
Prevalence and Characteristics of Phenicol-Oxazolidinone Resistance Genes in Enterococcus Faecalis and Enterococcus Faecium Isolated from Food-Producing Animals and Meat in Korea

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Identification of Phenicol-Oxazolidinone Resistant Gene
2.2. Antimicrobial-Resistant Pattern
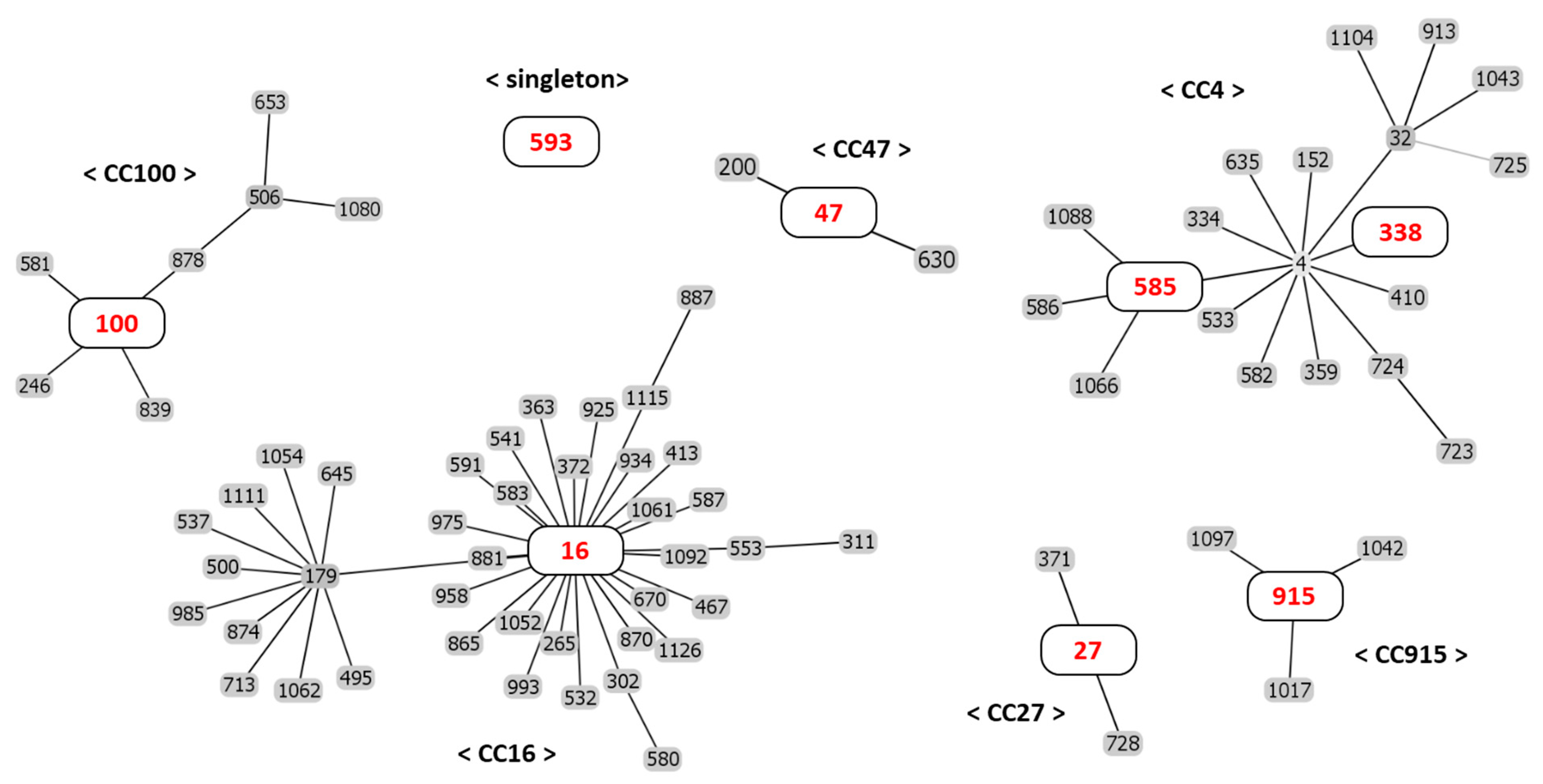
2.3. Molecular Typing by Multilocus Sequence Typing (MLST)
2.4. Transferability of Phenicol-Oxazolidinone Resistance Genes
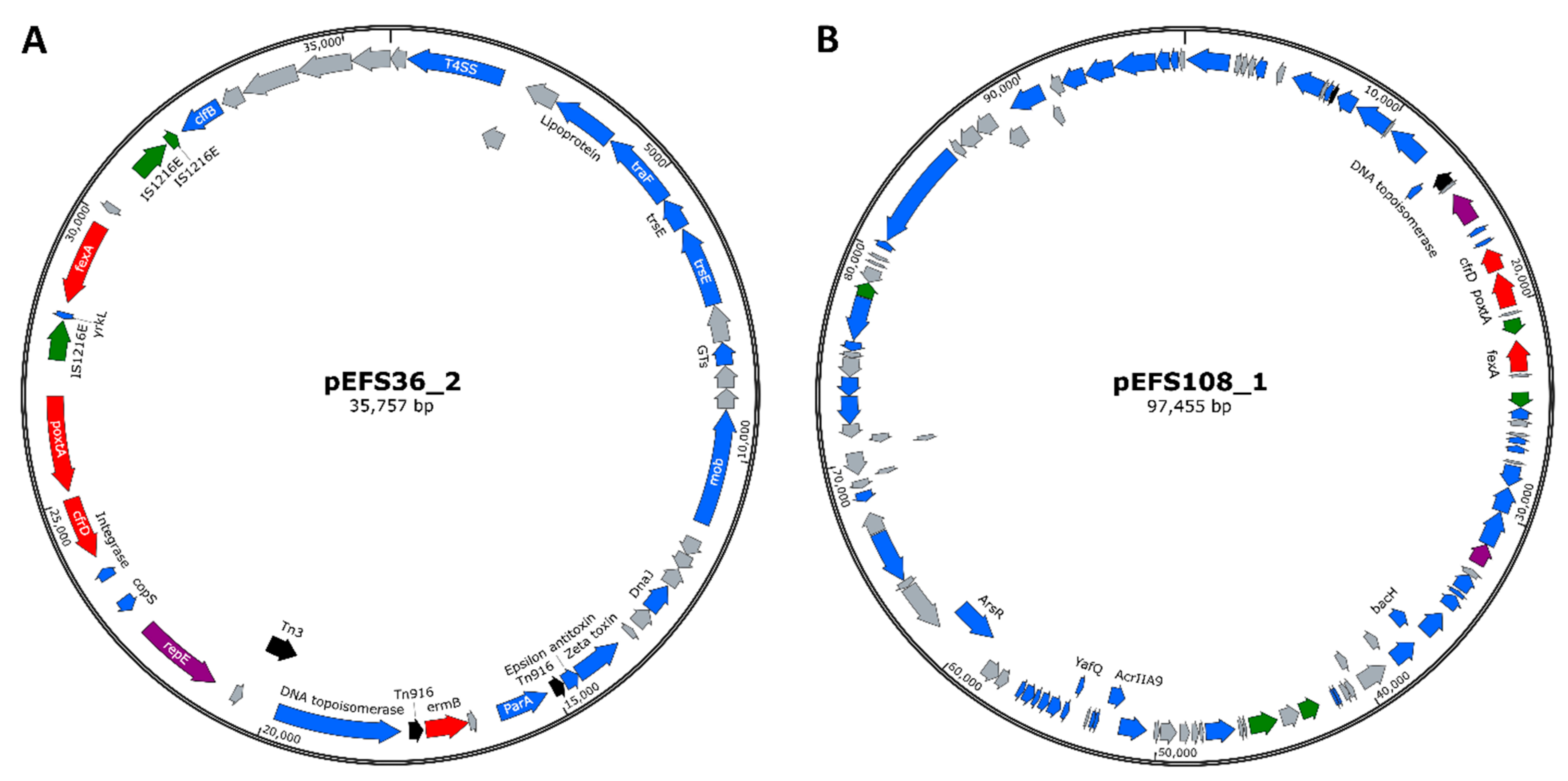
2.5. Complete Genome Sequencing
2.5.1. Genome Features of Three Isolates
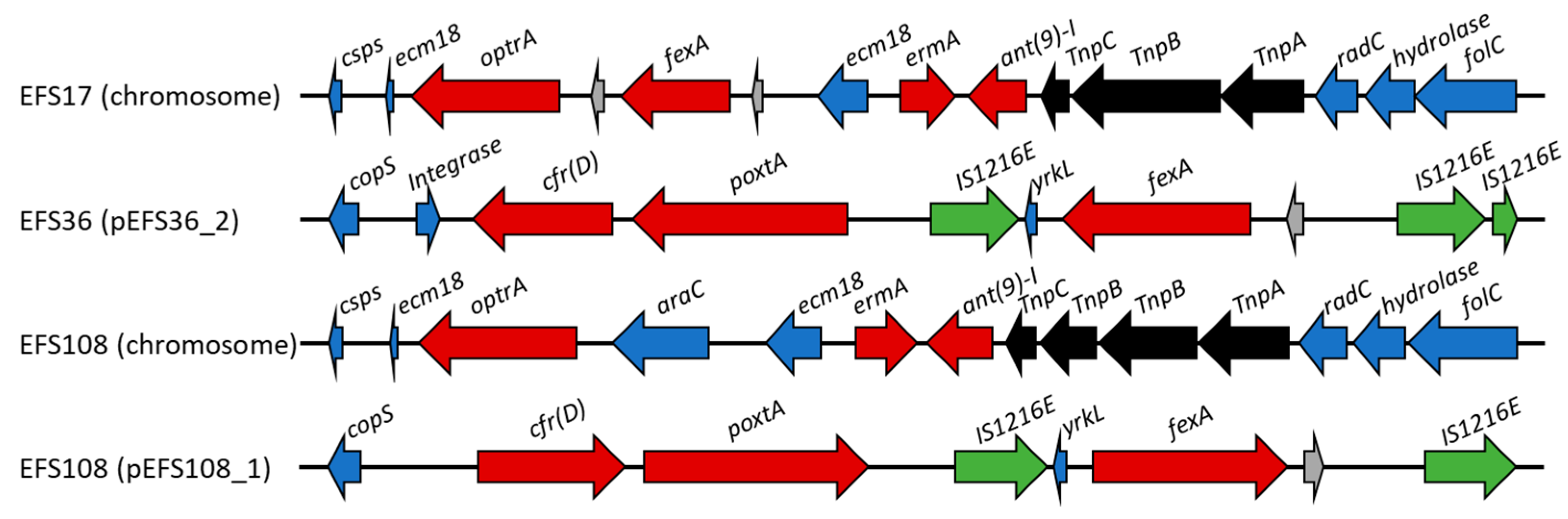
2.5.2. In Silico Identification of Antimicrobial Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Enterococcus Strains
4.2. Screening of Phenicol-Oxazolidinone Resistance Gene by Polymerase Chain Reaction (PCR)
4.3. Antimicrobial Susceptibility Testing
4.4. MLST Analysis
4.5. Conjugation
4.6. Complete Genome Sequencing
4.6.1. Genome Sequencing, Assembly, and Annotation
4.6.2. Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Na, S.H.; Moon, D.C.; Choi, M.J.; Oh, S.J.; Jung, D.Y.; Kang, H.Y.; Hyun, B.H.; Lim, S.K. Detection of oxazolidinone and phenicol resistant enterococcal isolates from duck feces and carcasses. Int. J. Food Microbiol. 2019, 293, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, F.; Bai, B.; Lin, Z.; Xu, G.; Chen, Z.; Sun, X.; Zheng, J.; Deng, Q.; Yu, Z. Linezolid Resistance in Enterococcus faecalis Associated With Urinary Tract Infections of Patients in a Tertiary Hospitals in China: Resistance Mechanisms, Virulence, and Risk Factors. Front. Public Health 2021, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Tamang, M.D.; Moon, D.C.; Kim, S.R.; Kang, H.Y.; Lee, K.; Nam, H.M.; Jang, G.C.; Lee, H.S.; Jung, S.C.; Lim, S.K. Detection of novel oxazolidinone and phenicol resistance gene optrA in enterococcal isolates from food animals and animal carcasses. Vet. Microbiol. 2017, 201, 252–256. [Google Scholar] [CrossRef]
- Bi, R.; Qin, T.; Fan, W.; Ma, P.; Gu, B. The emerging problem of linezolid-resistant enterococci. J. Glob. Antimicrob. Resist. 2018, 13, 11–19. [Google Scholar] [CrossRef]
- Bender, J.K.; Fleige, C.; Klare, I.; Werner, G. Development of a multiplex-PCR to simultaneously detect acquired linezolid resistance genes cfr, optrA and poxtA in enterococci of clinical origin. J. Microbiol. Methods 2019, 160, 101–103. [Google Scholar] [CrossRef]
- Yang, X.X.; Tian, T.T.; Qiao, W.; Tian, Z.; Yang, M.; Zhang, Y.; Li, J.Y. Prevalence and characterization of oxazolidinone and phenicol cross-resistance gene optrA in enterococci obtained from anaerobic digestion systems treating swine manure. Environ. Pollut. 2020, 267, 115540. [Google Scholar] [CrossRef]
- Hua, R.; Xia, Y.; Wu, W.; Yang, M.; Yan, J. Molecular epidemiology and mechanisms of 43 low-level linezolid-resistant enterococcus faecalis strains in Chongqing, China. Ann. Lab. Med. 2018, 39, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.H.; Cha, M.H.; Woo, G.J.; Chi, Y.M. Characterization of oxazolidinone and phenicol resistance genes in non-clinical enterococcal isolates from Korea. J. Glob. Antimicrob. Resist. 2021, 24, 363–369. [Google Scholar] [CrossRef]
- Egan, S.A.; Shore, A.C.; O’Connell, B.; Brennan, G.I.; Coleman, D.C. Linezolid resistance in Enterococcus faecium and Enterococcus faecalis from hospitalized patients in Ireland: High prevalence of the MDR genes optrA and poxtA in isolates with diverse genetic backgrounds. J. Antimicrob. Chemother. 2020, 75, 1704–1711. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Shan, X.; Li, D.; Schwarz, S.; Zhang, S.M.; Li, X.S.; Du, X.D. Analysis of a poxtA- And optrA-co-carrying conjugative multiresistance plasmid from Enterococcus faecalis. J. Antimicrob. Chemother. 2019, 74, 1771–1775. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Jeong, Y.S.; Chang, J.; Sung, H.; Kim, M.N. Emergence of optrA-mediated linezolid-nonsusceptible enterococcus faecalis in a tertiary care hospital. Ann. Lab. Med. 2020, 40, 322–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute of Food and Drug Safety Evaluation; Ministry of Agriculture, Food and Rural Affairs; Animal and Plant Quarantine Agency; Korea Animal Health Products Association. National Antibiotic Use and Resistance Monitoring in 2019. 2020. Available online: https://impfood.mfds.go.kr/CFBCC02F02/getCntntsDetail?cntntsSn=325548&cntntsMngId=00006 (accessed on 17 October 2021).
- Kim, M.H.; Moon, D.C.; Kim, S.J.; Mechesso, A.F.; Song, H.J.; Kang, H.Y.; Choi, J.H.; Yoon, S.S.; Lim, S.K. Nationwide surveillance on antimicrobial resistance profiles of enterococcus faecium and enterococcus faecalis isolated from healthy food animals in South Korea, 2010 to 2019. Microorganisms 2021, 9, 925. [Google Scholar] [CrossRef]
- Yoon, S.; Son, S.H.; Kim, Y.B.; Seo, K.W.; Lee, Y.J. Molecular characteristics of optrA-carrying Enterococcus faecalis from chicken meat in South Korea. Poult. Sci. 2020, 99, 6990–6996. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.H.; Yoon, S.; Kim, K.; Kim, Y.B.; Lee, Y.J. Comparative analysis of chloramphenicol-resistant enterococcus faecalis isolated from dairy companies in korea. Vet. Sci. 2021, 8, 143. [Google Scholar] [CrossRef]
- Kang, Z.Z.; Lei, C.W.; Kong, L.H.; Wang, Y.L.; Ye, X.L.; Ma, B.H.; Wang, X.C.; Li, C.; Zhang, Y.; Wang, H.N. Detection of transferable oxazolidinone resistance determinants in Enterococcus faecalis and Enterococcus faecium of swine origin in Sichuan Province, China. J. Glob. Antimicrob. Resist. 2019, 19, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Maasjost, J.; Mühldorfer, K.; De Jäckel, S.C.; Hafez, H.M. Antimicrobial susceptibility patterns of enterococcus faecalis and enterococcus faecium isolated from poultry flocks in Germany. Avian Dis. 2015, 59, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Na, S.H.; Moon, D.C.; Kim, M.H.; Kang, H.Y.; Kim, S.J.; Choi, J.H.; Mechesso, A.F.; Yoon, S.S.; Lim, S.K. Detection of the phenicol–oxazolidinone resistance gene poxta in enterococcus faecium and enterococcus faecalis from food-producing animals during 2008–2018 in Korea. Microorganisms 2020, 8, 1839. [Google Scholar] [CrossRef]
- Freitas, A.R.; Tedim, A.P.; Novais, C.; Lanza, V.F.; Peixe, L. Comparative genomics of global optrA-carrying Enterococcus faecalis uncovers a common chromosomal hotspot for optrA acquisition within a diversity of core and accessory genomes. Microb. Genom. 2020, 6, e000350. [Google Scholar] [CrossRef]
- de Jong, A.; Simjee, S.; El Garch, F.; Moyaert, H.; Rose, M.; Youala, M.; Dry, M. Antimicrobial susceptibility of enterococci recovered from healthy cattle, pigs and chickens in nine EU countries (EASSA Study) to critically important antibiotics. Vet. Microbiol. 2018, 216, 168–175. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.; Shen, Y.; Schwarz, S.; Cai, J.; Lv, Y.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J.; et al. Genetic environment of the transferable oxazolidinone/phenicol resistance gene optrA in Enterococcus faecalis isolates of human and animal origin. J. Antimicrob. Chemother. 2016, 71, 1466–1473. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Wang, X.; Yin, Y.; Li, S.; Zhang, Y.; Wang, Q.; Wang, H. Molecular characteristics of oxazolidinone resistance in enterococci from a multicenter study in China. BMC Microbiol. 2019, 19, 162. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S. Florfenicol-chloramphenicol exporter gene fexA is part of the novel transposon Tn558. Antimicrob. Agents Chemother. 2005, 49, 813–815. [Google Scholar] [CrossRef] [Green Version]
- Shan, X.; Li, X.S.; Wang, N.; Schwarz, S.; Zhang, S.M.; Li, D.; Du, X.D. Studies on the role of IS1216E in the formation and dissemination of poxtA-carrying plasmids in an Enterococcus faecium clade A1 isolate. J. Antimicrob. Chemother. 2020, 75, 3126–3130. [Google Scholar] [CrossRef]
- Darini, A.L.C.; Palepou, M.F.I.; Woodford, N. Effects of the movement of insertion sequences on the structure of vana glycopeptide resistance elements in Enterococcus faecium. Antimicrob. Agents Chemother. 2000, 44, 1362–1364. [Google Scholar] [CrossRef] [Green Version]
- Raze, D.; Dardenne, O.; Hallut, S.; Martinez-Bueno, M.; Coyette, J.; Ghuysen, J.M. The gene encoding the low-affinity penicillin-binding protein 3r in Enterococcus hirae S185R is borne on a plasmid carrying other antibiotic resistance determinants. Antimicrob. Agents Chemother. 1998, 42, 534–539. [Google Scholar] [CrossRef] [Green Version]
- Ciric, L.; Brouwer, M.S.M.; Mullany, P.; Roberts, A.P. Minocycline resistance in an oral Streptococcus infantis isolate is encoded by tet(S) on a novel small, low copy number plasmid. FEMS Microbiol. Lett. 2014, 353, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; CLSI: Wayne, PA, USA, 2014; Volume 32, ISBN 1562388975. [Google Scholar]
- Ruiz-Garbajosa, P.; Bonten, M.J.M.; Robinson, D.A.; Top, J.; Nallapareddy, S.R.; Torres, C.; Coque, T.M.; Cantón, R.; Baquero, F.; Murray, B.E.; et al. Multilocus sequence typing scheme for Enterococcus faecalis reveals hospital-adapted genetic complexes in a background of high rates of recombination. J. Clin. Microbiol. 2006, 44, 2220–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljahdali, N.H.; Khajanchi, B.K.; Weston, K.; Deck, J.; Cox, J.; Singh, R.; Gilbert, J.; Sanad, Y.M.; Han, J.; Nayak, R.; et al. Genotypic and phenotypic characterization of incompatibility group fib positive salmonella enterica serovar typhimurium isolates from food animal sources. Genes 2020, 11, 1307. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Hasman, H. PlasmidFinder and In Silico pMLST: Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). In Methods in Molecular Biology; H. Humana: New York, NY, USA, 2020; Volume 2075, pp. 285–294. [Google Scholar]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Source | Detected Resistance Genes by PCR | Transconjugants | |||||
|---|---|---|---|---|---|---|---|---|---|
| optrA | poxtA | cfr | fexA | optrA | poxtA | fexA | |||
| E. faecalis | EFS17 | Pork, meat | + | − | − | + | − | − | − |
| E. faecalis | EFS24 | Pork, meat | + | − | − | + | + | − | + |
| E. faecalis | EFS27 | Pork, meat | + | − | − | + | + | − | + |
| E. faecalis | EFS36 | Beef, meat | − | + | − | + | − | + | + |
| E. faecalis | EFS74 | Beef, meat | − | + | − | − | − | + | − |
| E. faecalis | EFS98 | Pig, slaughterhouse | − | + | − | + | − | − | − |
| E. faecalis | EFS99 | Pig, slaughterhouse | − | + | − | + | − | − | + |
| E. faecalis | EFS108 | Pig, slaughterhouse | + | + | − | + | − | + | + |
| E. faecalis | EFS117 | Cattle, slaughterhouse | + | − | − | − | − | − | − |
| E. faecalis | EFS147 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS151 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS153 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS154 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS158 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS253 | Pig, slaughterhouse | + | − | − | + | + | − | + |
| E. faecalis | EFS255 | Cattle, slaughterhouse | − | + | − | − | − | + | − |
| E. faecalis | EFS268 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS269 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS270 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecalis | EFS271 | Cattle, farm | + | − | − | + | + | − | + |
| E. faecium | EFM21 | Cattle, farm | − | + | − | − | − | − | − |
| E. faecium | EFM262 | Cattle, farm | − | + | − | − | − | − | − |
| Scheme 1. | Strain | Antimicrobial Resistance Pattern 1 | MIC 2 (mg/L) | |
|---|---|---|---|---|
| LZD | FFN | |||
| E. faecalis | EFS17 | TET, CIP, ERY, TYLT, LZD, GEN, KAN, STR, CHL, FFN | 8 | 64 |
| E. faecalis | EFS24 | TET, ERY, TYLT, LZD, GEN, KAN, STR, CHL, FFN | 8 | 64 |
| E. faecalis | EFS27 | TET, ERY, TYLT, KAN, STR, CHL, FFN | 4 | 64 |
| E. faecalis | EFS36 | ERY, TYLT, GEN, KAN, STR, CHL, FFN | 2 | 64 |
| E. faecalis | EFS74 | TET, FFN | 4 | 16 |
| E. faecalis | EFS98 | TET, ERY, TYLT, STR, FFN | 4 | 64 |
| E. faecalis | EFS99 | TET, ERY, TYLT, STR, FFN | 4 | 16 |
| E. faecalis | EFS108 | TET, ERY, TYLT, CHL, FFN | 2 | 64 |
| E. faecalis | EFS117 | TET, DAP, ERY, TYLT, FFN | − | 16 |
| E. faecalis | EFS147 | TET, ERY, TYLT, CHL, FFN | − | 32 |
| E. faecalis | EFS151 | TET, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS153 | TET, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS154 | TET, ERY, TYLT, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS158 | TET, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS253 | TET, ERY, TYLT, LZD, GEN, KAN, STR, CHL, FFN | 8 | 64 |
| E. faecalis | EFS255 | TET, ERY, TYLT, FFN | − | 16 |
| E. faecalis | EFS268 | TET, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS269 | TET, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS270 | TET, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecalis | EFS271 | TET, TGC, ERY, TYLT, KAN, STR, CHL, FFN | − | 32 |
| E. faecium | EFM21 | TET, CIP, DAP, ERY, TYLT, KAN, STR, CHL, FFN | 4 | 32 |
| E. faecium | EFM262 | TET, TGC, CIP, ERY, TYLT, KAN, STR, AMP, CHL, FFN | − | 32 |
| Strain | Allele | ST 1 | CC 2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| gdh | gyd | pstS | gki | aroE | xpt | yiqL | |||
| EFS17 | 8 | 7 | 7 | 4 | 4 | 4 | 1 | 585 | 4 |
| EFS24 | 5 | 1 | 1 | 3 | 7 | 7 | 6 | 16 | 16 |
| EFS27 | 39 | 2 | 49 | 45 | 7 | 2 | 17 | 915 | 915 |
| EFS36 | 8 | 7 | 7 | 5 | 4 | 14 | 1 | 338 | 4 |
| EFS74 | 19 | 1 | 24 | 22 | 19 | 17 | 14 | 47 | 47 |
| EFS98 | 34 | 2 | 17 | 37 | 29 | 23 | 6 | 100 | 100 |
| EFS99 | 34 | 2 | 17 | 37 | 29 | 23 | 6 | 100 | 100 |
| EFS108 | 3 | 2 | 7 | 10 | 10 | 2 | 7 | 27 | 27 |
| EFS117 | 5 | 1 | 1 | 3 | 7 | 7 | 6 | 16 | 16 |
| EFS147 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS151 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS153 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS154 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS158 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS253 | 5 | 1 | 1 | 3 | 7 | 7 | 6 | 16 | 16 |
| EFS255 | 34 | 2 | 17 | 37 | 29 | 23 | 6 | 100 | 100 |
| EFS268 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS269 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS270 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| EFS271 | 14 | 2 | 17 | 1 | 3 | 3 | 17 | 593 | Singleton |
| Genomic Features | EFS17 | EFS36 | EFS108 |
|---|---|---|---|
| Genome size (bp) | 2,914,315 | 3,095,754 | 3,093,936 |
| Chromosome size (bp) | 2,838,954 | 3,016,592 | 2,816,588 |
| Number of plasmids | 1 | 2 | 5 |
| Size range of plasmid(s) (bp) | 75,361 | 35,757–43,405 | 5120–97,455 |
| GC content (%) | 37.4 | 37.3 | 37.3 |
| Number of genes | 2876 | 3135 | 3053 |
| Number of coding genes | 2803 | 3057 | 2980 |
| Coding genes in chromosome | 2712 | 2964 | 2670 |
| Coding genes in plasmid(s) | 91 | 93 | 310 |
| Number of RNAs | 73 | 78 | 73 |
| Strain | Class | Antimicrobial Resistance Genes |
|---|---|---|
| EFS17 | aminocyclitol | ant(9)-Ia |
| aminoglycoside | aac(6′)-aph(2″), aph(3′)-III, str, ant(6)-Ia | |
| fluoroquinolone | parC | |
| folate pathway antagonist | dfrG | |
| lincosamide | lsaA, lsaE, lnuB, ermA, ermB | |
| macrolide | ermA, ermB | |
| oxazolidinone | optrA | |
| phenicol | fexA, optrA, cat | |
| pleuromutilin | lsaE | |
| streptogramin a | lsaA, lsaE | |
| streptogramin b | ermA, ermB | |
| tetracycline | tetM, tetL | |
| EFS36 | aminoglycoside | aac(6′)-aph(2″), aph(3′)-III, ant(6)-Ia |
| lincosamide | lsaE, cfr(D), lnuB, lsaA, ermB | |
| macrolide | ermB | |
| oxazolidinone | cfr(D), poxtA | |
| phenicol | fexA, poxtA, cfr(D) | |
| pleuromutilin | lsaE, cfr(D) | |
| streptogramin a | lsaA, lsaE, cfr(D) | |
| streptogramin b | ermB | |
| tetracycline | poxtA | |
| EFS108 | aminocyclitol | ant(9)-Ia |
| lincosamide | ermA, ermB, lsaA | |
| macrolide | ermA, ermB | |
| oxazolidinone | optrA, cfr(D), poxtA | |
| phenicol | fexA, optrA, cat | |
| streptogramin a | lsaA | |
| streptogramin b | ermA, ermB | |
| tetracycline | tetM, tetL |
| Target | Primer | Nucleotide Sequence (5′→3′) | Amplicon (bp) | Reference |
|---|---|---|---|---|
| cfr | cfr-F | TGC TAC AGG CGA CAT TGG AT | 137 | [9] |
| cfr-R | GAC GGT TGG CTA GAG CTT CA | |||
| optrA | optrA-F | ACC GGT GTC CTC TTT GTC AG | 369 | [9] |
| optrA-R | TCA ATG GAG TTA CGA TCG CCT T | |||
| poxtA | poxtA-F | TCA GAG CCG TAC TGA GCA AC | 167 | [9] |
| poxtA-R | CGT TTC TGG GTC AAG GTG GT | |||
| fexA | fexA-F | GTA CTT GTA GGT GCA ATT ACG GCT GA | 1272 | [24] |
| fexA-R | CGC ATC TGA GTA GGA CAT AGC GTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.; Shin, S.-W.; Kwak, H.-S.; Cha, M.-H.; Yang, S.-M.; Gwak, Y.-S.; Woo, G.-J.; Kim, H.-Y. Prevalence and Characteristics of Phenicol-Oxazolidinone Resistance Genes in Enterococcus Faecalis and Enterococcus Faecium Isolated from Food-Producing Animals and Meat in Korea. Int. J. Mol. Sci. 2021, 22, 11335. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111335
Kim E, Shin S-W, Kwak H-S, Cha M-H, Yang S-M, Gwak Y-S, Woo G-J, Kim H-Y. Prevalence and Characteristics of Phenicol-Oxazolidinone Resistance Genes in Enterococcus Faecalis and Enterococcus Faecium Isolated from Food-Producing Animals and Meat in Korea. International Journal of Molecular Sciences. 2021; 22(21):11335. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111335
Chicago/Turabian StyleKim, Eiseul, So-Won Shin, Hyo-Sun Kwak, Min-Hyeok Cha, Seung-Min Yang, Yoon-Soo Gwak, Gun-Jo Woo, and Hae-Yeong Kim. 2021. "Prevalence and Characteristics of Phenicol-Oxazolidinone Resistance Genes in Enterococcus Faecalis and Enterococcus Faecium Isolated from Food-Producing Animals and Meat in Korea" International Journal of Molecular Sciences 22, no. 21: 11335. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111335