
Effects of GLP-1RA and SGLT2i, Alone or in Combination, on Mouse Models of Type 2 Diabetes Representing Different Disease Stages

 ,
,
Abstract
:1. Introduction
2. Results
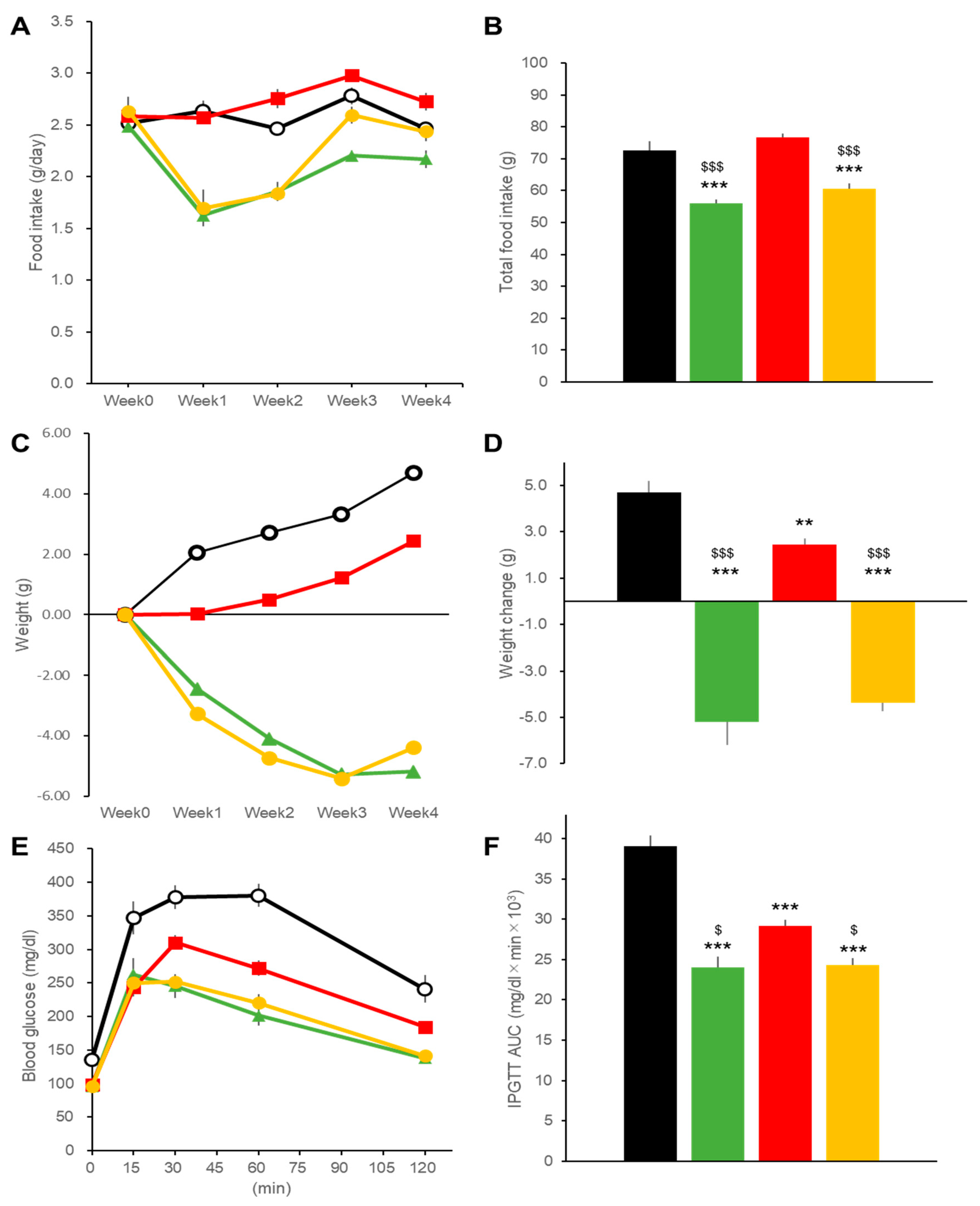
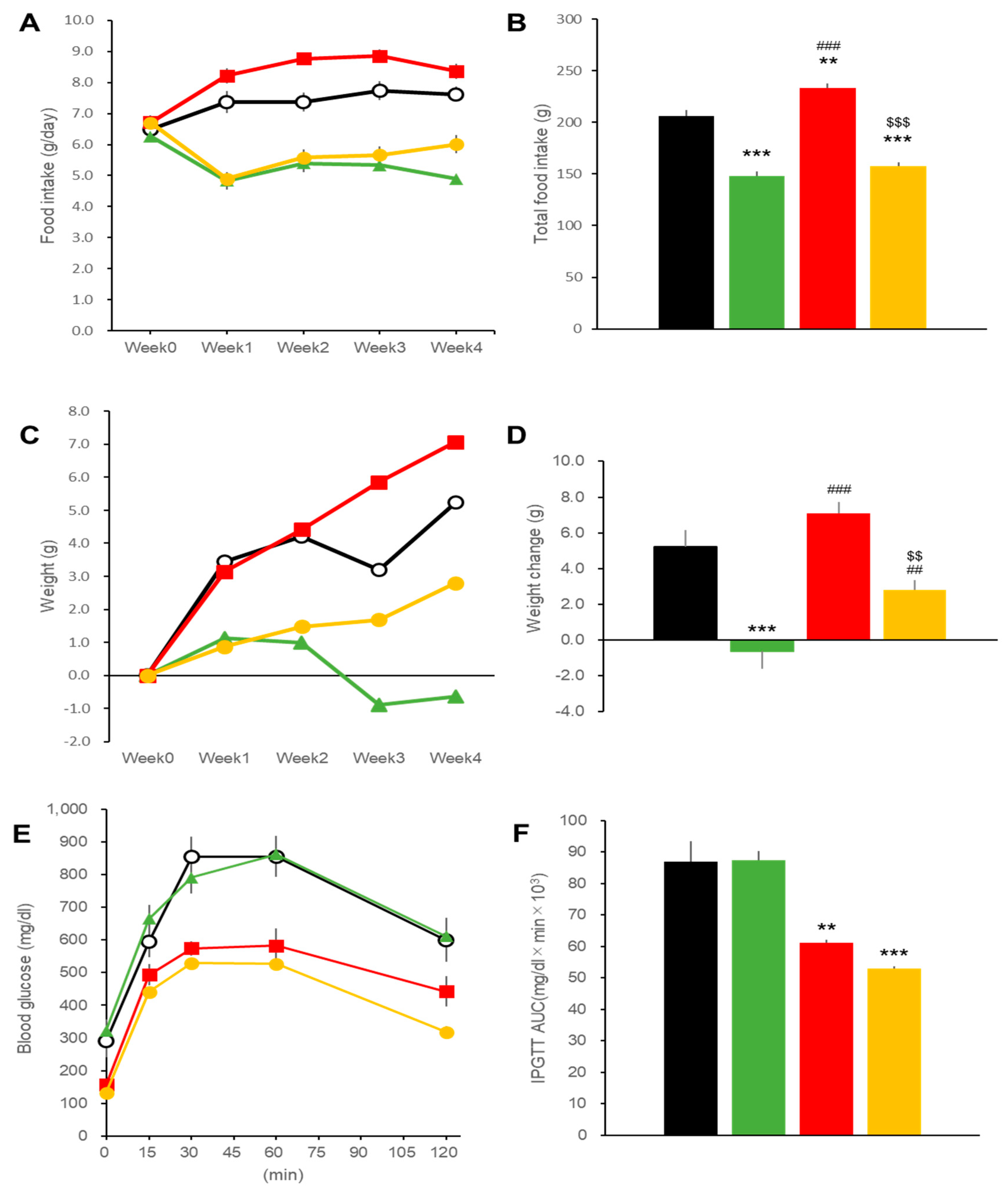
2.1. Effects on Body Weight and Blood Glucose in DIO Mice
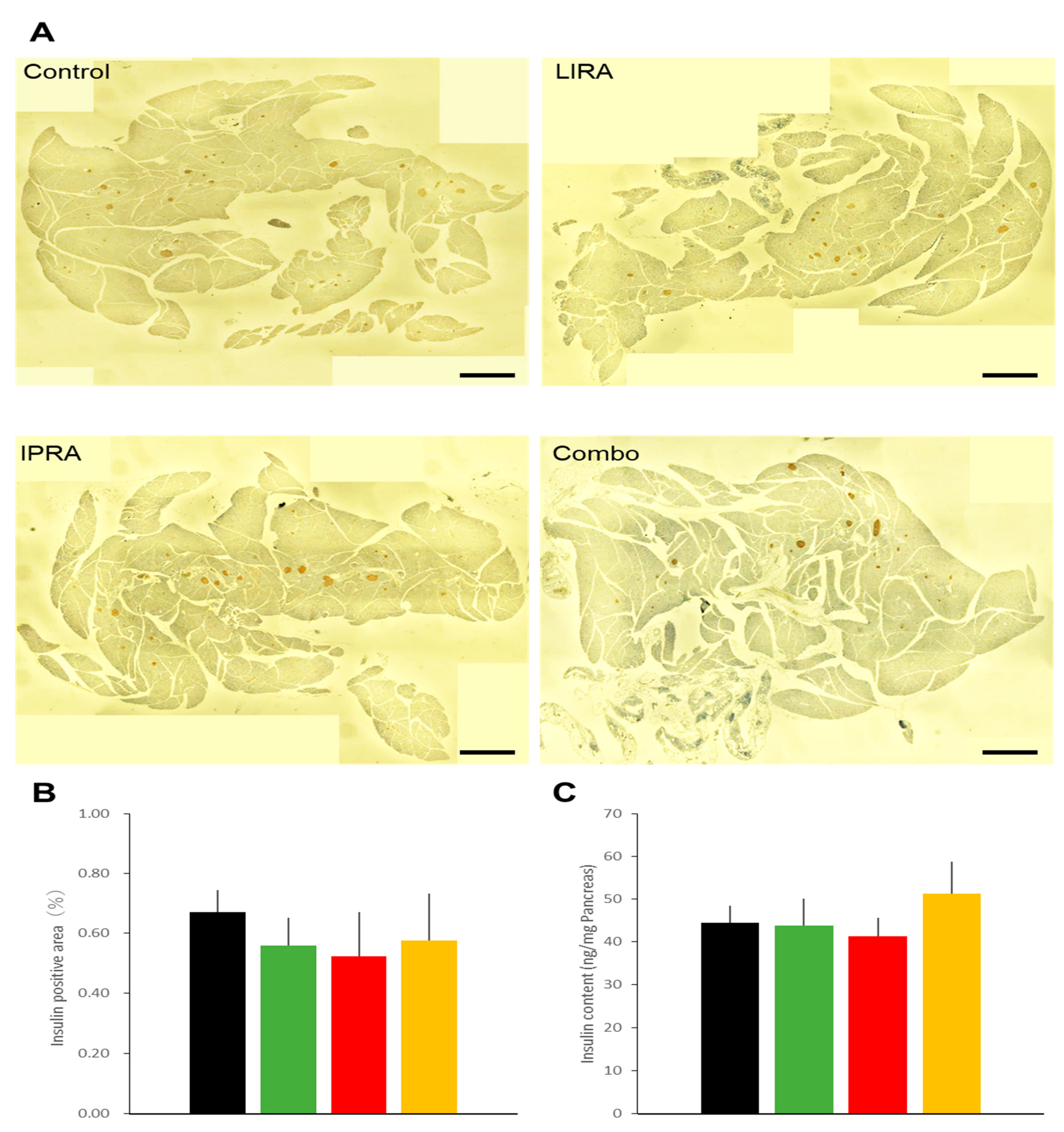
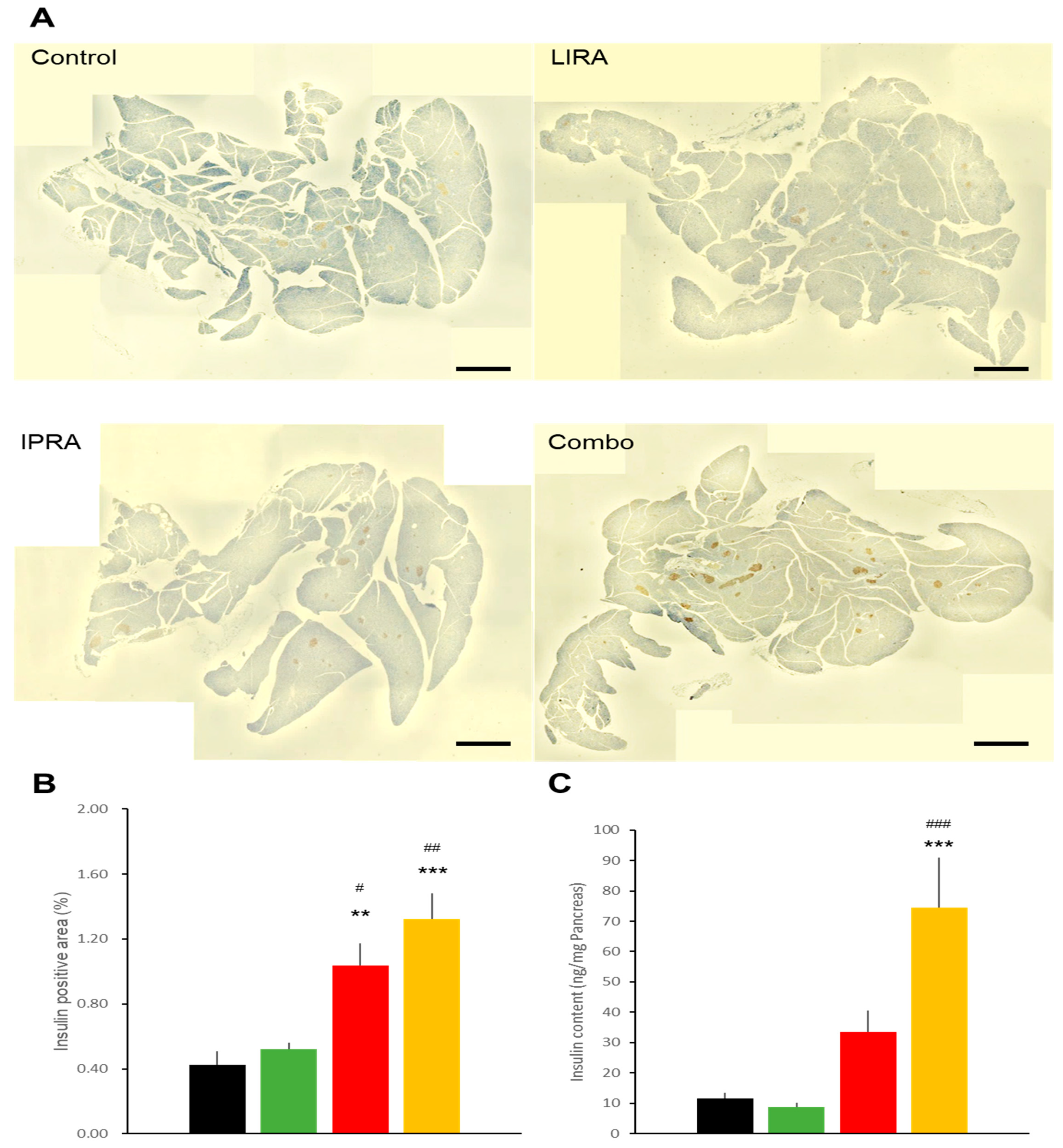
2.2. Pancreatic Effects in DIO Mice
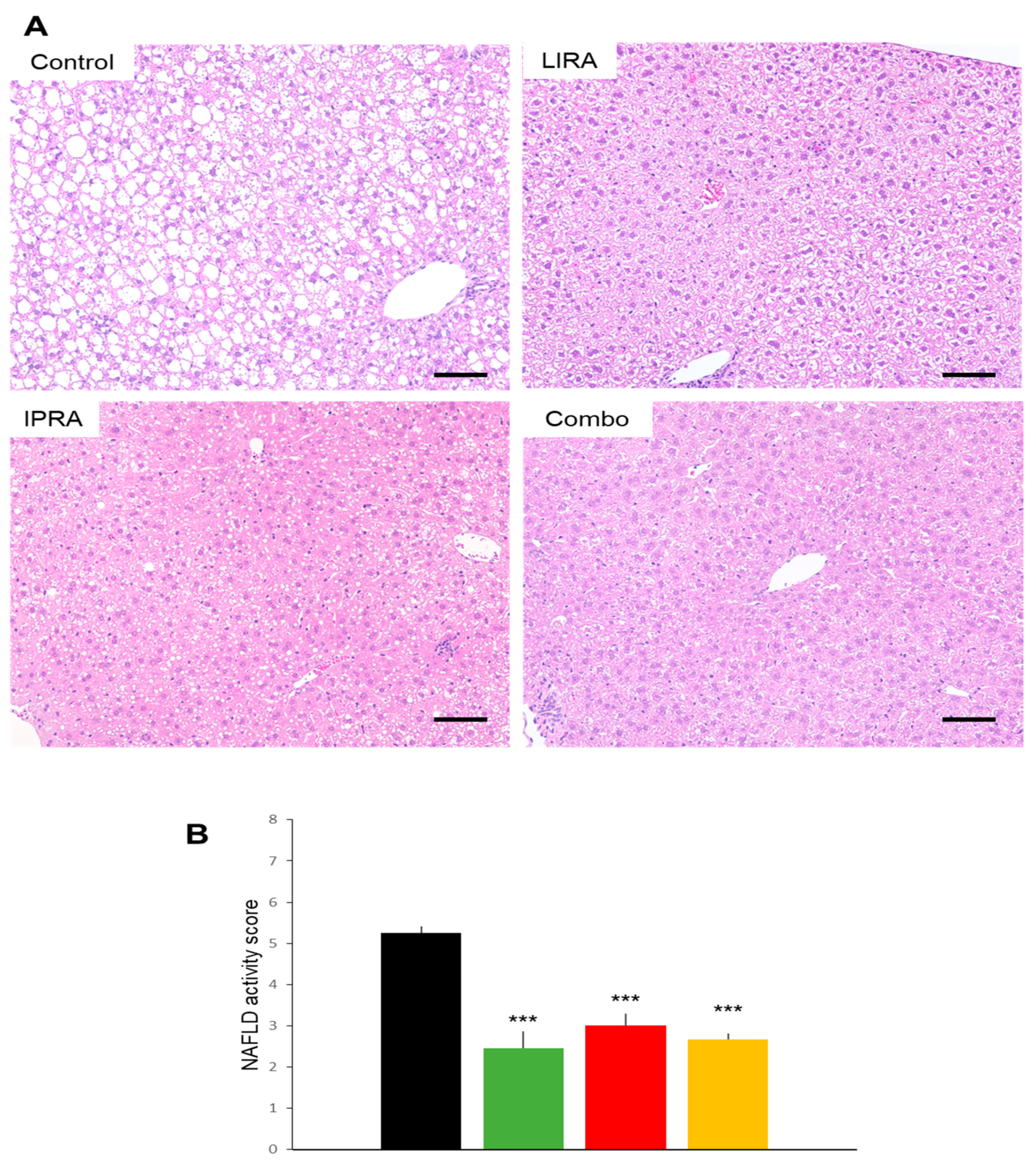
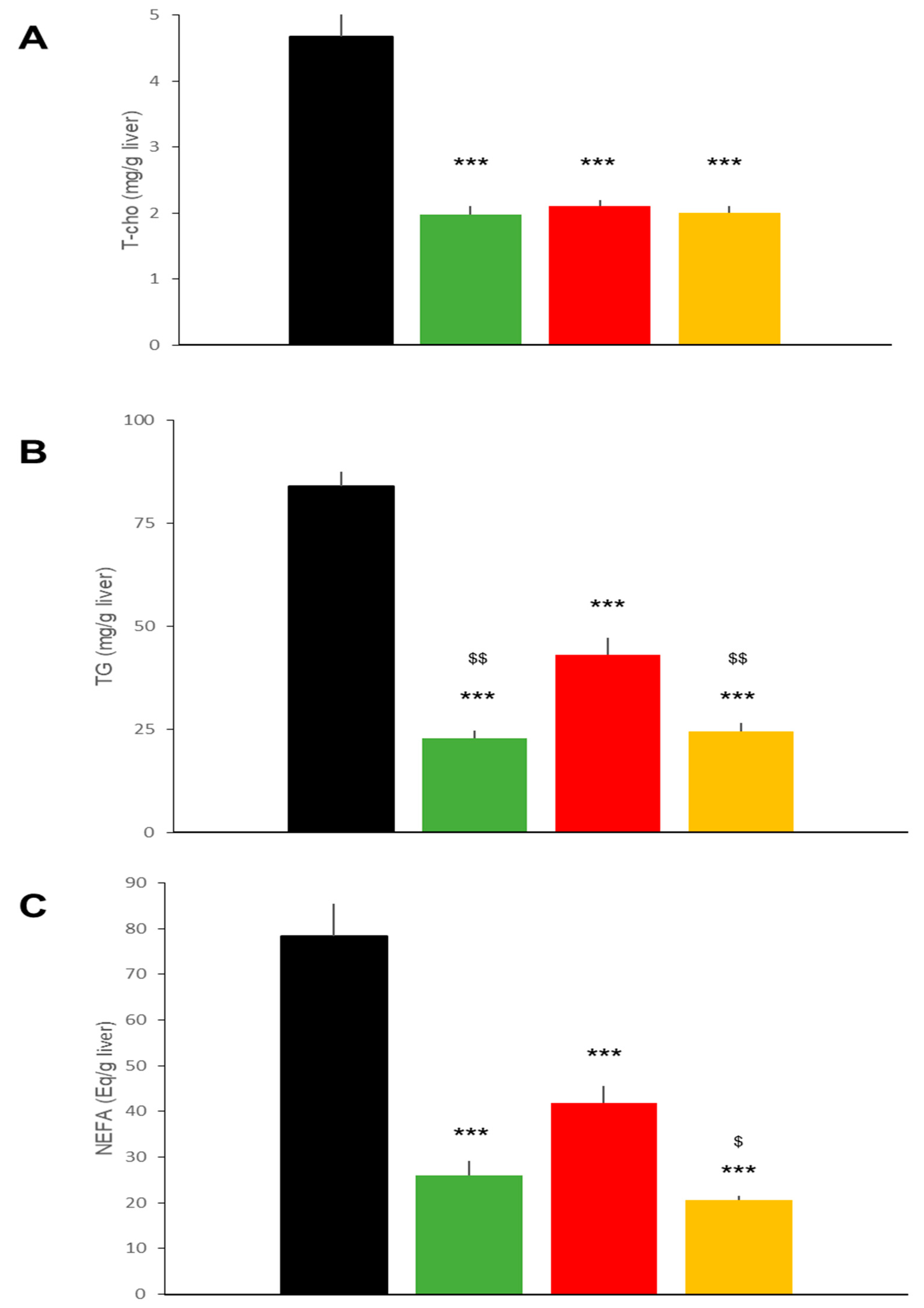
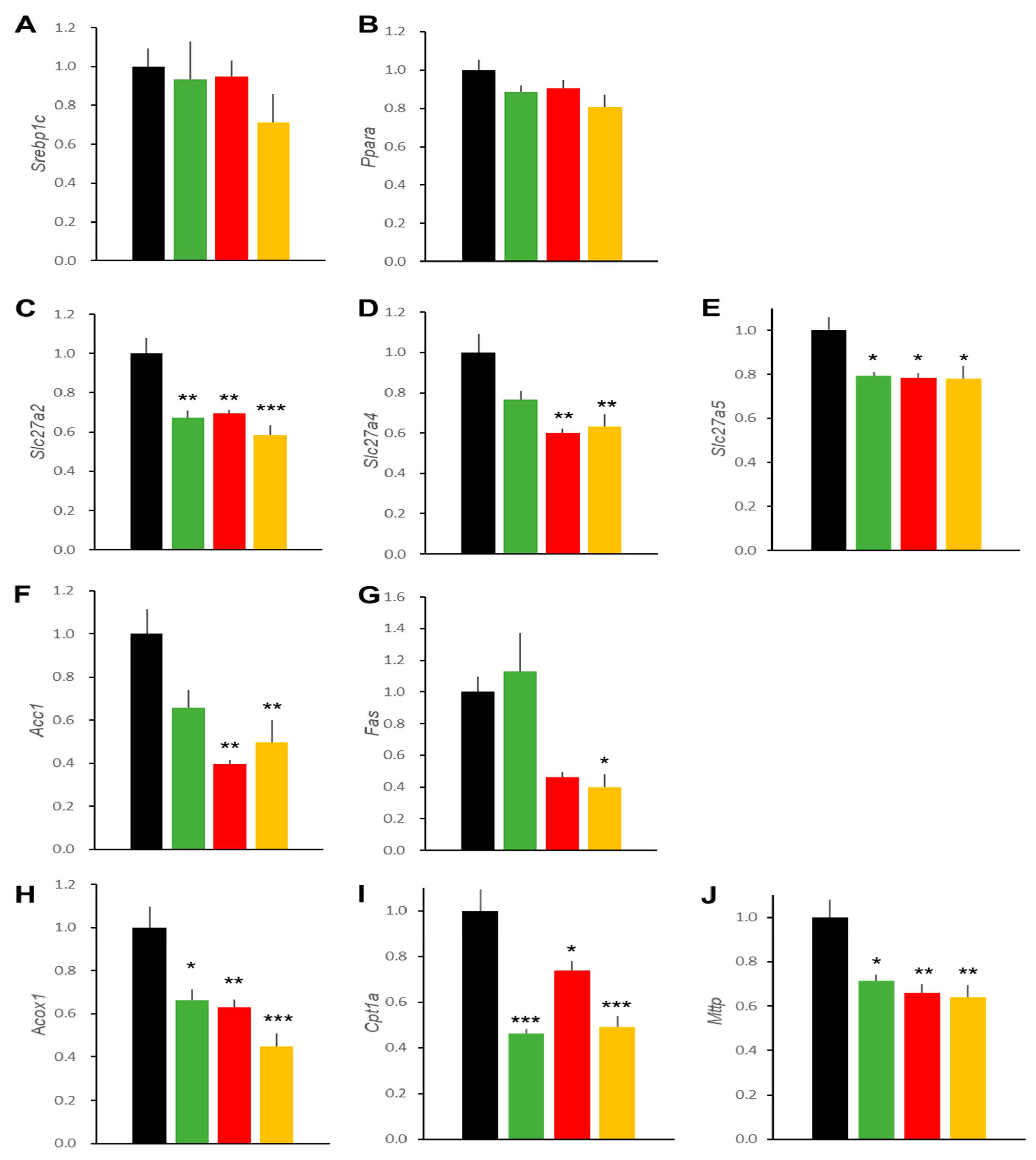
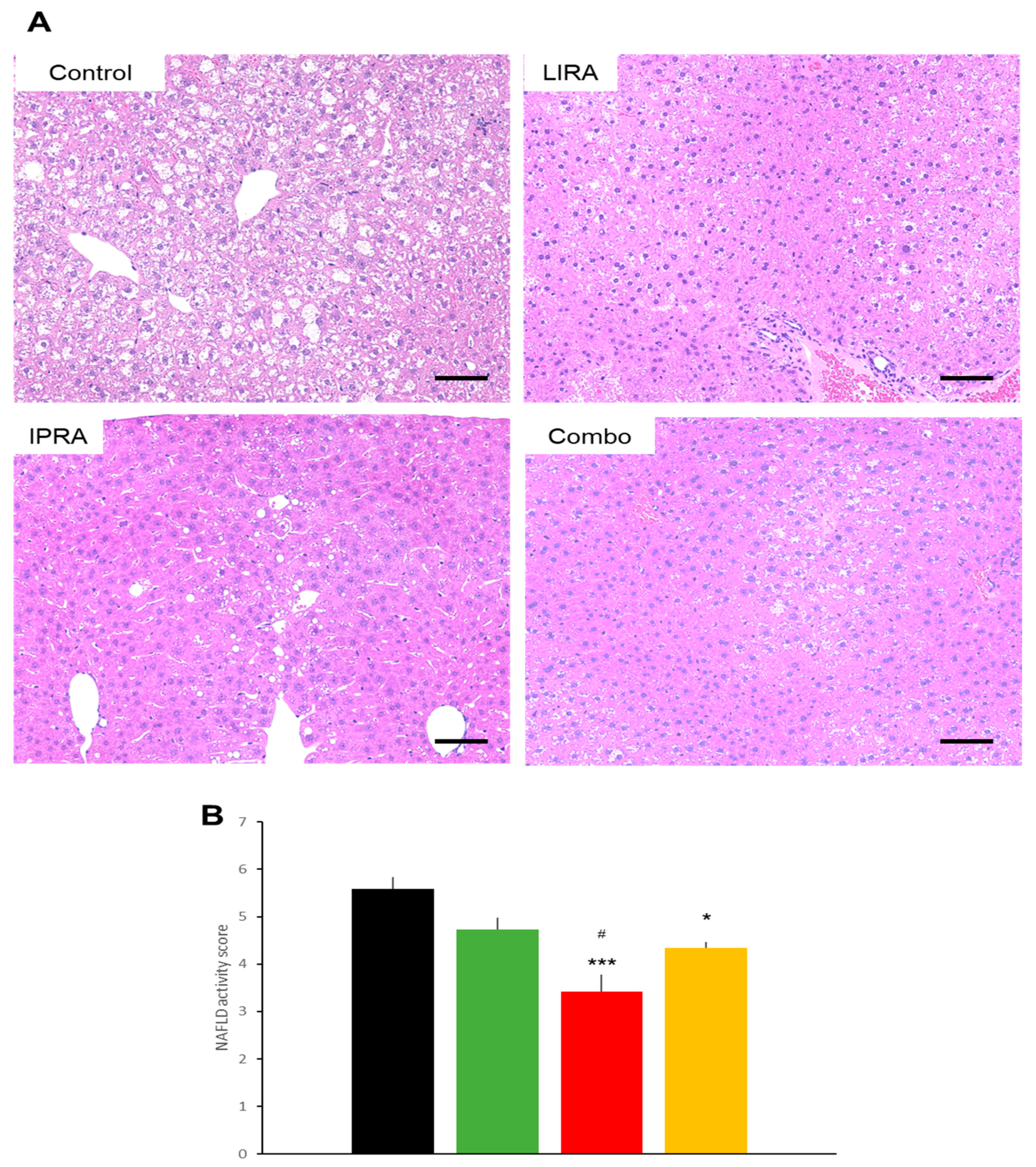
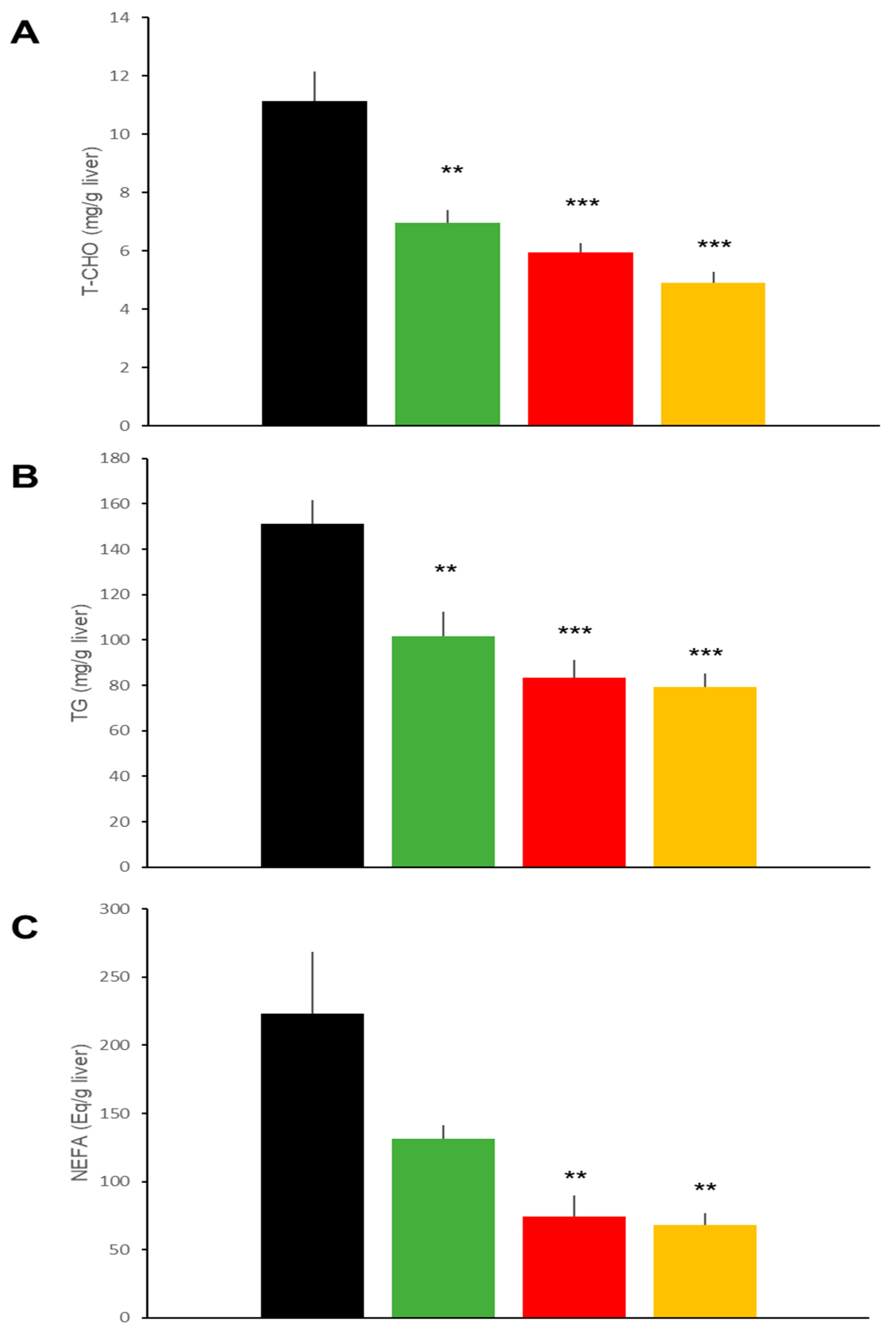
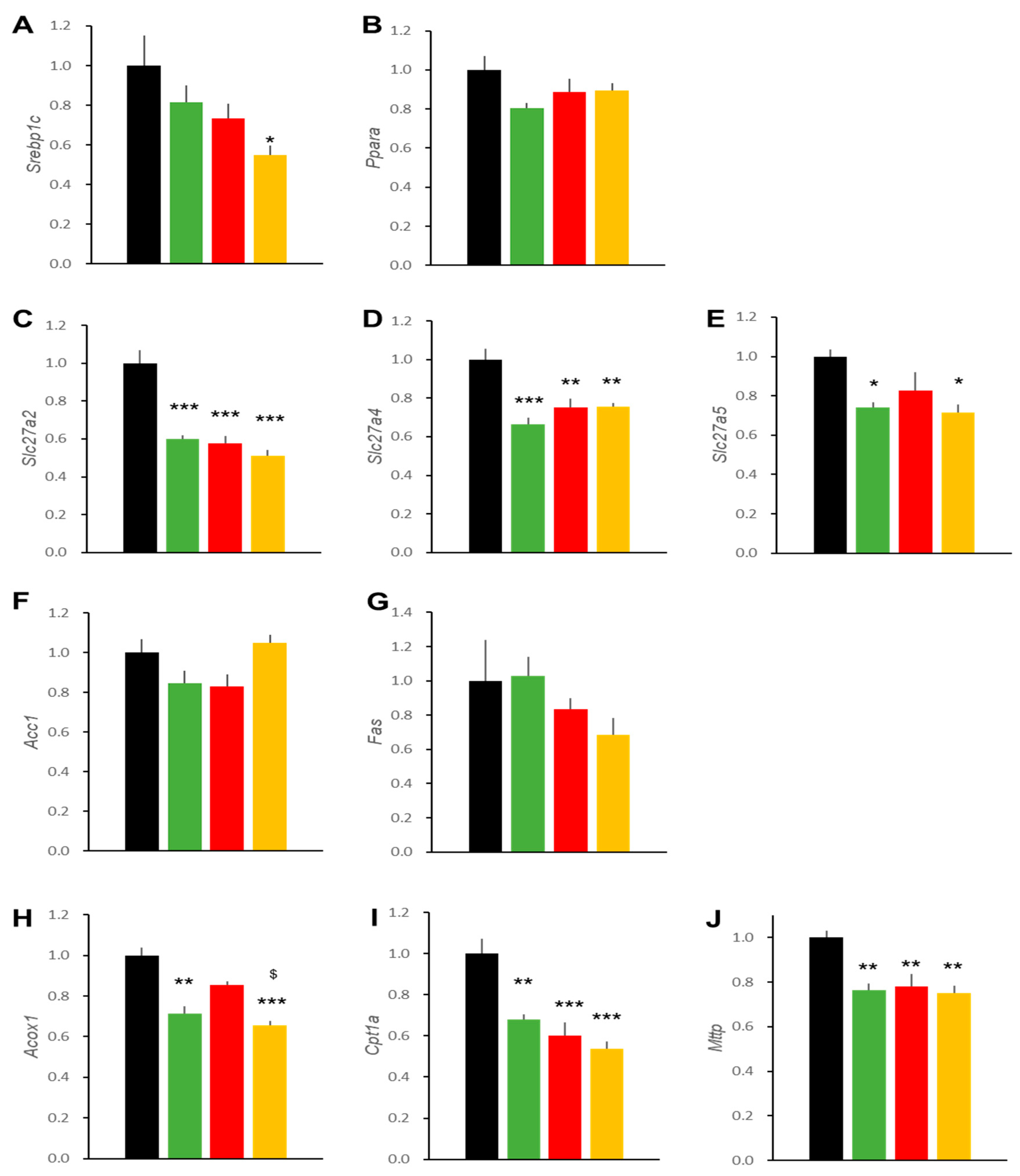
2.3. Hepatic Effects in DIO Mice
2.4. Effects on BW and Glycemia in db/db Mice
2.5. Pancreatic Effects in db/db Mice
2.6. Hepatic Effects in db/db Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histochemical Studies
4.3. Measurements of Serum Parameters, Hepatic Lipids, and Pancreatic Insulin Content
4.4. Quantitative Real Time Reverse Transcription PCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahren, B. Glucagon-like peptide-1 receptor agonists for type 2 diabetes: A rational drug development. J. Diabetes Investig. 2019, 10, 196–201. [Google Scholar] [CrossRef]
- Ferrannini, G.; Savarese, G.; Ryden, L. Sodium-glucose transporter inhibition in heart failure: From an unexpected side effect to a novel treatment possibility. Diabetes Res. Clin. Pract. 2021, 175, 108796. [Google Scholar] [CrossRef] [PubMed]
- Vincent, R.K.; Williams, D.M.; Evans, M. A look to the future in non-alcoholic fatty liver disease: Are glucagon-like peptide-1 analogues or sodium-glucose co-transporter-2 inhibitors the answer? Diabetes Obes. Metab. 2020, 22, 2227–2240. [Google Scholar] [CrossRef] [PubMed]
- Mirabelli, M.; Chiefari, E.; Caroleo, P.; Arcidiacono, B.; Corigliano, D.M.; Giuliano, S.; Brunetti, F.S.; Tanyolac, S.; Foti, D.P.; Puccio, L.; et al. Long-term effectiveness of liraglutide for weight management and glycemic control in type 2 diabetes. Int. J. Environ. Res. Public Health 2020, 17, 207. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Pharmacologic Approaches to Glycemic Treatment: Standards of Medical Care in Diabetes 2021. Diabetes Care. 2021, 44 (Suppl. 1), S111–S124. [Google Scholar] [CrossRef]
- Mantsiou, C.; Karagiannis, T.; Kakotrichi, P.; Malandris, K.; Avgerinos, I.; Liakos, A.; Tsapas, A.; Bekiari, E. Glucagon-like peptide-1 receptor agonists and sodium-glucose co-transporter-2 inhibitors as combination therapy for type 2 diabetes: A systematic review and meta-analysis. Diabetes Obes. Metab. 2020, 22, 1857–1868. [Google Scholar] [CrossRef]
- Gastaldelli, A.; Repetto, E.; Guja, C.; Hardy, E.; Han, J.; Jabbour, S.A.; Ferrannini, E. Exenatide and dapagliflozin combination improves markers of liver steatosis and fibrosis in patients with type 2 diabetes. Diabetes Obes. Metab. 2020, 22, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Harreiter, J.; Just, I.; Leutner, M.; Bastian, M.; Brath, H.; Schelkshorn, C.; Klepochova, R.; Krššák, M.; Kautzky-Willer, A. Combined exenatide and dapagliflozin has no additive effects on reduction of hepatocellular lipids despite better glycaemic control in patients with type 2 diabetes mellitus treated with metformin: EXENDA, a 24-week, prospective, randomized, placebo-controlled pilot trial. Diabetes Obes. Metab. 2021, 23, 1129–1139. [Google Scholar]
- Johnson, J.D.; Kushner, J.A. Endogenous Insulin: Its Role in the Initiation, Progression and Management of Diabetes. In The Endocrinologist; Society of Endocrinology: Bristol, UK, 2018; Volume 129, pp. 13–14. [Google Scholar]
- Boeder, S.; Edelman, S.V. Sodium-glucose co-transporter inhibitors as adjunctive treatment to insulin in type 1 diabetes: A review of randomized controlled trials. Diabetes Obes. Metab. 2019, 21 (Suppl. 2), 62–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kaneto, H.; Shimoda, M.; Hirukawa, H.; Okauchi, S.; Kohara, K.; Hamamoto, S.; Tawaramoto, K.; Hashiramoto, M.; Kaku, K. Protective effects of pioglitazone and/or liraglutide on pancreatic β-cells in db/db mice: Comparison of their effects between in an early and advanced stage of diabetes. Mol. Cell. Endocrinol. 2015, 400, 78–89. [Google Scholar] [CrossRef]
- Millar, P.; Pathak, N.; Parthsarathy, V.; Bjourson, A.J.; O’Kane, M.; Pathak, V.; Moffett, R.C.; Flatt, P.R.; Gault, V.A. Metabolic and neuroprotective effects of dapagliflozin and liraglutide in diabetic mice. J. Endocrinol. 2017, 234, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Imajo, K.; Kato, T.; Kessoku, T.; Ogawa, Y.; Tomeno, W.; Kato, S.; Mawatari, H.; Fujita, K.; Yoneda, M.; et al. The selective SGLT2 inhibitor ipragliflozin has a therapeutic effect on nonalcoholic steatohepatitis in mice. PLoS ONE 2016, 11, e0146337. [Google Scholar] [CrossRef] [Green Version]
- Kuhre, R.E.; Deacon, C.F.; Wewer Albrechtsen, N.J.; Holst, J.J. Do sodium-glucose co-transporter-2 inhibitors increase plasma glucagon by direct actions on the alpha cell? And does the increase matter for the associated incease in endogenous glucose production? Diebet Obes. Metab. 2021, 23, 2009–2019. [Google Scholar] [CrossRef]
- Ferrannini, E.; Muscelli, E.; Frascerra, S.; Baldi, S.; Mari, A.; Heise, T.; Broedl, U.C.; Woerle, H.-J. Metabolic response to sodium-glucose cotransporter 2 inhibition in type 2 diabetes patients. J. Clin. Investig. 2014, 124, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Mercovi, A.; Solius-Herrera, C.; Daniele, G.; Eldor, R.; Fiorentino, T.V.; Tripathy, D.; Xiong, J.; Perez, Z.; Norton, L.; Abdul-Gani, M.A.; et al. Dapagliflozin improves muscle insulin sensitivity bit enhances glucose production. J. Clin Investig. 2014, 124, 509–514. [Google Scholar]
- Ishihara, H.; Yamaguchi, S.; Nakao, I.; Sakatani, T. Ipragliflozin add-on therapy to a GLP-1 receptor agonist in Japanese patients with type 2 diabetes (AGATE): A 52-week open-label study. Diabetes Ther. 2018, 9, 1549–1567. [Google Scholar] [CrossRef] [Green Version]
- Kramer, C.K.; Zinman, B.; Choi, H.; Connelly, P.W.; Retnakaran, R. Impact of the glucagon assay when assessing the effects of chronic liraglutide therapy on glucagon secretion. J. Clin. Endocrinol. Metab. 2017, 102, 2729–2733. [Google Scholar] [CrossRef]
- Chae, H.; Augustin, R.; Gatineau, E.; Mayoux, E.; Bensellam, M.; Antonie, N.; Khattab, F.; Lai, B.K.; Brusa, D.; Stierstorfer, B.; et al. SGLT2 is not expressed in pancreatic alpha- and beta-cells, and its inhibition does not directly affect glucagon and insulin secretion in rodents and humans. Mol. Metab. 2020, 42, 101071. [Google Scholar] [CrossRef]
- Jojima, T.; Tomotsune, T.; Iijima, T.; Akimoto, K.; Suzuki, K.; Aso, Y. Empagliflozin (an SGLT2 inhibitor), alone or in combination with linagliptin (a DPP-4 inhibitor), prevents steatohepatitis in a novel mouse model of non-alcoholic steatohepatitis and diabetes. Diabetol. Metab. Syndr. 2016, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Clemmensen, C.; Finan, B.; Fischer, K.; Tom, R.Z.; Legutko, B.; Sehrer, L.; Heine, D.; Grassl, N.; Meyer, C.W.; Henderson, B.; et al. Dual melanocortin-4 receptor and GLP-1 receptor agonism amplifies metabolic benefits in diet-induced obese mice. EMBO Mol. Med. 2015, 7, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Madsen, M.S.A.; Holm, J.B.; Pallejà, A.; Wismann, P.; Fabricius, K.; Rigbolt, K.; Mikkelsen, M.; Sommer, M.; Jelsing, J.; Nielsen, H.B.; et al. Metabolic and gut microbiome changes following GLP-1 or dual GLP-1/GLP-2 receptor agonist treatment in diet-induced obese mice. Sci. Rep. 2019, 9, 15582. [Google Scholar] [CrossRef]
- Perakakis, N.; Stefanakis, K.; Feigh, M.; Veidal, S.S.; Mantzoros, C.S. Elafibranor and liraglutide improve differentially liver health and metabolism in a mouse model of non-alcoholic steatohepatitis. Liver Int. 2021, 41, 1853–1866. [Google Scholar] [CrossRef]
- Ye, X.; Qi, J.; Ren, G.; Xu, P.; Wu, Y.; Zhu, S.; Yu, D.; Li, S.; Wu, Q.; Muhi, R.L.; et al. Long-lasting anti-diabetic efficacy of PEGylated FGF-21 and liraglutide in treatment of type 2 diabetic mice. Endocrine 2015, 49, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Xu, R.X.; Cui, C.J.; Wang, Y.; Du, Y.; Chen, Z.G.; Yao, Y.H.; Ma, C.Y.; Zhu, C.G.; Guo, Y.L.; et al. Liraglutide downregulates hepatic LDL receptor and PCSK9 expression in HepG2 cells and db/db mice through a HNF-1a dependent mechanism. Cardiovasc. Diabetol. 2018, 17, 48. [Google Scholar] [CrossRef] [PubMed]
- Sumida, Y.; Yoneda, M.; Tokushige, K.; Kawanaka, M.; Fujii, H.; Yoneda, M.; Imajo, K.; Takahashi, H.; Eguchi, Y.; Ono, M. Antidiabetic therapy in the treatment of nonalcoholic steatohepatitis. Int. J. Mol. Sci. 2020, 21, 1907. [Google Scholar] [CrossRef] [Green Version]
- Kothari, S.; Dhami-Shah, H.; Shah, S.R. Antidiabetic drugs and statins in nonalcoholic fatty liver disease. J. Clin. Exp. Hepatol. 2019, 9, 723–730. [Google Scholar] [CrossRef] [Green Version]
- McLean, M.A.; Wong, C.K.; Campbell, J.E.; Hodson, D.J.; Trapp, S.; Drucker, D.J. Revisiting the complexity of GLP-1 action from sites of synthesis to receptor activation. Endocr. Rev. 2021, 42, 101–132. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.T.; Cheon, Y.; Li, Y.; Nara, T.Y. Mechanisms of regulation of gene expression by fatty acids. Lipids 2004, 39, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Decara, J.; Arrabal, S.; Beiroa, D.; Rivera, P.; Vargas, A.; Serrano, A.; Pavón, F.J.; Ballesteros, J.; Dieguez, C.; Nogueiras, R.; et al. Antiobesity efficacy of GLP-1 receptor agonist liraglutide is associated with peripheral tissue-specific modulation of lipid metabolic regulators. Biofactors 2016, 42, 600–611. [Google Scholar] [CrossRef]
- Mells, J.E.; Fu, P.P.; Sharma, S.; Olson, D.; Cheng, L.; Handy, J.A.; Saxena, N.K.; Sorescu, D.; Anania, F.A. Glp-1 analog, liraglutide, ameliorates hepatic steatosis and cardiac hypertrophy in C57BL/6J mice fed a Western diet. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G225–G235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | LIRA | IPRA | Combo |
|---|---|---|---|---|
| Insulin (ng/mL) | 0.40 ± 0.04 | 0.54 ± 0.14 | 0.46 ± 0.10 | 0.37 ± 0.11 |
| Glucagon (pmol/L) | 11.3 ± 0.8 | 11.0 ± 0.3 | 9.3 ± 1.1 | 11.0 ± 0.8 |
| AST (IU/L) | 57.0 ± 5.3 | 46.8 ± 3.2 | 60.0 ± 8.9 | 56.0 ± 4.4 |
| ALT (IU/L) | 36.7 ± 6.6 | 13.2 ± 0.50 | 22.2 ± 1.7 | 18.3 ± 2.5 |
| ** | * | |||
| T-cho (mg/dL) | 164.7 ± 3.9 | 130.3 ± 2.7 | 152.5 ± 0.9 | 120.0 ± 2.4 |
| *** $$$ | * | *** $$$ | ||
| TG (mg/dL) | 71.0 ± 5.5 | 68.3 ± 8.5 | 62.5 ± 6.6 | 58.7 ± 2.9 |
| Parameter | Control | LIRA | IPRA | Combo |
|---|---|---|---|---|
| Insulin (ng/mL) | 2.98 ± 0.16 | 1.90 ± 0.32 | 4.64 ± 0.60 | 5.34 ± 0.71 |
| # | *,## | |||
| Glucagon (pmol/L) | 12.5 ± 2.4 | 13.9 ± 3.2 | 17.2 ± 3.1 | 24.9 ± 2.4 |
| p = 0.0600 vs. control | ||||
| AST (IU/L) | 66.2 ± 3.9 | 58.7 ± 1.84 | 70.5 ± 4.48 | 51.8 ± 2.53 |
| $$ | ||||
| ALT (IU/L) | 73.5 ± 5.82 | 44.0 ± 2.2 | 65.7 ± 6.1 | 40.0 ± 2.9 |
| **,$$ | ***,$$ | |||
| T-cho (mg/dL) | 127.5 ± 6.76 | 109.8 ± 5.13 | 131.3 ± 3.6 | 113.0 ± 1.5 |
| $ | ||||
| TG (mg/dL) | 244.2 ± 20.0 | 147.2 ± 17.3 | 187.2 ± 28.8 | 107.8 ± 12.1 |
| * | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koike, M.; Saito, H.; Kohno, G.; Takubo, M.; Watanabe, K.; Ishihara, H. Effects of GLP-1RA and SGLT2i, Alone or in Combination, on Mouse Models of Type 2 Diabetes Representing Different Disease Stages. Int. J. Mol. Sci. 2021, 22, 11463. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111463
Koike M, Saito H, Kohno G, Takubo M, Watanabe K, Ishihara H. Effects of GLP-1RA and SGLT2i, Alone or in Combination, on Mouse Models of Type 2 Diabetes Representing Different Disease Stages. International Journal of Molecular Sciences. 2021; 22(21):11463. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111463
Chicago/Turabian StyleKoike, Masao, Hitoki Saito, Genta Kohno, Masahiro Takubo, Kentaro Watanabe, and Hisamitsu Ishihara. 2021. "Effects of GLP-1RA and SGLT2i, Alone or in Combination, on Mouse Models of Type 2 Diabetes Representing Different Disease Stages" International Journal of Molecular Sciences 22, no. 21: 11463. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111463