar] [CrossRef]
- Ortega, G.; Castano, D.; Diercks, T.; Millet, O. Carbohydrate affinity for the glucose-galactose binding protein is regulated by allosteric domain motions. J. Am. Chem. Soc. 2012, 134, 19869–19876. [Google Scholar] [CrossRef]
- Unione, L.; Ortega, G.; Mallagaray, A.; Corzana, F.; Perez-Castells, J.; Canales, A.; Jimenez-Barbero, J.; Millet, O. Unraveling the Conformational Landscape of Ligand Binding to Glucose/Galactose-Binding Protein by Paramagnetic NMR and MD Simulations. ACS Chem. Biol. 2016, 11, 2149–2157. [Google Scholar] [CrossRef]
- Fonin, A.V.; Stepanenko, O.V.; Povarova, O.I.; Volova, C.A.; Philippova, E.M.; Bublikov, G.S.; Kuznetsova, I.M.; Demchenko, A.P.; Turoverov, K.K. Spectral characteristics of the mutant form GGBP/H152C of D-glucose/D-galactose-binding protein labeled with fluorescent dye BADAN: Influence of external factors. PeerJ 2014, 2, e275. [Google Scholar] [CrossRef] [Green Version]
- Fonin, A.V.; Golikova, A.D.; Zvereva, I.A.; D’Auria, S.; Staiano, M.; Uversky, V.N.; Kuznetsova, I.M.; Turoverov, K.K. Osmolyte-Like Stabilizing Effects of Low GdnHCl Concentrations on d-Glucose/d-Galactose-Binding Protein. Int. J. Mol. Sci. 2017, 18, 2008. [Google Scholar] [CrossRef] [Green Version]
- D’Auria, S.; Ausili, A.; Marabotti, A.; Varriale, A.; Scognamiglio, V.; Staiano, M.; Bertoli, E.; Rossi, M.; Tanfani, F. Binding of glucose to the D-galactose/D-glucose-binding protein from Escherichia coli restores the native protein secondary structure and thermostability that are lost upon calcium depletion. J. Biochem. 2006, 139, 213–221. [Google Scholar] [CrossRef]
- D’Auria, S.; Varriale, A.; Gonnelli, M.; Saviano, M.; Staiano, M.; Rossi, M.; Strambini, G.B. Tryptophan phosphorescence studies of the D-galactose/D-glucose-binding protein from Escherichia coli provide a molecular portrait with structural and dynamics features of the protein. J. Proteome Res. 2007, 6, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Fonin, A.V.; Silonov, S.A.; Sitdikova, A.K.; Kuznetsova, I.M.; Uversky, V.N.; Turoverov, K.K. Structure and Conformational Properties of d-Glucose/d-Galactose-Binding Protein in Crowded Milieu. Molecules 2017, 22, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangopadhyay, J.P.; Grabarek, Z.; Ikemoto, N. Fluorescence probe study of Ca2+-dependent interactions of calmodulin with calmodulin-binding peptides of the ryanodine receptor. Biochem. Biophys. Res. Commun. 2004, 323, 760–768. [Google Scholar] [CrossRef]
- Hammarstrom, P.; Owenius, R.; Martensson, L.G.; Carlsson, U.; Lindgren, M. High-resolution probing of local conformational changes in proteins by the use of multiple labeling: Unfolding and self-assembly of human carbonic anhydrase II monitored by spin, fluorescent, and chemical reactivity probes. Biophys. J. 2001, 80, 2867–2885. [Google Scholar] [CrossRef] [Green Version]
- Holt, A.; Koehorst, R.B.; Rutters-Meijneke, T.; Gelb, M.H.; Rijkers, D.T.; Hemminga, M.A.; Killian, J.A. Tilt and rotation angles of a transmembrane model peptide as studied by fluorescence spectroscopy. Biophys. J. 2009, 97, 2258–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pospisil, P.; Luxem, K.E.; Ener, M.; Sykora, J.; Kocabova, J.; Gray, H.B.; Vlcek, A., Jr.; Hof, M. Fluorescence quenching of (dimethylamino)naphthalene dyes Badan and Prodan by tryptophan in cytochromes P450 and micelles. J. Phys. Chem. B 2014, 118, 10085–10091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsalkova, T.N.; Davydova, N.Y.; Halpert, J.R.; Davydov, D.R. Mechanism of interactions of alpha-naphthoflavone with cytochrome P450 3A4 explored with an engineered enzyme bearing a fluorescent probe. Biochemistry 2007, 46, 106–119. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.S.; Vararattanavech, A.; Plasencia, I.; Greisen, P.J.; Bomholt, J.; Torres, J.; Emneus, J.; Helix-Nielsen, C. Interaction between sodium dodecyl sulfate and membrane reconstituted aquaporins: A comparative study of spinach SoPIP2;1 and E. coli AqpZ. Biochim. Biophys. Acta 2011, 1808, 2600–2607. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, C.M.; Bujalowski, P.J.; Ma, L.; Anderson, J.; Epstein, H.F.; Oberhauser, A.F. Tracking UNC-45 chaperone-myosin interaction with a titin mechanical reporter. Biophys. J. 2012, 102, 2212–2219. [Google Scholar] [CrossRef] [Green Version]
- Kipp, R.A.; Case, M.A.; Wist, A.D.; Cresson, C.M.; Carrell, M.; Griner, E.; Wiita, A.; Albiniak, P.A.; Chai, J.; Shi, Y.; et al. Molecular targeting of inhibitor of apoptosis proteins based on small molecule mimics of natural binding partners. Biochemistry 2002, 41, 7344–7349. [Google Scholar] [CrossRef]
- Nguyen, A.H.; Nguyen, V.T.; Kamio, Y.; Higuchi, H. Single-molecule visualization of environment-sensitive fluorophores inserted into cell membranes by staphylococcal gamma-hemolysin. Biochemistry 2006, 45, 2570–2576. [Google Scholar] [CrossRef]
- Stewart, R.C.; VanBruggen, R. Phosphorylation and binding interactions of CheY studied by use of Badan-labeled protein. Biochemistry 2004, 43, 8766–8777. [Google Scholar] [CrossRef]
- Kim, Y.E.; Chen, J.; Chan, J.R.; Langen, R. Engineering a polarity-sensitive biosensor for time-lapse imaging of apoptotic processes and degeneration. Nat. Methods 2010, 7, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Schindel, C.; Zitzer, A.; Schulte, B.; Gerhards, A.; Stanley, P.; Hughes, C.; Koronakis, V.; Bhakdi, S.; Palmer, M. Interaction of Escherichia coli hemolysin with biological membranes. A study using cysteine scanning mutagenesis. Eur. J. Biochem. 2001, 268, 800–808. [Google Scholar] [CrossRef]
- Fischermeier, E.; Pospisil, P.; Sayed, A.; Hof, M.; Solioz, M.; Fahmy, K. Dipolar Relaxation Dynamics at the Active Site of an ATPase Regulated by Membrane Lateral Pressure. Angew. Chem. Int. Ed. 2017, 56, 1269–1272. [Google Scholar] [CrossRef]
- Biswas, S.; Mukherjee, S.K.; Chowdhury, P.K. Crowder-Induced Rigidity in a Multidomain Protein: Insights from Solvation. J. Phys. Chem. B 2016, 120, 12501–12510. [Google Scholar] [CrossRef]
- Stepanenko, O.V.; Fonin, A.V.; Stepanenko, O.V.; Staiano, M.; D’Auria, S.; Kuznetsova, I.M.; Turoverov, K.K. Tryptophan residue of the D-galactose/D-glucose-binding protein from E. Coli localized in its active center does not contribute to the change in intrinsic fluorescence upon glucose binding. J. Fluoresc. 2015, 25, 87–94. [Google Scholar] [CrossRef]
- Fonin, A.V.; Sulatskaya, A.I.; Kuznetsova, I.M.; Turoverov, K.K. Fluorescence of dyes in solutions with high absorbance. Inner filter effect correction. PLoS ONE 2014, 9, e103878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonin, A.V.; Kuznetsova, I.M.; Turoverov, K.K. Spectral properties of BADAN in solutions with different polarities. J. Mol. Struct. 2015, 1090, 107–111. [Google Scholar] [CrossRef]
- Marabotti, A.; Ausili, A.; Staiano, M.; Scire, A.; Tanfani, F.; Parracino, A.; Varriale, A.; Rossi, M.; D’Auria, S. Pressure affects the structure and the dynamics of the D-galactose/D-glucose-binding protein from Escherichia coli by perturbing the C-terminal domain of the protein. Biochemistry 2006, 45, 11885–11894. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, V.; Scire, A.; Aurilia, V.; Staiano, M.; Crescenzo, R.; Palmucci, C.; Bertoli, E.; Rossi, M.; Tanfani, F.; D’Auria, S. A strategic fluorescence labeling of D-galactose/D-glucose-binding protein from Escherichia coli helps to shed light on the protein structural stability and dynamics. J. Proteome Res. 2007, 6, 4119–4126. [Google Scholar] [CrossRef]
- Piszczek, G.; D’Auria, S.; Staiano, M.; Rossi, M.; Ginsburg, A. Conformational stability and domain coupling in D-glucose/D-galactose-binding protein from Escherichia coli. Biochem. J. 2004, 381, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Marabotti, A.; Herman, P.; Staiano, M.; Varriale, A.; de Champdore, M.; Rossi, M.; Gryczynski, Z.; D’Auria, S. Pressure effect on the stability and the conformational dynamics of the D-Galactose/D-Glucose-binding protein from Escherichia coli. Proteins 2006, 62, 193–201. [Google Scholar] [CrossRef]
- Lim, W.K.; Rosgen, J.; Englander, S.W. Urea, but not guanidinium, destabilizes proteins by forming hydrogen bonds to the peptide group. Proc. Natl. Acad. Sci. USA 2009, 106, 2595–2600. [Google Scholar] [CrossRef] [Green Version]
- Mason, P.E.; Neilson, G.W.; Enderby, J.E.; Saboungi, M.L.; Dempsey, C.E.; MacKerell, A.D., Jr.; Brady, J.W. The structure of aqueous guanidinium chloride solutions. J. Am. Chem. Soc. 2004, 126, 11462–11470. [Google Scholar] [CrossRef]
- Carr, J.K.; Buchanan, L.E.; Schmidt, J.R.; Zanni, M.T.; Skinner, J.L. Structure and dynamics of urea/water mixtures investigated by vibrational spectroscopy and molecular dynamics simulation. J. Phys. Chem. B 2013, 117, 13291–13300. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Viga, A.; Woutersen, S. Protein Denaturation with Guanidinium: A 2D-IR Study. J. Phys. Chem. Lett. 2013, 4, 3397–3401. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tsumoto, K.; Umetsu, M.; Nakanishi, T.; Yasutake, Y.; Sakai, N.; Yao, M.; Tanaka, I.; Arakawa, T.; Kumagai, I. Structural evidence for guanidine-protein side chain interactions: Crystal structure of CutA from Pyrococcus horikoshii in 3 M guanidine hydrochloride. Biochem. Biophys. Res. Commun. 2004, 323, 185–191. [Google Scholar] [CrossRef]
- Dunbar, J.; Yennawar, H.P.; Banerjee, S.; Luo, J.; Farber, G.K. The effect of denaturants on protein structure. Protein Sci. 1997, 6, 1727–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolosa, L.; Gryczynski, I.; Eichhorn, L.R.; Dattelbaum, J.D.; Castellano, F.N.; Rao, G.; Lakowicz, J.R. Glucose sensor for low-cost lifetime-based sensing using a genetically engineered protein. Anal. Biochem. 1999, 267, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Harayama, S.; Bollinger, J.; Iino, T.; Hazelbauer, G.L. Characterization of the mgl operon of Escherichia coli by transposon mutagenesis and molecular cloning. J. Bacteriol. 1983, 153, 408–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotman, B.; Ganesan, A.K.; Guzman, R. Transport systems for galactose and galactosides in Escherichia coli. II. Substrate and inducer specificities. J. Mol. Biol. 1968, 36, 247–260. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Turoverov, K.K.; Haitlina, S.Y.; Pinaev, G.P. Ultra-violet fluorescence of actin. Determination of native actin content in actin preparations. FEBS Lett. 1976, 62, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Sillen, A.; Engelborghs, Y. The Correct Use of “Average” Fluorescence Parameters. Photochem. Photobiol. 1998, 67, 475–486. [Google Scholar]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006; p. 954. [Google Scholar]
- Anufrieva, Y.V.; Gotlib, Y.Y.; Krakovyak, M.G.; Pautov, V.D. Polarized luminescence as used for analyses of high frequency twisting vibrations in macromolecules. Polym. Sci. USSR 1976, 18, 3132–3141. [Google Scholar] [CrossRef]
- Tcherkasskaya, O.; Ptitsyn, O.B.; Knutson, J.R. Nanosecond dynamics of tryptophans in different conformational states of apomyoglobin proteins. Biochemistry 2000, 39, 1879–1889. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonin, A.V.; Silonov, S.A.; Antifeeva, I.A.; Stepanenko, O.V.; Stepanenko, O.V.; Fefilova, A.S.; Povarova, O.I.; Gavrilova, A.A.; Kuznetsova, I.M.; Turoverov, K.K. Photophysical Properties of BADAN Revealed in the Study of GGBP Structural Transitions. Int. J. Mol. Sci. 2021, 22, 11113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011113
Fonin AV, Silonov SA, Antifeeva IA, Stepanenko OV, Stepanenko OV, Fefilova AS, Povarova OI, Gavrilova AA, Kuznetsova IM, Turoverov KK. Photophysical Properties of BADAN Revealed in the Study of GGBP Structural Transitions. International Journal of Molecular Sciences. 2021; 22(20):11113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011113
Chicago/Turabian StyleFonin, Alexander V., Sergey A. Silonov, Iuliia A. Antifeeva, Olga V. Stepanenko, Olesya V. Stepanenko, Anna S. Fefilova, Olga I. Povarova, Anastasia A. Gavrilova, Irina M. Kuznetsova, and Konstantin K. Turoverov. 2021. "Photophysical Properties of BADAN Revealed in the Study of GGBP Structural Transitions" International Journal of Molecular Sciences 22, no. 20: 11113. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011113