
In Leishmania major, the Homolog of the Oncogene PES1 May Play a Critical Role in Parasite Infectivity

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
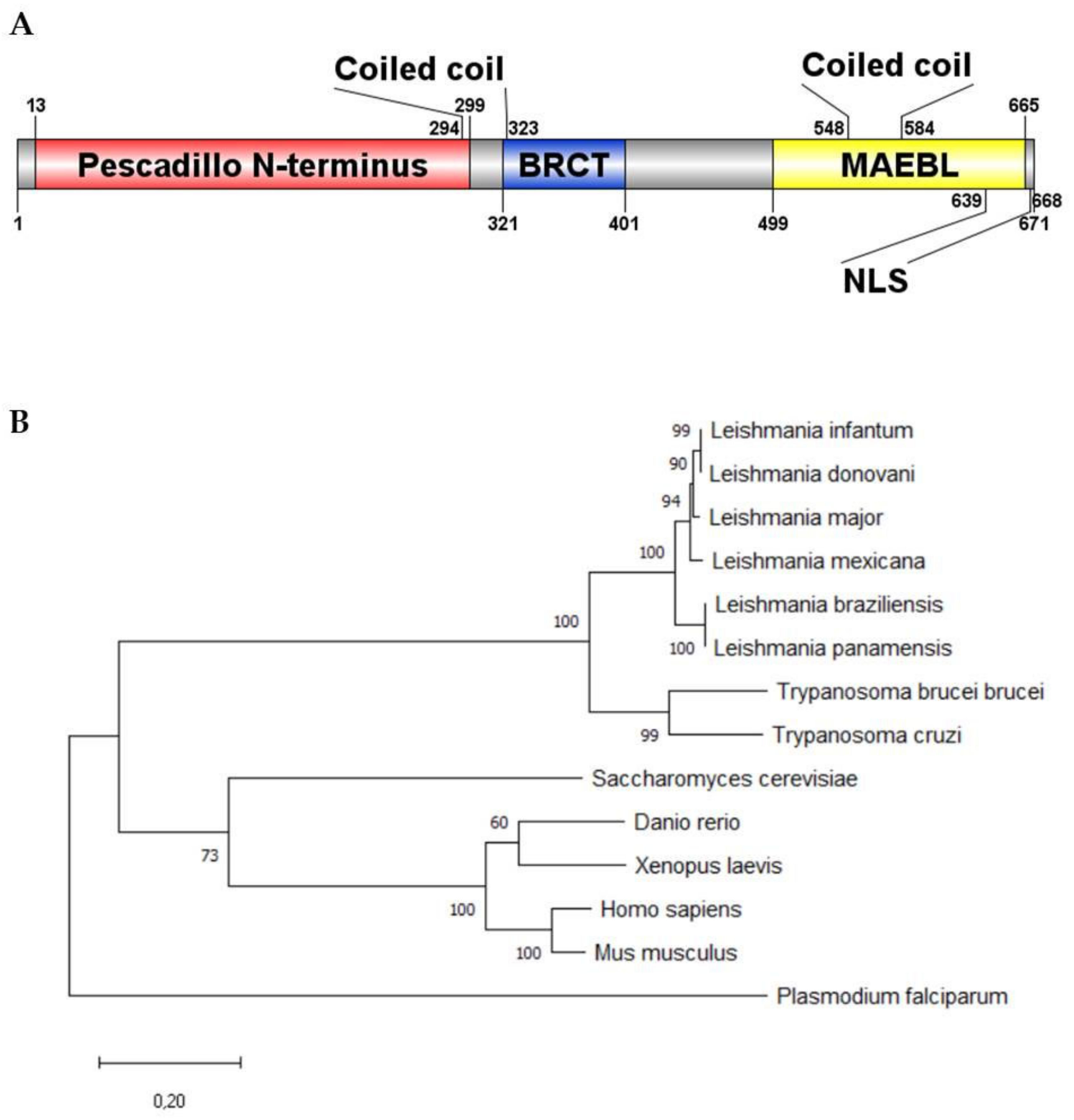
2.1. LmjPES Is Highly Conserved among Trypanosomatid Cluster
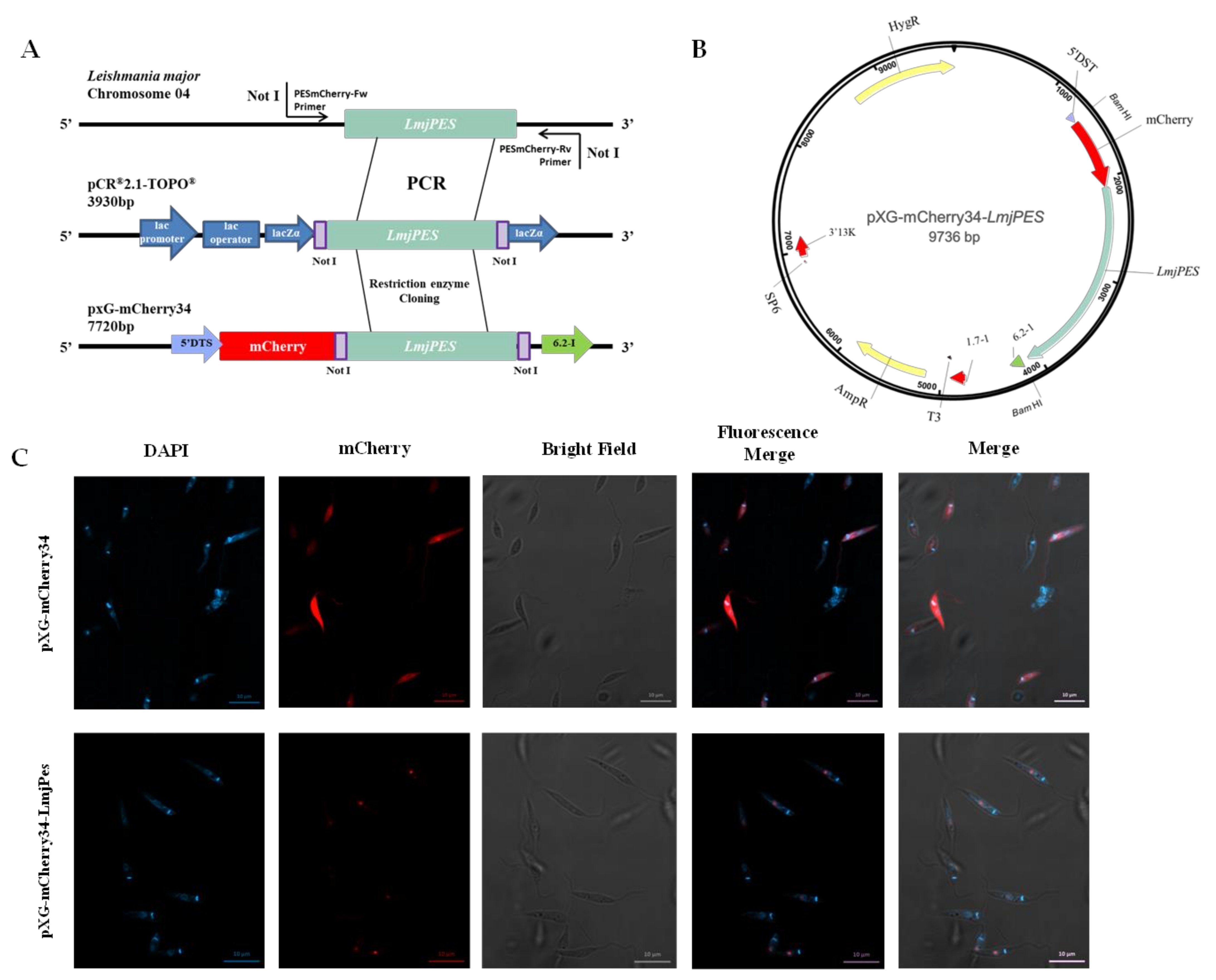
2.2. LmjPES Is a Protein Located in the Nucleus
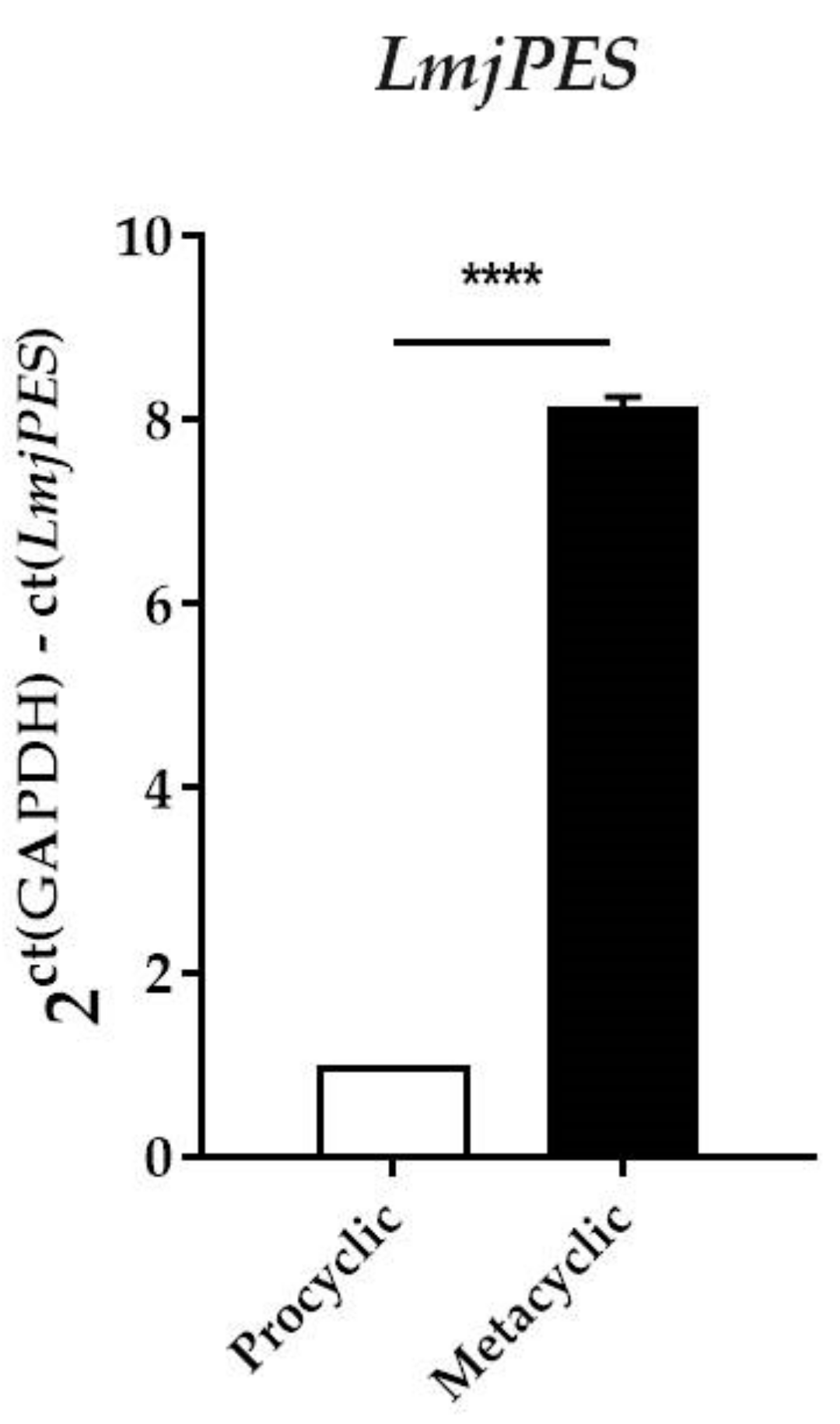
2.3. LmjPES Gene Expression was Highly Expressed in the Metacyclic Stage
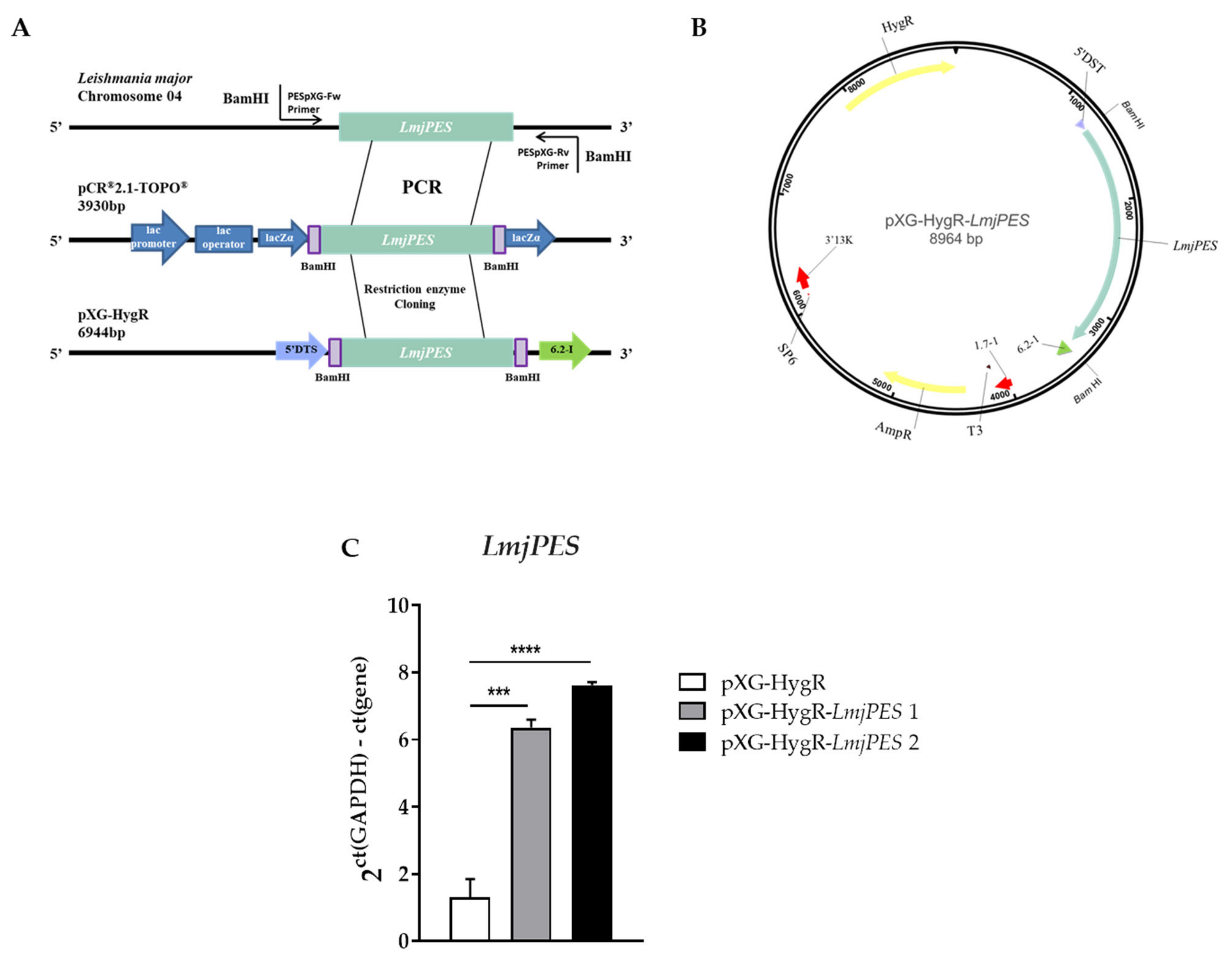
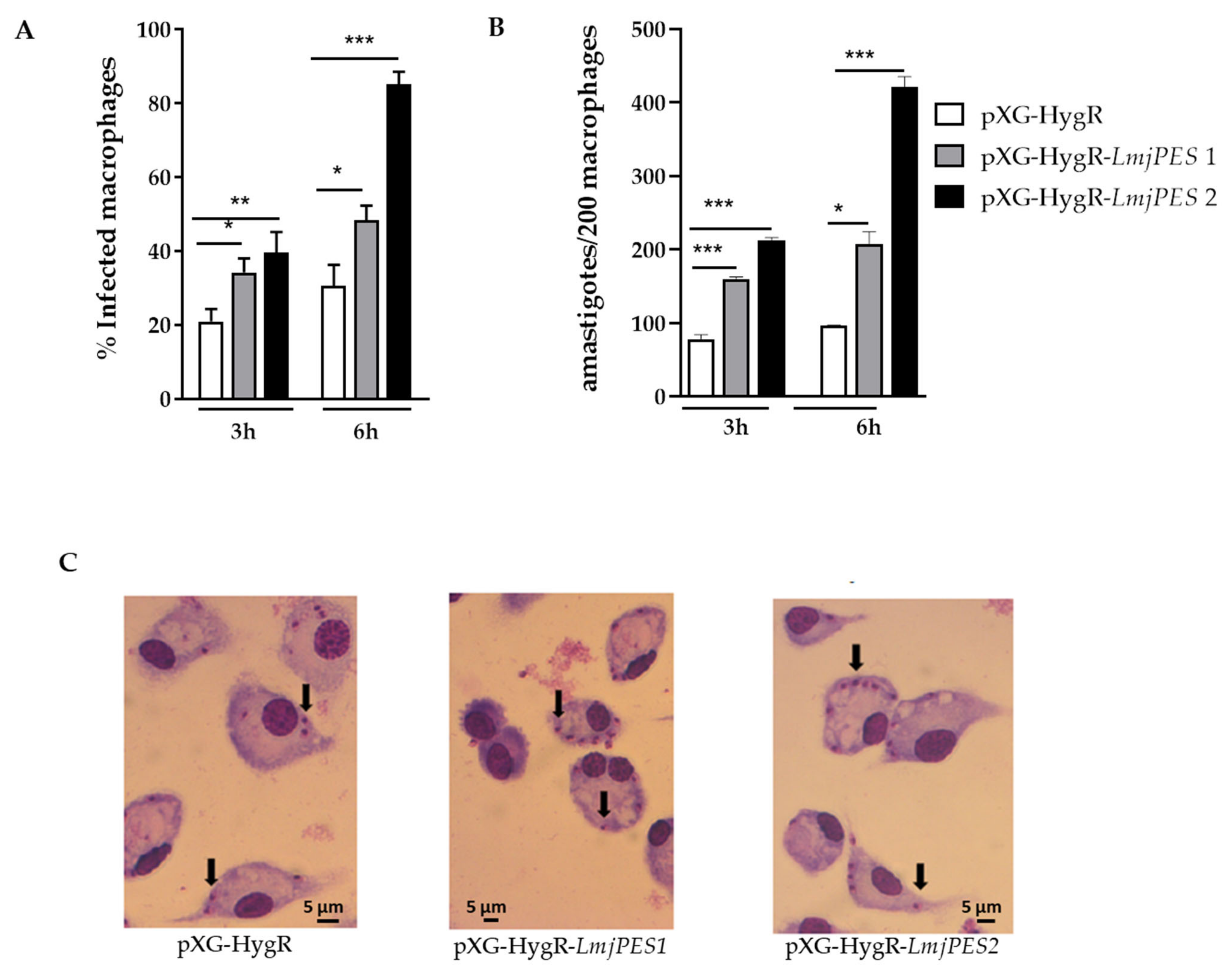
2.4. LmjPES Overexpression Increased the Infectivity Rates In Vitro
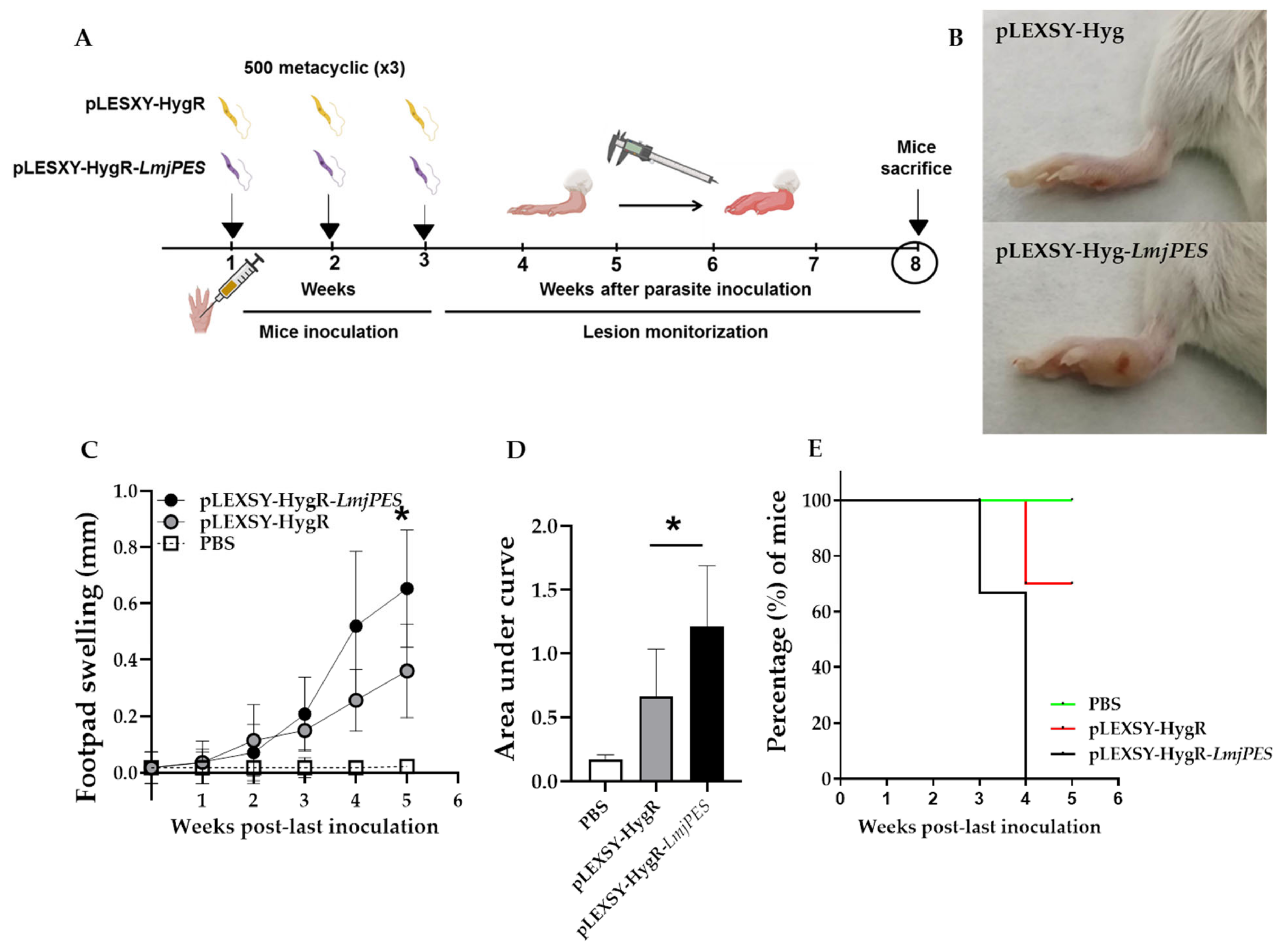
2.5. LmjPES Overexpressing Parasites Exhibited Higher and Faster Footpad Inflammation in BALB/c Mice than Non-Overexpressing Parasites
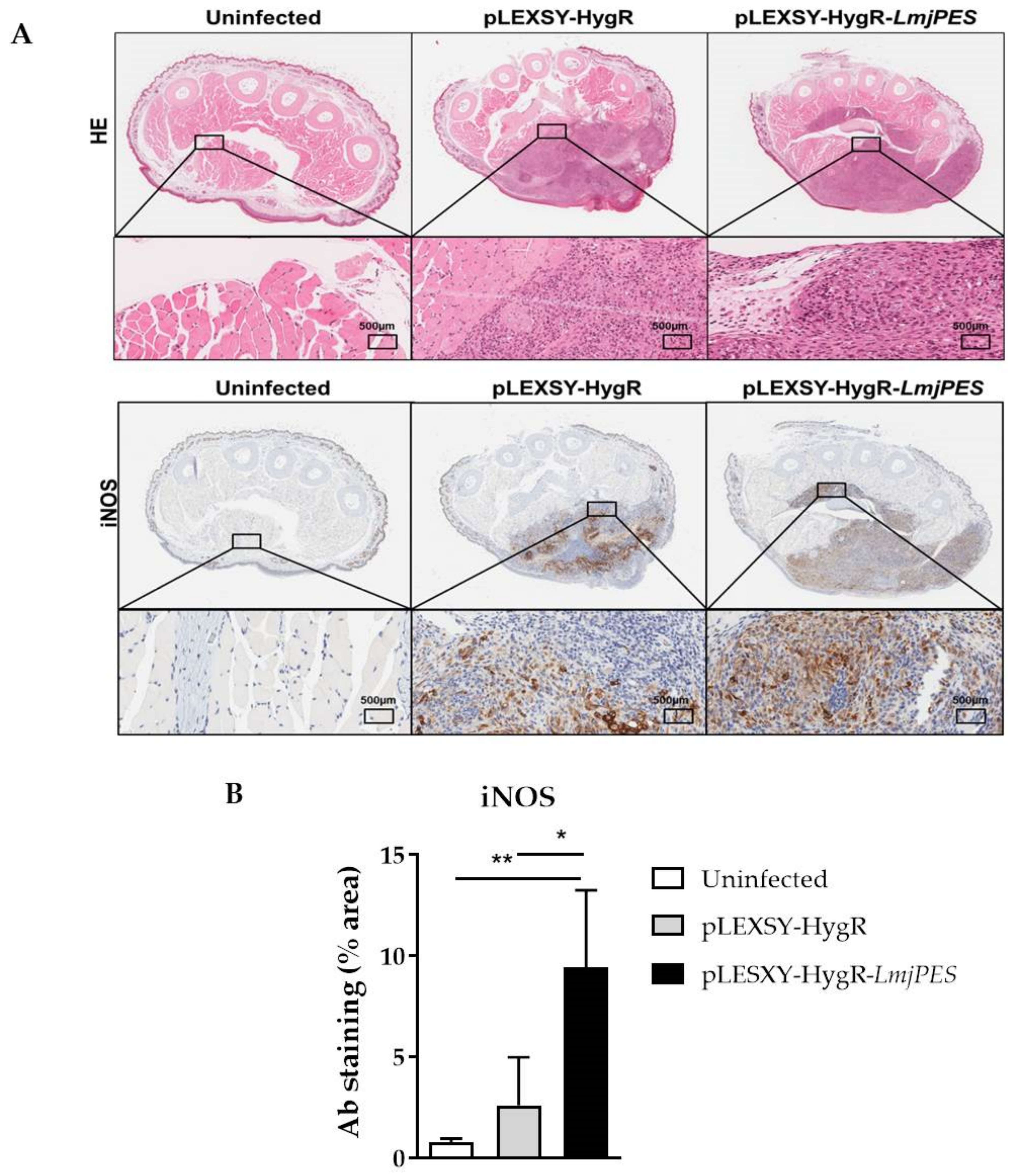
2.6. In Vivo Infection with LmjPES Overexpressing Parasites Induced a Higher Production of iNOS in the Inoculation Area
3. Discussion
4. Materials and Methods
4.1. Nucleotide and Amino Acid Sequence Alignments
4.2. Assessment of Secondary Structure and Post-Translational Modifications
4.3. Phylogenetic Analysis
4.4. Parasites and Animals
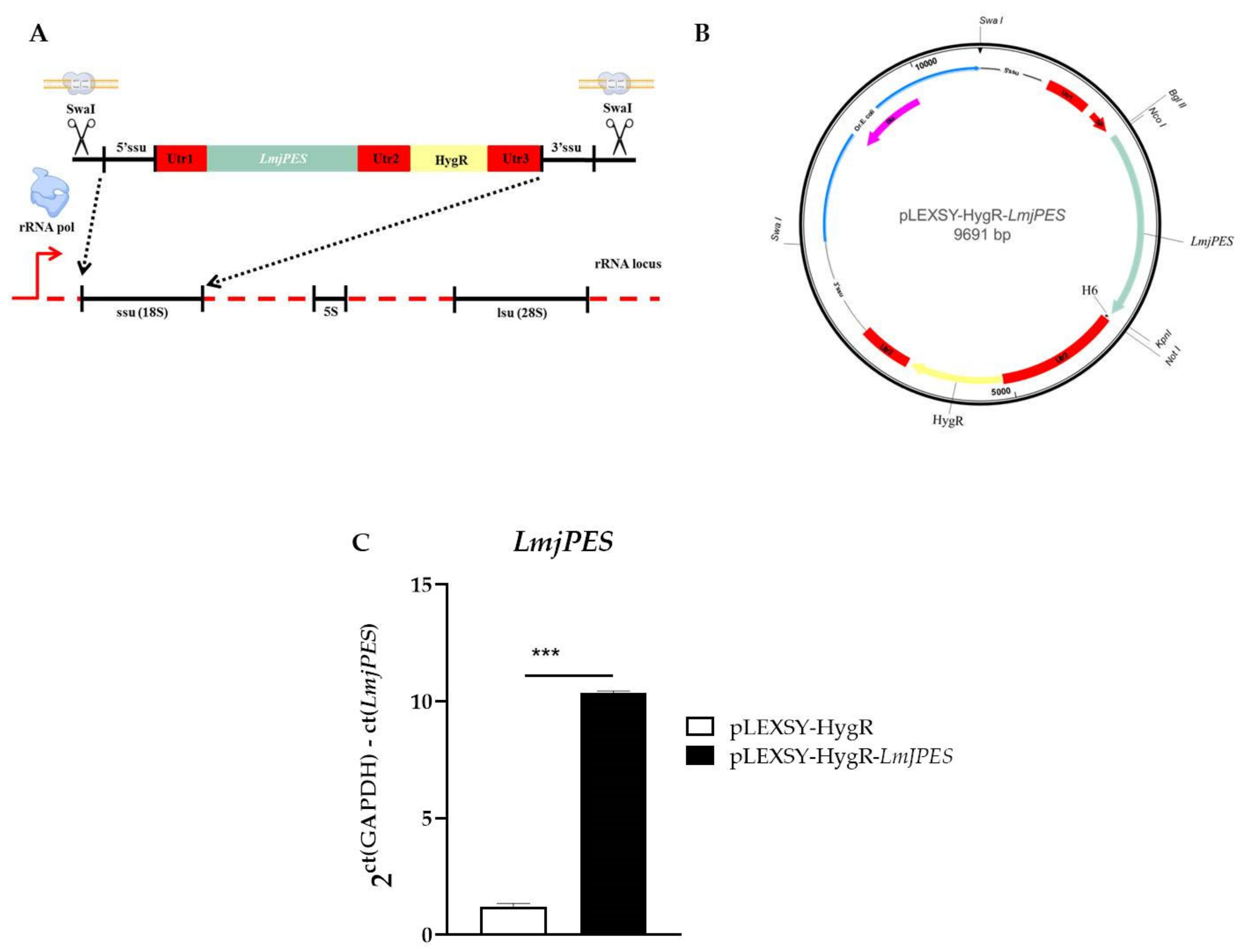
4.5. Genetic Manipulation of Leishmania major
4.6. Fluorescence Microscopy
4.7. In Vitro Infections
4.8. In Vivo Infections
4.9. RNA Extraction and Gene Expression Analysis
4.10. Immunohistochemistry Analysis:
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molyneux, D.H.; Asamoa-Bah, A.; Fenwick, A.; Savioli, L.; Hotez, P. The history of the neglected tropical disease movement. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 169–175. [Google Scholar] [CrossRef]
- Engels, D.; Zhou, X.-N. Neglected tropical diseases: An effective global response to local poverty-related disease priorities. Infect. Dis. Poverty 2020, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine (US) Forum on Microbial Threats. The Causes and Impacts of Neglected Tropical and Zoonotic Diseases: Opportunities for Integrated Intervention Strategies. Washington (DC): National Academies Press (US); 2011. Workshop Overview. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK62526/?log$=activity (accessed on 23 September 2021).
- World Health Organization. Observatory, T.G.H. Leishmaniasis . Available online: https://www.who.int/data/gho/data/themes/topics/gho-ntd-leishmaniasis (accessed on 23 September 2021).
- World Health Organization. Leishmaniasis. Available online: https://www.who.int/health-topics/leishmaniasis#tab=tab_1 (accessed on 23 September 2021).
- Balanã-Fouce, R.; Prada, C.F.; Requena, J.M.; Cushman, M.; Pommier, Y.; Álvarez-Velilla, R.; Escudero-Martínez, J.M.; Calvo-Álvarez, E.; Pérez-Pertejo, Y.; Reguera, R.M. Indotecan (LMP400) and AM13-55: Two novel indenoisoquinolines show potential for treating visceral leishmaniasis. Antimicrob. Agents Chemother. 2012, 56, 5264–5270. [Google Scholar] [CrossRef] [Green Version]
- Cojean, S.; Houze, S.; Haouchine, D.; Huteau, F.; Lariven, S.; Hubert, V.; Michard, F.; Bories, C.; Pratlong, F.; Le Bras, J.; et al. Leishmania resistance to miltefosine associated with genetic marker. Emerg. Infect. Dis. 2012, 18, 704–706. [Google Scholar] [CrossRef]
- Coelho, A.C.; Boisvert, S.; Mukherjee, A.; Leprohon, P.; Corbeil, J.; Ouellette, M. Multiple mutations in heterogeneous miltefosine-resistant Leishmania major population as determined by whole genome sequencing. PLoS Negl. Trop. Dis. 2012, 6, e1512. [Google Scholar] [CrossRef] [Green Version]
- Sundar, S.; Singh, B. Emerging therapeutic targets for treatment of leishmaniasis. Expert Opin. Ther. Targets 2018, 22, 467–486. [Google Scholar] [CrossRef]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef] [Green Version]
- Dannheisig, D.P.; Bächle, J.; Tasic, J.; Keil, M.; Pfister, A.S. The Wnt/β-Catenin Pathway is Activated as a Novel Nucleolar Stress Response. J. Mol. Biol. 2021, 433, 166719. [Google Scholar] [CrossRef]
- Ahn, C.S.; Cho, H.K.; Lee, D.-H.; Sim, H.-J.; Kim, S.-G.; Pai, H.-S. Functional characterization of the ribosome biogenesis factors PES, BOP1, and WDR12 (PeBoW), and mechanisms of defective cell growth and proliferation caused by PeBoW deficiency in Arabidopsis. J. Exp. Bot. 2016, 67, 5217–5232. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.; Wang, B.; Meng, Z.; Zhao, J.; Jin, X. PES1 is transcriptionally regulated by BRD4 and promotes cell proliferation and glycolysis in hepatocellular carcinoma. Int. J. Biochem. Cell Biol. 2018, 104, 1–8. [Google Scholar] [CrossRef]
- Lerch-Gaggl, A.; Haque, J.; Li, J.; Ning, G.; Traktman, P.; Duncan, S.A. Pescadillo is essential for nucleolar assembly, ribosome biogenesis, and mammalian cell proliferation. J. Biol. Chem. 2002, 277, 45347–45355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gessert, S.; Maurus, D.; Rössner, A.; Kühl, M. Pescadillo is required for Xenopus laevis eye development and neural crest migration. Dev. Biol. 2007, 310, 99–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.K.; Ahn, C.S.; Lee, H.-S.; Kim, J.-K.; Pai, H.-S. Pescadillo plays an essential role in plant cell growth and survival by modulating ribosome biogenesis. Plant J. 2013, 76, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Feng, Q.; Su, Y.; Dong, B.; Wu, J.; Meng, L.; Qu, L.; Shou, C. Transcriptional regulation of PES1 expression by c-Jun in colon cancer. PLoS ONE 2012, 7, e42253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, W.; Qu, L.; Meng, L.; Liu, C.; Wu, J.; Shou, C. PES1 regulates sensitivity of colorectal cancer cells to anticancer drugs. Biochem. Biophys. Res. Commun. 2013, 431, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Li, J.; Han, Y.; Lin, J.; Niu, C.; Zhou, Z.; Yuan, B.; Huang, K.; Li, J.; Jiang, K.; et al. PES1 promotes breast cancer by differentially regulating ERα and ERβ. J. Clin. Invest. 2012, 122, 2857–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Zhang, Y.; Chen, X.; Wang, Y.; Wu, P.; Wu, C.; Chen, D. microRNA-1271 impedes the development of prostate cancer by downregulating PES1 and upregulating ERβ. J. Transl. Med. 2020, 18, 209. [Google Scholar] [CrossRef]
- Shen, J.; Cowen, L.E.; Griffin, A.M.; Chan, L.; Köhler, J.R. The Candida albicans pescadillo homolog is required for normal hypha-to-yeast morphogenesis and yeast proliferation. Proc. Natl. Acad. Sci. USA 2008, 105, 20918–20923. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Musiyenko, A.; Cioffi, E.; Oldenburg, A.; Adams, B.; Bitko, V.; Krishna, S.S.; Barik, S. A zinc-binding dual-specificity YVH1 phosphatase in the malaria parasite, Plasmodium falciparum, and its interaction with the nuclear protein, pescadillo. Mol. Biochem. Parasitol. 2004, 133, 297–310. [Google Scholar] [CrossRef]
- Brar, G.A.; Weissman, J.S. Ribosome profiling reveals the what, when, where and how of protein synthesis. Nat. Rev. Mol. Cell Biol. 2015, 16, 651–664. [Google Scholar] [CrossRef] [Green Version]
- Thomson, E.; Ferreira-Cerca, S.; Hurt, E. Eukaryotic ribosome biogenesis at a glance. J. Cell Sci. 2013, 126, 4815–4821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulon, S.; Westman, B.J.; Hutten, S.; Boisvert, F.-M.; Lamond, A.I. The nucleolus under stress. Mol. Cell 2010, 40, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Calvillo, S.; Florencio-Martínez, L.E.; Nepomuceno-Mejía, T. Nucleolar Structure and Function in Trypanosomatid Protozoa. Cells 2019, 8, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacas, A.; Fernández-Rubio, C.; Algarabel, M.; Peña-Guerrero, J.; Larrea, E.; Rocha Formiga, F.; García-Sosa, A.T.; Nguewa, P.A. The Novel Serine/Threonine Protein Kinase LmjF.22.0810 from Leishmania major may be Involved in the Resistance to Drugs such as Paromomycin. Biomolecules 2019, 9, 723. [Google Scholar] [CrossRef] [Green Version]
- Sádlová, J.; Price, H.P.; Smith, B.A.; Votỳpka, J.; Volf, P.; Smith, D.F. The stage-regulated HASPB and SHERP proteins are essential for differentiation of the protozoan parasite Leishmania major in its sand fly vector, Phlebotomus papatasi. Cell. Microbiol. 2010, 12, 1765–1779. [Google Scholar] [CrossRef] [Green Version]
- Elias, M.C.; Marques-Porto, R.; Freymüller, E.; Schenkman, S. Transcription rate modulation through the Trypanosoma cruzi life cycle occurs in parallel with changes in nuclear organisation. Mol. Biochem. Parasitol. 2001, 112, 79–90. [Google Scholar] [CrossRef]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The genome of the African trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, N.M.; Myler, P.J.; Bartholomeu, D.C.; Nilsson, D.; Aggarwal, G.; Tran, A.N.; Ghedin, E.; Worthey, E.A.; Delcher, A.L.; Blandin, G.; et al. The genome sequence of Trypanosoma cruzi, etiologic agent of chagas disease. Science 2005, 309, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auende, M.L.; Amsterdam, A.; Becker, T.; Kawakami, K.; Gaiano, N. Insertional mutagenesis in zebrafish identifies two novel genes, pescadillo and dead eye, essential for embryonic development. Genes Dev. 1996, 10, 3141–3155. [Google Scholar] [CrossRef] [Green Version]
- Oeffinger, M.; Leung, A.; Lamond, A.; Tollervey, D.; Lueng, A. Yeast Pescadillo is required for multiple activities during 60S ribosomal subunit synthesis. RNA 2002, 8, 626–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uppuluri, P.; Chaturvedi, A.K.; Jani, N.; Pukkila-Worley, R.; Monteagudo, C.; Mylonakis, E.; Köhler, J.R.; Lopez Ribota, J.L. Physiologic expression of the Candida albicans pescadillo homolog is required for virulence in a murine model of hematogenously disseminated candidiasis. Eukaryot. Cell 2012, 11, 1552–1556. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Gustafsson, J.-Å. Targeting PES1 for restoring the ERα/ERβ ratio in breast cancer. J. Clin. Invest. 2012, 122, 2771–2773. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, Y.; Jarell, A.D.; Flaman, J.M.; Foltz, G.; Schuster, J.; Sopher, B.L.; Irvin, D.K.; Kanning, K.; Kornblum, H.I.; Nelson, P.S.; et al. Pescadillo, a novel cell cycle regulatory protein abnormally expressed in malignant cells. J. Biol. Chem. 2001, 276, 6656–6665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, J.; Boger, S.; Li, J.; Duncan, S. a The murine Pes1 gene encodes a nuclear protein containing a BRCT domain. Genomics 2000, 70, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Lapik, Y.R.; Fernandes, C.J.; Lau, L.F.; Pestov, D.G. Physical and functional interaction between Pes1 and Bop1 in mammalian ribosome biogenesis. Mol. Cell 2004, 15, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Sahasranaman, A.; Jakovljevic, J.; Schleifman, E.; Woolford, J.L., Jr. Interactions among Ytm1, Erb1, and Nop7 required for assembly of the Nop7-subcomplex in yeast preribosomes. Mol. Biol. Cell 2008, 19, 2844–2856. [Google Scholar] [CrossRef]
- Grimm, T.; Hölzel, M.; Rohrmoser, M.; Harasim, T.; Malamoussi, A.; Gruber-Eber, A.; Kremmer, E.; Eick, D. Dominant-negative Pes1 mutants inhibit ribosomal RNA processing and cell proliferation via incorporation into the PeBoW-complex. Nucleic Acids Res. 2006, 34, 3030–3043. [Google Scholar] [CrossRef] [Green Version]
- Callebaut, I.; Mornon, J.P. From BRCA1 to RAP1: A widespread BRCT module closely associated with DNA repair. FEBS Lett. 1997, 400, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Coquelle, N.; Green, R.; Glover, J.N.M. Impact of BRCA1 BRCT domain missense substitutions on phosphopeptide recognition. Biochemistry 2011, 50, 4579–4589. [Google Scholar] [CrossRef]
- Sun, L.; Huang, Y.; Edwards, R.A.; Yang, S.; Blackford, A.N.; Niedzwiedz, W.; Glover, J.N.M. Structural Insight into BLM Recognition by TopBP1. Structure 2017, 25, 1582–1588.e3. [Google Scholar] [CrossRef] [Green Version]
- Pessetto, Z.Y.; Yan, Y.; Bessho, T.; Natarajan, A. Inhibition of BRCT(BRCA1)-phosphoprotein interaction enhances the cytotoxic effect of olaparib in breast cancer cells: A proof of concept study for synthetic lethal therapeutic option. Breast Cancer Res. Treat. 2012, 134, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Kappe, S.H.; Noe, A.R.; Fraser, T.S.; Blair, P.L.; Adams, J.H. A family of chimeric erythrocyte binding proteins of malaria parasites. Proc. Natl. Acad. Sci. USA 1998, 95, 1230–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavsar, R.B.; Makley, L.N.; Tsonis, P.A. The other lives of ribosomal proteins. Hum. Genomics 2010, 4, 327–344. [Google Scholar] [CrossRef]
- Reithinger, R.; Dujardin, J.-C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous leishmaniasis. Lancet. Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Sacks, D.; Noben-Trauth, N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol. 2002, 2, 845–858. [Google Scholar] [CrossRef]
- Vacas, A.; Fernández-Rubio, C.; Larrea, E.; Peña-Guerrero, J.; Nguewa, P.A. LmjF.22.0810 from Leishmania major Modulates the Th2-Type Immune Response and Is Involved in Leishmaniasis Outcome. Biomedicines 2020, 8, 452. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.G.; Souza-Testasicca, M.C.; Vago, J.P.; Figueiredo, A.B.; Canavaci, A.M.C.; Perucci, L.O.; Ferreira, T.P.T.; Coelho, E.A.F.; Gonçalves, D.U.; Rocha, M.O.C.; et al. Annexin A1 Is Involved in the Resolution of Inflammatory Responses during Leishmania braziliensis Infection. J. Immunol. 2017, 198, 3227–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, F.J.S.; Schleicher, U.; Mattner, J.; Alber, G.; Bogdan, C. Cytokines, signaling pathways, and effector molecules required for the control of Leishmania (Viannia) braziliensis in mice. Infect. Immun. 2007, 75, 3823–3832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cangussú, S.D.; de Souza, C.C.; Campos, C.F.; Vieira, L.Q.; Afonso, L.C.C.; Arantes, R.M.E. Histopathology of Leishmania major infection: Revisiting L. major histopathology in the ear dermis infection model. Mem. Inst. Oswaldo Cruz 2009, 104, 918–922. [Google Scholar] [CrossRef]
- De Trez, C.; Magez, S.; Akira, S.; Ryffel, B.; Carlier, Y.; Muraille, E. iNOS-producing inflammatory dendritic cells constitute the major infected cell type during the chronic Leishmania major infection phase of C57BL/6 resistant mice. PLoS Pathog. 2009, 5, e1000494. [Google Scholar] [CrossRef]
- Castro Gomes, C.M.; Sousa, M.G.T.; Menezes, J.P.B.; Batista, M.C.; Lima, A.C.S.; Belda, W.J.; Bradshaw, D.; Gama, M.E.A.; Laurenti, M.D.; Silveira, F.T.; et al. Immunopathological characterization of human cutaneous leishmaniasis lesions caused by Leishmania (Viannia) spp. in Amazonian Brazil. Parasitol. Res. 2017, 116, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Stenger, S.; Donhauser, N.; Thüring, H.; Röllinghoff, M.; Bogdan, C. Reactivation of latent leishmaniasis by inhibition of inducible nitric oxide synthase. J. Exp. Med. 1996, 183, 1501–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olekhnovitch, R.; Ryffel, B.; Müller, A.J.; Bousso, P. Collective nitric oxide production provides tissue-wide immunity during Leishmania infection. J. Clin. Invest. 2014, 124, 1711–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Requena, J.M.; Alonso, C.; Soto, M. Evolutionarily conserved proteins as prominent immunogens during Leishmania infections. Parasitol. Today 2000, 16, 246–250. [Google Scholar] [CrossRef]
- Cordeiro-Da-Silva, A.; Borges, M.C.; Guilvard, E.; Ouaissi, A. Dual role of the Leishmania major ribosomal protein S3a homologue in regulation of T- and B-cell activation. Infect. Immun. 2001, 69, 6588–6596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iborra, S.; Parody, N.; Abánades, D.R.; Bonay, P.; Prates, D.; Novais, F.O.; Barral-Netto, M.; Alonso, C.; Soto, M. Vaccination with the Leishmania major ribosomal proteins plus CpG oligodeoxynucleotides induces protection against experimental cutaneous leishmaniasis in mice. Microbes Infect. 2008, 10, 1133–1141. [Google Scholar] [CrossRef]
- Ramírez, L.; Iborra, S.; Cortés, J.; Bonay, P.; Alonso, C.; Barral-Netto, M.; Soto, M. BALB/c mice vaccinated with Leishmania major ribosomal proteins extracts combined with CpG oligodeoxynucleotides become resistant to disease caused by a secondary parasite challenge. J. Biomed. Biotechnol. 2010, 2010, 181690. [Google Scholar] [CrossRef] [PubMed]
- Green, S.J.; Crawford, R.M.; Hockmeyer, J.T.; Meltzer, M.S.; Nacy, C.A. Leishmania major amastigotes initiate the L-arginine-dependent killing mechanism in IFN-gamma-stimulated macrophages by induction of tumor necrosis factor-alpha. J. Immunol. 1990, 145, 4290–4297. [Google Scholar]
- Stenger, S.; Thüring, H.; Röllinghoff, M.; Bogdan, C. Tissue expression of inducible nitric oxide synthase is closely associated with resistance to Leishmania major. J. Exp. Med. 1994, 180, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yu, L.; Zhang, H.; Wu, J.; Yuan, J.; Li, X.; Li, M. Down-regulation of pescadillo inhibits proliferation and tumorigenicity of breast cancer cells. Cancer Sci. 2009, 100, 2255–2260. [Google Scholar] [CrossRef]
- James, A.; Wang, Y.; Raje, H.; Rosby, R.; DiMario, P. Nucleolar stress with and without p53. Nucleus 2014, 5, 402–426. [Google Scholar] [CrossRef] [Green Version]
- Názer, E.; Sánchez, D.O. Nucleolar accumulation of RNA binding proteins induced by Actinomycin D is functional in Trypanosoma cruzi and Leishmania mexicana but not in T. brucei. PLoS ONE 2011, 6, e24184. [Google Scholar] [CrossRef]
- Názer, E.; Verdún, R.E.; Sánchez, D.O. Nucleolar localization of RNA binding proteins induced by actinomycin D and heat shock in Trypanosoma cruzi. PLoS ONE 2011, 6, e19920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslett, M.; Aurrecoechea, C.; Berriman, M.; Brestelli, J.; Brunk, B.P.; Carrington, M.; Depledge, D.P.; Fischer, S.; Gajria, B.; Gao, X.; et al. TriTrypDB: A functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 2010, 38, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Schlessinger, A.; Punta, M.; Yachdav, G.; Kajan, L.; Rost, B. Improved disorder prediction by combination of orthogonal approaches. PLoS ONE 2009, 4, e4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yachdav, G.; Kloppmann, E.; Kajan, L.; Hecht, M.; Goldberg, T.; Hamp, T.; Hönigschmid, P.; Schafferhans, A.; Roos, M.; Bernhofer, M.; et al. PredictProtein--an open resource for online prediction of protein structural and functional features. Nucleic Acids Res. 2014, 42, W337–W343. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.-Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. In Proceedings of the Pacific Symposium on Biocomputing, Kauai, HI, USA, 3–7 January 2002; pp. 310–322. [Google Scholar]
- The MYC Predictor. Available online: https://mendel.imp.ac.at/myristate/SUPLpredictor.htm (accessed on 23 September 2021).
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices From Protein Sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Fernández-Rubio, C.; Campbell, D.; Vacas, A.; Ibañez, E.; Moreno, E.; Espuelas, S.; Calvo, A.; Palop, J.A.; Plano, D.; Sanmartin, C.; et al. Leishmanicidal activities of novel methylseleno-imidocarbamates. Antimicrob. Agents Chemother. 2015, 59, 5705–5713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Rubio, C.; Larrea, E.; Peña Guerrero, J.; Sesma Herrero, E.; Gamboa, I.; Berrio, C.; Plano, D.; Amin, S.; Sharma, A.K.; Nguewa, P.A. Leishmanicidal Activity of Isoselenocyanate Derivatives. Antimicrob. Agents Chemother. 2019, 63, e00904-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacas, A.; Sugden, C.; Velasco-Rodriguez, O.; Algarabel-Olona, M.; Pena-Guerrero, J.; Larrea, E.; Fernandez-Rubio, C.; Nguewa, P.A. Construction of Two mCherry Plasmids (pXG-mCherry) for Transgenic Leishmania: Valuable Tools for Future Molecular Analysis. J. Parasitol. Res. 2017, 2017, 1964531. [Google Scholar] [CrossRef] [Green Version]
- Cruz, A.; Coburn, C.M.; Beverley, S.M. Double targeted gene replacement for creating null mutants. Proc. Natl. Acad. Sci. USA 1991, 88, 7170–7174. [Google Scholar] [CrossRef] [Green Version]
- Sacks, D.L.; Peter, V. Perkins Identification of an Infective Stage of Leishmania Promastigotes. Science 2011, 223, 1417–1419. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward (Fw) Primer (5′→3′) | Reverse (Rv) Primer (5′→3′) |
|---|---|---|
| LmjPESmCherry | GCGGCCGCTATTATGGTCACATAAGAAGCAG | GCGGCCGCATTACTGCACCCACTT |
| LmjPESpXG | GCCGGATCCCCACCAATGGTCCATAAGAAGCAGGCA | GCCGGATCCTTACTGCACCCACTTGGGCAG |
| LmjPESpLEXSY | CCATGGGAAATGGTCCATAAGAAGCAGGCA | GGT ACC CTG CAC CCA CTT GGG CAG TTT |
| pLEXSY-INT | CCGACTGCAACA AGGTGTAG | CAT CTA TAG AGA AGT ACA CGT AAA AG |
| qLmjPES | GAGATGGACATGGAGGACGA | TCATCTCGCGCTG |
| qGAPDH | CATCAAGTGCGTGAAGGCGC | CGTCGGCGAGTACTCGTGCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algarabel, M.; Fernández-Rubio, C.; Musilova, K.; Peña-Guerrero, J.; Vacas, A.; Larrea, E.; Nguewa, P.A. In Leishmania major, the Homolog of the Oncogene PES1 May Play a Critical Role in Parasite Infectivity. Int. J. Mol. Sci. 2021, 22, 12592. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212592
Algarabel M, Fernández-Rubio C, Musilova K, Peña-Guerrero J, Vacas A, Larrea E, Nguewa PA. In Leishmania major, the Homolog of the Oncogene PES1 May Play a Critical Role in Parasite Infectivity. International Journal of Molecular Sciences. 2021; 22(22):12592. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212592
Chicago/Turabian StyleAlgarabel, Miriam, Celia Fernández-Rubio, Katerina Musilova, José Peña-Guerrero, Andrés Vacas, Esther Larrea, and Paul A. Nguewa. 2021. "In Leishmania major, the Homolog of the Oncogene PES1 May Play a Critical Role in Parasite Infectivity" International Journal of Molecular Sciences 22, no. 22: 12592. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212592