
7,8-Dihydroxiflavone Maintains Retinal Functionality and Protects Various Types of RGCs in Adult Rats with Optic Nerve Transection

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
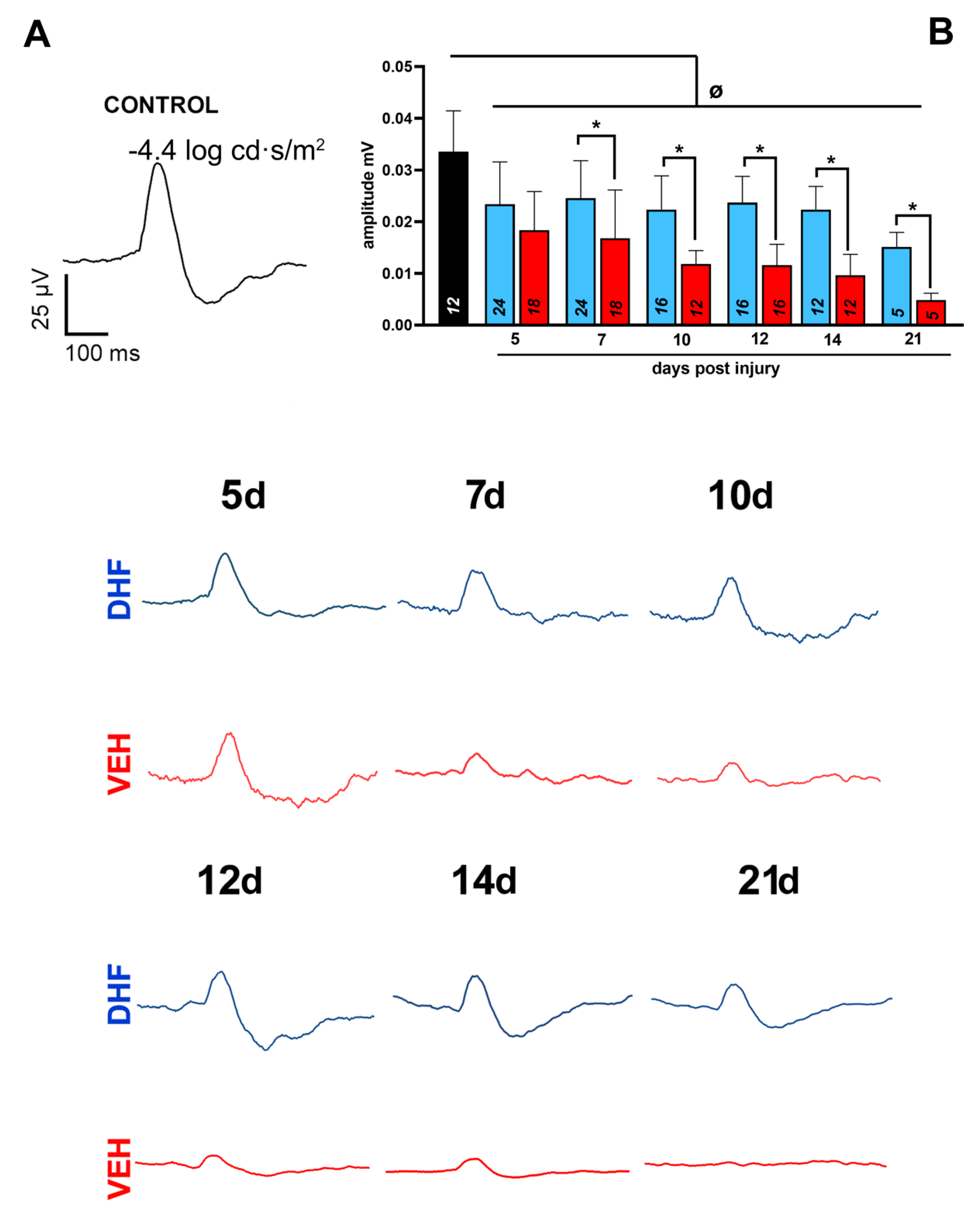
2.1. Alterations of the Main ERG Waves
2.1.1. Scotopic Threshold Responses
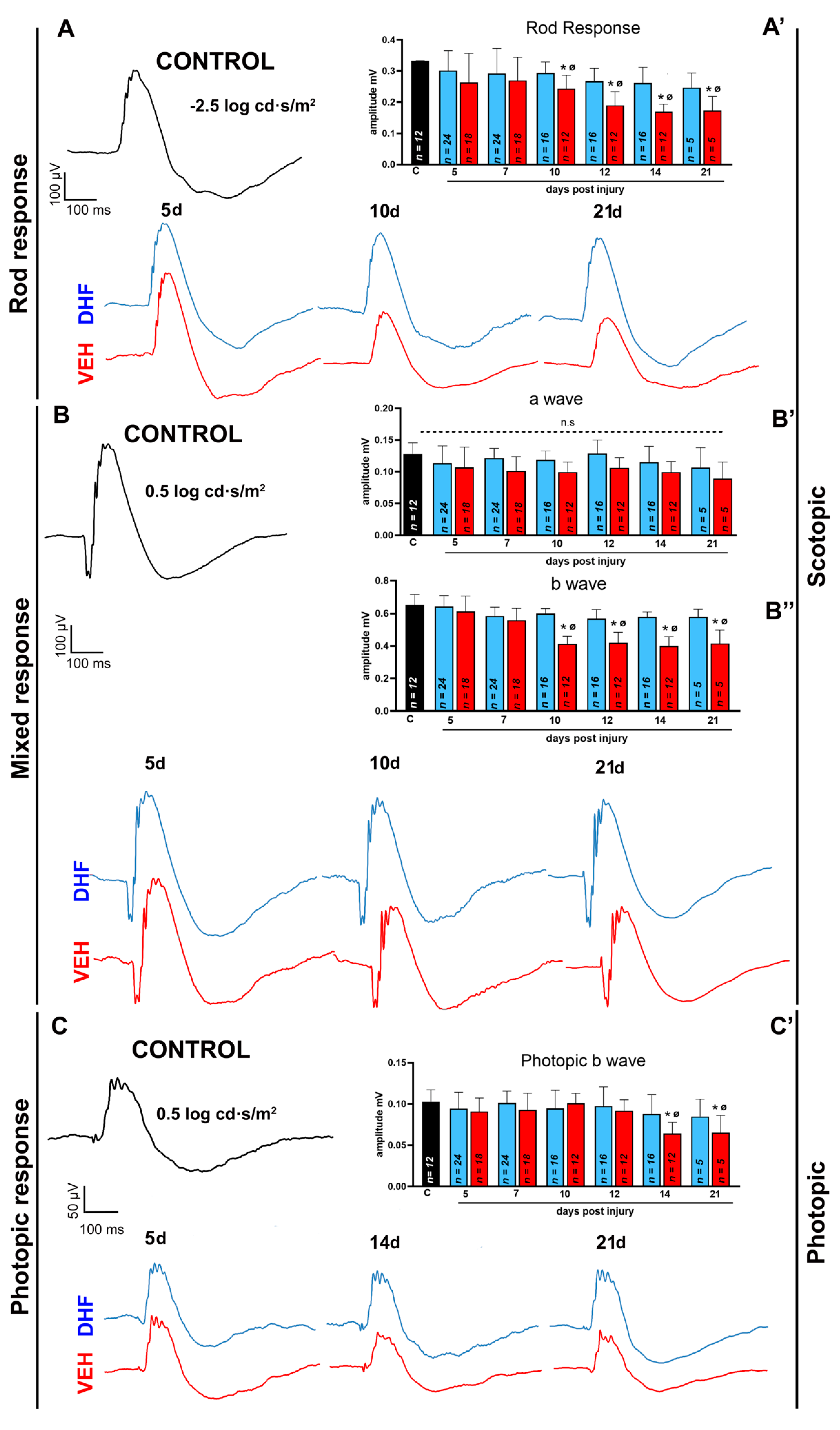
2.1.2. Rod Response
2.1.3. Mixed Response
2.1.4. Photopic b-wave
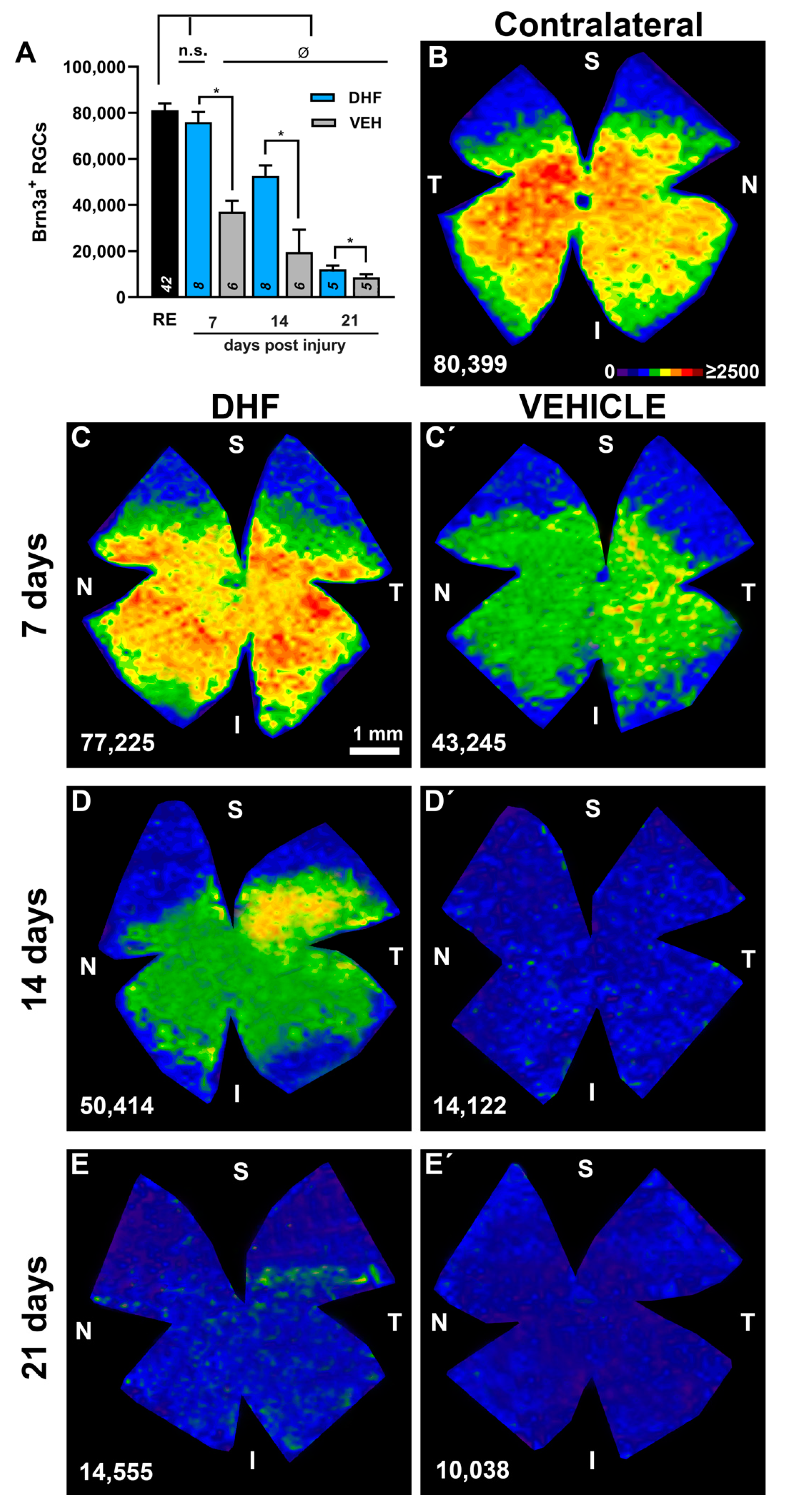
2.2. RGC Survival at Different Time-Intervals
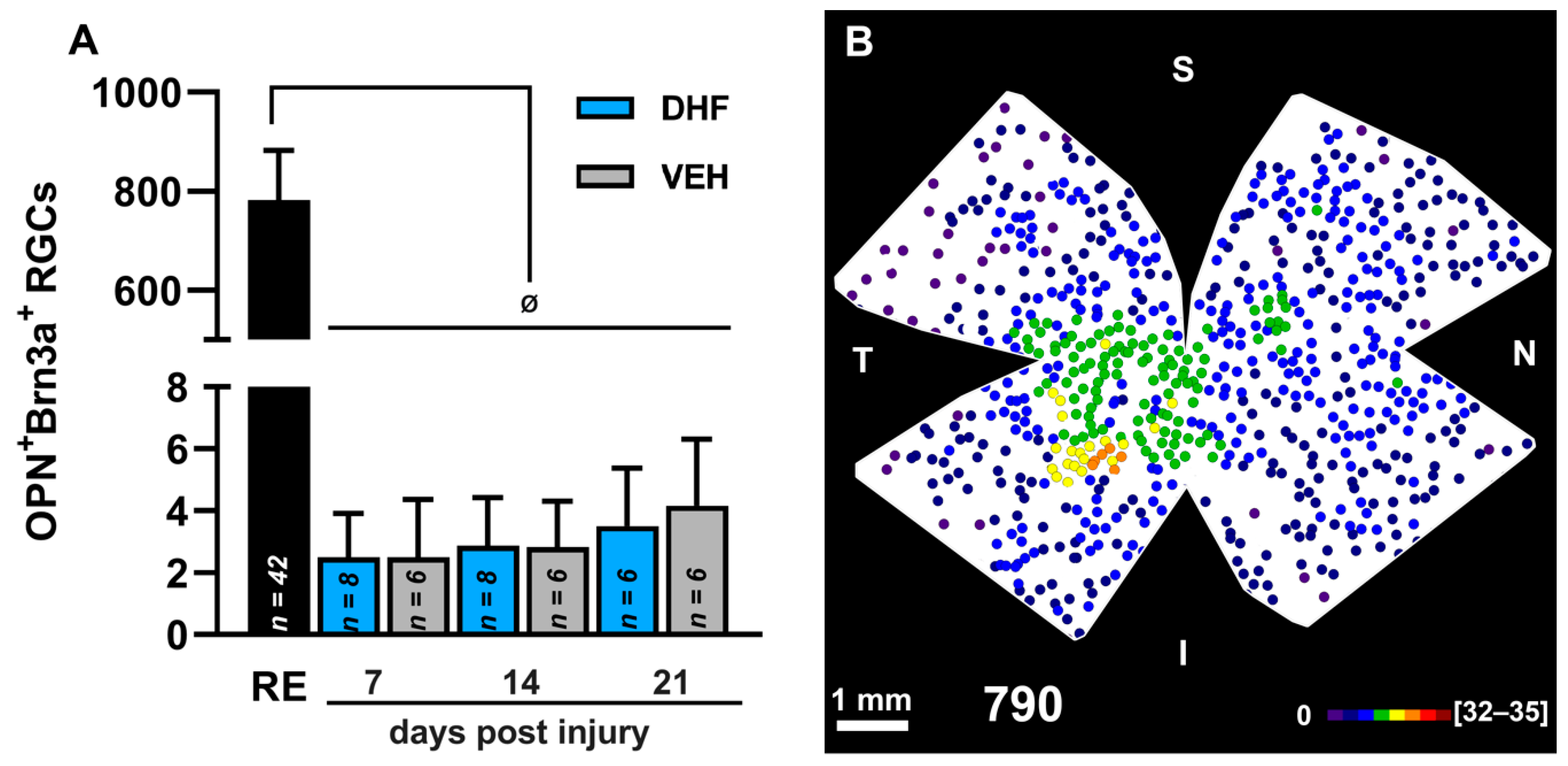
2.2.1. Brn3a+ RGCs
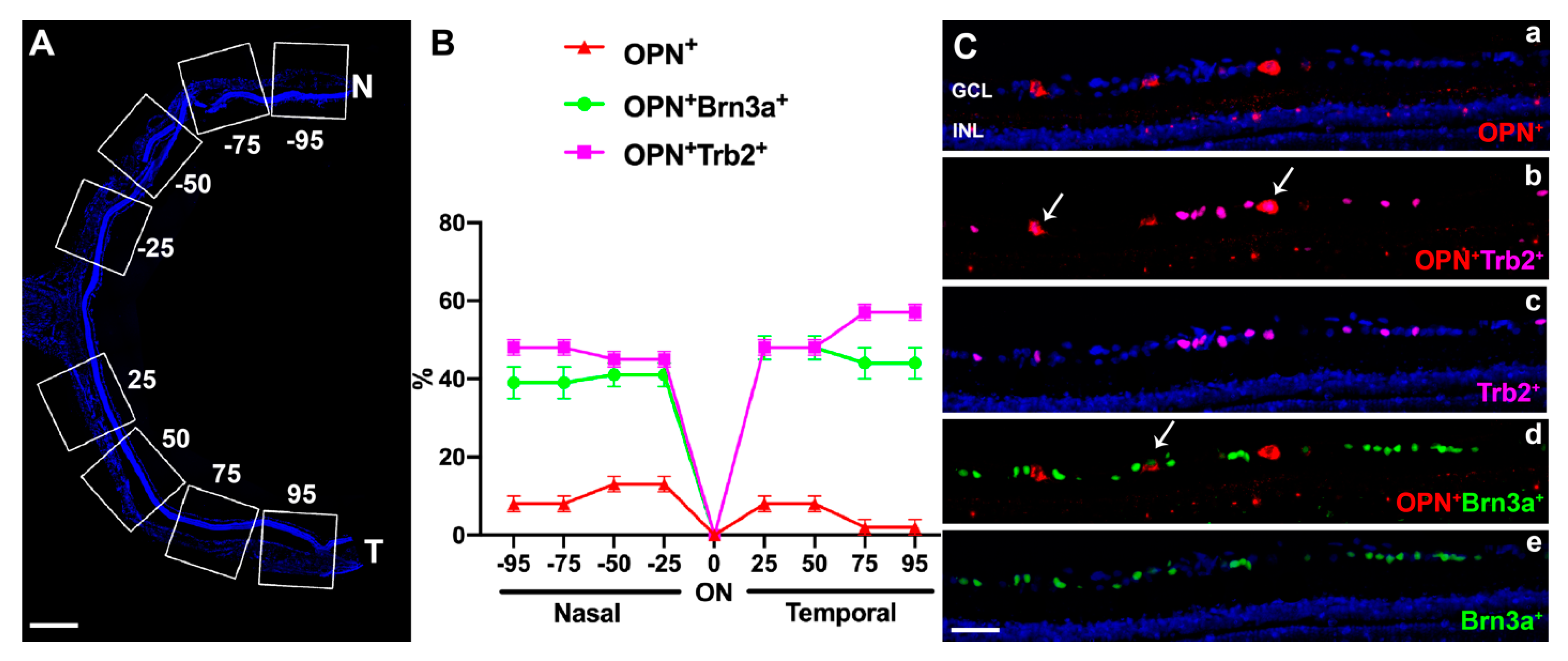
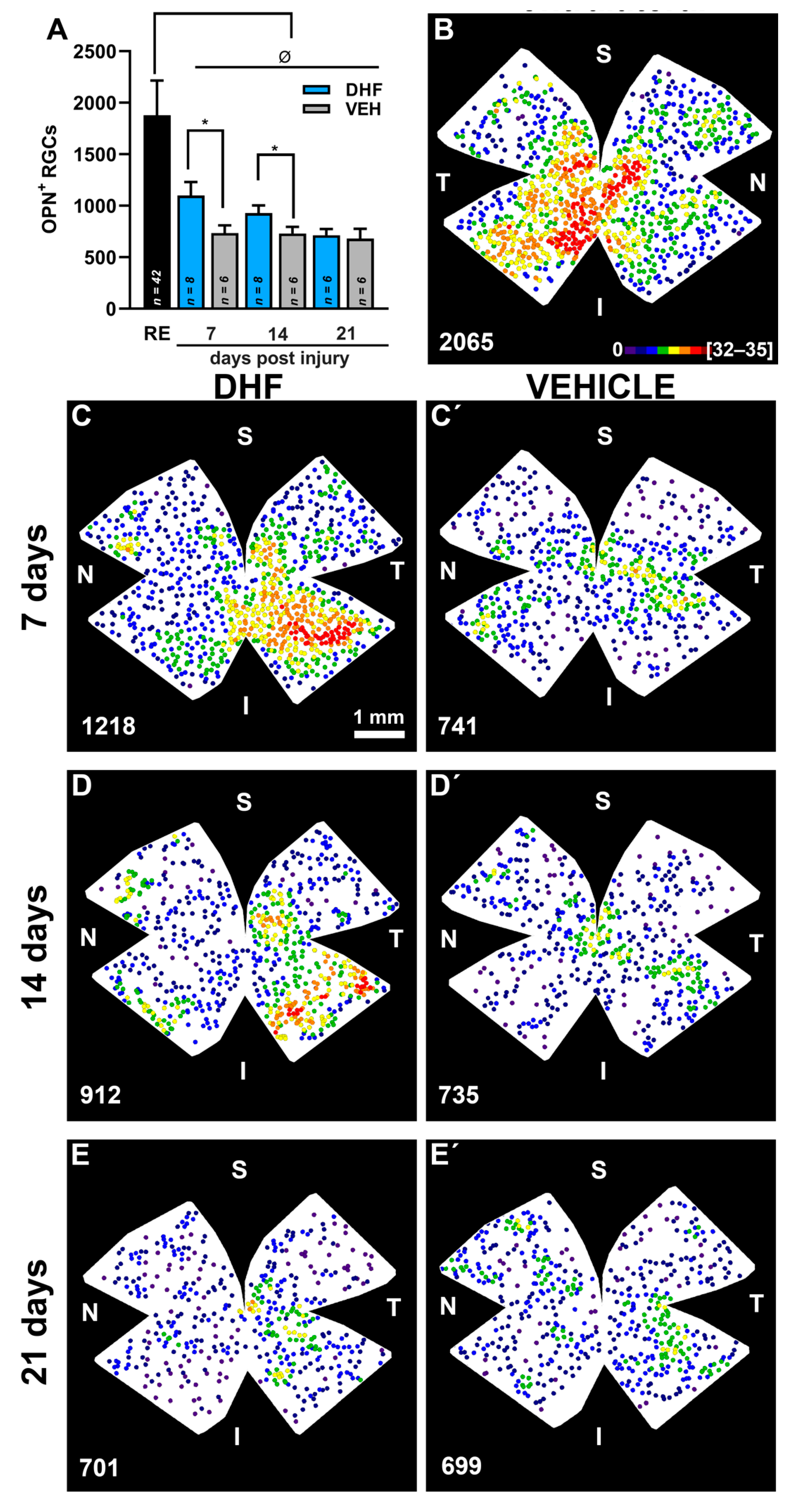
2.2.2. OPN+RGCs
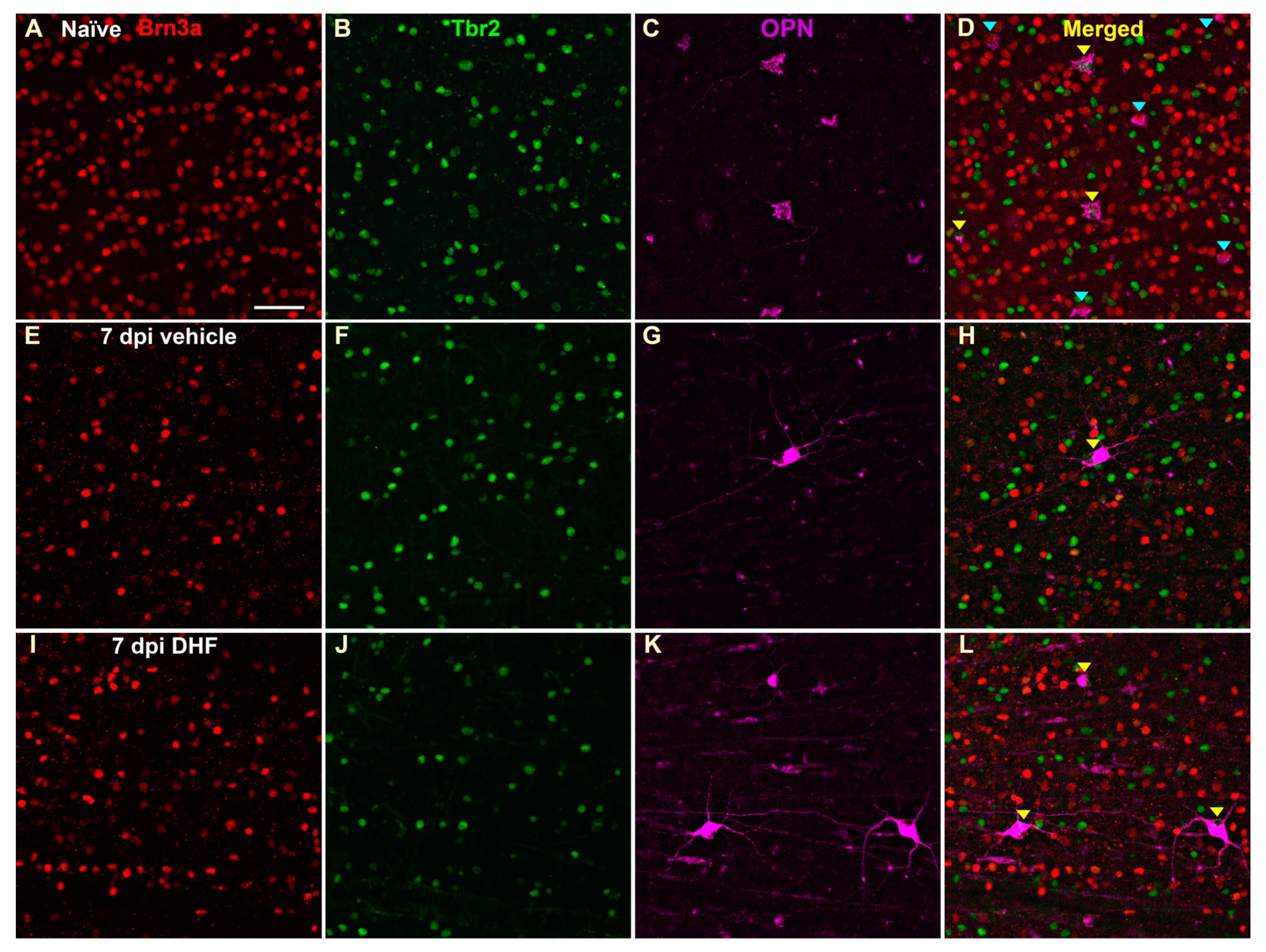
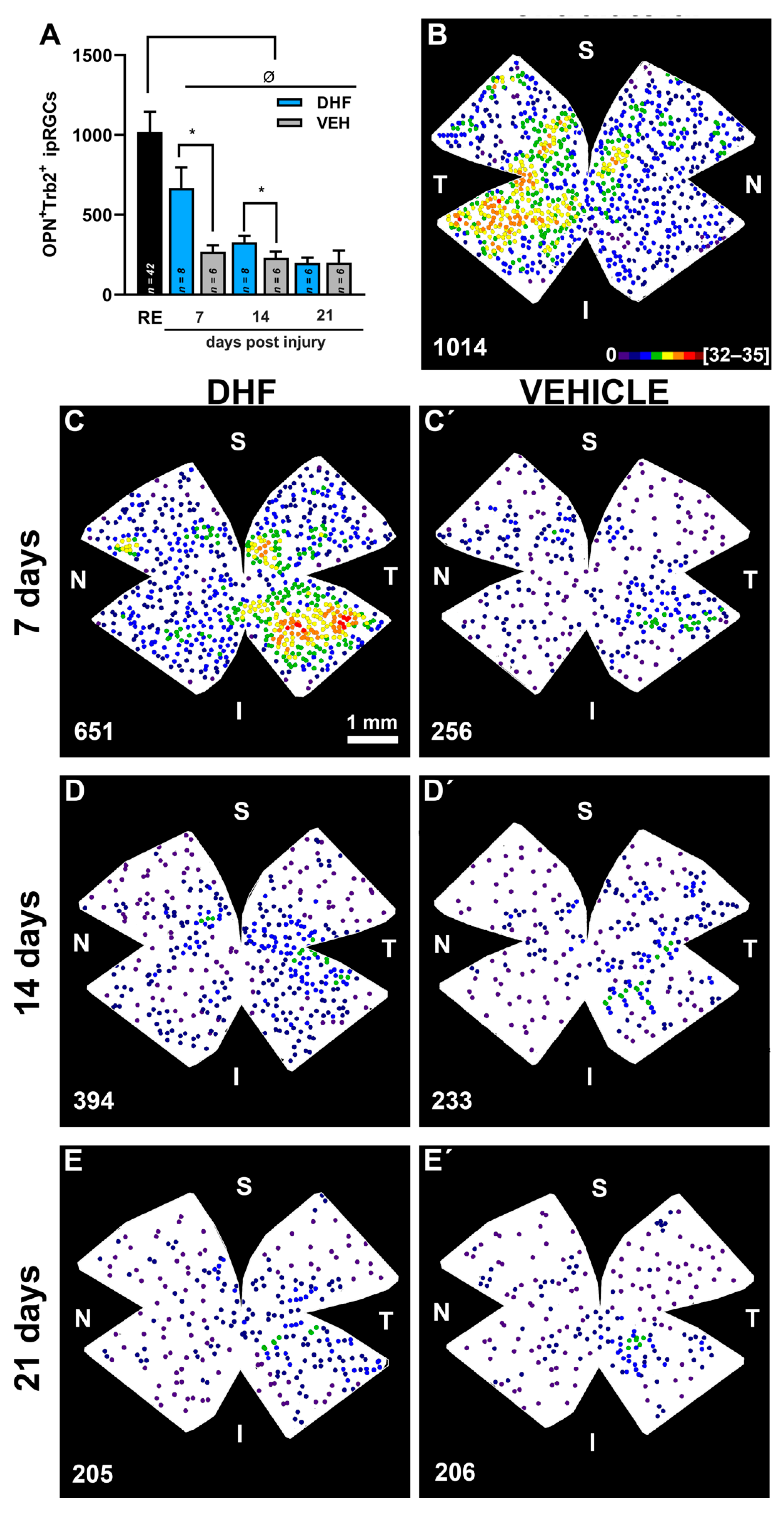
2.2.3. OPN+Tbr2+RGCs
3. Discussion
3.1. DHF Neuroprotection Involves Function of Ganglion Cells and Second Order Neurons
3.2. Albino Rat Retinal Distribution and Total Numbers of Brn3a, α-like, and M4-like RGCs
3.3. Resilience of α-RGCs and M4-like ipRGCs to IONT
3.4. DHF Prevents IONT-Induced RGC Loss
3.5. Possible Mechanisms of Action of DHF
3.6. Translational Relevance
3.7. Present Limitations
4. Materials and Methods
4.1. Animals
4.2. Axotomy of the Left Optic Nerve
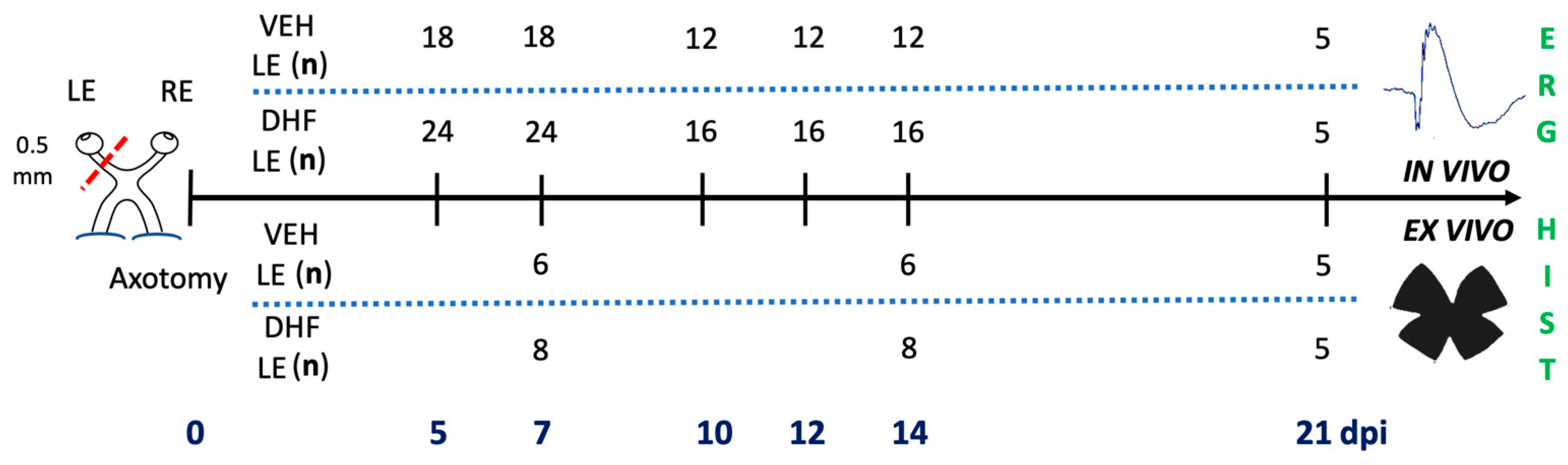
4.3. Experimental Design and Drug Administration
4.4. Electroretinography (ERG)
4.5. Animal Processing and Immunocytochemistry to Identify Different RGC Types
4.6. Assessment of Retinal Ganglion Cell Types
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguayo, A.J.; Vidal-Sanz, M.; Villegas-Pérez, M.P.; Bray, G.M. Growth and connectivity of axotomized retinal neurons in adult rats with optic nerves substituted by PNS grafts linking the eye and the midbrain. Ann. N. Y. Acad. Sci. 1987, 495, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McKerracher, L.; Vidal-Sanz, M.; Essagian, C.; Aguayo, A.J. Selective impairment of slow axonal transport after optic nerve injury in adult rats. J. Neurosci. 1990, 10, 2834–2841. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, S.J.O.; Sauvé, Y.; Avilés-Trigueros, M.; Vidal-Sanz, M.; Lund, R.D. Extent and Duration of Recovered Pupillary Light Reflex Following Retinal Ganglion Cell Axon Regeneration through Peripheral Nerve Grafts Directed to the Pretectum in Adult Rats. Exp. Neurol. 1998, 154, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Sanz, M.; Avilés-Trigueros, M.; Whiteley, S.J.O.; Sauvé, Y.; Lund, R.D. Reinnervation of the pretectum in adult rats by regenerated retinal ganglion cell axons: Anatomical and functional studies. Prog. Brain Res. 2002, 137, 443–452. [Google Scholar]
- Sasaki, H.; Coffey, P.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Young, M.J.; Lund, R.D.; Fukuda, Y. Light induced EEG desynchronization and behavioral arousal in rats with restored retinocollicular projection by peripheral nerve graft. Neurosci. Lett. 1996, 218, 45–48. [Google Scholar] [CrossRef]
- Avilés-Trigueros, M.; Sauvé, Y.; Lund, R.D.; Vidal-Sanz, M. Selective innervation of retinorecipient brainstem nuclei by retinal ganglion cell axons regenerating through peripheral nerve grafts in adult rats. J. Neurosci. 2000, 20, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Schlamp, C.L.; Johnson, E.C.; Li, Y.; Morrison, J.C.; Nickells, R.W. Changes in Thy1 gene expression associated with damaged retinal ganglion cells. Mol. Vis. 2001, 7, 192–201. [Google Scholar]
- Casson, R.J.; Chidlow, G.; Wood, J.P.M.; Vidal-Sanz, M.; Osborne, N.N. The effect of retinal ganglion cell injury on light-induced photoreceptor degeneration. Invest. Ophthalmol. Vis. Sci. 2004, 45, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Chidlow, G.; Casson, R.J.; Sobrado-Calvo, P.; Vidal-Sanz, M.; Osborne, N.N. Measurement of retinal injury in the rat after optic nerve transection: An RT-PCR study. Mol. Vis. 2005, 11, 387–396. [Google Scholar] [PubMed]
- Stutzki, H.; Leibig, C.; Andreadaki, A.; Fischer, D.; Zeck, G. Inflammatory stimulation preserves physiological properties of retinal ganglion cells after optic nerve injury. Front. Cell. Neurosci. 2014, 8, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas-Perez, M.P.; Vidal-Sanz, M.; Bray, G.M.; Aguayo, A.J. Influences of peripheral nerve grafts on the survival and regrowth of axotomized retinal ganglion cells in adult rats. J. Neurosci. 1988, 8, 265–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas-Pérez, M.-P.; Vidal-Sanz, M.; Rasminsky, M.; Bray, G.M.; Aguayo, A.J. Rapid and protracted phases of retinal ganglion cell loss follow axotomy in the optic nerve of adult rats. J. Neurobiol. 1993, 24, 23–36. [Google Scholar] [CrossRef]
- Rovere, G.; Nadal-Nicolás, F.M.; Agudo-Barriuso, M.; Sobrado-Calvo, P.; Nieto-López, L.; Nucci, C.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Comparison of Retinal Nerve Fiber Layer Thinning and Retinal Ganglion Cell Loss After Optic Nerve Transection in Adult Albino Rats. Investig. Opthalmology Vis. Sci. 2015, 56, 4487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal-Sanz, M.; Lafuente, M.P.; Mayor, S.; de Imperial, J.M.; Villegas-Pérez, M.P. Retinal ganglion cell death induced by retinal ischemia. neuroprotective effects of two alpha-2 agonists. Surv. Ophthalmol. 2001, 45 (Suppl. 3), S261–S267. [Google Scholar] [CrossRef]
- Lafuente, M.P.; Villegas-Pérez, M.P.; Sobrado-Calvo, P.; García-Avilés, A.; Miralles de Imperial, J.; Vidal-Sanz, M. Neuroprotective effects of alpha(2)-selective adrenergic agonists against ischemia-induced retinal ganglion cell death. Invest. Ophthalmol. Vis. Sci. 2001, 42, 2074–2084. [Google Scholar] [PubMed]
- Lafuente López-Herrera, M.P.; Mayor-Torroglosa, S.; Miralles de Imperial, J.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Transient ischemia of the retina results in altered retrograde axoplasmic transport: Neuroprotection with brimonidine. Exp. Neurol. 2002, 178, 243–258. [Google Scholar] [CrossRef]
- Lafuente, M.P.; Villegas-Pérez, M.P.; Mayor, S.; Aguilera, M.E.; Miralles de Imperial, J.; Vidal-Sanz, M. Neuroprotective effects of brimonidine against transient ischemia-induced retinal ganglion cell death: A dose response in vivo study. Exp. Eye Res. 2002, 74, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Trigueros, M.; Mayor-Torroglosa, S.; García-Avilés, A.; Lafuente, M.P.; Rodríguez, M.E.; De Imperial, J.M.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Transient ischemia of the retina results in massive degeneration of the retinotectal projection: Long-term neuroprotection with brimonidine. Exp. Neurol. 2003, 184, 767–777. [Google Scholar] [CrossRef]
- Mayor-Torroglosa, S.; De La Villa, P.; Rodríguez, M.E.; Lafuente López-Herrera, M.P.; Avilés-Trigueros, M.; García-Avilés, A.; Miralles De Imperial, J.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Ischemia results 3 months later in altered ERG, degeneration of inner layers, and deafferented tectum: Neuroprotection with brimonidine. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3825–3835. [Google Scholar] [CrossRef] [PubMed]
- Lönngren, U.; Näpänkangas, U.; Lafuente, M.; Mayor, S.; Lindqvist, N.; Vidal-Sanz, M.; Hallböök, F. The growth factor response in ischemic rat retina and superior colliculus after brimonidine pre-treatment. Brain Res. Bull. 2006, 71, 208–218. [Google Scholar] [CrossRef]
- Vidal-Sanz, M.; de la Villa, P.; Avilés-Trigueros, M.; Mayor-Torroglosa, S.; Salinas-Navarro, M.; Alarcón-Martínez, L.; Villegas-Pérez, M.P. Neuroprotection of retinal ganglion cell function and their central nervous system targets. Eye 2007, 21, S42–S45. [Google Scholar] [CrossRef]
- Parrilla-Reverter, G.; Agudo, M.; Nadal-Nicolás, F.; Alarcón-Martínez, L.; Jiménez-López, M.; Salinas-Navarro, M.; Sobrado-Calvo, P.; Bernal-Garro, J.M.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Time-course of the retinal nerve fibre layer degeneration after complete intra-orbital optic nerve transection or crush: A comparative study. Vis. Res. 2009, 49, 2808–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindqvist, N.; Peinado-Ramón, P.; Vidal-Sanz, M.; Hallböök, F. GDNF, Ret, GFRα1 and 2 in the adult rat retino-tectal system after optic nerve transection. Exp. Neurol. 2004, 187, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Di Polo, A.; Aigner, L.J.; Dunn, R.J.; Bray, G.M.; Aguayo, A.J. Prolonged delivery of brain-derived neurotrophic factor by adenovirus-infected Müller cells temporarily rescues injured retinal ganglion cells. Proc. Natl. Acad. Sci. USA 1998, 95, 3978–3983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Migallón, M.C.; Nadal-Nicolás, F.M.; Jiménez-López, M.; Sobrado-Calvo, P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Brain derived neurotrophic factor maintains Brn3a expression in axotomized rat retinal ganglion cells. Exp. Eye Res. 2011, 92, 260–267. [Google Scholar] [CrossRef]
- Sánchez-Migallón, M.C.; Valiente-Soriano, F.J.; Nadal-Nicolás, F.M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Apoptotic retinal ganglion cell death after optic nerve transection or crush in mice: Delayed RGC loss with BDNF or a caspase 3 inhibitor. Investig. Ophthalmol. Vis. Sci. 2016, 57, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Migallón, M.C.; Valiente-Soriano, F.J.; Salinas-Navarro, M.; Nadal-Nicolás, F.M.; Jiménez-López, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Nerve fibre layer degeneration and retinal ganglion cell loss long term after optic nerve crush or transection in adult mice. Exp. Eye Res. 2018, 170, 40–50. [Google Scholar] [CrossRef]
- Valiente-Soriano, F.J.; Nadal-Nicolás, F.M.; Salinas-Navarro, M.; Jiménez-López, M.; Bernal-Garro, J.M.; Villegas-Pérez, M.P.; Agudo-Barriuso, M.; Vidal-Sanz, M. BDNF Rescues RGCs But Not Intrinsically Photosensitive RGCs in Ocular Hypertensive Albino Rat Retinas. Investig. Opthalmol. Vis. Sci. 2015, 56, 1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovere, G.; Nadal-Nicolás, F.M.; Wang, J.; Bernal-Garro, J.M.; García-Carrillo, N.; Villegas-Pérez, M.P.; Agudo-Barriuso, M.; Vidal-Sanz, M. Melanopsin-containing or non-melanopsin-containing retinal ganglion cells response to acute ocular hypertension with or without brain-derived neurotrophic factor neuroprotection. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6652–6661. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Sanz, M.; Galindo-Romero, C.; Valiente-Soriano, F.J.; Nadal-Nicolás, F.M.; Ortin-Martinez, A.; Rovere, G.; Salinas-Navarro, M.; Lucas-Ruiz, F.; Sanchez-Migallon, M.C.; Sobrado-Calvo, P.; et al. Shared and Differential Retinal Responses against Optic Nerve Injury and Ocular Hypertension. Front. Neurosci. 2017, 11, 235. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Sanz, M.; Lafuente, M.; Sobrado-Calvo, P.; SellÉs-Navarro, I.; Rodriguez, E.; Mayor-Torroglosa, S.; Villegas-PÉrez, M.P. Death and neuroprotection of retinal ganglion cells after different types of injury. Neurotox. Res. 2000, 2, 215–227. [Google Scholar] [CrossRef]
- Lucas-Ruiz, F.; Galindo-Romero, C.; Salinas-Navarro, M.; González-Riquelme, M.J.; Vidal-Sanz, M.; Agudo Barriuso, M. Systemic and Intravitreal Antagonism of the TNFR1 Signaling Pathway Delays Axotomy-Induced Retinal Ganglion Cell Loss. Front. Neurosci. 2019, 13, 1096. [Google Scholar] [CrossRef] [Green Version]
- Romano, G.L.; Amato, R.; Lazzara, F.; Porciatti, V.; Chou, T.-H.; Drago, F.; Bucolo, C. P2X7 receptor antagonism preserves retinal ganglion cells in glaucomatous mice. Biochem. Pharmacol. 2020, 180, 114199. [Google Scholar] [CrossRef]
- Conti, F.; Romano, G.L.; Eandi, C.M.; Toro, M.D.; Rejdak, R.; Di Benedetto, G.; Lazzara, F.; Bernardini, R.; Drago, F.; Cantarella, G.; et al. Brimonidine is Neuroprotective in Animal Paradigm of Retinal Ganglion Cell Damage. Front. Pharmacol. 2021, 12, 705405. [Google Scholar] [CrossRef] [PubMed]
- Lazzara, F.; Amato, R.; Platania, C.B.M.; Conti, F.; Chou, T.-H.; Porciatti, V.; Drago, F.; Bucolo, C. 1α,25-dihydroxyvitamin D(3) protects retinal ganglion cells in glaucomatous mice. J. Neuroinflamm. 2021, 18, 206. [Google Scholar] [CrossRef] [PubMed]
- Emili, M.; Guidi, S.; Uguagliati, B.; Giacomini, A.; Bartesaghi, R.; Stagni, F. Treatment with the flavonoid 7,8-Dihydroxyflavone: A promising strategy for a constellation of body and brain disorders. Crit. Rev. Food Sci. Nutr. 2020. [Google Scholar] [CrossRef]
- Liu, X.; Qi, Q.; Xiao, G.; Li, J.; Luo, H.R.; Ye, K. O-methylated metabolite of 7,8-dihydroxyflavone activates TrkB receptor and displays antidepressant activity. Pharmacology 2013, 91, 185–200. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Villegas, B.; Di Pierdomenico, J.; Gallego-Ortega, A.; Galindo-Romero, C.; Martínez-de-la-Casa, J.M.; García-Feijoo, J.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Systemic treatment with 7,8-Dihydroxiflavone activates TtkB and affords protection of two different retinal ganglion cell populations against axotomy in adult rats. Exp. Eye Res. 2021, 210, 108694. [Google Scholar] [CrossRef]
- Galindo-Romero, C.; Vidal-Villegas, B.; Asís-Martínez, J.; Lucas-Ruiz, F.; Gallego-Ortega, A.; Vidal-Sanz, M. 7,8-Dihydroxiflavone protects adult rat axotomized retinal ganglion cells through MAPK/ERK and PI3K/AKT activation. Int. J. Mol. Sci. 2021, 22, 10896. [Google Scholar] [CrossRef] [PubMed]
- Segal, R.A.; Greenberg, M.E. Intracellular signaling pathways activated by neurotrophic factors. Annu. Rev. Neurosci. 1996, 19, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Huxlin, K.R.; Goodchild, A.K. Retinal ganglion cells in the albino rat: Revised morphological classification. J. Comp. Neurol. 1997, 385, 309–323. [Google Scholar] [CrossRef]
- Perry, V.H. The ganglion cell layer of the retina of the rat: A Golgi study. Proc. R. Soc. London Ser. B Biol. Sci. 1979, 204, 363–375. [Google Scholar] [CrossRef]
- Dreher, B.; Sefton, A.J.; Ni, S.Y.; Nisbett, G. The morphology, number, distribution and central projections of Class I retinal ganglion cells in albino and hooded rats. Brain. Behav. Evol. 1985, 26, 10–48. [Google Scholar] [CrossRef]
- Peichl, L. Alpha and delta ganglion cells in the rat retina. J. Comp. Neurol. 1989, 286, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Peichl, L. Alpha ganglion cells in mammalian retinae: Common properties, species differences, and some comments on other ganglion cells. Vis. Neurosci. 1991, 7, 155–169. [Google Scholar] [CrossRef]
- Peichl, L.; Ott, H.; Boycott, B.B. Alpha ganglion cells in mammalian retinae. Proc. R. Soc. London Ser. B Biol. Sci. 1987, 231, 169–197. [Google Scholar] [CrossRef]
- Krieger, B.; Qiao, M.; Rousso, D.L.; Sanes, J.R.; Meister, M. Four alpha ganglion cell types in mouse retina: Function, structure, and molecular signatures. PLoS ONE 2017, 12, e0180091. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Qiao, M.; Bei, F.; Kim, I.J.; He, Z.; Sanes, J.R. Subtype-Specific regeneration of retinal ganglion cells following axotomy: Effects of osteopontin and mtor signaling. Neuron 2015, 85, 1244–1256. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.M.; Shekhar, K.; Whitney, I.E.; Jacobi, A.; Benhar, I.; Hong, G.; Yan, W.; Adiconis, X.; Arnold, M.E.; Lee, J.M.; et al. Single-Cell Profiles of Retinal Ganglion Cells Differing in Resilience to Injury Reveal Neuroprotective Genes. Neuron 2019, 104, 1039–1055.e12. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Villegas, B.; Gallego-Ortega, A.; Miralles de Imperial-Ollero, J.A.; Martínez de la Casa, J.M.; García Feijoo, J.; Vidal-Sanz, M. Photosensitive ganglion cells: A diminutive, yet essential population. Arch. Soc. Esp. Oftalmol. 2021, 96, 299–315. [Google Scholar] [CrossRef]
- Reifler, A.N.; Chervenak, A.P.; Dolikian, M.E.; Benenati, B.A.; Meyers, B.S.; Demertzis, Z.D.; Lynch, A.M.; Li, B.Y.; Wachter, R.D.; Abufarha, F.S.; et al. The rat retina has five types of ganglion-cell photoreceptors. Exp. Eye Res. 2015, 130, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pack, W.; Hill, D.D.; Wong, K.Y. Melatonin modulates M4-type ganglion-cell photoreceptors. Neuroscience 2015, 303, 178–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, C.-A.; Li, H.; Zhang, Z.; Kiyama, T.; Panda, S.; Hattar, S.; Ribelayga, C.P.; Mills, S.L.; Wang, S.W. T-box transcription regulator Tbr2 is essential for the formation and maintenance of Opn4/melanopsin-expressing intrinsically photosensitive retinal ganglion cells. J. Neurosci. 2014, 34, 13083–13095. [Google Scholar] [CrossRef]
- Sweeney, N.T.; Tierney, H.; Feldheim, D.A. Tbr2 is required to generate a neural circuit mediating the pupillary light reflex. J. Neurosci. 2014, 34, 5447–5453. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-K.; Kiyama, T.; Weber, N.; Whitaker, C.M.; Pan, P.; Badea, T.C.; Massey, S.C.; Mao, C.-A. Characterization of Tbr2-expressing retinal ganglion cells. J. Comp. Neurol. 2021, 529, 3513–3532. [Google Scholar] [CrossRef]
- Estevez, M.E.; Fogerson, P.M.; Ilardi, M.C.; Borghuis, B.G.; Chan, E.; Weng, S.; Auferkorte, O.N.; Demb, J.B.; Berson, D.M. Form and Function of the M4 Cell, an Intrinsically Photosensitive Retinal Ganglion Cell Type Contributing to Geniculocortical Vision. J. Neurosci. 2012, 32, 13608–13620. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.M.; Alam, N.M.; Chen, S.; Kofuji, P.; Li, W.; Prusky, G.T.; Hattar, S. A Role for Melanopsin in Alpha Retinal Ganglion Cells and Contrast Detection. Neuron 2014, 82, 781–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoda, T.; Okabe, Y.; Schmidt, T.M. Overlapping morphological and functional properties between M4 and M5 intrinsically photosensitive retinal ganglion cells. J. Comp. Neurol. 2020, 528, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Ortega, A.; Salinas-Navarro, M.; Valiente-Soriano, F.; Aviles-Trigueros, M.; de la Villa, P.; Vidal-Sanz, M. Quantification of M4 intrinsically photosensitive retinal ganglion cells in pigmented mice. Ophthalmic Res. 2021, 64, 1–120. [Google Scholar] [CrossRef]
- Smeds, L.; Takeshita, D.; Turunen, T.; Tiihonen, J.; Westö, J.; Martyniuk, N.; Seppänen, A.; Ala-Laurila, P. Paradoxical Rules of Spike Train Decoding Revealed at the Sensitivity Limit of Vision. Neuron 2019, 104, 576–587.e11. [Google Scholar] [CrossRef]
- Sonoda, T.; Lee, S.K.; Birnbaumer, L.; Schmidt, T.M. Melanopsin Phototransduction Is Repurposed by ipRGC Subtypes to Shape the Function of Distinct Visual Circuits. Neuron 2018, 99, 754–767.e4. [Google Scholar] [CrossRef]
- Sexton, T.J.; Bleckert, A.; Turner, M.H.; Van Gelder, R.N. Type I intrinsically photosensitive retinal ganglion cells of early post-natal development correspond to the M4 subtype. Neural Dev. 2015, 10, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadal-Nicolás, F.M.; Sobrado-Calvo, P.; Jiménez-López, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Long-Term Effect of Optic Nerve Axotomy on the Retinal Ganglion Cell Layer. Investig. Opthalmol. Vis. Sci. 2015, 56, 6095. [Google Scholar] [CrossRef]
- Vidal-Villegas, B.; Di Pierdomenico, J.; De Imperial-Ollero, J.A.M.; Ortín-Martínez, A.; Nadal-Nicolás, F.M.; Bernal-Garro, J.M.; Navarro, N.C.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Melanopsin+RGCs are fully resistant to NMDA-Induced excitotoxicity. Int. J. Mol. Sci. 2019, 20, 3012. [Google Scholar] [CrossRef] [Green Version]
- VanderWall, K.B.; Lu, B.; Alfaro, J.S.; Allsop, A.R.; Carr, A.S.; Wang, S.; Meyer, J.S. Differential susceptibility of retinal ganglion cell subtypes in acute and chronic models of injury and disease. Sci. Rep. 2020, 10, 17359. [Google Scholar] [CrossRef]
- Daniel, S.; Clark, A.F.; McDowell, C.M. Subtype-specific response of retinal ganglion cells to optic nerve crush. Cell Death Discov. 2018, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Alarcón-Martínez, L.; de la Villa, P.; Avilés-Trigueros, M.; Blanco, R.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Short and long term axotomy-induced ERG changes in albino and pigmented rats. Mol. Vis. 2009, 15, 2373–2383. [Google Scholar]
- Alarcón-Martínez, L.; Avilés-Trigueros, M.; Galindo-Romero, C.; Valiente-Soriano, J.; Agudo-Barriuso, M.; Villa, P. de la Villegas-Pérez, M.P.; Vidal-Sanz, M. ERG changes in albino and pigmented mice after optic nerve transection. Vis. Res. 2010, 50, 2176–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salinas-Navarro, M.; Alarcón-Martínez, L.; Valiente-Soriano, F.J.; Jiménez-López, M.; Mayor-Torroglosa, S.; Avilés-Trigueros, M.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Ocular hypertension impairs optic nerve axonal transport leading to progressive retinal ganglion cell degeneration. Exp. Eye Res. 2010, 90, 168–183. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Ortega, A.; Norte-Muñoz, M.; Miralles de Imperial-Ollero, J.A.; Bernal-Garro, J.M.; Valiente-Soriano, F.J.; de la Villa Polo, P.; Avilés-Trigueros, M.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Functional and morphological alterations in a glaucoma model of acute ocular hypertension. Prog. Brain Res. 2020, 256, 1–29. [Google Scholar] [CrossRef]
- Nadal-Nicolás, F.M.; Jiménez-López, M.; Sobrado-Calvo, P.; Nieto-López, L.; Cánovas-Martínez, I.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo, M. Brn3a as a marker of retinal ganglion cells: Qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3860–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadal-Nicolás, F.M.; Jiménez-López, M.; Salinas-Navarro, M.; Sobrado-Calvo, P.; Alburquerque-Béjar, J.J.; Vidal-Sanz, M.; Agudo-Barriuso, M. Whole Number, Distribution and Co-Expression of Brn3 Transcription Factors in Retinal Ganglion Cells of Adult Albino and Pigmented Rats. PLoS ONE 2012, 7, e49830. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Nicolás, F.M.; Salinas-Navarro, M.; Jiménez-López, M.; Sobrado-Calvo, P.; Villegas-Pérez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Displaced retinal ganglion cells in albino and pigmented rats. Front. Neuroanat. 2014, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Bui, B.V.; Fortune, B. Ganglion cell contributions to the rat full-field electroretinogram. J. Physiol. 2004, 555, 153–173. [Google Scholar] [CrossRef] [Green Version]
- Sieving, P.A.; Frishman, L.J.; Steinberg, R.H. Scotopic threshold response of proximal retina in cat. J. Neurophysiol. 1986, 56, 1049–1061. [Google Scholar] [CrossRef]
- Frishman, L.J.; Steinberg, R.H. Intraretinal analysis of the threshold dark-adapted ERG of cat retina. J. Neurophysiol. 1989, 61, 1221–1232. [Google Scholar] [CrossRef]
- Frishman, L.J.; Shen, F.F.; Du, L.; Robson, J.G.; Harwerth, R.S.; Smith, E.L., 3rd; Carter-Dawson, L.; Crawford, M.L. The scotopic electroretinogram of macaque after retinal ganglion cell loss from experimental glaucoma. Invest. Ophthalmol. Vis. Sci. 1996, 37, 125–141. [Google Scholar]
- Bui, B.V.; Edmunds, B.; Cioffi, G.A.; Fortune, B. The gradient of retinal functional changes during acute intraocular pressure elevation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 202–213. [Google Scholar] [CrossRef]
- Fortune, B.; Bui, B.V.; Morrison, J.C.; Johnson, E.C.; Dong, J.; Cepurna, W.O.; Jia, L.; Barber, S.; Cioffi, G.A. Selective ganglion cell functional loss in rats with experimental glaucoma. Invest. Ophthalmol. Vis. Sci. 2004, 45, 1854–1862. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Navarro, M.; Alarcón-Martínez, L.; Valiente-Soriano, F.J.; Ortín-Martínez, A.; Jiménez-López, M.; Avilés-Trigueros, M.; Villegas-Pérez, M.P.; de la Villa, P.; Vidal-Sanz, M. Functional and morphological effects of laser-induced ocular hypertension in retinas of adult albino Swiss mice. Mol. Vis. 2009, 15, 2578–2598. [Google Scholar] [PubMed]
- Cuenca, N.; Pinilla, I.; Fernández-Sánchez, L.; Salinas-Navarro, M.; Alarcón-Martínez, L.; Avilés-Trigueros, M.; de la Villa, P.; Miralles de Imperial, J.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Changes in the inner and outer retinal layers after acute increase of the intraocular pressure in adult albino Swiss mice. Exp. Eye Res. 2010, 91, 273–285. [Google Scholar] [CrossRef]
- Gargini, C.; Bisti, S.; Demontis, G.C.; Valter, K.; Stone, J.; Cervetto, L. Electroretinogram changes associated with retinal upregulation of trophic factors: Observations following optic nerve section. Neuroscience 2004, 126, 775–783. [Google Scholar] [CrossRef]
- Agudo, M.; Pérez-Marín, M.C.; Lönngren, U.; Sobrado, P.; Conesa, A.; Cánovas, I.; Salinas-Navarro, M.; Miralles-Imperial, J.; Hallböök, F.; Vidal-Sanz, M. Time course profiling of the retinal transcriptome after optic nerve transection and optic nerve crush. Mol. Vis. 2008, 14, 1050–1063. [Google Scholar]
- Agudo, M.; Pérez-Marín, M.C.; Sobrado-Calvo, P.; Lönngren, U.; Salinas-Navarro, M.; Cánovas, I.; Nadal-Nicolás, F.M.; Miralles-Imperial, J.; Hallböök, F.; Vidal-Sanz, M. Immediate upregulation of proteins belonging to different branches of the apoptotic cascade in the retina after optic nerve transection and optic nerve crush. Investig. Ophthalmol. Vis. Sci. 2009, 50, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Asís-Martínez, A.; Gallego-Ortega, A.; Di Pierdomenico, J.; Valiente-Soriano, F.; Salinas-Navarro, M.; Vidal-Sanz, M. Long-term study of the outer retina after optic nerve transection. Ophthalmic Res. 2021, 64. [Google Scholar] [CrossRef]
- Ortín-Martínez, A.; Jimenez-Lopez, M.; Nadal-Nicolás, F.M.; Salinas-Navarro, M.; Alarcon-Martínez, L.; Sauve, Y.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Automated quantification and topographical distribution of the whole population of S- and L-cones in adult albino and pigmented rats. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3171–3183. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Nicolás, F.M.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Two methods to trace retinal ganglion cells with fluorogold: From the intact optic nerve or by stereotactic injection into the optic tract. Exp Eye Res. 2015, 131, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleckert, A.; Schwartz, G.W.; Turner, M.H.; Rieke, F.; Wong, R.O.L. Visual space is represented by nonmatching topographies of distinct mouse retinal ganglion cell types. Curr. Biol. 2014, 24, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal-Sanz, M.; Nadal-Nicolás, F.; Valiente-Soriano, F.; Agudo-Barriuso, M.; Villegas-Pérez, M. Identifying specific RGC types may shed light on their idiosyncratic responses to neuroprotection. Neural Regen. Res. 2015, 10, 1228. [Google Scholar] [CrossRef]
- Vidal-Sanz, M.; Valiente-Soriano, F.J.; Ortín-Martínez, A.; Nadal-Nicolás, F.M.; Jiménez-López, M.; Salinas-Navarro, M.; Alarcón-Martínez, L.; García-Ayuso, D.; Avilés-Trigueros, M.; Agudo-Barriuso, M.; et al. Retinal neurodegeneration in experimental glaucoma. Prog. Brain Res. 2015, 220, 1–35. [Google Scholar] [CrossRef]
- Agudo-Barriuso, M.; Nadal-Nicolás, F.; Madeira, M.; Rovere, G.; Vidal-Villegas, B.; Vidal-Sanz, M. Melanopsin expression is an indicator of the well-being of melanopsin-expressing retinal ganglion cells but not of their viability. Neural Regen. Res. 2016, 11, 1243. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.A.; Madison, R.D. Axotomized mouse retinal ganglion cells containing melanopsin show enhanced survival, but not enhanced axon regrowth into a peripheral nerve graft. Vis. Res. 2004, 44, 2667–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez de Sevilla Müller, L.; Sargoy, A.; Rodriguez, A.R.; Brecha, N.C. Melanopsin ganglion cells are the most resistant retinal ganglion cell type to axonal injury in the rat retina. PLoS ONE 2014, 9, e93274. [Google Scholar] [CrossRef]
- Nadal-Nicolás, F.M.; Madeira, M.H.; Salinas-Navarro, M.; Jiménez-López, M.; Galindo-Romero, C.; Ortín-Martínez, A.; Santiago, A.R.; Vidal-Sanz, M.; Agudo-Barriuso, M. Transient downregulation of melanopsin expression after retrograde tracing or optic nerve injury in adult rats. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4309–4323. [Google Scholar] [CrossRef] [Green Version]
- DeParis, S.; Caprara, C.; Grimm, C. Intrinsically photosensitive retinal ganglion cells are resistant to N-methyl-D-aspartic acid excitotoxicity. Mol. Vis. 2012, 18, 2814–2827. [Google Scholar]
- Honda, S.; Namekata, K.; Kimura, A.; Guo, X.; Harada, C.; Murakami, A.; Matsuda, A.; Harada, T. Survival of Alpha and Intrinsically Photosensitive Retinal Ganglion Cells in NMDA-Induced Neurotoxicity and a Mouse Model of Normal Tension Glaucoma. Invest. Ophthalmol. Vis. Sci. 2019, 60, 3696–3707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Ayuso, D.; Galindo-Romero, C.; Di Pierdomenico, J.; Vidal-Sanz, M.; Agudo-Barriuso, M.; Villegas Pérez, M.P. Light-induced retinal degeneration causes a transient downregulation of melanopsin in the rat retina. Exp. Eye Res. 2017, 161, 10–16. [Google Scholar] [CrossRef]
- Le Bail, J.C.; Varnat, F.; Nicolas, J.C.; Habrioux, G. Estrogenic and antiproliferative activities on MCF-7 human breast cancer cells by flavonoids. Cancer Lett. 1998, 130, 209–216. [Google Scholar] [CrossRef]
- Zhang, R.; Kang, K.A.; Piao, M.J.; Ko, D.O.; Wang, Z.H.; Chang, W.Y.; You, H.J.; Lee, I.K.; Kim, B.J.; Kang, S.S.; et al. Preventive effect of 7,8-dihydroxyflavone against oxidative stress induced genotoxicity. Biol. Pharm. Bull. 2009, 32, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chua, K.-W.; Chua, C.C.; Yu, H.; Pei, A.; Chua, B.H.L.; Hamdy, R.C.; Xu, X.; Liu, C.-F. Antioxidant activity of 7,8-dihydroxyflavone provides neuroprotection against glutamate-induced toxicity. Neurosci. Lett. 2011, 499, 181–185. [Google Scholar] [CrossRef]
- Lucas-Ruiz, F.; Galindo-Romero, C.; Albaladejo-García, V.; Vidal-Sanz, M.; Agudo-Barriuso, M. Mechanisms implicated in the contralateral effect in the central nervous system after unilateral injury: Focus on the visual system. Neural Regen. Res. 2021, 16, 2125–2131. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, G.-Y.; Hyun, J.W.; Hwang, H.J.; Kim, N.D.; Kim, B.-W.; Choi, Y.H. 7,8-Dihydroxyflavone exhibits anti-inflammatory properties by downregulating the NF-κB and MAPK signaling pathways in lipopolysaccharide-treated RAW264.7 cells. Int. J. Mol. Med. 2012, 29, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Park, C.; Hwang, H.J.; Kim, B.W.; Kim, G.-Y.; Kim, C.M.; Kim, N.D.; Choi, Y.H. 7,8-Dihydroxyflavone attenuates the release of pro-inflammatory mediators and cytokines in lipopolysaccharide-stimulated BV2 microglial cells through the suppression of the NF-κB and MAPK signaling pathways. Int. J. Mol. Med. 2014, 33, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.-W.; Liu, X.; Yepes, M.; Shepherd, K.R.; Miller, G.W.; Liu, Y.; Wilson, W.D.; Xiao, G.; Blanchi, B.; Sun, Y.E.; et al. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc. Natl. Acad. Sci. USA 2010, 107, 2687–2692. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Obianyo, O.; Chan, C.B.; Huang, J.; Xue, S.; Yang, J.J.; Zeng, F.; Goodman, M.; Ye, K. Biochemical and biophysical investigation of the brain-derived neurotrophic factor mimetic 7,8-dihydroxyflavone in the binding and activation of the TrkB receptor. J. Biol. Chem. 2014, 289, 27571–27584. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Xiang, Z.; Zhu, X.; Ai, Z.; Shen, J.; Huang, T.; Liu, L.; Ji, W.; Li, T. Neuroprotective Effects of 7, 8-dihydroxyflavone on Midbrain Dopaminergic Neurons in MPP(+)-treated Monkeys. Sci. Rep. 2016, 6, 34339. [Google Scholar] [CrossRef] [PubMed]
- Kong, A.W.; Della Santina, L.; Ou, Y. Probing ON and OFF Retinal Pathways in Glaucoma Using Electroretinography. Transl. Vis. Sci. Technol. 2020, 9, 14. [Google Scholar] [CrossRef]
- Vidal-Sanz, M.; Bray, G.M.; Villegas-Pérez, M.P.; Thanos, S.; Aguayo, A.J. Axonal regeneration and synapse formation in the superior colliculus by retinal ganglion cells in the adult rat. J. Neurosci. 1987, 7, 2894–2909. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.C.; Johnson, E.C.; Cepurna, W.O.; Funk, R.H. Microvasculature of the rat optic nerve head. Invest. Ophthalmol. Vis. Sci. 1999, 40, 1702–1709. [Google Scholar] [PubMed]
- Sugiyama, K.; Gu, Z.B.; Kawase, C.; Yamamoto, T.; Kitazawa, Y. Optic nerve and peripapillary choroidal microvasculature of the rat eye. Invest. Ophthalmol. Vis. Sci. 1999, 40, 3084–3090. [Google Scholar] [PubMed]
- Valiente-Soriano, F.J.; Ortín-Martínez, A.; Pierdomenico, J.D.; García-Ayuso, D.; Gallego-Ortega, A.; Miralles de Imperial-Ollero, J.A.; Jiménez-López, M.; Villegas-Pérez, M.P.; Wheeler, L.A.; Vidal-Sanz, M. Topical brimonidine or intravitreal bdnf, cntf, or bfgf protect cones against phototoxicity. Transl. Vis. Sci. Technol. 2019, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortín-Martínez, A.; Valiente-Soriano, F.J.; García-Ayuso, D.; Alarcón-Martínez, L.; Jiménez-López, M.; Bernal-Garro, J.M.; Nieto-López, L.; Nadal-Nicolás, F.M.; Villegas-Péez, M.P.; Wheeler, L.A.; et al. A novel in vivo model of focal light emitting diode-induced cone-photoreceptor phototoxicity: Neuroprotection afforded by brimonidine, BDNF, PEDF or bFGF. PLoS ONE 2014, 9, e113798. [Google Scholar] [CrossRef]
- Ortín-Martínez, A.; Nadal-Nicolás, F.M.; Jimeńez-López, M.; Alburquerque-Béjar, J.J.; Nieto-Loṕez, L.; Garcia-Ayuso, D.; Villegas-Pérez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Number and distribution of mouse retinal cone photoreceptors: Differences between an albino (Swiss) and a pigmented (C57/BL6) strain. PLoS ONE 2014, 9, e102392. [Google Scholar] [CrossRef]
- Ortín-Martínez, A.; Salinas-Navarro, M.; Nadal-Nicolás, F.M.; Jiménez-López, M.; Valiente-Soriano, F.J.; García-Ayuso, D.; Bernal-Garro, J.M.; Avilés-Trigueros, M.; Agudo-Barriuso, M.; Villegas-Pérez, M.P.; et al. Laser-induced ocular hypertension in adult rats does not affect non-RGC neurons in the ganglion cell layer but results in protracted severe loss of cone-photoreceptors. Exp. Eye Res. 2015, 132, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Galindo-Romero, C.; Jiménez-López, M.; García-Ayuso, D.; Salinas-Navarro, M.; Nadal-Nicolás, F.M.; Agudo-Barriuso, M.; Villegas-Pérez, M.P.; Avilés-Trigueros, M.; Vidal-Sanz, M. Number and spatial distribution of intrinsically photosensitive retinal ganglion cells in the adult albino rat. Exp. Eye Res. 2013, 108, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Di Pierdomenico, J.; Scholz, R.; Valiente-Soriano, F.J.; Sánchez-Migallón, M.C.; Vidal-Sanz, M.; Langmann, T.; Agudo-Barriuso, M.; García-Ayuso, D.; Villegas-Pérez, M.P. Neuroprotective effects of FGF2 and minocycline in two animal models of inherited retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4392–4403. [Google Scholar] [CrossRef] [Green Version]
- Miralles de Imperial-Ollero, J.A.; Gallego-Ortega, A.; Norte-Muñoz, M.; Di Pierdomenico, J.; Valiente-Soriano, F.J.; Vidal-Sanz, M. An in vivo model of focal light emitting diode-induced cone photoreceptor phototoxicity in adult pigmented mice: Protection with bFGF. Exp. Eye Res. 2021, 211, 108746. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brn3a | OPN | OPN-Trb2 | OPN-Brn3a | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RE | 7 d | 14 d | 21 d | RE | 7 d | 14 d | 21 d | RE | 7 d | 14 d | 21 d | RE | 7 d | 14 d | 21 d | |
| RE (n) | 42 | 42 | 42 | 42 | ||||||||||||
| Mean | 81,085 | 1917 | 1015 | 782 | ||||||||||||
| SD | 3056 | 229 | 120 | 100 | ||||||||||||
| VEH (n) | 6 | 6 | 5 | 6 | 6 | 5 | 6 | 6 | 5 | 6 | 6 | 5 | ||||
| Mean | 37,121 * | 22,754 *‡ | 8626 *‡ | 735 * | 729 * | 679 * | 269 * | 233 * | 202 * | 3 * | 3 * | 4 * | ||||
| SD | 4312 | 4739 | 1295 | 73 | 64 | 97 | 43 | 39 | 75 | 2 | 1 | 2 | ||||
| % | 100 | 45.7 | 28.1 | 10.6 | 100 | 38.4 | 38.0 | 35.4 | 100 | 26.5 | 22.9 | 19.9 | 100 | 0.4 | 0.4 | 0.5 |
| DHF (n) | 6 | 6 | 5 | 6 | 6 | 5 | 6 | 5 | 5 | 6 | 5 | 5 | ||||
| Mean | 76,051 | 52,620 *†‡ | 12,089 *†‡ | 1115 *† | 956 *†‡ | 714 *‡ | 669 *† | 341 *†‡ | 205 *‡ | 3 * | 3 * | 4 * | ||||
| SD | 4312 | 4581 | 1591 | 148 | 60 | 60 | 127 | 41 | 32 | 1 | 2 | 2 | ||||
| % | 100 | 93.8 | 64.9 | 14.9 | 100 | 58.1 | 49.8 | 37.2 | 100 | 65.9 | 33.7 | 20.2 | 100 | 0.4 | 0.4 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallego-Ortega, A.; Vidal-Villegas, B.; Norte-Muñoz, M.; Salinas-Navarro, M.; Avilés-Trigueros, M.; Villegas-Pérez, M.P.; Vidal-Sanz, M. 7,8-Dihydroxiflavone Maintains Retinal Functionality and Protects Various Types of RGCs in Adult Rats with Optic Nerve Transection. Int. J. Mol. Sci. 2021, 22, 11815. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111815
Gallego-Ortega A, Vidal-Villegas B, Norte-Muñoz M, Salinas-Navarro M, Avilés-Trigueros M, Villegas-Pérez MP, Vidal-Sanz M. 7,8-Dihydroxiflavone Maintains Retinal Functionality and Protects Various Types of RGCs in Adult Rats with Optic Nerve Transection. International Journal of Molecular Sciences. 2021; 22(21):11815. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111815
Chicago/Turabian StyleGallego-Ortega, Alejandro, Beatriz Vidal-Villegas, María Norte-Muñoz, Manuel Salinas-Navarro, Marcelino Avilés-Trigueros, María Paz Villegas-Pérez, and Manuel Vidal-Sanz. 2021. "7,8-Dihydroxiflavone Maintains Retinal Functionality and Protects Various Types of RGCs in Adult Rats with Optic Nerve Transection" International Journal of Molecular Sciences 22, no. 21: 11815. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111815