
Chronic Rhinosinusitis and Alzheimer’s Disease—A Possible Role for the Nasal Microbiome in Causing Neurodegeneration in the Elderly


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Nasal Microbiota
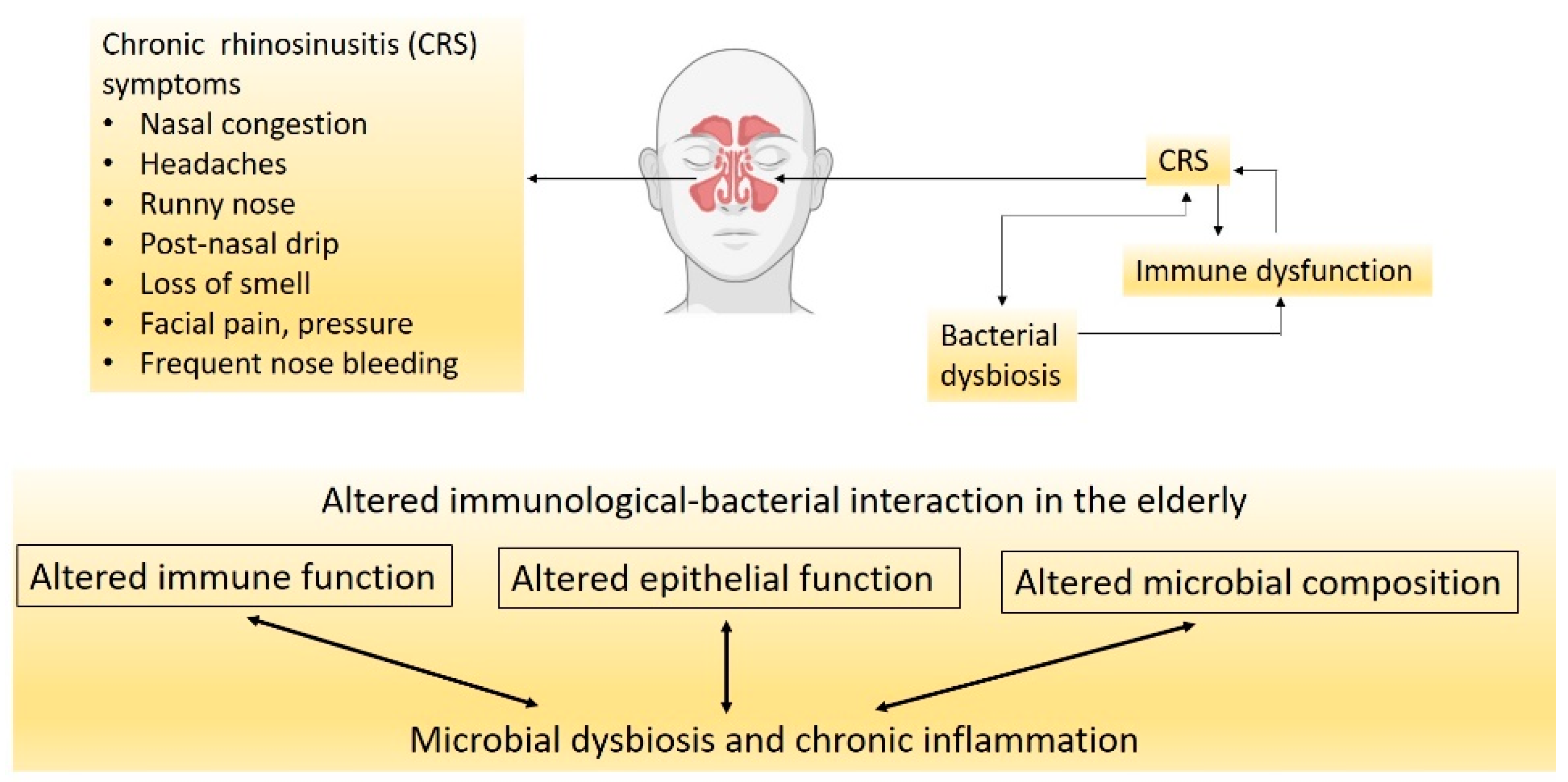
3. CRS, Nasal Microbiota and Their Influence on Neurological Health
4. Alzheimer’s Disease
5. CRS and AD—How Close Are They?
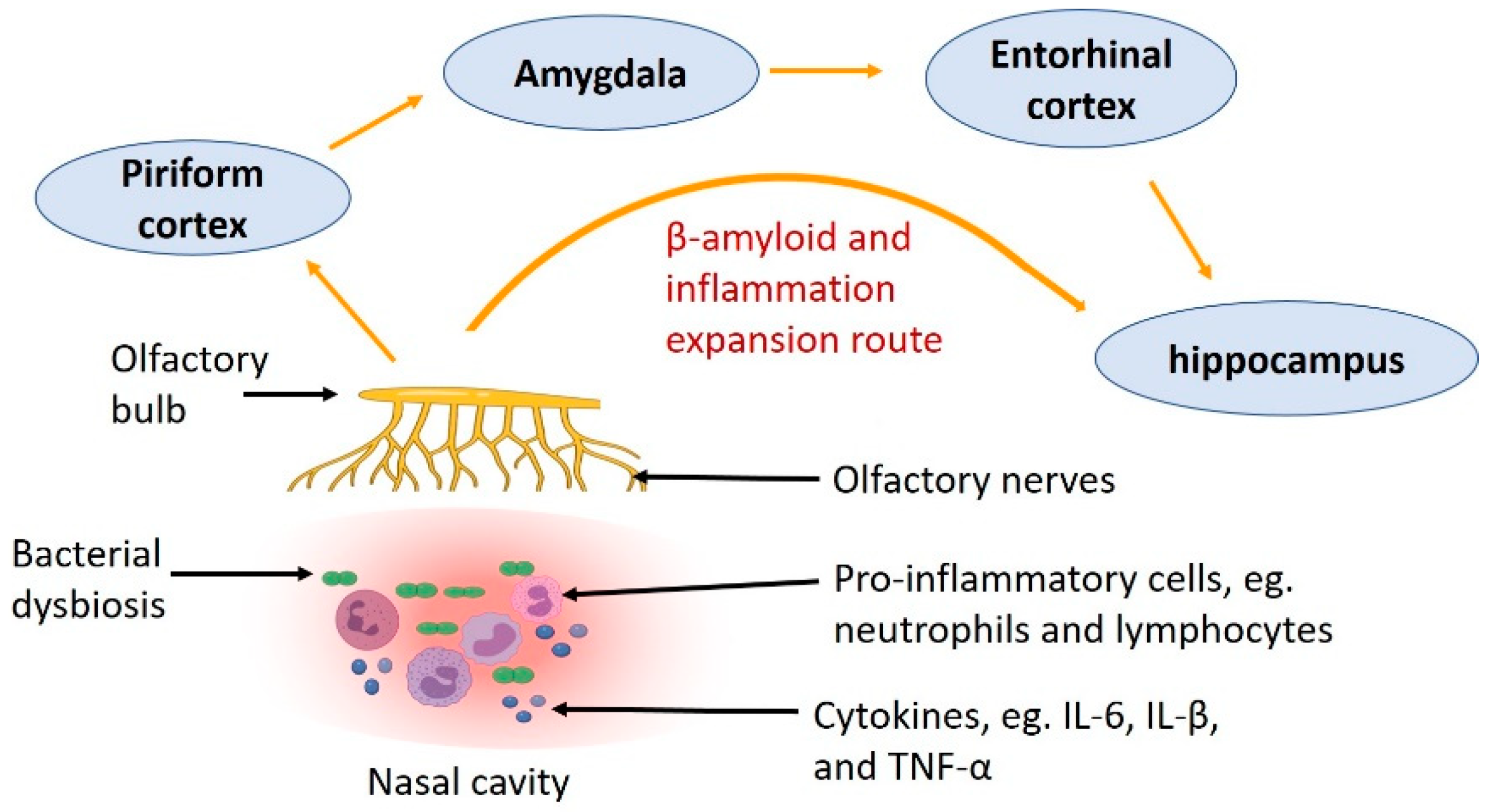
6. Expansion Route of Aβ—From the Nose to the Brain
7. Potential Mechanisms Linking Nasal Microbiota and AD
8. Future Research
Author Contributions
Funding
Conflicts of Interest
References
- Baquero, F.; Nombela, C. The microbiome as a human organ. Clin. Microbiol. Infect. 2012, 18 (Suppl. 4), 2–4. [Google Scholar] [CrossRef] [Green Version]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of Probiotic Supplementation on Cognitive Function and Metabolic Status in Alzheimer’s Disease: A Randomized, Double-Blind and Controlled Trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.-H.; Kuo, C.-W.; Hsieh, K.-H.; Shieh, M.-J.; Peng, C.-W.; Chen, Y.-C.; Chang, Y.-L.; Huang, Y.-Z.; Chen, C.-C.; Chang, P.-K.; et al. Probiotics Alleviate the Progressive Deterioration of Motor Functions in a Mouse Model of Parkinson’s Disease. Brain Sci. 2020, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Morshedi, M.; Hashemi, R.; Moazzen, S.; Sahebkar, A.; Hosseinifard, E.-S. Immunomodulatory and anti-inflammatory effects of probiotics in multiple sclerosis: A systematic review. J. Neuroinflammation 2019, 16, 231. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Li, J.; Gui, X.; Shi, X.; Bao, Z.; Han, H.; Li, M.D. Updated review of research on the gut microbiota and their relation to depression in animals and human beings. Mol. Psychiatry 2020, 25, 2759–2772. [Google Scholar] [CrossRef]
- Collins, S.M. A role for the gut microbiota in IBS. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Fazlollahi, M.; Lee, T.D.; Andrade, J.; Oguntuyo, K.; Chun, Y.; Grishina, G.; Grishin, A.; Bunyavanich, S. The nasal microbiome in asthma. J. Allergy Clin. Immunol. 2018, 142, 834–843.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutin, S.; Graeber, S.Y.; Weitnauer, M.; Panitz, J.; Stahl, M.; Clausznitzer, D.; Kaderali, L.; Einarsson, G.; Tunney, M.M.; Elborn, J.S.; et al. Comparison of microbiomes from different niches of upper and lower airways in children and adolescents with cystic fibrosis. PLoS ONE 2015, 10, e0116029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dando, S.J.; Mackay-Sim, A.; Norton, R.; Currie, B.J.; St John, J.A.; Ekberg, J.A.; Batzloff, M.; Ulett, G.C.; Beacham, I.R. Pathogens penetrating the central nervous system: Infection pathways and the cellular and molecular mechanisms of invasion. Clin. Microbiol. Rev. 2014, 27, 691–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lane, A.P. Chronic rhinosinusitis as a multifactorial inflammatory disorder. Curr. Infect. Dis. Rep. 2011, 13, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, F.; Schlosser, R.J.; Storck, K.A.; Ganjaei, K.G.; Rowan, N.R.; Soler, Z.M. Effects of endoscopic sinus surgery on objective and subjective measures of cognitive dysfunction in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2019, 9, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Rowan, N.R.; Schlosser, R.J.; Storck, K.A.; Ganjaei, K.G.; Soler, Z.M. The impact of medical therapy on cognitive dysfunction in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2019, 9, 738–745. [Google Scholar] [CrossRef] [PubMed]
- Andualem, Z.; Gizaw, Z.; Bogale, L.; Dagne, H. Indoor bacterial load and its correlation to physical indoor air quality parameters in public primary schools. Multidiscip. Respir. Med. 2019, 14, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, N. The nose and paranasal sinuses physiology and anatomy. Adv. Drug Deliv. Rev. 2001, 51, 5–19. [Google Scholar] [CrossRef]
- Mellert, T.K.; Getchell, M.L.; Sparks, L.; Getchell, T.V. Characterization of the immune barrier in human olfactory mucosa. Otolaryngol. Head Neck Surg. 1992, 106, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, K.; Reichert, J.L.; Hoier, S.; Schachenreiter, J.; Duller, S.; Moissl-Eichinger, C.; Schöpf, V. The nasal microbiome mirrors and potentially shapes olfactory function. Sci. Rep. 2018, 8, 1296. [Google Scholar] [CrossRef] [Green Version]
- Di Stadio, A.; Costantini, C.; Renga, G.; Pariano, M.; Ricci, G.; Romani, L. The Microbiota/Host Immune System Interaction in the Nose to Protect from COVID-19. Life 2020, 10, 345. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial community variation in human body habitats across space and time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Biesbroek, G.; Bosch, A.A.; Wang, X.; Keijser, B.J.; Veenhoven, R.H.; Sanders, E.A.; Bogaert, D. The impact of breastfeeding on nasopharyngeal microbial communities in infants. Am. J. Respir. Crit. Care Med. 2014, 190, 298–308. [Google Scholar] [CrossRef]
- Gritz, E.C.; Bhandari, V. The human neonatal gut microbiome: A brief review. Front. Pediatr. 2015, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Mueller, N.T.; Bakacs, E.; Combellick, J.; Grigoryan, Z.; Dominguez-Bello, M.G. The infant microbiome development: Mom matters. Trends Mol. Med. 2015, 21, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Bassis, C.M.; Tang, A.L.; Young, V.B.; Pynnonen, M.A. The nasal cavity microbiota of healthy adults. Microbiome 2014, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Stearns, J.C.; Davidson, C.J.; McKeon, S.; Whelan, F.J.; Fontes, M.E.; Schryvers, A.B.; Bowdish, D.M.; Kellner, J.D.; Surette, M.G. Culture and molecular-based profiles show shifts in bacterial communities of the upper respiratory tract that occur with age. ISME J. 2015, 9, 1246–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitri-Pinheiro, S.; Soares, R.; Barata, P. The Microbiome of the Nose—Friend or Foe? Allergy Rhinol. 2020, 11, 2152656720911605. [Google Scholar] [CrossRef] [Green Version]
- Riccardi, N.; Rotulo, G.A.; Castagnola, E. Definition of Opportunistic Infections in Immunocompromised Children on the Basis of Etiologies and Clinical Features: A Summary for Practical Purposes. Curr. Pediatr. Rev. 2019, 15, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Mahdavinia, M.; Keshavarzian, A.; Tobin, M.C.; Landay, A.L.; Schleimer, R.P. A comprehensive review of the nasal microbiome in chronic rhinosinusitis (CRS). Clin. Exp. Allergy 2016, 46, 21–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cope, E.K.; Goldberg, A.N.; Pletcher, S.D.; Lynch, S.V. Compositionally and functionally distinct sinus microbiota in chronic rhinosinusitis patients have immunological and clinically divergent consequences. Microbiome 2017, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ta, L.D.H.; Yap, G.C.; Tay, C.J.X.; Lim, A.S.M.; Huang, C.-H.; Chu, C.W.; De Sessions, P.F.; Shek, L.P.; Goh, A.; Van Bever, H.P.S.; et al. Establishment of the nasal microbiota in the first 18 months of life: Correlation with early-onset rhinitis and wheezing. J. Allergy Clin. Immunol. 2018, 142, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Bomar, L.; Brugger, S.D.; Lemon, K.P. Bacterial microbiota of the nasal passages across the span of human life. Curr. Opin. Microbiol. 2018, 41, 8–14. [Google Scholar] [CrossRef]
- Ramakrishnan, V.R.; Feazel, L.M.; Gitomer, S.A.; Ir, D.; Robertson, C.E.; Frank, D.N. The microbiome of the middle meatus in healthy adults. PLoS ONE 2013, 8, e85507. [Google Scholar] [CrossRef] [PubMed]
- Whelan, F.J.; Verschoor, C.P.; Stearns, J.C.; Rossi, L.; Luinstra, K.; Loeb, M.; Smieja, M.; Johnstone, J.; Surette, M.G.; Bowdish, D.M. The loss of topography in the microbial communities of the upper respiratory tract in the elderly. Ann. Am. Thorac. Soc. 2014, 11, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Schenck, L.P.; Surette, M.G.; Bowdish, D.M. Composition and immunological significance of the upper respiratory tract microbiota. FEBS Lett. 2016, 590, 3705–3720. [Google Scholar] [CrossRef]
- Desrosiers, M.; Evans, G.A.; Keith, P.K.; Wright, E.D.; Kaplan, A.; Bouchard, J.; Ciavarella, A.; Doyle, P.W.; Javer, A.R.; Leith, E.S.; et al. Canadian clinical practice guidelines for acute and chronic rhinosinusitis. Allergy Asthma Clin. Immunol. 2011, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedaghat, A.R. Chronic Rhinosinusitis. Am. Fam Physician 2017, 96, 500–506. [Google Scholar]
- Hamilos, D.L.M.D. Drivers of chronic rhinosinusitis: Inflammation versus infection. J. Allergy Clin. Immunol. 2015, 136, 1454–1459. [Google Scholar] [CrossRef] [Green Version]
- Stevens, W.W.; Lee, R.J.; Schleimer, R.P.; Cohen, N.A. Chronic rhinosinusitis pathogenesis. J. Allergy Clin. Immunol. 2015, 136, 1442–1453. [Google Scholar] [CrossRef] [Green Version]
- Abreu, N.A.; Nagalingam, N.A.; Song, Y.; Roediger, F.C.; Pletcher, S.D.; Goldberg, A.N.; Lynch, S.V. Sinus microbiome diversity depletion and Corynebacterium tuberculostearicum enrichment mediates rhinosinusitis. Sci. Transl. Med. 2012, 4, 151ra124. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.H.; Hong, S.J.; Han, B.; Lee, S.H.; Suh, L.; Norton, J.; Lin, D.; Conley, D.B.; Chandra, R.; Kern, R.C.; et al. Age-related differences in the pathogenesis of chronic rhinosinusitis. J. Allergy Clin. Immunol. 2012, 129, 858–860.e2. [Google Scholar] [CrossRef] [Green Version]
- Busse, P.J.; Mathur, S.K. Age-related changes in immune function: Effect on airway inflammation. J. Allergy Clin. Immunol. 2010, 126, 690–699. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.C.; Chan, K.N.; Hu, W.H.; Lam, W.K.; Ling, Z.; Tipoe, G.L.; June, S.U.N.; Leung, R.; Tsang, K.W. The effect of aging on nasal mucociliary clearance, beat frequency, and ultrastructure of respiratory cilia. Am. J. Respir. Crit. Care Med. 2001, 163, 983–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proença de Oliveira-Maul, J.; Barbosa de Carvalho, H.; Goto, D.M.; Maia, R.M.; Fló, C.; Barnabé, V.; Franco, D.R.; Benabou, S.; Perracini, M.R.; Jacob-Filho, W.; et al. Aging, diabetes, and hypertension are associated with decreased nasal mucociliary clearance. Chest 2013, 143, 1091–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubb, B.R.; Livraghi-Butrico, A.; Rogers, T.D.; Yin, W.; Button, B.; Ostrowski, L.E. Reduced mucociliary clearance in old mice is associated with a decrease in Muc5b mucin. Am. J. Physiology. Lung Cell. Mol. Physiol. 2016, 310, L860–L867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baptist, A.P.; Nyenhuis, S. Rhinitis in the Elderly. Immunol. Allergy Clin. N. Am. 2016, 36, 343–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, P.A.; Wise, S.K.; Nieto, D.; Panella, N.; Aiken, A.; DelGaudio, J.M. Intranasal volume increases with age: Computed tomography volumetric analysis in adults. Laryngoscope 2016, 126, 2212–2215. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, M., Jr.; Lane, A.P. Innate immunity of the sinonasal cavity and its role in chronic rhinosinusitis. Otolaryngol. Head Neck Surg. 2007, 136, 348–356. [Google Scholar] [CrossRef]
- Lane, A.P. The role of innate immunity in the pathogenesis of chronic rhinosinusitis. Curr. Allergy Asthma Rep. 2009, 9, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, A.; Patadia, M.; Stankiewicz, J. Neurological Complications of Acute and Chronic Sinusitis. Curr. Neurol. Neurosci. Rep. 2018, 18, 5. [Google Scholar] [CrossRef]
- Snowdon, D.A.; Greiner, L.H.; Mortimer, J.A.; Riley, K.P.; Greiner, P.A.; Markesbery, W.R. Brain Infarction and the Clinical Expression of Alzheimer Disease: The Nun Study. JAMA 1997, 277, 813–817. [Google Scholar] [CrossRef]
- Alzheimer’s Association. 2016 Alzheimer’s disease facts and figures. Alzheimers Dement. 2016, 12, 459–509. [Google Scholar] [CrossRef]
- Bature, F.; Guinn, B.-A.; Pang, D.; Pappas, Y. Signs and symptoms preceding the diagnosis of Alzheimer’s disease: A systematic scoping review of literature from 1937 to 2016. BMJ Open 2017, 7, e015746. [Google Scholar] [CrossRef] [Green Version]
- Panza, F.; Seripa, D.; D’Onofrio, G.; Frisardi, V.; Solfrizzi, V.; Mecocci, P.; Pilotto, A. Neuropsychiatric Symptoms, Endophenotypes, and Syndromes in Late-Onset Alzheimer’s Disease: Focus on APOE Gene. Int. J. Alzheimers Dis. 2011, 2011, 721457. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Hu, N.; Tan, M.S.; Yu, J.T.; Tan, L. Behavioral and psychological symptoms in Alzheimer’s disease. BioMed Res. Int. 2014, 2014, 927804. [Google Scholar] [CrossRef]
- Guerreiro, R.; Bras, J. The age factor in Alzheimer’s disease. Genome Med. 2015, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.Y.; Snyder, P.J.; Wu, W.C.; Zhang, M.; Echeverria, A.; Alber, J. Pathophysiologic relationship between Alzheimer’s disease, cerebrovascular disease, and cardiovascular risk: A review and synthesis. Alzheimers Dement. Diagn. Assess. Dis. Monit. 2017, 7, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Croze, M.L.; Zimmer, L. Ozone Atmospheric Pollution and Alzheimer’s Disease: From Epidemiological Facts to Molecular Mechanisms. J. Alzheimers Dis. 2018, 62, 503–522. [Google Scholar] [CrossRef]
- Lotz, S.K.; Blackhurst, B.M.; Reagin, K.L.; Funk, K.E. Microbial Infections Are a Risk Factor for Neurodegenerative Diseases. Front. Cell. Neurosci. 2021, 15, 246. [Google Scholar] [CrossRef]
- Subramaniapillai, S.; Almey, A.; Natasha Rajah, M.; Einstein, G. Sex and gender differences in cognitive and brain reserve: Implications for Alzheimer’s disease in women. Front. Neuroendocrinol. 2021, 60, 100879. [Google Scholar] [CrossRef]
- Bretsky, P.M.; Buckwalter, J.G.; Seeman, T.E.; Miller, C.A.; Poirier, J.; Schellenberg, G.D.; Finch, C.E.; Henderson, V.W. Evidence for an interaction between apolipoprotein E genotype, gender, and Alzheimer disease. Alzheimer Dis. Assoc. Disord. 1999, 13, 216–221. [Google Scholar] [CrossRef]
- Gilsanz, P.; Lee, C.; Corrada, M.M.; Kawas, C.H.; Quesenberry, C.P., Jr.; Whitmer, R.A. Reproductive period and risk of dementia in a diverse cohort of health care members. Neurology 2019, 92, e2005–e2014. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Blalock, E.M.; Chen, K.C.; Stromberg, A.J.; Norris, C.M.; Kadish, I.; Kraner, S.D.; Porter, N.M.; Landfield, P.W. Harnessing the power of gene microarrays for the study of brain aging and Alzheimer’s disease: Statistical reliability and functional correlation. Ageing Res. Rev. 2005, 4, 481–512. [Google Scholar] [CrossRef]
- Roses, A.D. Apolipoprotein E is a relevant susceptibility gene that affects the rate of expression of Alzheimer’s disease. Neurobiol. Aging 1994, 15 (Suppl. 2), S165–S167. [Google Scholar] [CrossRef]
- Selkoe, D.J. Alzheimer’s disease. In the beginning. Nature 1991, 354, 432–433. [Google Scholar] [CrossRef]
- Castellani, R.J.; Smith, M.A. Compounding artefacts with uncertainty, and an amyloid cascade hypothesis that is ‘too big to fail’. J. Pathol. 2011, 224, 147–152. [Google Scholar] [CrossRef]
- Yoon, S.-S.; Jo, S.A. Mechanisms of Amyloid-β Peptide Clearance: Potential Therapeutic Targets for Alzheimer’s Disease. Biomol. Ther. 2012, 20, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.J.; Munsell, L.Y.; Morris, J.C.; Swarm, R.; Yarasheski, K.E.; Holtzman, D.M. Human amyloid-beta synthesis and clearance rates as measured in cerebrospinal fluid in vivo. Nat. Med. 2006, 12, 856–861. [Google Scholar] [CrossRef]
- Sun, X.; Chen, W.-D.; Wang, Y.-D. β-Amyloid: The key peptide in the pathogenesis of Alzheimer’s disease. Front. Pharmacol. 2015, 6, 221. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, U.; Nilson, A.N.; Kayed, R. The Role of Amyloid-β Oligomers in Toxicity, Propagation, and Immunotherapy. EBioMedicine 2016, 6, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Sanchez, D.; De la Rosa-Prieto, C.; Ubeda-Banon, I.; Martinez-Marcos, A. Interneurons, tau and amyloid-β in the piriform cortex in Alzheimer’s disease. Brain Struct. Funct. 2015, 220, 2011–2025. [Google Scholar] [CrossRef]
- Jung, H.J.; Lee, J.Y.; Choi, Y.S.; Choi, H.G.; Wee, J.H. Chronic rhinosinusitis and progression of cognitive impairment in dementia. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2021, 138, 147–151. [Google Scholar] [CrossRef]
- Chung, S.-D.; Hung, S.-H.; Lin, H.-C.; Kang, J.-H. Dementia is associated with chronic rhinosinusitis: A population-based case-controlled study. Am. J. Rhinol. Allergy 2015, 29, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Tarasidis, G.S.; DeConde, A.S.; Mace, J.C.; Ashby, S.; Smith, T.L.; Orlandi, R.R.; Alt, J.A. Cognitive dysfunction associated with pain and quality of life in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2015, 5, 1004–1009. [Google Scholar] [CrossRef]
- Matsui, T.; Arai, H.; Nakajo, M.; Maruyama, M.; Ebihara, S.; Sasaki, H.; Yoshida, Y.-i. Role of Chronic Sinusitis in Cognitive Functioning in the Elderly. J. Am. Geriatr. Soc. 2003, 51, 1818–1819. [Google Scholar] [CrossRef]
- Jafari, A.; de Lima Xavier, L.; Bernstein, J.D.; Simonyan, K.; Bleier, B.S. Association of Sinonasal Inflammation With Functional Brain Connectivity. JAMA Otolaryngol.-Head Neck Surg. 2021, 147, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Yasue, M.; Sugiura, S.; Uchida, Y.; Otake, H.; Teranishi, M.; Sakurai, T.; Toba, K.; Shimokata, H.; Ando, F.; Otsuka, R.; et al. Prevalence of Sinusitis Detected by Magnetic Resonance Imaging in Subjects with Dementia or Alzheimer’s Disease. Curr. Alzheimer Res. 2015, 12, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Brook, C.D.; Kuperstock, J.E.; Rubin, S.J.; Ryan, M.W.; Platt, M.P. The Association of Allergic Sensitization with Radiographic Sinus Opacification. Am. J. Rhinol. Allergy 2017, 31, 12–15. [Google Scholar] [CrossRef]
- Lal, D.; Keim, P.; Delisle, J.; Barker, B.; Rank, M.A.; Chia, N.; Schupp, J.M.; Gillece, J.D.; Cope, E.K. Mapping and comparing bacterial microbiota in the sinonasal cavity of healthy, allergic rhinitis, and chronic rhinosinusitis subjects. Int. Forum Allergy Rhinol. 2017, 7, 561–569. [Google Scholar] [CrossRef]
- Wee, J.H.; Yoo, D.M.; Byun, S.H.; Hong, S.J.; Park, M.W.; Choi, H.G. Association between neurodegenerative dementia and chronic rhinosinusitis: A nested case-control study using a national health screening cohort. Medicine 2020, 99, e22141. [Google Scholar] [CrossRef]
- Daulatzai, M.A. Quintessential Risk Factors: Their Role in Promoting Cognitive Dysfunction and Alzheimer’s Disease. Neurochem. Res. 2012, 37, 2627–2658. [Google Scholar] [CrossRef]
- Ziehn, M.O.; Avedisian, A.A.; Tiwari-Woodruff, S.; Voskuhl, R.R. Hippocampal CA1 atrophy and synaptic loss during experimental autoimmune encephalomyelitis, EAE. Lab. Investig. 2010, 90, 774–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.-Y.; Tan, M.-S.; Yu, J.-T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar] [CrossRef] [PubMed]
- Yarchoan, M.; Louneva, N.; Xie, S.X.; Swenson, F.J.; Hu, W.; Soares, H.; Trojanowski, J.Q.; Lee, V.M.Y.; Kling, M.A.; Shaw, L.M.; et al. Association of plasma C-reactive protein levels with the diagnosis of Alzheimer’s disease. J. Neurol. Sci. 2013, 333, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Kaur, D.; Sharma, V.; Deshmukh, R. Activation of microglia and astrocytes: A roadway to neuroinflammation and Alzheimer’s disease. Inflammopharmacology 2019, 27, 663–677. [Google Scholar] [CrossRef]
- Broe, G.A.; Grayson, D.A.; Creasey, H.M.; Waite, L.M.; Casey, B.J.; Bennett, H.P.; Brooks, W.S.; Halliday, G.M. Anti-inflammatory Drugs Protect Against Alzheimer Disease at Low Doses. Arch. Neurol. 2000, 57, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Brandtzaeg, P. Cells Producing Immunoglobulins and other Immune Factors in Human Nasal Mucosa. In Protides of the Biological Fluids; Peeters, H., Ed.; Elsevier: Amsterdam, The Netherland, 1985; Volume 32, pp. 363–366. [Google Scholar]
- Brescia, G.; Barion, U.; Zanotti, C.; Giacomelli, L.; Martini, A.; Marioni, G. The prognostic role of serum eosinophil and basophil levels in sinonasal polyposis. Int. Forum Allergy Rhinol. 2017, 7, 261–267. [Google Scholar] [CrossRef]
- Hulse, K.E.; Stevens, W.W.; Tan, B.K.; Schleimer, R.P. Pathogenesis of nasal polyposis. Clin. Exp. Allergy 2015, 45, 328–346. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.H.; Liang, K.L.; May, L.; Lane, A.P. Tumor necrosis factor alpha inhibits olfactory regeneration in a transgenic model of chronic rhinosinusitis-associated olfactory loss. Am. J. Rhinol. Allergy 2010, 24, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Swardfager, W.; Lanctôt, K.; Rothenburg, L.; Wong, A.; Cappell, J.; Herrmann, N. A meta-analysis of cytokines in Alzheimer’s disease. Biol. Psychiatry 2010, 68, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Lennard, C.M.; Mann, E.A.; Sun, L.L.; Chang, A.S.; Bolger, W.E. Interleukin-1 beta, interleukin-5, interleukin-6, interleukin-8, and tumor necrosis factor-alpha in chronic sinusitis: Response to systemic corticosteroids. Am. J. Rhinol. 2000, 14, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Scheckenbach, K.; Wagenmann, M. Cytokine Patterns and Endotypes in Acute and Chronic Rhinosinusitis. Curr. Allergy Asthma Rep. 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Kou, W.; Hu, G.H.; Yao, H.B.; Wang, X.; Shen, Y.; Kang, H.Y.; Hong, S.L. Regulation of Transforming Growth Factor-β1 Activation and Expression in the Tissue Remodeling Involved in Chronic Rhinosinusitis. ORL 2012, 74, 172–178. [Google Scholar] [CrossRef] [PubMed]
- von Bernhardi, R.; Cornejo, F.; Parada, G.E.; Eugenín, J. Role of TGFβ signaling in the pathogenesis of Alzheimer’s disease. Front. Cell. Neurosci. 2015, 9, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Isla, T.; Price, J.L.; McKeel, D.W., Jr.; Morris, J.C.; Growdon, J.H.; Hyman, B.T. Profound loss of layer II entorhinal cortex neurons occurs in very mild Alzheimer’s disease. J. Neurosci. 1996, 16, 4491–4500. [Google Scholar] [CrossRef]
- Du, A.T.; Schuff, N.; Kramer, J.H.; Ganzer, S.; Zhu, X.P.; Jagust, W.J.; Miller, B.L.; Reed, B.R.; Mungas, D.; Yaffe, K.; et al. Higher atrophy rate of entorhinal cortex than hippocampus in AD. Neurology 2004, 62, 422–427. [Google Scholar] [CrossRef]
- Velayudhan, L.; Proitsi, P.; Westman, E.; Muehlboeck, J.S.; Mecocci, P.; Vellas, B.; Tsolaki, M.; Kłoszewska, I.; Soininen, H.; Spenger, C.; et al. Entorhinal cortex thickness predicts cognitive decline in Alzheimer’s disease. J. Alzheimers Dis. 2013, 33, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Cross, D.J.; Anzai, Y.; Petrie, E.C.; Martin, N.; Richards, T.L.; Maravilla, K.R.; Peskind, E.R.; Minoshima, S. Loss of olfactory tract integrity affects cortical metabolism in the brain and olfactory regions in aging and mild cognitive impairment. J. Nucl. Med. 2013, 54, 1278–1284. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Rao, X.; Gao, Y.; Wang, J.; Xu, F. Amyloid-β deposition and olfactory dysfunction in an Alzheimer’s disease model. J. Alzheimers Dis. 2013, 37, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Sanchez, D.; Flores-Cuadrado, A.; Ubeda-Bañon, I.; de la Rosa-Prieto, C.; Martinez-Marcos, A. Interneurons in the human olfactory system in Alzheimer’s disease. Exp. Neurol. 2016, 276, 13–21. [Google Scholar] [CrossRef]
- Attems, J.; Lintner, F.; Jellinger, K.A. Olfactory involvement in aging and Alzheimer’s disease: An autopsy study. J. Alzheimer’s Dis. 2005, 7, 149–157. [Google Scholar] [CrossRef]
- Clavaguera, F.; Hench, J.; Goedert, M.; Tolnay, M. Invited review: Prion-like transmission and spreading of tau pathology. Neuropathol. Appl. Neurobiol. 2015, 41, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Imamura, F.; Ito, A.; LaFever, B.J. Subpopulations of Projection Neurons in the Olfactory Bulb. Front. Neural Circuits 2020, 14. [Google Scholar] [CrossRef]
- Yoo, S.-J.; Son, G.; Bae, J.; Kim, S.Y.; Yoo, Y.K.; Park, D.; Baek, S.Y.; Chang, K.-A.; Suh, Y.-H.; Lee, Y.-B.; et al. Longitudinal profiling of oligomeric Aβ in human nasal discharge reflecting cognitive decline in probable Alzheimer’s disease. Sci. Rep. 2020, 10, 11234. [Google Scholar] [CrossRef]
- Litvack, J.R.; Fong, K.; Mace, J.; James, K.E.; Smith, T.L. Predictors of Olfactory Dysfunction in Patients With Chronic Rhinosinusitis. Laryngoscope 2008, 118, 2225–2230. [Google Scholar] [CrossRef] [Green Version]
- Kern, R.C. Chronic sinusitis and anosmia: Pathologic changes in the olfactory mucosa. Laryngoscope 2000, 110, 1071–1077. [Google Scholar] [CrossRef]
- Alt, J.A.; Mace, J.C.; Smith, T.L.; Soler, Z.M. Endoscopic sinus surgery improves cognitive dysfunction in patients with chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2016, 6, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- Alzheimer’s Association. 2021 Alzheimer’s disease facts and figures. Alzheimers Dement. 2021, 17, 327–406. [Google Scholar] [CrossRef]
- Manabe, T.; Fujikura, Y.; Mizukami, K.; Akatsu, H.; Kudo, K. Pneumonia-associated death in patients with dementia: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0213825. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.H.; Huang, Y.C.; Chen, C.J.; Lin, P.Y.; Chang, L.Y.; Chiu, C.H.; Tsao, K.C.; Huang, C.G.; Lin, T.Y. Chlamydial pneumonia in children requiring hospitalization: Effect of mixed infection on clinical outcome. J. Microbiol. Immunol. Infect. 2005, 38, 117–122. [Google Scholar]
- Mulvihill, N.T.; Foley, J.B. Inflammation in acute coronary syndromes. Heart 2002, 87, 201–204. [Google Scholar] [CrossRef]
- Heller, J.P.; Rusakov, D.A. Morphological plasticity of astroglia: Understanding synaptic microenvironment. Glia 2015, 63, 2133–2151. [Google Scholar] [CrossRef]
- Fischer, H.G.; Reichmann, G. Brain dendritic cells and macrophages/microglia in central nervous system inflammation. J. Immunol. 2001, 166, 2717–2726. [Google Scholar] [CrossRef] [Green Version]
- Gérard, H.C.; Dreses-Werringloer, U.; Wildt, K.S.; Deka, S.; Oszust, C.; Balin, B.J.; Frey, W.H., 2nd; Bordayo, E.Z.; Whittum-Hudson, J.A.; Hudson, A.P. Chlamydophila (Chlamydia) pneumoniae in the Alzheimer’s brain. FEMS Immunol. Med. Microbiol. 2006, 48, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Emery, D.C.; Shoemark, D.K.; Batstone, T.E.; Waterfall, C.M.; Coghill, J.A.; Cerajewska, T.L.; Davies, M.; West, N.X.; Allen, S.J. 16S rRNA Next Generation Sequencing Analysis Shows Bacteria in Alzheimer’s Post-Mortem Brain. Front. Aging Neurosci. 2017, 9, 195. [Google Scholar] [CrossRef]
- Zhao, Y.; Cong, L.; Lukiw, W.J. Lipopolysaccharide (LPS) Accumulates in Neocortical Neurons of Alzheimer’s Disease (AD) Brain and Impairs Transcription in Human Neuronal-Glial Primary Co-cultures. Front. Aging Neurosci. 2017, 9, 407. [Google Scholar] [CrossRef]
- Shibuya, Y.; Kawakatsu, S.; Hayashi, H.; Kobayashi, R.; Suzuki, A.; Sato, C.; Otani, K. Comparison of entorhinal cortex atrophy between early-onset and late-onset Alzheimer’s disease using the VSRAD, a specific and sensitive voxel-based morphometry. Int. J. Geriatr. Psychiatry 2013, 28, 372–376. [Google Scholar] [CrossRef]
- Piromchai, P.; Kasemsiri, P.; Laohasiriwong, S.; Thanaviratananich, S. Chronic rhinosinusitis and emerging treatment options. Int. J. Gen. Med. 2013, 6, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Barshak, M.B.; Durand, M.L. The role of infection and antibiotics in chronic rhinosinusitis. Laryngoscope Investig. Otolaryngol. 2017, 2, 36–42. [Google Scholar] [CrossRef]
- Fairbanks, D.N. Pocket Guide to Antimicrobial Therapy in Otolaryngology; American Academy of Otolaryngology-Head and Neck Surgery Foundatin, Incorporated: Alexandria, VA, USA, 1983. [Google Scholar]
- Kuang, L.; Hashimoto, K.; Huang, E.J.; Gentry, M.S.; Zhu, H. Frontotemporal dementia non-sense mutation of progranulin rescued by aminoglycosides. Hum. Mol. Genet. 2020, 29, 624–634. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrass, S.; Yi, C.; Chen, H. Chronic Rhinosinusitis and Alzheimer’s Disease—A Possible Role for the Nasal Microbiome in Causing Neurodegeneration in the Elderly. Int. J. Mol. Sci. 2021, 22, 11207. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011207
Harrass S, Yi C, Chen H. Chronic Rhinosinusitis and Alzheimer’s Disease—A Possible Role for the Nasal Microbiome in Causing Neurodegeneration in the Elderly. International Journal of Molecular Sciences. 2021; 22(20):11207. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011207
Chicago/Turabian StyleHarrass, Sanaa, Chenju Yi, and Hui Chen. 2021. "Chronic Rhinosinusitis and Alzheimer’s Disease—A Possible Role for the Nasal Microbiome in Causing Neurodegeneration in the Elderly" International Journal of Molecular Sciences 22, no. 20: 11207. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011207