
The Collagen Receptor uPARAP in Malignant Mesothelioma: A Potential Diagnostic Marker and Therapeutic Target

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
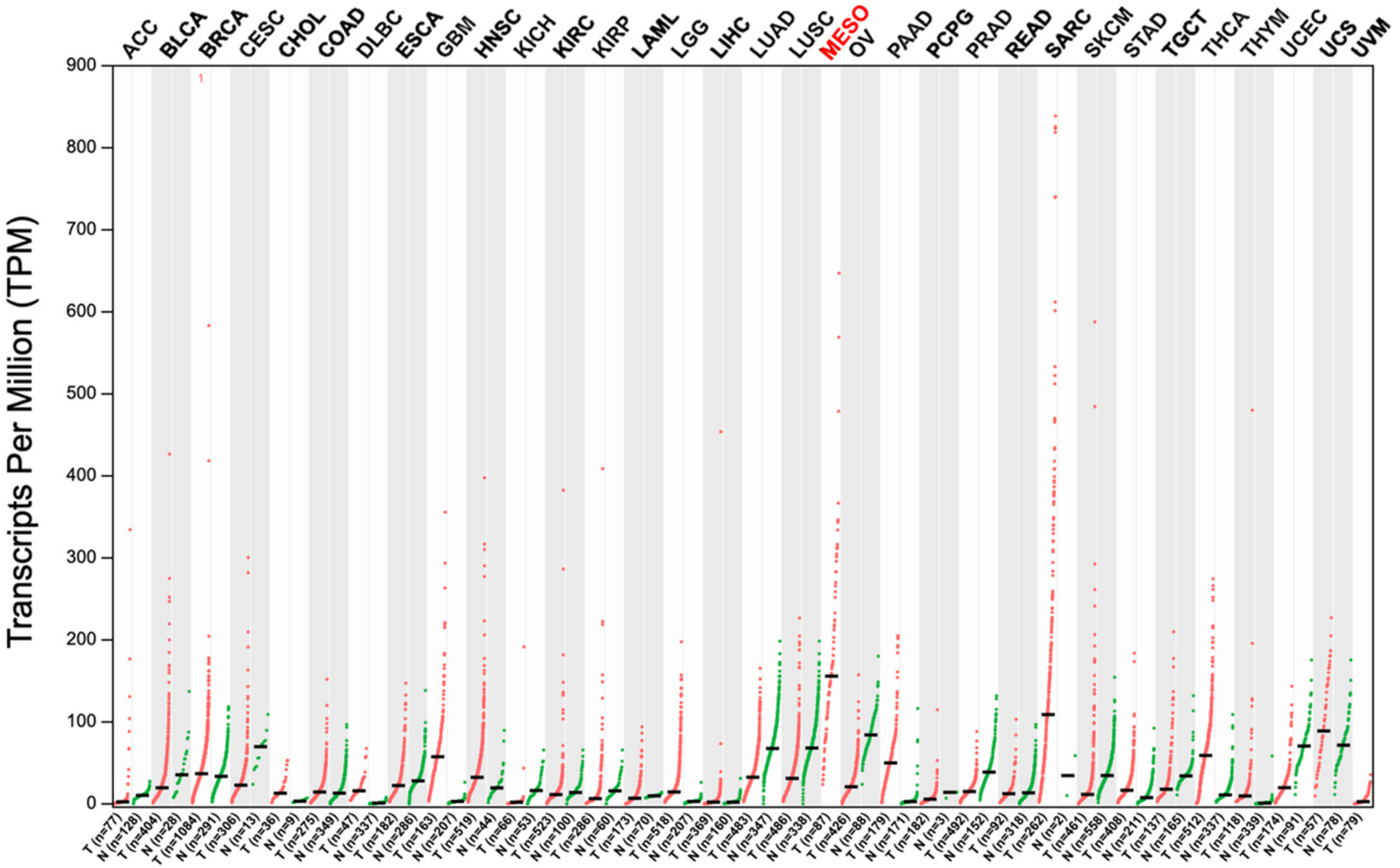
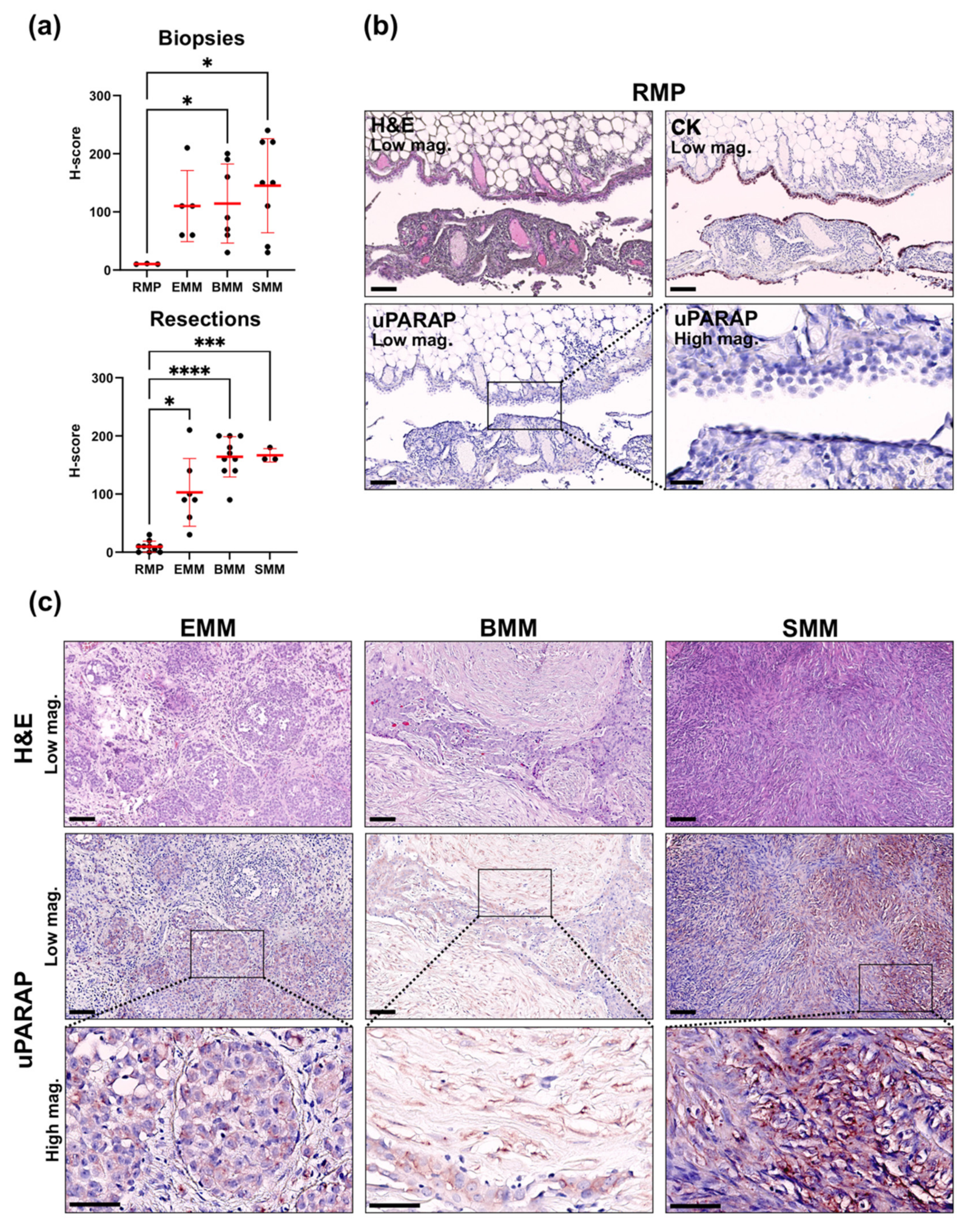
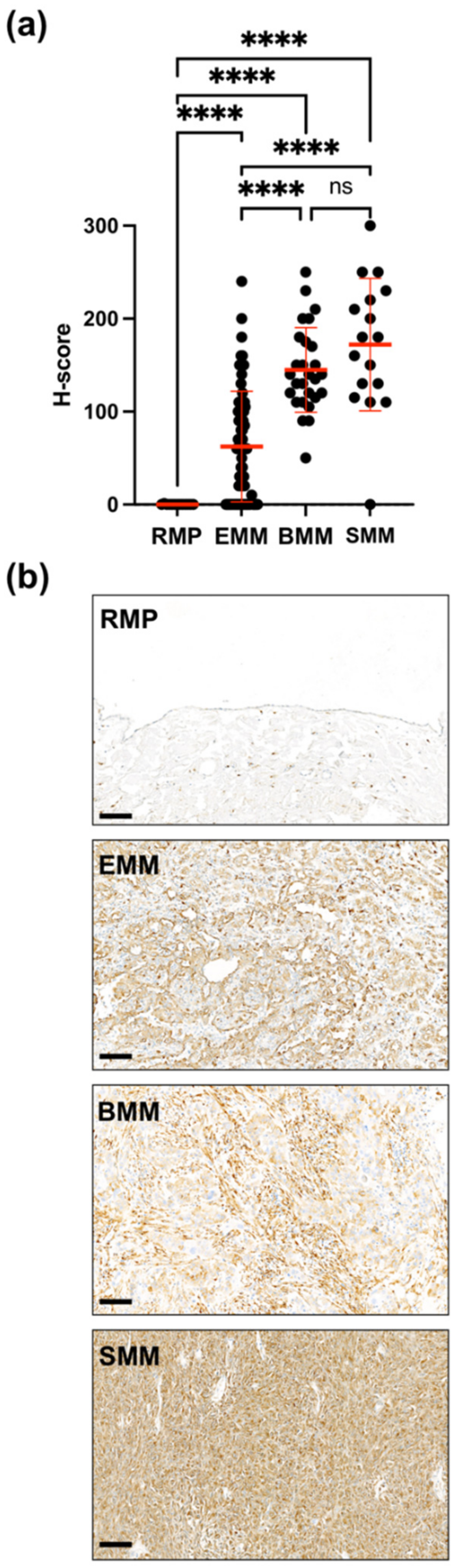
2.1. Upregulation of uPARAP in Malignant Mesothelioma
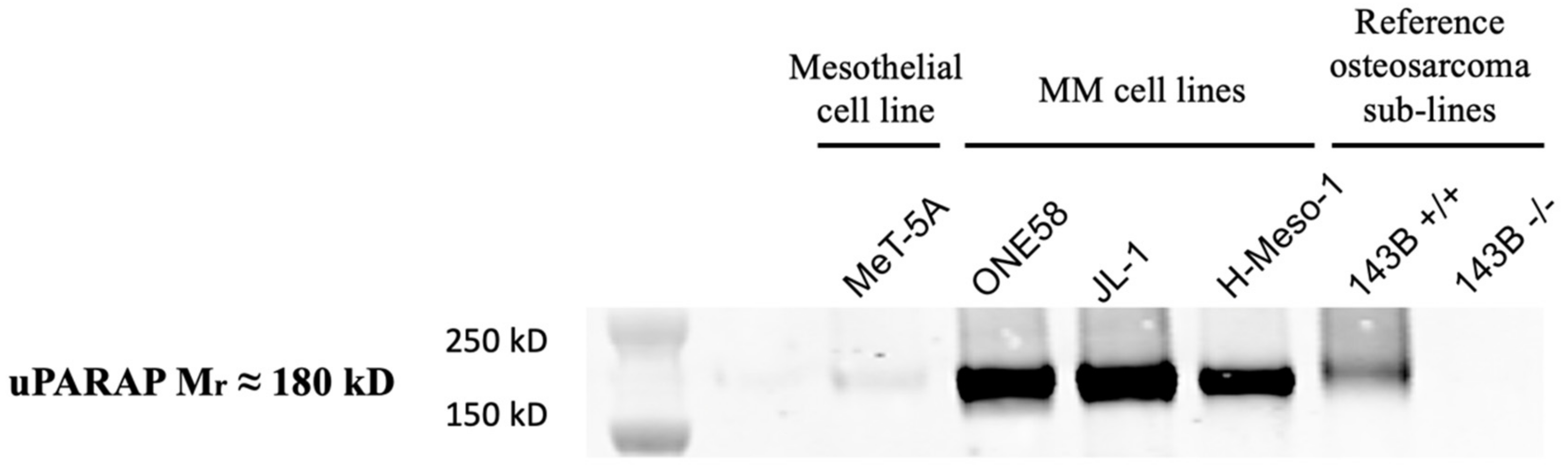
2.2. uPARAP Expression in Mesothelioma Cell Lines
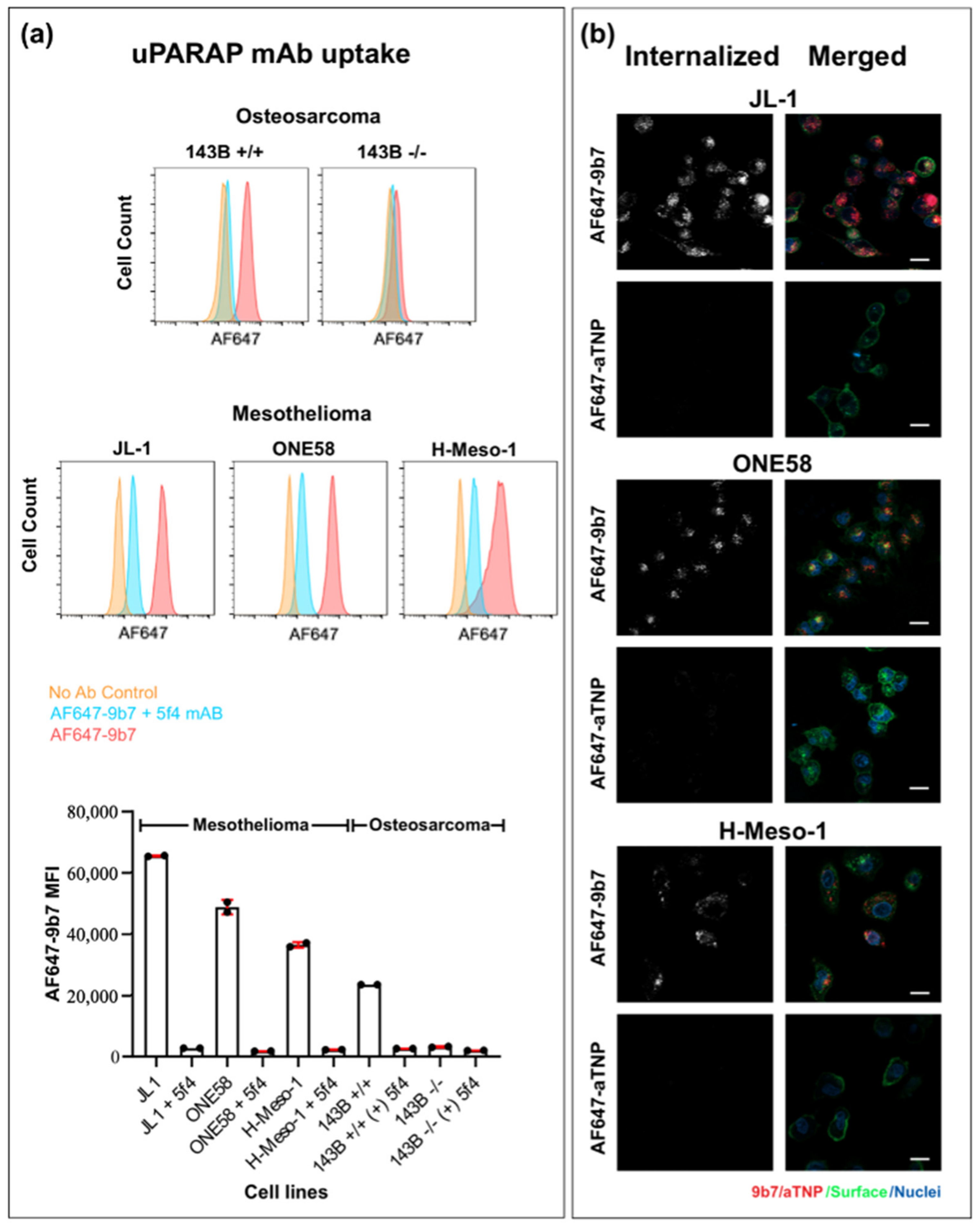
2.3. Internalization of a uPARAP-Directed Antibody in Malignant Mesothelioma Cells
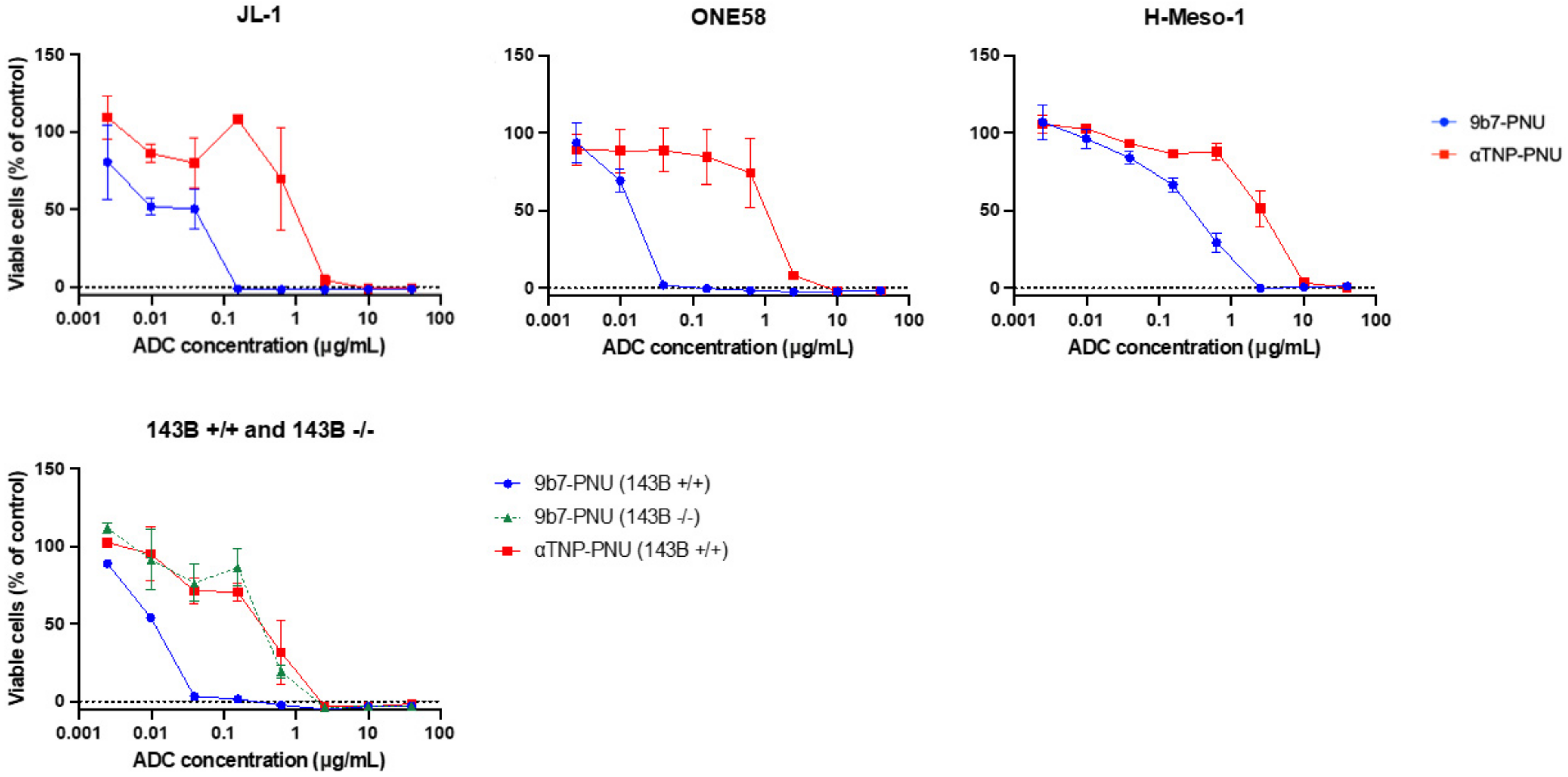
2.4. uPARAP-Specific Eradication of MM Cells with a uPARAP-Directed ADC
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Antibodies, Fluorescence Labeling and Preparation of ADCs
4.3. Tissue Samples
4.4. Histology and Immunohistochemistry
4.5. Evaluation of Staining
4.6. Western Blot
4.7. Confocal Microscopy
4.8. Flow Cytometry
4.9. Assay for Cellular Sensitivity to ADCs
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carbone, M.; Adusumilli, P.S.; Alexander, H.R.; Baas, P.; Bardelli, F.; Bononi, A.; Bueno, R.; Felley-Bosco, E.; Galateau-Salle, F.; Jablons, D.; et al. Mesothelioma: Scientific Clues for Prevention, Diagnosis, and Therapy. CA Cancer J. Clin. 2019, 69, 402–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cakiroglu, E.; Senturk, S. Genomics and Functional Genomics of Malignant Pleural Mesothelioma. Int. J. Mol. Sci. 2020, 21, 6342. [Google Scholar] [CrossRef] [PubMed]
- Scherpereel, A.; Wallyn, F.; Albelda, S.M.; Munck, C. Novel Therapies for Malignant Pleural Mesothelioma. Lancet Oncol. 2018, 19, e161–e172. [Google Scholar] [CrossRef]
- Baas, P.; Scherpereel, A.; Nowak, A.K.; Fujimoto, N.; Peters, S.; Tsao, A.S.; Mansfield, A.S.; Popat, S.; Jahan, T.; Antonia, S.; et al. First-Line Nivolumab plus Ipilimumab in Unresectable Malignant Pleural Mesothelioma (CheckMate 743): A Multicentre, Randomised, Open-Label, Phase 3 Trial. Lancet 2021, 397, 375–386. [Google Scholar] [CrossRef]
- Hu, Z.I.; Ghafoor, A.; Sengupta, M.; Hassan, R. Malignant Mesothelioma: Advances in Immune Checkpoint Inhibitor and Mesothelin-Targeted Therapies. Cancer 2021, 127, 1010–1020. [Google Scholar] [CrossRef]
- Hassan, R.; Blumenschein, G.R.; Moore, K.N.; Santin, A.D.; Kindler, H.L.; Nemunaitis, J.J.; Seward, S.M.; Thomas, A.; Kim, S.K.; Rajagopalan, P.; et al. First-in-Human, Multicenter, Phase I Dose-Escalation and Expansion Study of Anti-Mesothelin Antibody-Drug Conjugate Anetumab Ravtansine in Advanced or Metastatic Solid Tumors. J. Clin. Oncol. 2020, 38, 1824–1835. [Google Scholar] [CrossRef]
- Clarke, J.; Chu, S.-C.; Siu, L.L.; Machiels, J.-P.; Markman, B.; Heinhuis, K.; Millward, M.; Lolkema, M.; Patel, S.P.; de Souza, P.; et al. Abstract B057: BMS-986148, an Anti-Mesothelin Antibody-Drug Conjugate (ADC), Alone or in Combination with Nivolumab Demonstrates Clinical Activity in Patients with Select Advanced Solid Tumors. Mol. Cancer Ther. 2019, 18, B057. [Google Scholar] [CrossRef]
- Scales, S.J.; Gupta, N.; Pacheco, G.; Firestein, R.; French, D.M.; Koeppen, H.; Rangell, L.; Barry-Hamilton, V.; Luis, E.; Chuh, J.; et al. An Antimesothelin-Monomethyl Auristatin E Conjugate with Potent Antitumor Activity in Ovarian, Pancreatic, and Mesothelioma Models. Mol. Cancer Ther. 2014, 13, 2630–2640. [Google Scholar] [CrossRef] [Green Version]
- Weekes, C.D.; Lamberts, L.E.; Borad, M.J.; Voortman, J.; McWilliams, R.R.; Diamond, J.R.; de Vries, E.G.E.; Verheul, H.M.; Lieu, C.H.; Kim, G.P.; et al. Phase I Study of DMOT4039A, an Antibody–Drug Conjugate Targeting Mesothelin, in Patients with Unresectable Pancreatic or Platinum-Resistant Ovarian Cancer. Mol. Cancer Ther. 2016, 15, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Chia, P.-L.; Parakh, S.; Tsao, M.-S.; Pham, N.-A.; Gan, H.K.; Cao, D.; Burvenich, I.J.G.; Rigopoulos, A.; Reilly, E.B.; John, T.; et al. Targeting and Efficacy of Novel MAb806-Antibody-Drug Conjugates in Malignant Mesothelioma. Pharmaceuticals 2020, 13, E289. [Google Scholar] [CrossRef]
- Patnaik, A.; Meric-Bernstam, F.; Rocha Lima, C.M.S.P.; Robert, F.; Dowlati, A.; Kindler, H.L.; Davar, D.; Powell, S.F.; Garfin, P.M.; Balmanoukian, A.S. SGN228-001: A Phase I Open-Label Dose-Escalation, and Expansion Study of SGN-CD228A in Select Advanced Solid Tumors. JCO 2020, 38, TPS3652. [Google Scholar] [CrossRef]
- Criscitiello, C.; Morganti, S.; Curigliano, G. Antibody–Drug Conjugates in Solid Tumors: A Look into Novel Targets. J. Hematol. Oncol. 2021, 14, 20. [Google Scholar] [CrossRef]
- Madsen, D.H.; Ingvarsen, S.; Jürgensen, H.J.; Melander, M.C.; Kjøller, L.; Moyer, A.; Honoré, C.; Madsen, C.A.; Garred, P.; Burgdorf, S.; et al. The Non-Phagocytic Route of Collagen Uptake: A Distinct Degradation Pathway. J. Biol. Chem. 2011, 286, 26996–27010. [Google Scholar] [CrossRef] [Green Version]
- Engelholm, L.H.; List, K.; Netzel-Arnett, S.; Cukierman, E.; Mitola, D.J.; Aaronson, H.; Kjøller, L.; Larsen, J.K.; Yamada, K.M.; Strickland, D.K.; et al. UPARAP/Endo180 Is Essential for Cellular Uptake of Collagen and Promotes Fibroblast Collagen Adhesion. J. Cell Biol. 2003, 160, 1009–1015. [Google Scholar] [CrossRef]
- Melander, M.C.; Jürgensen, H.J.; Madsen, D.H.; Engelholm, L.H.; Behrendt, N. The Collagen Receptor UPARAP/Endo180 in Tissue Degradation and Cancer (Review). Int. J. Oncol. 2015, 47, 1177–1188. [Google Scholar] [CrossRef] [Green Version]
- Engelholm, L.H.; Ingvarsen, S.; Jürgensen, H.J.; Hillig, T.; Madsen, D.H.; Nielsen, B.S.; Behrendt, N. The Collagen Receptor UPARAP/Endo180. Front. Biosci. 2009, 14, 2103–2114. [Google Scholar] [CrossRef]
- Nielsen, C.F.; van Putten, S.M.; Lund, I.K.; Melander, M.C.; Nørregaard, K.S.; Jürgensen, H.J.; Reckzeh, K.; Christensen, K.R.; Ingvarsen, S.Z.; Gårdsvoll, H.; et al. The Collagen Receptor UPARAP/Endo180 as a Novel Target for Antibody-Drug Conjugate Mediated Treatment of Mesenchymal and Leukemic Cancers. Oncotarget 2017, 8, 44605–44624. [Google Scholar] [CrossRef]
- Hmeljak, J.; Sanchez-Vega, F.; Hoadley, K.A.; Shih, J.; Stewart, C.; Heiman, D.; Tarpey, P.; Danilova, L.; Drill, E.; Gibb, E.A.; et al. Integrative Molecular Characterization of Malignant Pleural Mesothelioma. Cancer Discov. 2018, 8, 1548–1565. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Program—National Cancer Institute. Available online: https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga (accessed on 22 August 2021).
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A Web Server for Cancer and Normal Gene Expression Profiling and Interactive Analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- GEPIA. Available online: http://gepia.cancer-pku.cn/detail.php?gene=MRC2 (accessed on 15 June 2021).
- Husain, A.N.; Colby, T.V.; Ordóñez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement from the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Sulek, J.; Wagenaar-Miller, R.A.; Shireman, J.; Molinolo, A.; Madsen, D.H.; Engelholm, L.H.; Behrendt, N.; Bugge, T.H. Increased Expression of the Collagen Internalization Receptor UPARAP/Endo180 in the Stroma of Head and Neck Cancer. J. Histochem. Cytochem. 2007, 55, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Madsen, D.H.; Engelholm, L.H.; Ingvarsen, S.; Hillig, T.; Wagenaar-Miller, R.A.; Kjøller, L.; Gårdsvoll, H.; Høyer-Hansen, G.; Holmbeck, K.; Bugge, T.H.; et al. Extracellular Collagenases and the Endocytic Receptor, Urokinase Plasminogen Activator Receptor-Associated Protein/Endo180, Cooperate in Fibroblast-Mediated Collagen Degradation. J. Biol. Chem. 2007, 282, 27037–27045. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, B.S.; Rank, F.; Engelholm, L.H.; Holm, A.; Danø, K.; Behrendt, N. Urokinase Receptor-Associated Protein (UPARAP) Is Expressed in Connection with Malignant as Well as Benign Lesions of the Human Breast and Occurs in Specific Populations of Stromal Cells. Int. J. Cancer 2002, 98, 656–664. [Google Scholar] [CrossRef]
- Galateau-Salle, F.; Churg, A.; Roggli, V.; Travis, W.D.; World Health Organization Committee for Tumors of the Pleura. The 2015 World Health Organization Classification of Tumors of the Pleura: Advances since the 2004 Classification. J. Thorac. Oncol. 2016, 11, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Schulte, J.J.; Chapel, D.B.; Attanoos, R.; Brcic, L.; Burn, J.; Butnor, K.J.; Chang, N.; Chen, H.; Dacic, S.; De Perrot, M.; et al. Comparison of Nuclear Grade, Necrosis, and Histologic Subtype Between Biopsy and Resection in Pleural Malignant Mesothelioma. Am. J. Clin. Pathol. 2021, 156, 989–999. [Google Scholar] [CrossRef]
- List, K.; Høyer-Hansen, G.; Rønne, E.; Danø, K.; Behrendt, N. Different Mechanisms Are Involved in the Antibody Mediated Inhibition of Ligand Binding to the Urokinase Receptor: A Study Based on Biosensor Technology. This Work Was Supported by the Danish Cancer Society. J. Immunol. Methods 1999, 222, 125–133. [Google Scholar] [CrossRef]
- Holte, D.; Lyssikatos, J.P.; Valdiosera, A.M.; Swinney, Z.; Sisodiya, V.; Sandoval, J.; Lee, C.; Aujay, M.A.; Tchelepi, R.B.; Hamdy, O.M.; et al. Evaluation of PNU-159682 Antibody Drug Conjugates (ADCs). Bioorgan. Med. Chem. Lett. 2020, 30, 127640. [Google Scholar] [CrossRef]
- Engelholm, L.H.; Melander, M.C.; Hald, A.; Persson, M.; Madsen, D.H.; Jürgensen, H.J.; Johansson, K.; Nielsen, C.; Nørregaard, K.S.; Ingvarsen, S.Z.; et al. Targeting a Novel Bone Degradation Pathway in Primary Bone Cancer by Inactivation of the Collagen Receptor UPARAP/Endo180. J. Pathol. 2016, 238, 120–133. [Google Scholar] [CrossRef]
- Tada, Y.; Suzuki, T.; Shimada, H.; Hiroshima, K.; Tatsumi, K.; Tagawa, M. Molecular-Targeted Therapy for Malignant Mesothelioma. PLEURA 2015, 2, 2373997515600403. [Google Scholar] [CrossRef] [Green Version]
- Joubert, N.; Beck, A.; Dumontet, C.; Denevault-Sabourin, C. Antibody–Drug Conjugates: The Last Decade. Pharmaceuticals 2020, 13, 245. [Google Scholar] [CrossRef]
- Jin, Y.; Schladetsch, M.A.; Huang, X.; Balunas, M.J.; Wiemer, A.J. Stepping Forward in Antibody-Drug Conjugate Development. Pharmacol. Ther. 2021, 107917. [Google Scholar] [CrossRef] [PubMed]
- Affatato, R.; Mendogni, P.; Del Gobbo, A.; Ferrero, S.; Ricci, F.; Broggini, M.; Rosso, L. Establishment and Characterization of Patient-Derived Xenografts (PDXs) of Different Histology from Malignant Pleural Mesothelioma Patients. Cancers 2020, 12, 3846. [Google Scholar] [CrossRef] [PubMed]
- Ingvarsen, S.Z.; Gårdsvoll, H.; van Putten, S.; Nørregaard, K.S.; Krigslund, O.; Meilstrup, J.A.; Tran, C.; Jürgensen, H.J.; Melander, M.C.; Nielsen, C.H.; et al. Tumor Cell MT1-MMP Is Dispensable for Osteosarcoma Tumor Growth, Bone Degradation and Lung Metastasis. Sci. Rep. 2020, 10, 19138. [Google Scholar] [CrossRef] [PubMed]
- Lund, I.K.; Jögi, A.; Rønø, B.; Rasch, M.G.; Lund, L.R.; Almholt, K.; Gårdsvoll, H.; Behrendt, N.; Rømer, J.; Høyer-Hansen, G. Antibody-Mediated Targeting of the Urokinase-Type Plasminogen Activator Proteolytic Function Neutralizes Fibrinolysis In Vivo. J. Biol. Chem. 2008, 283, 32506–32515. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, A.G.; Sauter, J.L.; Nowak, A.K.; Kindler, H.L.; Gill, R.R.; Remy-Jardin, M.; Armato, S.G.; Fernandez-Cuesta, L.; Bueno, R.; Alcala, N.; et al. EURACAN/IASLC Proposals for Updating the Histologic Classification of Pleural Mesothelioma: Towards a More Multidisciplinary Approach. J. Thorac. Oncol. 2020, 15, 29–49. [Google Scholar] [CrossRef]
- Rosen, L.E.; Karrison, T.; Ananthanarayanan, V.; Gallan, A.J.; Adusumilli, P.S.; Alchami, F.S.; Attanoos, R.; Brcic, L.; Butnor, K.J.; Galateau-Sallé, F.; et al. Nuclear Grade and Necrosis Predict Prognosis in Malignant Epithelioid Pleural Mesothelioma: A Multi-Institutional Study. Mod. Pathol. 2018, 31, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Detre, S.; Saclani Jotti, G.; Dowsett, M. A “Quickscore” Method for Immunohistochemical Semiquantitation: Validation for Oestrogen Receptor in Breast Carcinomas. J. Clin. Pathol. 1995, 48, 876–878. [Google Scholar] [CrossRef] [Green Version]
- Zimling, Z.G.; Sørensen, J.B.; Gerds, T.A.; Bech, C.; Andersen, C.B.; Santoni-Rugiu, E. Low ERCC1 Expression in Malignant Pleural Mesotheliomas Treated with Cisplatin and Vinorelbine Predicts Prolonged Progression-Free Survival. J. Thorac. Oncol. 2012, 7, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, J.N.; Santoni-Rugiu, E.; Sørensen, J.B. Changes in Epidermal Growth Factor Receptor Expression during Chemotherapy in Non-Small Cell Lung Cancer. Cancer Chemother. Pharmacol. 2014, 73, 131–137. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ADC | ONE58, EC50 (95% CI) | JL-1, EC50 (95% CI) | H-Meso-1, EC50 (95% CI) | 143B +/+, EC50 (95% CI) | 143B -/-, EC50 (95% CI) |
|---|---|---|---|---|---|
| 9b7-PNU | 0.014 (0.011–0.016) | 0.055 (0.021–0.090) | 0.26 (0.174–0.339) | 0.012 (0.011–0.013) | 0.40 (0.233–0.558) |
| aTNP-PNU | 1.5 (0.749–2.212) | 1.1 (0.587–1.648) | 2.6 (1.842–3.370) | 0.33 (0.129–0.528) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çakılkaya, P.; Sørensen, R.R.; Jürgensen, H.J.; Krigslund, O.; Gårdsvoll, H.; Nielsen, C.F.; Santoni-Rugiu, E.; Behrendt, N.; Engelholm, L.H. The Collagen Receptor uPARAP in Malignant Mesothelioma: A Potential Diagnostic Marker and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 11452. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111452
Çakılkaya P, Sørensen RR, Jürgensen HJ, Krigslund O, Gårdsvoll H, Nielsen CF, Santoni-Rugiu E, Behrendt N, Engelholm LH. The Collagen Receptor uPARAP in Malignant Mesothelioma: A Potential Diagnostic Marker and Therapeutic Target. International Journal of Molecular Sciences. 2021; 22(21):11452. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111452
Chicago/Turabian StyleÇakılkaya, Pınar, Rikke Raagaard Sørensen, Henrik Jessen Jürgensen, Oliver Krigslund, Henrik Gårdsvoll, Christoffer F. Nielsen, Eric Santoni-Rugiu, Niels Behrendt, and Lars H. Engelholm. 2021. "The Collagen Receptor uPARAP in Malignant Mesothelioma: A Potential Diagnostic Marker and Therapeutic Target" International Journal of Molecular Sciences 22, no. 21: 11452. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111452