
Novel and Annotated Long Noncoding RNAs Associated with Ischemia in the Human Heart

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
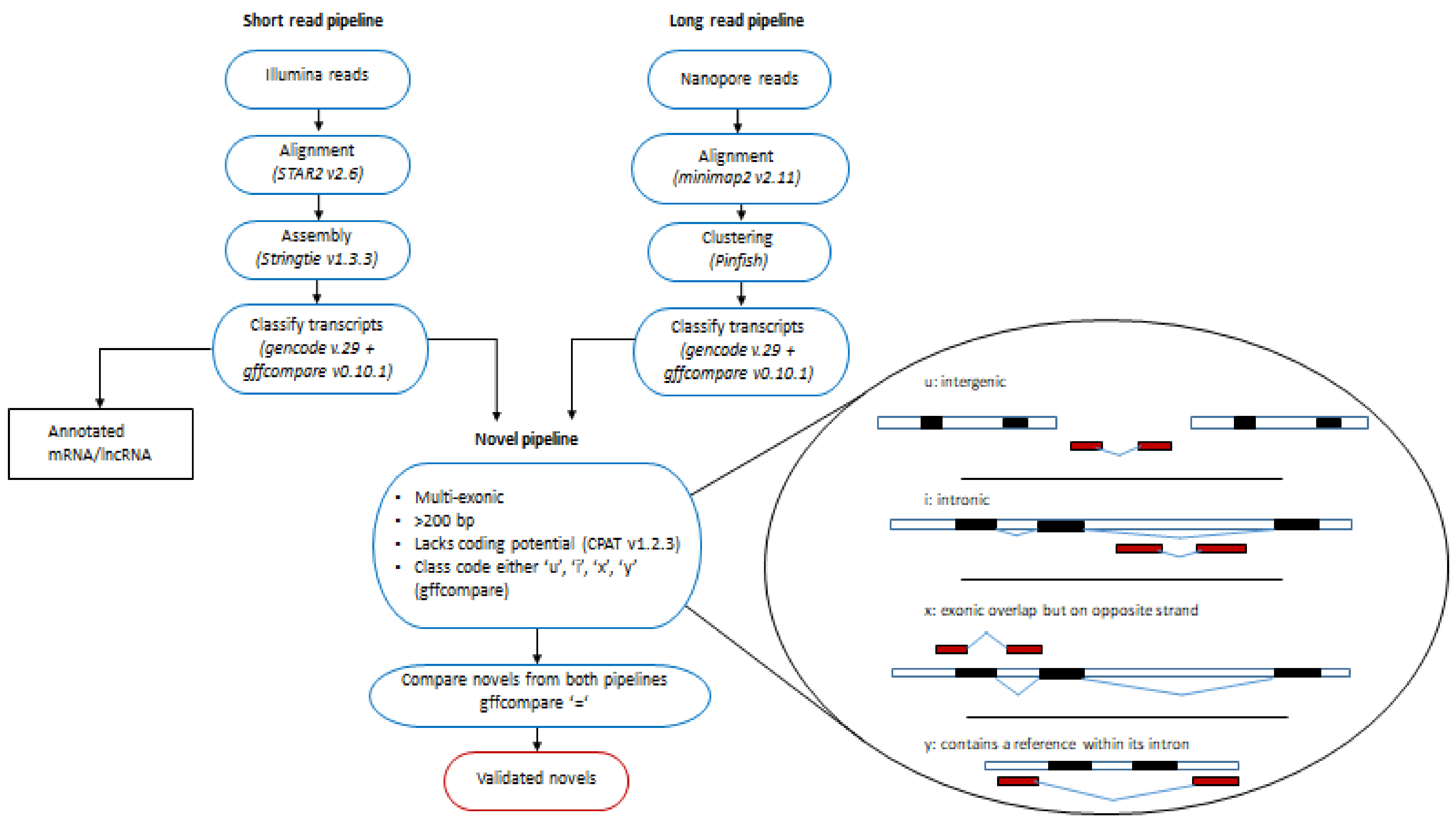
2.1. Protein Coding, Annotated and Novel lncRNAs Associated with Ischemia Identified with Illumina Short-Read Sequencing
2.2. Validating Novel lncRNAs in the Human Left Ventricle with Nanopore Long-Read Technology
2.3. Evolutionary Conservation of lncRNAs
2.4. Identifying Gene Networks Associated with Ischemia
2.5. Overlap of Annotated and Novel lncRNAs with Cis-eQTLs
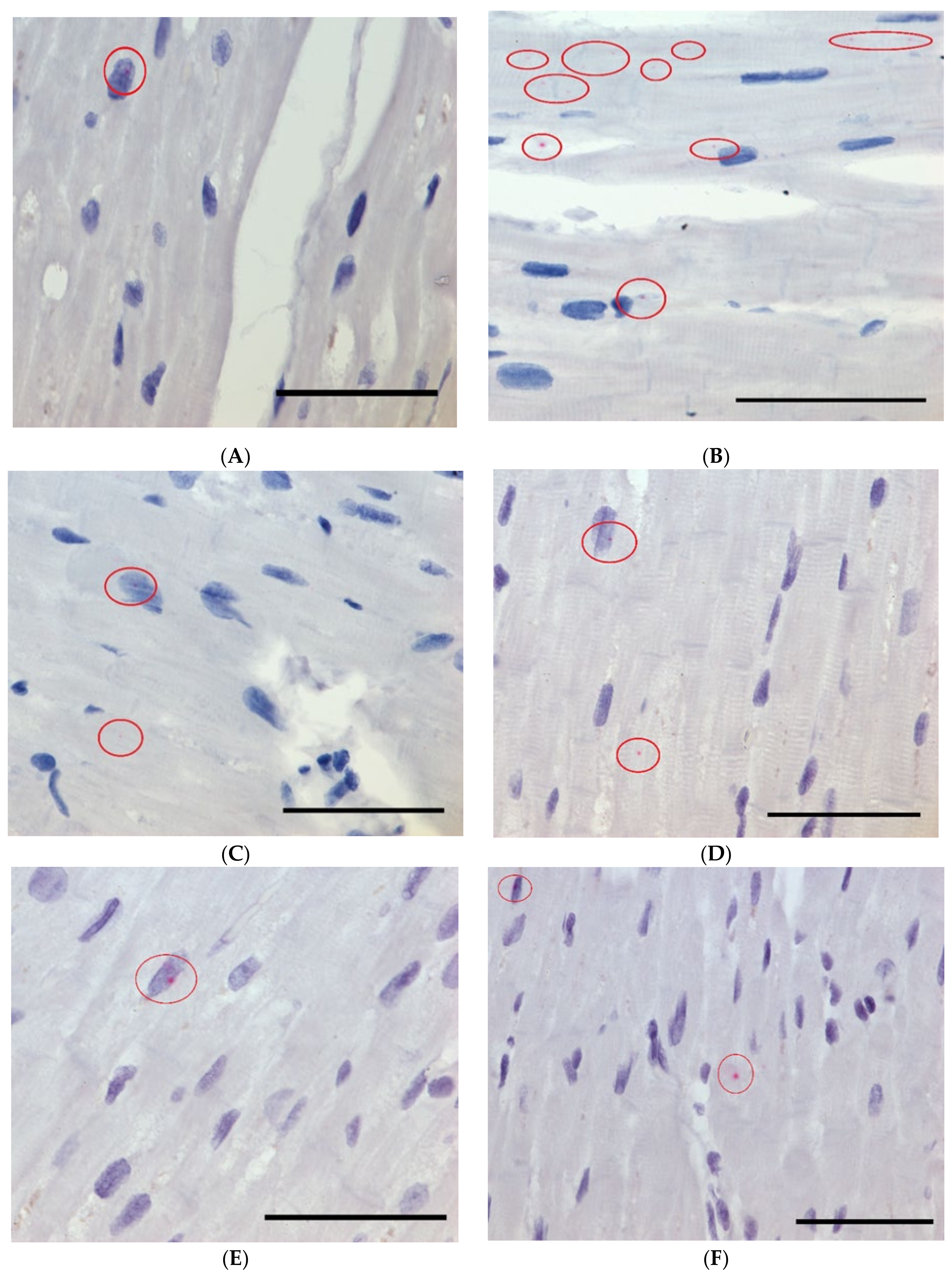
2.6. Subcellular Localization of Ischemia-Associated lncRNAs with RNA Scope

3. Discussion
4. Materials and Methods
4.1. Illumina Sequencing and QC Analysis
4.2. Nanopore Long-Read Sequencing
4.3. Identification and Validation of Novel lncRNAs
4.4. Conservation of Novel lncRNAs
4.5. Differential Expression
4.6. WGCNA: Weighted Correlation Network Analysis
4.7. Ingenuity Pathway Analysis (IPA)
4.8. Expression Quantitative Trait Locus (eQTLs)
4.9. Subcellular Localization of lncRNAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernstein, B.E.; Birney, E.; Dunham, I.; Green, E.D.; Gunter, C.; Snyder, M. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nat. Cell Biol. 2009, 458, 223–227. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [Green Version]
- Hon, C.-C.; Ramilowski, J.A.; Harshbarger, J.; Bertin, N.; Rackham, O.J.L.; Gough, J.; Denisenko, E.; Schmeier, S.; Poulsen, T.M.; Severin, J.; et al. An atlas of human long non-coding RNAs with accurate 5′ ends. Nat. Cell Biol. 2017, 543, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraczkowska, W.; Jagodziński, P.P. The long non-coding RNA landscape of atherosclerotic plaques. Mol. Diagn. Ther. 2019, 23, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Danese, E.; Montagnana, M. An historical approach to the diagnostic biomarkers of acute coronary syndrome. Ann. Transl. Med. 2016, 4, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddic, L.A.; Sigurdsson, M.; Chang, T.-W.; Mazaika, E.; Heydarpour, M.; Shernan, S.K.; Seidman, C.E.; Seidman, J.G.; Aranki, S.F.; Body, S.C.; et al. The long noncoding RNA landscape of the ischemic human left ventricle. Circ. Cardiovasc. Genet. 2017, 10, e001534. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, L.; Song, J.; Wang, Z.; Huang, X.; Guo, Z.; Chen, F.; Zhao, X. Long noncoding RNA MALAT1 mediates cardiac fibrosis in experimental postinfarct myocardium mice model. J. Cell. Physiol. 2019, 234, 2997–3006. [Google Scholar] [CrossRef]
- Gast, M.; Rauch, B.; Haghikia, A.; Nakagawa, S.; Haas, J.; Stroux, A.; Schmidt, D.; Schumann, P.; Weiss, S.; Jensen, L.; et al. Long noncoding RNA NEAT1 modulates immune cell functions and is suppressed in early onset myocardial infarction patients. Cardiovasc. Res. 2019, 115, 1886–1906. [Google Scholar] [CrossRef]
- Liu, L.; An, X.; Li, Z.; Song, Y.; Li, L.; Zuo, S.; Liu, N.; Yang, G.; Wang, H.; Cheng, X.; et al. The H19 long noncoding RNA is a novel negative regulator of cardiomyocyte hypertrophy. Cardiovasc. Res. 2016, 111, 56–65. [Google Scholar] [CrossRef]
- Zhu, Y.; Feng, Z.; Jian, Z.; Xiao, Y. Long noncoding RNA TUG1 promotes cardiac fibroblast transformation to myofibroblasts via miR-29c in chronic hypoxia. Mol. Med. Rep. 2018, 18, 3451–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seal, R.L.; Chen, L.L.; Griffiths-Jones, S.; Lowe, T.M.; Mathews, M.B.; O’Reilly, D.; Pierce, A.J.; Stadler, P.F.; Ulitsky, I.; Wolin, S.L.; et al. A guide to naming human non-coding RNA genes. EMBO J. 2020, 39, e103777. [Google Scholar] [CrossRef] [PubMed]
- Cambier, L.; Lacampagne, A.; Auffray, C.; Pomiès, P. Fem1a is a mitochondrial protein up-regulated upon ischemia-reperfusion injury. FEBS Lett. 2009, 583, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Gupta, I.; Singh, K.; Varshney, N.K.; Khan, S. Delineating crosstalk mechanisms of the ubiquitin proteasome system that regulate apoptosis. Front. Cell Dev. Biol. 2018, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Kalfon, R.; Friedman, T.; Eliachar, S.; Shofti, R.; Haas, T.; Koren, L.; Moskovitz, J.D.; Hai, T.; Aronheim, A. JDP2 and ATF3 deficiencies dampen maladaptive cardiac remodeling and preserve cardiac function. PLoS ONE 2019, 14, e0213081. [Google Scholar] [CrossRef] [PubMed]
- Blevins, M.A.; Huang, M.; Zhao, R. The role of CtBP1 in oncogenic processes and its potential as a therapeutic target. Mol. Cancer Ther. 2017, 16, 981–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mailloux, R.J.; Gardiner, D.; O’Brien, M. 2-Oxoglutarate dehydrogenase is a more significant source of O2(-)/H2O2 than pyruvate dehydrogenase in cardiac and liver tissue. Free. Radic. Biol. Med. 2016, 97, 501–512. [Google Scholar] [CrossRef]
- Olmedo, I.; Pino, G.; Riquelme, J.A.; Aranguiz, P.; Díaz, M.C.; López-Crisosto, C.; Lavandero, S.; Donoso, P.; Pedrozo, Z.; Sánchez, G. Inhibition of the proteasome preserves Mitofusin-2 and mitochondrial integrity, protecting cardiomyocytes during ischemia-reperfusion injury. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165659. [Google Scholar] [CrossRef]
- Wu, Y.; Xia, Y.; Li, P.; Qu, H.-Q.; Liu, Y.; Yang, Y.; Lin, J.; Zheng, M.; Tian, L.; Wu, Z.; et al. Role of the ADCY9 gene in cardiac abnormalities of the Rubinstein-Taybi syndrome. Orphanet J. Rare Dis. 2020, 15, 101–110. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Xu, Y.; Wang, B.; Jia, C.; Wang, L.; Sun, H.; Zhao, H.; Wang, Z.; Zou, Q.; et al. lncRNA PCAT19 negatively regulates p53 in non-small cell lung cancer. Oncol. Lett. 2019, 18, 6795–6800. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, M.J.; Mukhopadhyay, S.; Hoofnagle, M.; Chabasse, C.; Sarkar, R. Tumor suppressor protein p53 negatively regulates ischemia-induced angiogenesis and arteriogenesis. J. Vasc. Surg. 2018, 68, 222S–233S.e1. [Google Scholar] [CrossRef]
- Ounzain, S.; Micheletti, R.; Arnan, C.; Plaisance, I.; Cecchi, D.; Schroen, B.; Reverter, F.; Alexanian, M.; Gonzales, C.; Ng, S.-Y.; et al. CARMEN, a human super enhancer-associated long noncoding RNA controlling cardiac specification, differentiation and homeostasis. J. Mol. Cell. Cardiol. 2015, 89, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Coppiello, G.; Collantes, M.; Sirerol-Piquer, M.S.; Vandenwijngaert, S.; Schoors, S.; Swinnen, M.; Vandersmissen, I.; Herijgers, P.; Topal, B.; Van Loon, J.; et al. Meox2/Tcf15 heterodimers program the heart capillary endothelium for cardiac fatty acid uptake. Circulation 2015, 131, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Kurotsu, S.; Osakabe, R.; Isomi, M.; Tamura, F.; Sadahiro, T.; Muraoka, N.; Kojima, H.; Haginiwa, S.; Tani, H.; Nara, K.; et al. Distinct expression patterns of Flk1 and Flt1 in the coronary vascular system during development and after myocardial infarction. Biochem. Biophys. Res. Commun. 2018, 495, 884–891. [Google Scholar] [CrossRef]
- Travisano, S.I.; Oliveira, V.L.; Prados, B.; Grego-Bessa, J.; Piñeiro-Sabarís, R.; Bou, V.; Gomez, M.J.; Sanchez-Cabo, F.; MacGrogan, M. Coronary arterial development is regulated by a Dll4-Jag1-EphrinB2 signaling cascade. eLife 2019, 8, e49977. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Belyaeva, O.V.; Korkina, O.V.; Stetsenko, A.V.; Kedishvili, N.Y. Human retinol dehydrogenase 13 (RDH13) is a mitochondrial short-chain dehydrogenase/reductase with a retinaldehyde reductase activity. FEBS J. 2007, 275, 138–147. [Google Scholar] [CrossRef]
- Mendler, L.; Braun, T.; Muller, S. The ubiquitin-Like SUMO system and heart function: From development to disease. Circ. Res. 2018, 118, 1524–4571. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.C. The MicroRNA biology of the mammalian nucleus. Mol. Ther. Nucleic Acids 2014, 3, e188. [Google Scholar] [CrossRef]
- Xie, Y.-H.; Hu, J. Suppression of long non-coding RNA PCAT19 inhibits glioma cell proliferation and invasion, and increases cell apoptosis through regulation of MELK targeted by miR-142-5p. Genes Genom. 2020, 42, 1299–1310. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Guo, J.; Zhang, W. lncRNA PCAT19 promotes the proliferation of laryngocarcinoma cells via modulation of the miR-182/PDK4 axis. J. Cell Biochem. 2019, 120, 12810–12821. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The reference human genome annotation for The ENCODE project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Tuteja, G. TissueEnrich: Tissue-specific gene enrichment analysis. Bioinformatics 2019, 35, 1966–1967. [Google Scholar] [CrossRef]
- Hardwick, S.; Chen, W.Y.; Wong, T.; Deveson, I.; Blackburn, J.; Andersen, S.B.; Nielsen, S.B.A.L.K.; Mattick, J.S.; Mercer, T. Spliced synthetic genes as internal controls in RNA sequencing experiments. Nat. Methods 2016, 13, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.-P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef] [PubMed]
- Siepel, A.; Bejerano, G.; Pedersen, J.S.; Hinrichs, A.S.; Hou, M.; Rosenbloom, K.; Clawson, H.; Spieth, J.; Hillier, L.W.; Richards, S.; et al. Evolutionarily conserved elements in vertebrate, insect, worm, and yeast genomes. Genome Res. 2005, 15, 1034–1050. [Google Scholar] [CrossRef] [Green Version]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2016, 4. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative Name | Chr’me | Start | Stop | Strand | # Exons | Mean TPM |
|---|---|---|---|---|---|---|
| VTCN1-AS | chr1 | 117128696 | 117143589 | + | 2 | 1.68 |
| LINC02934 | chr2 | 37489457 | 37605898 | + | 3 | 0.22 |
| LINC02935 | chr3 | 15894181 | 16137554 | + | 4 | 0.32 |
| ADCY5-AS | chr3 | 123335278 | 123338361 | + | 3 | 0.47 |
| LINC02936 | chr5 | 107778856 | 107781422 | + | 2 | 0.34 |
| LINC02937 | chr6 | 157328269 | 157363141 | + | 4 | 0.37 |
| LINC02938 | chr8 | 94223489 | 94228144 | − | 3 | 0.30 |
| PDGFD-AS | chr11 | 104071819 | 104093201 | + | 4 | 0.33 |
| DHRS1-AS | chr14 | 24271210 | 24299055 | + | 3 | 2.03 |
| RWDD3-DT | chr1 | 95247358 | 95256066 | + | 2 | 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ward, Z.; Schmeier, S.; Saddic, L.; Sigurdsson, M.I.; Cameron, V.A.; Pearson, J.; Miller, A.; Morley-Bunker, A.; Gorham, J.; Seidman, J.G.; et al. Novel and Annotated Long Noncoding RNAs Associated with Ischemia in the Human Heart. Int. J. Mol. Sci. 2021, 22, 11324. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111324
Ward Z, Schmeier S, Saddic L, Sigurdsson MI, Cameron VA, Pearson J, Miller A, Morley-Bunker A, Gorham J, Seidman JG, et al. Novel and Annotated Long Noncoding RNAs Associated with Ischemia in the Human Heart. International Journal of Molecular Sciences. 2021; 22(21):11324. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111324
Chicago/Turabian StyleWard, Zoe, Sebastian Schmeier, Louis Saddic, Martin I. Sigurdsson, Vicky A. Cameron, John Pearson, Allison Miller, Arthur Morley-Bunker, Josh Gorham, Jonathan G. Seidman, and et al. 2021. "Novel and Annotated Long Noncoding RNAs Associated with Ischemia in the Human Heart" International Journal of Molecular Sciences 22, no. 21: 11324. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111324