
Developmental Toxicology of Metal Mixtures in Drosophila: Unique Properties of Potency and Interactions of Mercury Isoforms



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
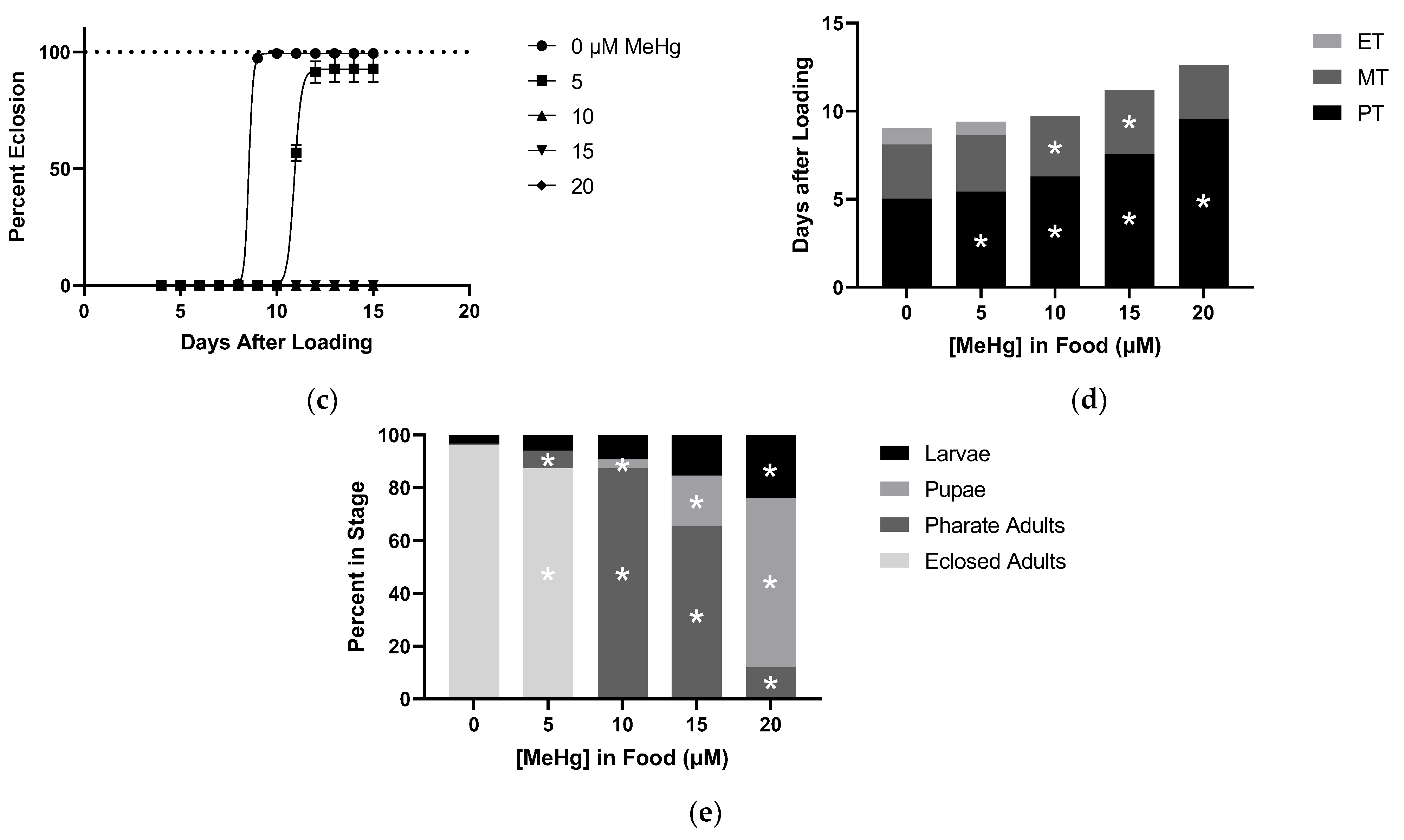
2.1. Effects of MeHg on Developmental Time Course
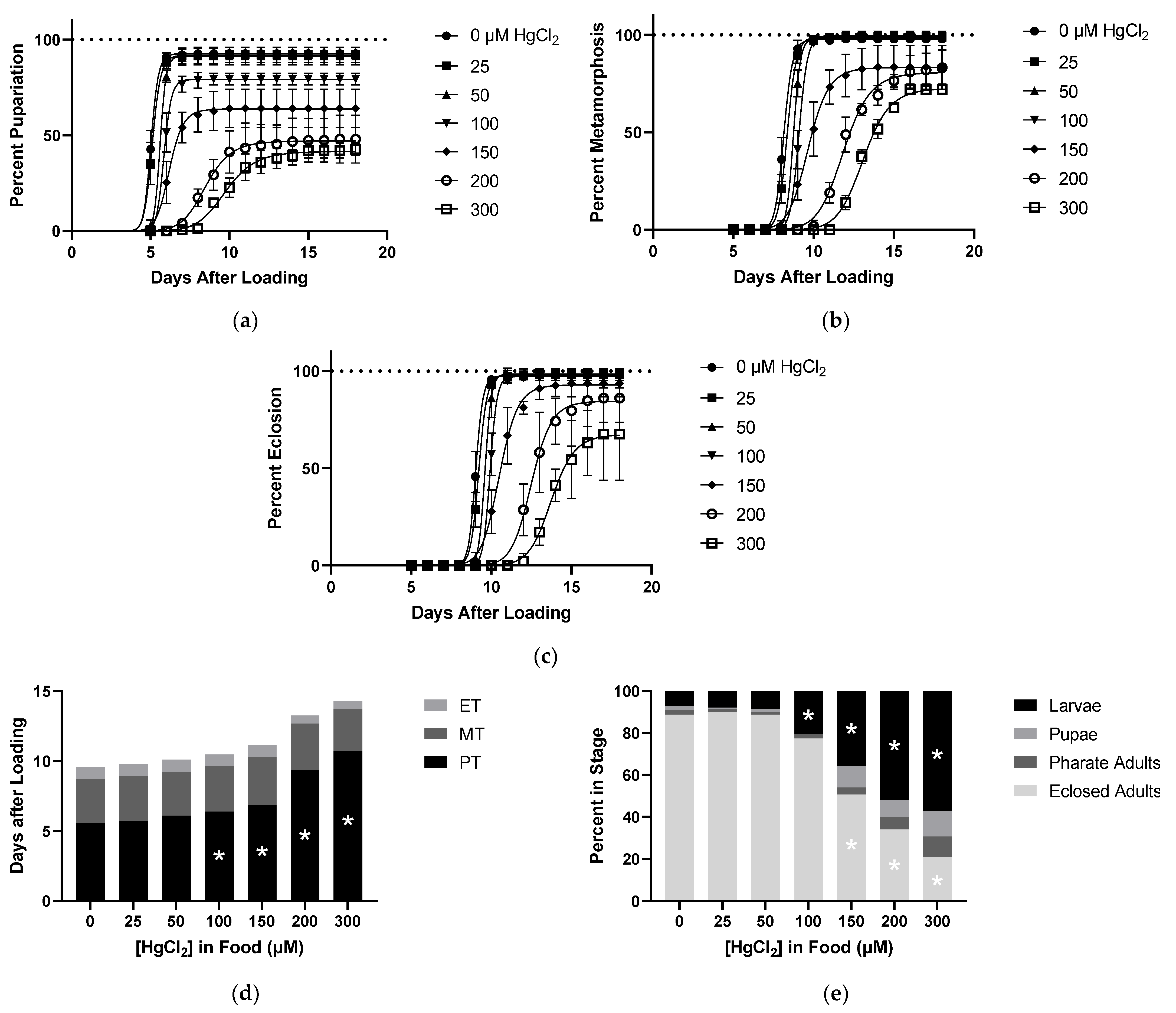
2.2. Effects of HgCl2 on Developmental Time Course
2.3. Effects of As on Developmental Time Course
2.4. Effects of Pb on Developmental Time Course
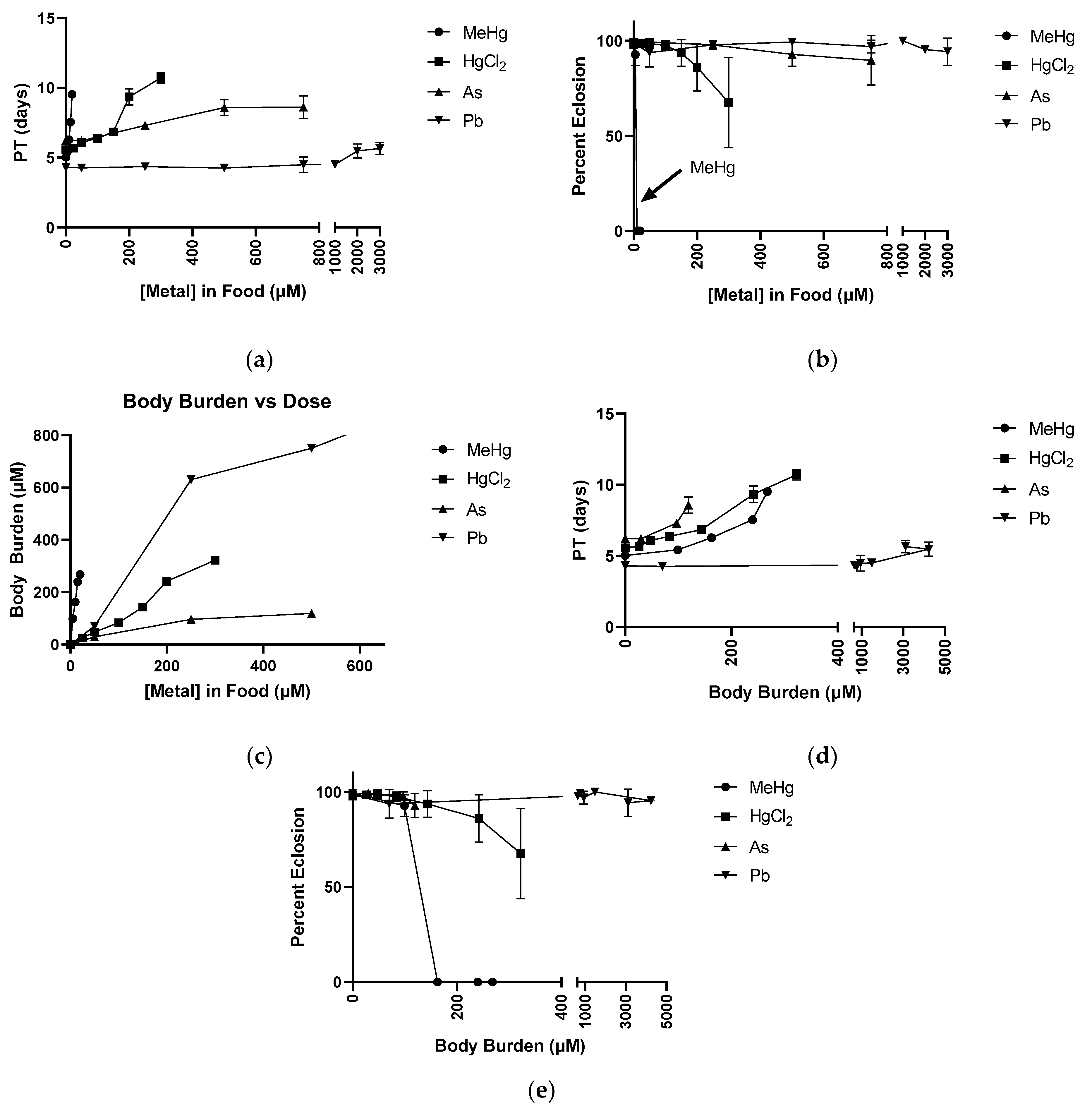
2.5. Dose Response with Respect to Body Burden for MeHg, HgCl2, As, and Pb
2.6. Effects of MeHg and HgCl2 Mixtures on Developmental Time Course
2.7. Effects of MeHg and As Mixtures on Developmental Time Course
2.8. Effects of MeHg and Pb mixtures on Developmental Time Course
2.9. Eclosion and Body Burden Assessment of MeHg and HgCl2 Mixtures
3. Discussion
4. Materials and Methods
4.1. Drosophila Stocks
4.2. Developmental Time Course and Body Burden Measurements
4.3. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cobbina, S.J.; Chen, Y.; Zhou, Z.; Wu, X.; Feng, W.; Wang, W.; Mao, G.; Xu, H.; Zhang, Z.; Wu, X.; et al. Low concentration toxic metal mixture interactions: Effects on essential and non-essential metals in brain, liver, and kidneys of mice on sub-chronic exposure. Chemosphere 2015, 132, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Dirilgen, N. Mercury and lead: Assessing the toxic effects on growth and metal accumulation by Lemna minor. Ecotoxicol. Environ. Saf. 2011, 74, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Heyes, A.; Rowe, C.L.; Conrad, P. Differential Patterns of Accumulation and Retention of Dietary Trace Elements Associated with Coal Ash During Larval Development and Metamorphosis of an Amphibian. Arch. Environ. Contam. Toxicol. 2014, 66, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Rebolledo, U.A.; Páez-Osuna, F.; Fernández, R. Single and mixture toxicity of As, Cd, Cr, Cu, Fe, Hg, Ni, Pb, and Zn to the rotifer Proales similis under different salinities. Environ. Pollut. 2021, 271, 116357. [Google Scholar] [CrossRef]
- Zhou, Q.; Gu, Y.; Yue, X.; Mao, G.; Wang, Y.; Su, H.; Xu, J.; Shi, H.; Zou, B.; Zhao, J.; et al. Combined toxicity and underlying mechanisms of a mixture of eight heavy metals. Mol. Med. Rep. 2016, 15, 859–866. [Google Scholar] [CrossRef]
- Jadhav, S.H.; Sarkar, S.N.; Patil, R.D.; Tripathi, H.C. Effects of Subchronic Exposure via Drinking Water to a Mixture of Eight Water-Contaminating Metals: A Biochemical and Histopathological Study in Male Rats. Arch. Environ. Contam. Toxicol. 2007, 53, 667–677. [Google Scholar] [CrossRef]
- Saghazadeh, A.; Rezaei, N. Systematic review and meta-analysis links autism and toxic metals and highlights the impact of country development status: Higher blood and erythrocyte levels for mercury and lead, and higher hair antimony, cadmium, lead, and mercury. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 340–368. [Google Scholar] [CrossRef]
- Kenston, S.S.F.; Su, H.; Li, Z.; Kong, L.; Wang, Y.; Song, X.; Gu, Y.; Barber, T.; Aldinger, J.; Hua, Q.; et al. The systemic toxicity of heavy metal mixtures in rats. Toxicol. Res. 2018, 7, 396–407. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, R.; Ramond, A.; O’Keeffe, L.; Shahzad, S.; Kunutsor, S.; Muka, T.; Gregson, J.; Willeit, P.; Warnakula, S.; Khan, H.; et al. Environmental toxic metal contaminants and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2018, 362, k3310. [Google Scholar] [CrossRef] [Green Version]
- Agency for Toxic Substances and Disease Registry. 2019. Available online: https://www.atsdr.cdc.gov/spl/ (accessed on 10 August 2021).
- van Wijngaarden, E.; Winters, P.C.; Cory-Slechta, D.A. Blood lead levels in relation to cognitive function in older U.S. adults. NeuroToxicology 2011, 32, 110–115. [Google Scholar] [CrossRef]
- Jia, Q.; Du, G.; Li, Y.; Wang, Z.; Xie, J.; Guangming, Y.; Yin, G.; Zhang, S.; Gao, Y.; Zhou, F.; et al. Pb2+ modulates ryanodine receptors from the endoplasmic reticulum in rat brain. Toxicol. Appl. Pharmacol. 2018, 338, 103–111. [Google Scholar] [CrossRef]
- Moore, C.L.; Flanigan, T.J.; Law, C.D.; Loukotková, L.; Woodling, K.A.; da Costa, G.G.; Fitzpatrick, S.C.; Ferguson, S.A. Developmental neurotoxicity of inorganic arsenic exposure in Sprague-Dawley rats. Neurotoxicology Teratol. 2019, 72, 49–57. [Google Scholar] [CrossRef]
- Wu, M.; Yan, C.; Xu, J.; Wu, W.; Li, H.; Zhou, X. Umbilical cord blood mercury levels in China. J. Environ. Sci. 2013, 25, 386–392. [Google Scholar] [CrossRef]
- Kolbinger, V.; Engström, K.; Berger, U.; Bose-O’Reilly, S. Polymorphisms in potential mercury transporter ABCC2 and neurotoxic symptoms in populations exposed to mercury vapor from goldmining. Environ. Res. 2019, 176, 108512. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P. Developmental neurotoxicity of industrial chemicals: A silent pandemic. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Model. Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonini, N.M.; Fortini, M.E. Human Neurodegenerative Disease Modeling Using Drosophila. Annu. Rev. Neurosci. 2003, 26, 627–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akins, J.M.; Schroeder, J.A.; Brower, D.L.; Aposhian, H.V. Evaluation of Drosophila melanogaster as an alternative animal for studying the neurotoxicity of heavy metals. BioMetals 1992, 5, 111–120. [Google Scholar] [CrossRef]
- Goldstein, S.H.; Babich, H. Differential effects of arsenite and arsenale to Drosophila melanogaster in a combined adult/developmental toxicity assay. Bull. Environ. Contam. Toxicol. 1989, 42, 276–282. [Google Scholar] [CrossRef]
- Cohn, J.; Widzowski, D.; Cory-Slechta, D.A. Lead retards development of Drosophila melanogaster. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1992, 102, 45–49. [Google Scholar] [CrossRef]
- Rand, M.D.; Vorojeikina, D.; Peppriell, A.; Gunderson, J.; Prince, L.M. Drosophotoxicology: Elucidating Kinetic and Dynamic Pathways of Methylmercury Toxicity in a Drosophila Model. Front. Genet. 2019, 10, 666. [Google Scholar] [CrossRef] [Green Version]
- Abnoos, H.; Fereidoni, M.; Mahdavi-Shahri, N.; Haddad, F.; Jalal, R. Developmental study of mercury effects on the fruit fly (Drosophila melanogaster). Interdiscip. Toxicol. 2013, 6, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Rand, M.D. Drosophotoxicology: The growing potential for Drosophila in neurotoxicology. Neurotoxicology Teratol. 2010, 32, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Morozova, T.V.; Hussain, Y.N.; Luoma, S.E.; McCoy, L.; Yamamoto, A.; Mackay, T.F.; Anholt, R.R. The Genetic Basis for Variation in Sensitivity to Lead Toxicity in Drosophila melanogaster. Environ. Health Perspect. 2016, 124, 1062–1070. [Google Scholar] [CrossRef] [Green Version]
- Frat, L.; Chertemps, T.; Pesce, E.; Bozzolan, F.; Dacher, M.; Planelló, R.; Herrero, O.; Llorente, L.; Moers, D.; Siaussat, D. Single and mixed exposure to cadmium and mercury in Drosophila melanogaster: Molecular responses and impact on post-embryonic development. Ecotoxicol. Environ. Saf. 2021, 220, 112377. [Google Scholar] [CrossRef] [PubMed]
- Simmons-Willis, T.A.; Koh, A.S.; Clarkson, T.W.; Ballatori, N. Transport of a neurotoxicant by molecular mimicry: The methylmercury–l-cysteine complex is a substrate for human L-type large neutral amino acid transporter (LAT) 1 and LAT2. Biochem. J. 2002, 367, 239–246. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Transport of Inorganic Mercury and Methylmercury in Target Tissues and Organs. J. Toxicol. Environ. Health Part B 2010, 13, 385–410. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L.; Pan, J. The role of L-type amino acid transporter 1 in human tumors. Intractable Rare Dis. Res. 2015, 4, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, L.; Galluccio, M.; Scalise, M.; Parravicini, C.; Palazzolo, L.; Eberini, I.; Indiveri, C. Novel insights into the transport mechanism of the human amino acid transporter LAT1 (SLC7A5). Probing critical residues for substrate translocation. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Boado, R.J.; Li, J.Y.; Chu, C.; Ogoshi, F.; Wise, P.; Pardridge, W.M. Site-directed mutagenesis of cysteine residues of large neutral amino acid transporter LAT1. Biochim. Biophys. Acta (BBA) Biomembr. 2005, 1715, 104–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Zhao, J.; Li, Y.; Fan, Y.; Zhu, N.; Gao, Y.; Li, B.; Liu, H.; Li, Y.-F. Demethylation of methylmercury in growing rice plants: An evidence of self-detoxification. Environ. Pollut. 2016, 210, 113–120. [Google Scholar] [CrossRef]
- Li, H.; Lin, X.; Zhao, J.; Cui, L.; Wang, L.; Gao, Y.; Li, B.; Chen, C.; Li, Y.-F. Intestinal Methylation and Demethylation of Mercury. Bull. Environ. Contam. Toxicol. 2019, 102, 597–604. [Google Scholar] [CrossRef]
- Jaszczak, J.; Wolpe, J.B.; Bhandari, R.; Jaszczak, R.G.; Halme, A. Growth Coordination During Drosophila melanogaster Imaginal Disc Regeneration Is Mediated by Signaling Through the Relaxin Receptor Lgr3 in the Prothoracic Gland. Genetics 2016, 204, 703–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddiford, L.M. Hormone receptors and the regulation of insect metamorphosis. Receptor 1993, 3, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittmers, L.E.; Wallgren, J.; Alich, A.; Aufderheide, A.C.; Rapp, G. Lead in Bone. IV. Distribution of Lead in the Human Skeleton. Arch. Environ. Health Int. J. 1988, 43, 381–391. [Google Scholar] [CrossRef]
- Castoldi, A.F.; Onishchenko, N.; Johansson, C.H.; Coccini, T.; Roda, E.; Vahter, M.; Ceccatelli, S.; Manzo, L. Neurodevelopmental toxicity of methylmercury: Laboratory animal data and their contribution to human risk assessment. Regul. Toxicol. Pharmacol. 2008, 51, 215–229. [Google Scholar] [CrossRef]
- Groth, E. Ranking the contributions of commercial fish and shellfish varieties to mercury exposure in the United States: Implications for risk communication. Environ. Res. 2010, 110, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, T.W.; Magos, L. The Toxicology of Mercury and Its Chemical Compounds. Crit. Rev. Toxicol. 2006, 36, 609–662. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.; Clarkson, T.W.; Marsh, D.O.; Amin-Zaki, L.; Tikriti, S.; Myers, G.G. Dose-response analysis of infants prenatally exposed to methyl mercury: An application of a single compartment model to single-strand hair analysis. Environ. Res. 1989, 49, 318–332. [Google Scholar] [CrossRef]
- Maull, E.A.; Ahsan, H.; Edwards, J.; Longnecker, M.; Navas-Acien, A.; Pi, J.; Silbergeld, E.K.; Styblo, M.; Tseng, C.-H.; Thayer, K.A.; et al. Evaluation of the Association between Arsenic and Diabetes: A National Toxicology Program Workshop Review. Environ. Health Perspect. 2012, 120, 1658–1670. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.A.; Khan, M.H.; Haque, M. Arsenic contamination in groundwater in Bangladesh: Implications and challenges for healthcare policy. Risk Manag. Healthc. Policy 2018, 11, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, W.P.; Chu, H.M.; How, S.W.; Fong, J.M.; Yeh, S.; Lin, C.S. Prevalence of Skin Cancer in an Endemic Area of Chronic Arsenicism in Taiwan. J. Natl. Cancer Inst. 1968, 40, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.G.M.; Opoka, R.; Kane, D.; Cartwright, I.L. Investigating Arsenic Susceptibility from a Genetic Perspective in Drosophila Reveals a Key Role for Glutathione Synthetase. Toxicol. Sci. 2008, 107, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Loewe, S. The problem of synergism and antagonism of combined drugs. Arzneimittelforschung 1953, 3, 285–290. [Google Scholar] [PubMed]
- Bliss, C.I. The Toxicity of Poisons Applied Jointly. Ann. Appl. Biol. 1939, 26, 585–615. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beamish, C.R.; Love, T.M.; Rand, M.D. Developmental Toxicology of Metal Mixtures in Drosophila: Unique Properties of Potency and Interactions of Mercury Isoforms. Int. J. Mol. Sci. 2021, 22, 12131. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212131
Beamish CR, Love TM, Rand MD. Developmental Toxicology of Metal Mixtures in Drosophila: Unique Properties of Potency and Interactions of Mercury Isoforms. International Journal of Molecular Sciences. 2021; 22(22):12131. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212131
Chicago/Turabian StyleBeamish, Catherine R., Tanzy M. Love, and Matthew D. Rand. 2021. "Developmental Toxicology of Metal Mixtures in Drosophila: Unique Properties of Potency and Interactions of Mercury Isoforms" International Journal of Molecular Sciences 22, no. 22: 12131. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212131