
Substrate-Dependent Trans-Stimulation of Organic Cation Transporter 2 Activity

 , ,
, ,
Abstract
:1. Introduction
2. Results
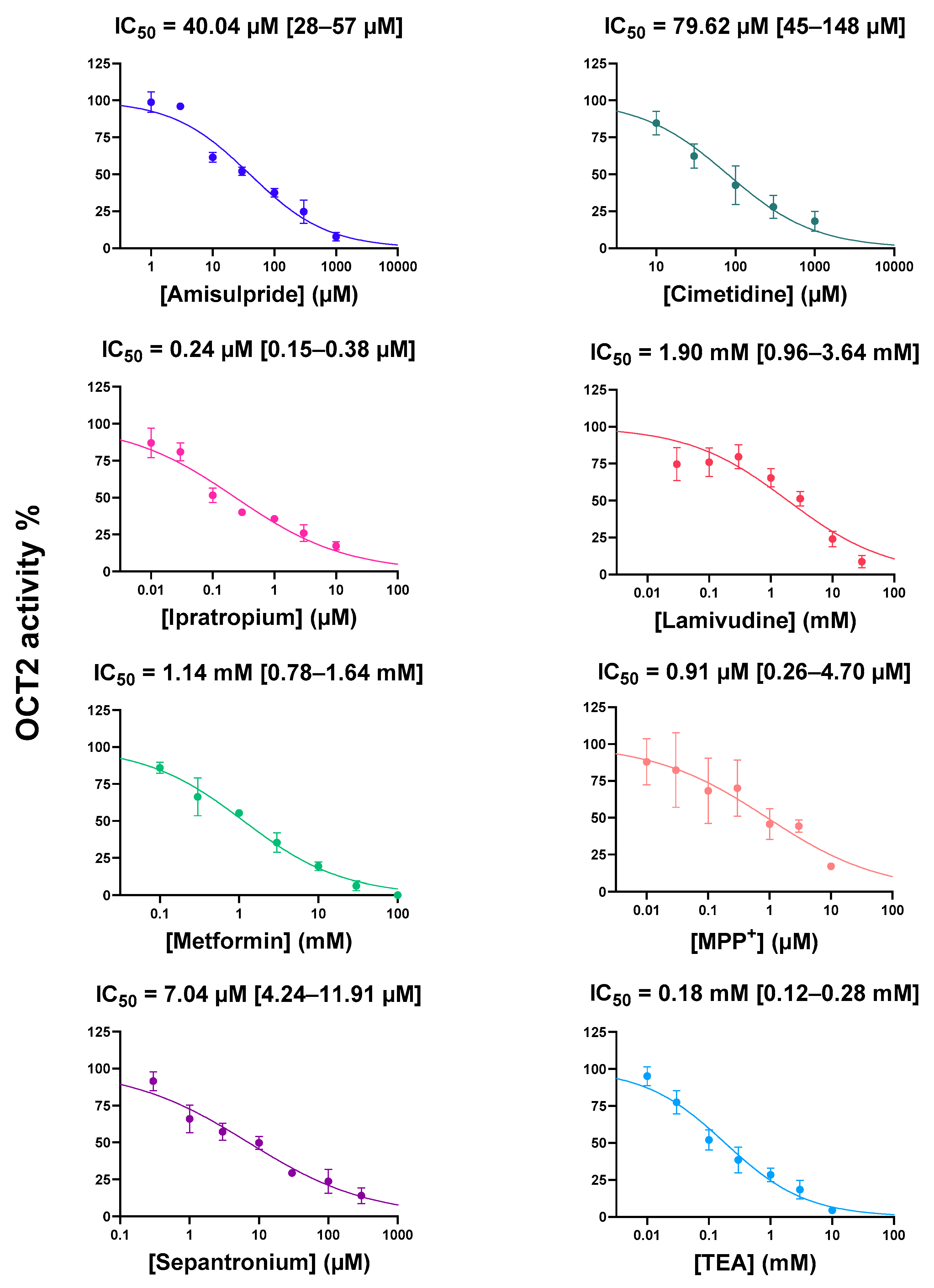
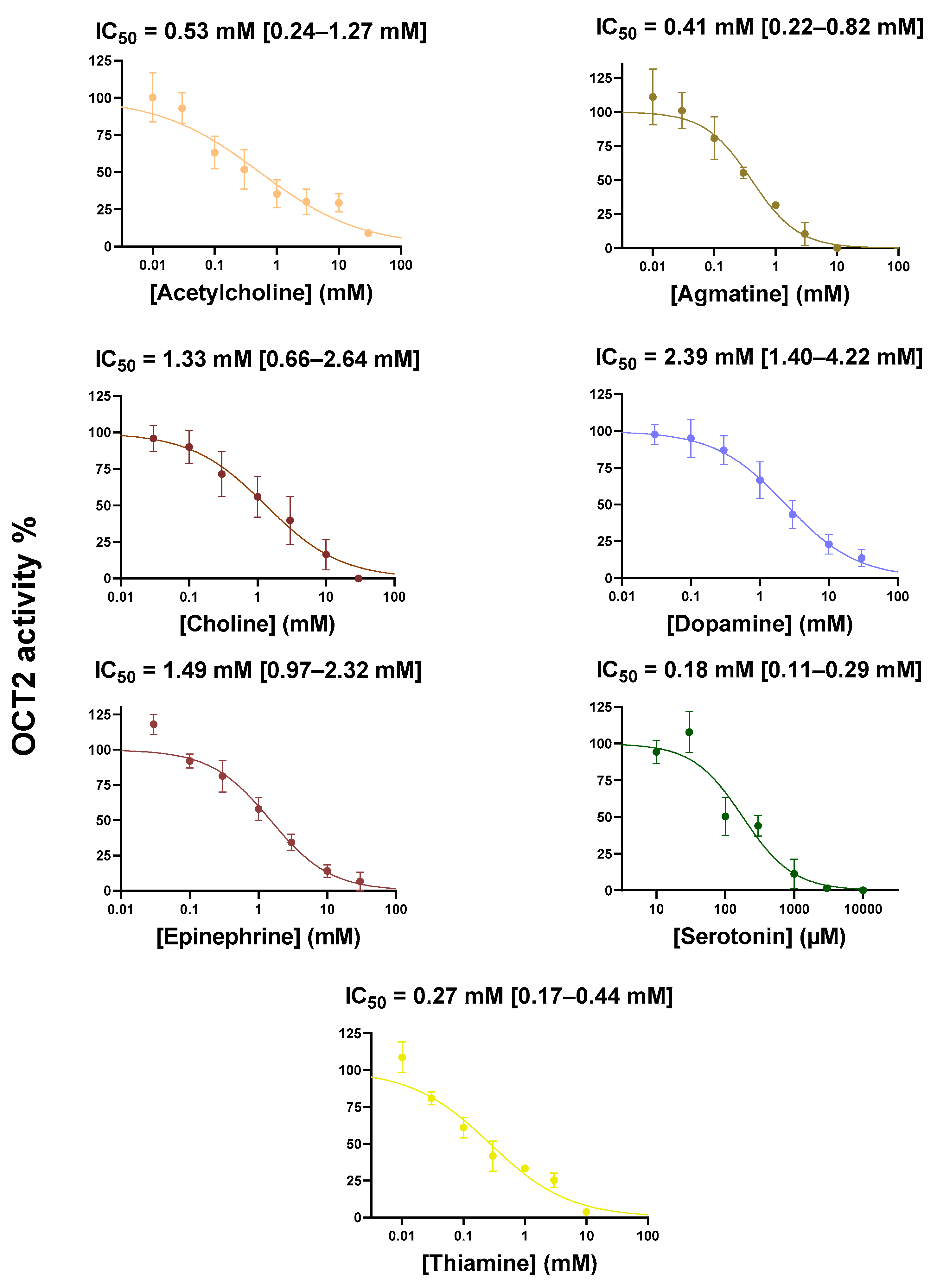
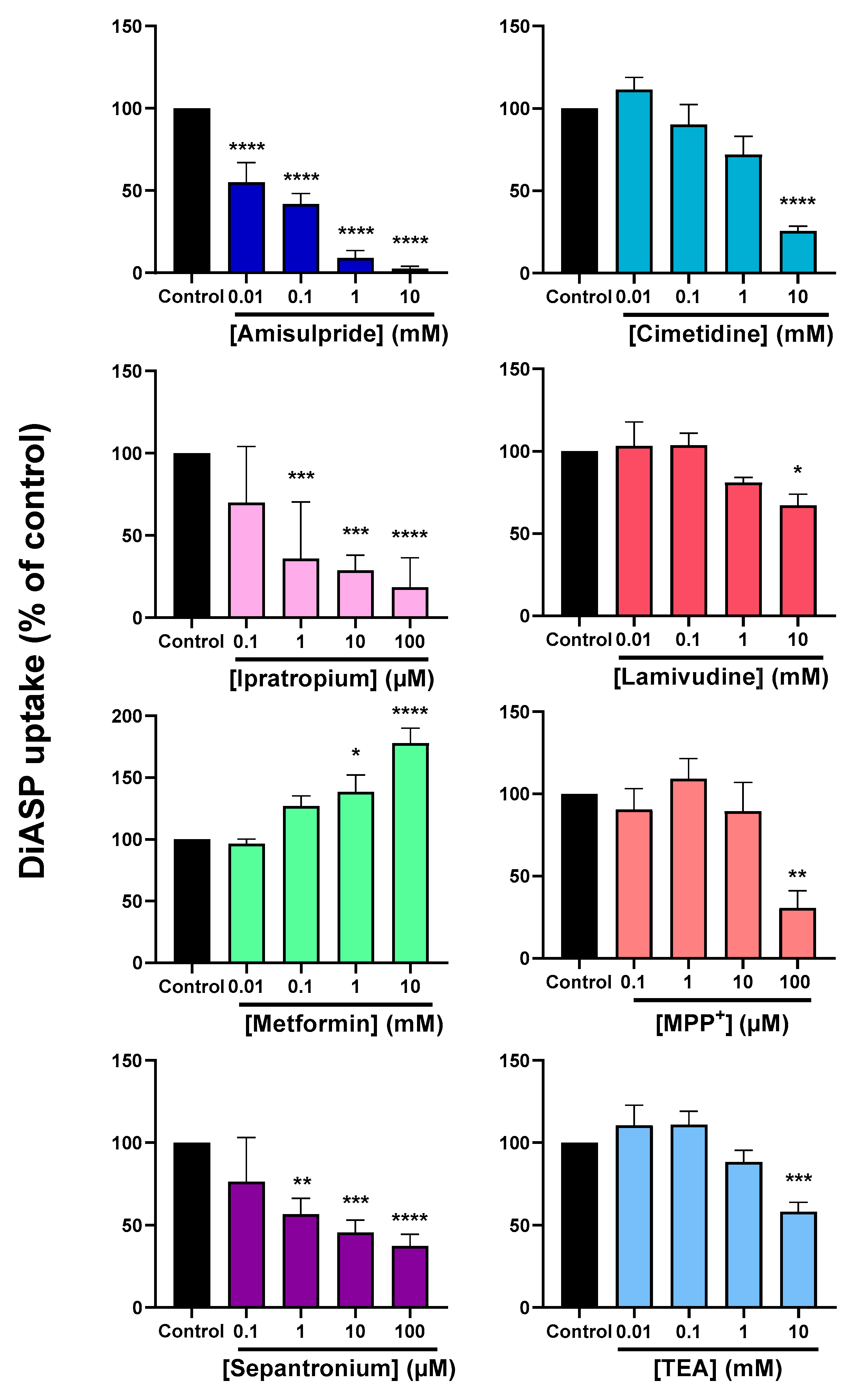
2.1. Cis-Inhibition of OCT2 Activity by OCT2 Substrates
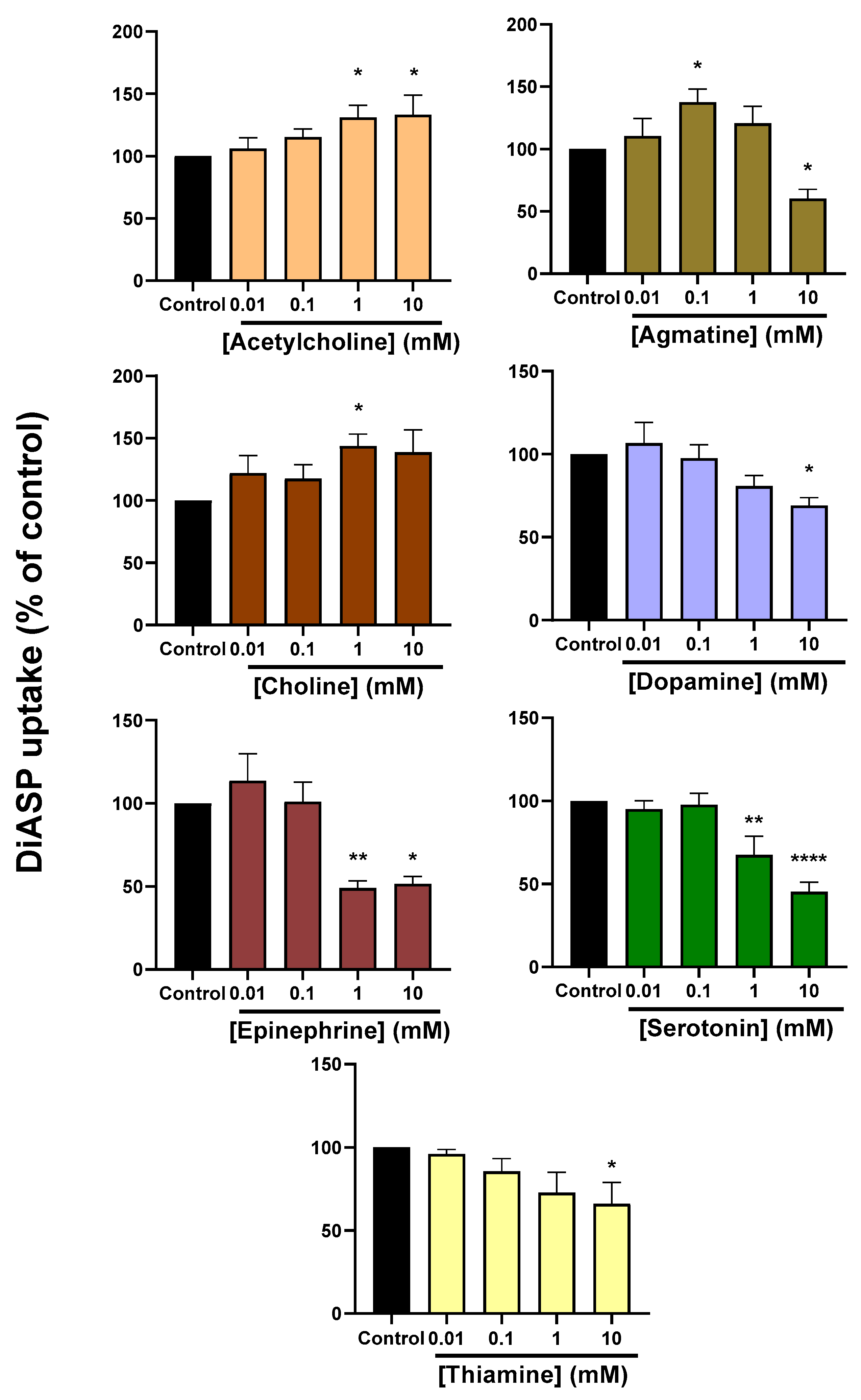
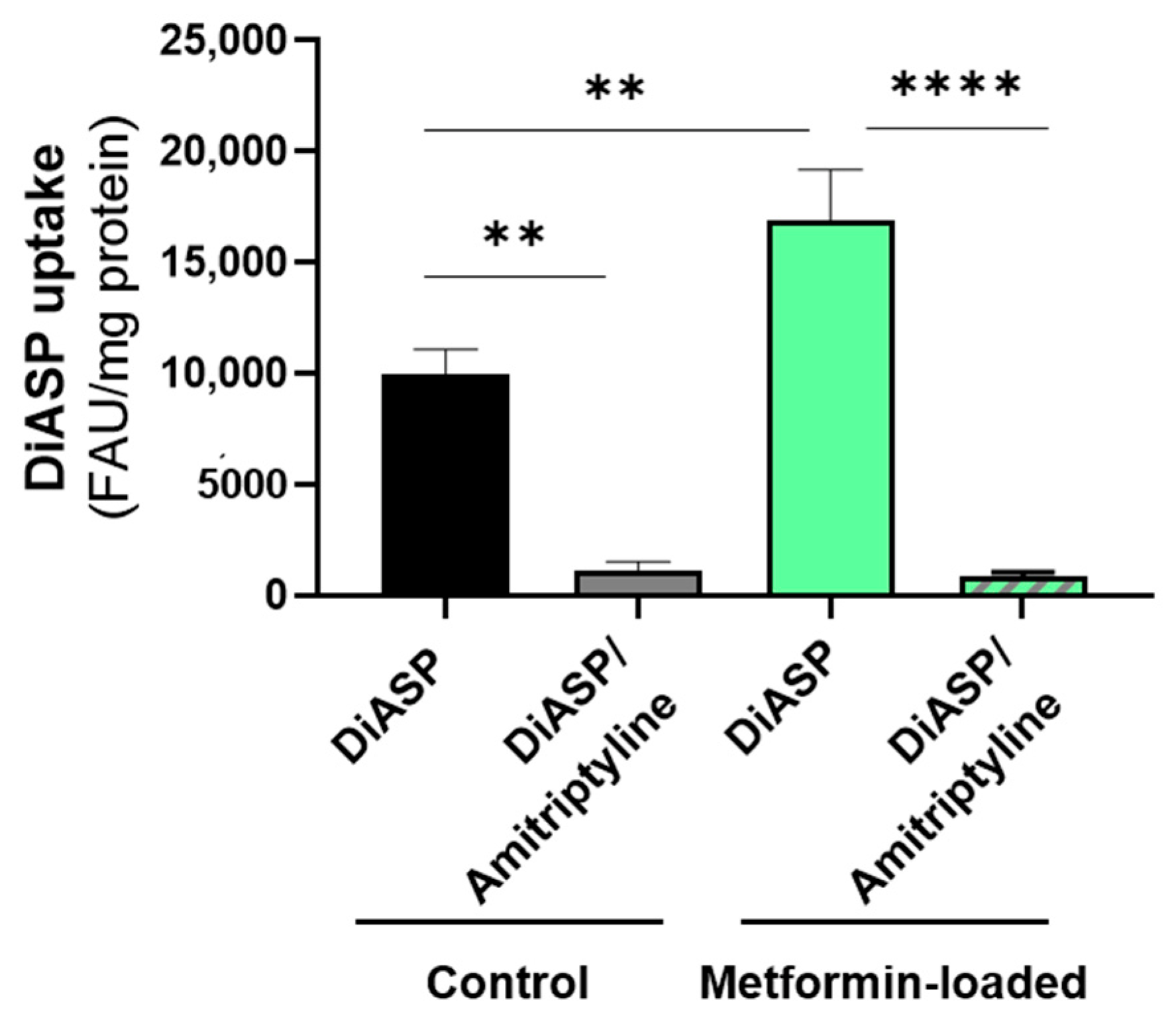
2.2. Trans-Stimulation of OCT2 Activity by OCT2 Substrates
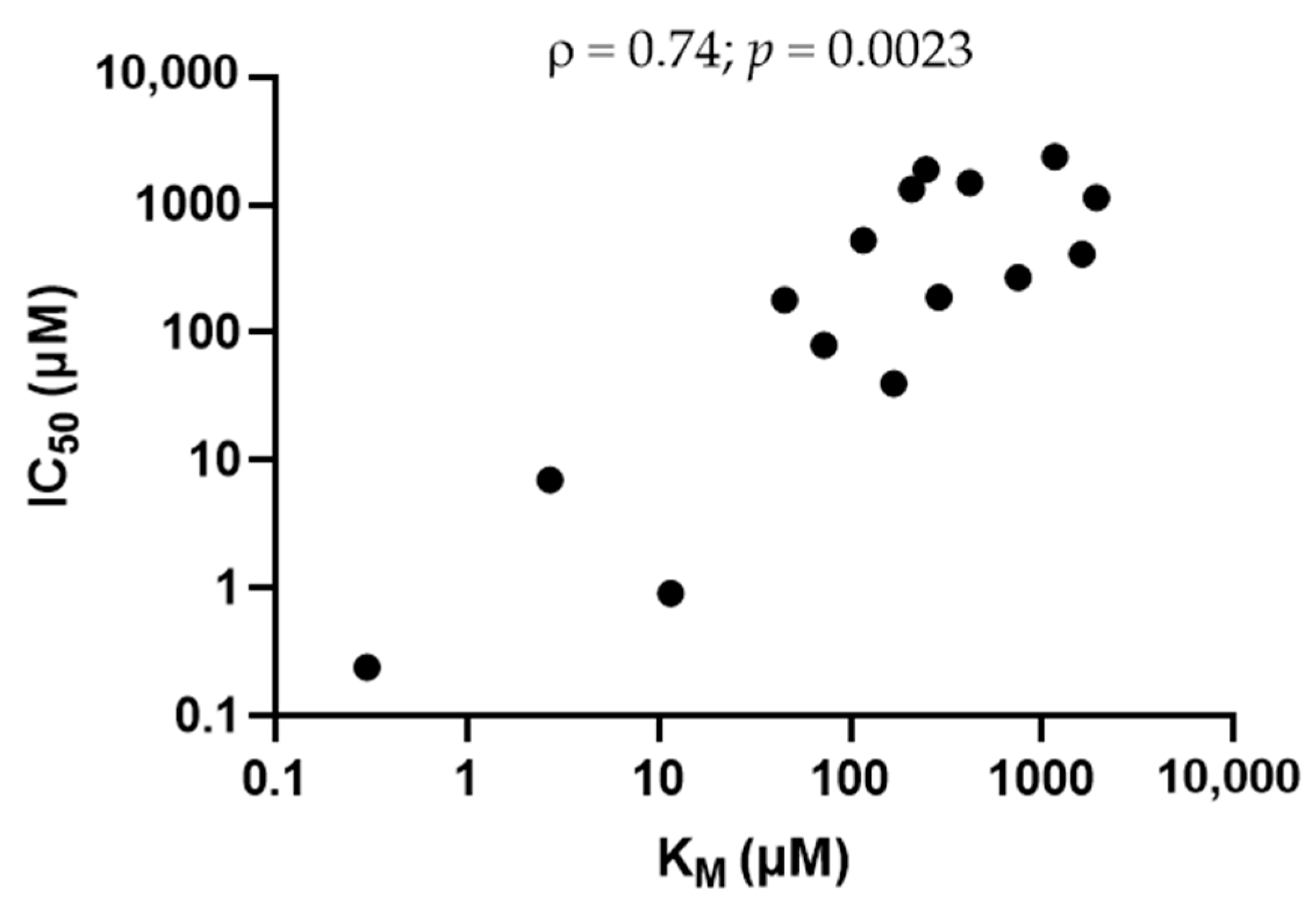
2.3. Physico-Chemical Parameters Associated with Trans-Stimulation of OCT2 Activity by OCT2 Substrates
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. OCT2 Activity Assays
4.3.1. Cis-Inhibition Assays
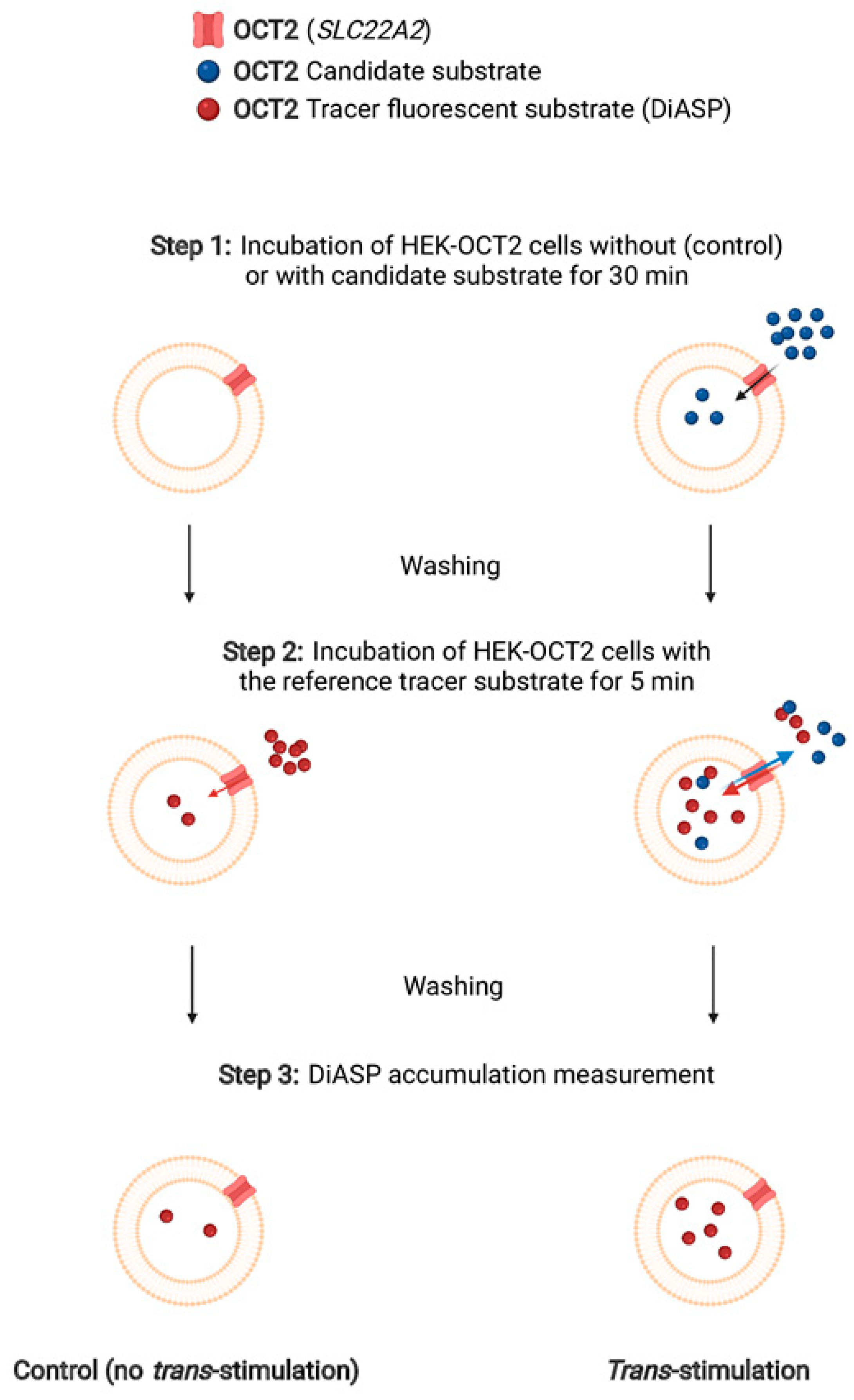
4.3.2. Trans-Stimulation Assays
4.4. Molecular Descriptors Generation
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pizzagalli, M.D.; Bensimon, A.; Superti-Furga, G. A guide to plasma membrane solute carrier proteins. FEBS J. 2021, 288, 2784–2835. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, K.M.; Huang, S.M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M.; et al. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef]
- Brecht, K.; Schäfer, A.M.; Meyer Zu Schwabedissen, H.E. Uptake Transporters of the SLC21, SLC22A, and SLC15A Families in Anticancer Therapy-Modulators of Cellular Entry or Pharmacokinetics? Cancers 2020, 12, 2263. [Google Scholar] [CrossRef] [PubMed]
- Nies, A.T.; Koepsell, H.; Damme, K.; Schwab, M. Organic cation transporters (OCTs, MATEs), in vitro and in vivo evidence for the importance in drug therapy. Handb. Exp. Pharmacol. 2011, 201, 105–167. [Google Scholar] [CrossRef]
- Lee, S.C.; Arya, V.; Yang, X.; Volpe, D.A.; Zhang, L. Evaluation of transporters in drug development: Current status and contemporary issues. Adv. Drug Deliv. Rev. 2017, 116, 100–118. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, D.; Schömig, E. Gene structures of the human non-neuronal monoamine transporters EMT and OCT2. Hum. Genet. 2000, 106, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.H. Molecular and cellular physiology of organic cation transporter 2. Am. J. Physiol. Ren. Physiol. 2019, 317, F1669–F1679. [Google Scholar] [CrossRef] [PubMed]
- Budiman, T.; Bamberg, E.; Koepsell, H.; Nagel, G. Mechanism of electrogenic cation transport by the cloned organic cation transporter 2 from rat. J. Biol. Chem. 2000, 275, 29413–29420. [Google Scholar] [CrossRef] [Green Version]
- Koepsell, H. Multiple binding sites in organic cation transporters require sophisticated procedures to identify interactions of novel drugs. Biol. Chem. 2019, 400, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Samodelov, S.L.; Kullak-Ublick, G.A.; Gai, Z.; Visentin, M. Organic Cation Transporters in Human Physiology, Pharmacology, and Toxicology. Int. J. Mol. Sci. 2020, 21, 7890. [Google Scholar] [CrossRef]
- Motohashi, H.; Inui, K. Multidrug and toxin extrusion family SLC47: Physiological, pharmacokinetic and toxicokinetic importance of MATE1 and MATE2-K. Mol. Asp. Med. 2013, 34, 661–668. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Hucke, A.; Rinschen, M.M.; Bauer, O.B.; Sperling, M.; Karst, U.; Köppen, C.; Sommer, K.; Schröter, R.; Ceresa, C.; Chiorazzi, A.; et al. An integrative approach to cisplatin chronic toxicities in mice reveals importance of organic cation-transporter-dependent protein networks for renoprotection. Arch. Toxicol. 2019, 93, 2835–2848. [Google Scholar] [CrossRef]
- Burckhardt, G. Drug transport by Organic Anion Transporters (OATs). Pharmacol. Ther. 2012, 136, 106–130. [Google Scholar] [CrossRef]
- Dvorak, V.; Wiedmer, T.; Ingles-Prieto, A.; Altermatt, P.; Batoulis, H.; Bärenz, F.; Bender, E.; Digles, D.; Dürrenberger, F.; Heitman, L.H.; et al. An Overview of Cell-Based Assay Platforms for the Solute Carrier Family of Transporters. Front. Pharmacol. 2021, 12, 722889. [Google Scholar] [CrossRef]
- Gründemann, D.; Hahne, C.; Berkels, R.; Schömig, E. Agmatine is efficiently transported by non-neuronal monoamine transporters extraneuronal monoamine transporter (EMT) and organic cation transporter 2 (OCT2). J. Pharmacol. Exp. Ther. 2003, 304, 810–817. [Google Scholar] [CrossRef] [Green Version]
- Dresser, M.J.; Xiao, G.; Leabman, M.K.; Gray, A.T.; Giacomini, K.M. Interactions of n-tetraalkylammonium compounds and biguanides with a human renal organic cation transporter (hOCT2). Pharm. Res. 2002, 19, 1244–1247. [Google Scholar] [CrossRef]
- Hagos, Y.; Schley, G.; Schödel, J.; Krick, W.; Burckhardt, G.; Willam, C.; Burckhardt, B.C. α-Ketoglutarate-related inhibitors of HIF prolyl hydroxylases are substrates of renal organic anion transporters 1 (OAT1) and 4 (OAT4). Pflugers Arch. 2012, 464, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Blazquez, A.G.; Briz, O.; Romero, M.R.; Rosales, R.; Monte, M.J.; Vaquero, J.; Macias, R.I.; Cassio, D.; Marin, J.J. Characterization of the role of ABCG2 as a bile acid transporter in liver and placenta. Mol. Pharmacol. 2012, 81, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Severance, A.C.; Sandoval, P.J.; Wright, S.H. Correlation between Apparent Substrate Affinity and OCT2 Transport Turnover. J. Pharmacol. Exp. Ther. 2017, 362, 405–412. [Google Scholar] [CrossRef]
- Bourdet, D.L.; Pritchard, J.B.; Thakker, D.R. Differential substrate and inhibitory activities of ranitidine and famotidine toward human organic cation transporter 1 (hOCT1; SLC22A1), hOCT2 (SLC22A2), and hOCT3 (SLC22A3). J. Pharmacol. Exp. Ther. 2005, 315, 1288–1297. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Guerrero, L.J.; Wright, S.H. Substrate-dependent inhibition of human MATE1 by cationic ionic liquids. J. Pharmacol. Exp. Ther. 2013, 346, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.N.; Wright, S.H. Multiple mechanisms of ligand interaction with the human organic cation transporter, OCT2. Am. J. Physiol. Ren. Physiol. 2013, 304, F56–F67. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, A.M.; Bock, T.; Meyer Zu Schwabedissen, H.E. Establishment and Validation of Competitive Counterflow as a Method To Detect Substrates of the Organic Anion Transporting Polypeptide 2B1. Mol. Pharm. 2018, 15, 5501–5513. [Google Scholar] [CrossRef]
- McKinney, T.D.; Hosford, M.A. Organic cation transport by rat hepatocyte basolateral membrane vesicles. Am. J. Physiol. 1992, 263 Pt 1, G939–G946. [Google Scholar] [CrossRef]
- Takahashi, K.; Nakamura, N.; Terada, T.; Okano, T.; Futami, T.; Saito, H.; Inui, K.I. Interaction of beta-lactam antibiotics with H+/peptide cotransporters in rat renal brush-border membranes. J. Pharmacol. Exp. Ther. 1998, 286, 1037–1042. [Google Scholar]
- Takano, M.; Inui, K.; Okano, T.; Saito, H.; Hori, R. Carrier-mediated transport systems of tetraethylammonium in rat renal brush-border and basolateral membrane vesicles. Biochim. et Biophys. Acta 1984, 773, 113–124. [Google Scholar] [CrossRef]
- Cisternino, S.; Chapy, H.; André, P.; Smirnova, M.; Debray, M.; Scherrmann, J.M. Coexistence of passive and proton antiporter-mediated processes in nicotine transport at the mouse blood-brain barrier. AAPS J. 2013, 15, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Chapy, H.; André, P.; Declèves, X.; Scherrmann, J.M.; Cisternino, S. A polyspecific drug/proton antiporter mediates diphenhydramine and clonidine transport at the mouse blood-retinal barrier. Br. J. Pharmacol. 2015, 172, 4714–4725. [Google Scholar] [CrossRef]
- Dresser, M.J.; Gray, A.T.; Giacomini, K.M. Kinetic and selectivity differences between rodent, rabbit, and human organic cation transporters (OCT1). J. Pharmacol. Exp. Ther. 2000, 292, 1146–1152. [Google Scholar]
- Apiwattanakul, N.; Sekine, T.; Chairoungdua, A.; Kanai, Y.; Nakajima, N.; Sophasan, S.; Endou, H. Transport properties of nonsteroidal anti-inflammatory drugs by organic anion transporter 1 expressed in Xenopus laevis oocytes. Mol. Pharmacol. 1999, 55, 847–854. [Google Scholar] [PubMed]
- Müller, J.P.; Keufgens, L.; Gründemann, D. Hyperosmolarity stimulates transporter-mediated insertion of estrone sulfate into the plasma membrane, but inhibits the uptake by SLC10A1 (NTCP). Biochem. Pharmacol. 2021, 186, 114484. [Google Scholar] [CrossRef]
- Chapy, H.; Goracci, L.; Vayer, P.; Parmentier, Y.; Carrupt, P.A.; Declèves, X.; Scherrmann, J.M.; Cisternino, S.; Cruciani, G. Pharmacophore-based discovery of inhibitors of a novel drug/proton antiporter in human brain endothelial hCMEC/D3 cell line. Br. J. Pharmacol. 2015, 172, 4888–4904. [Google Scholar] [CrossRef] [PubMed]
- Lips, K.S.; Volk, C.; Schmitt, B.M.; Pfeil, U.; Arndt, P.; Miska, D.; Ermert, L.; Kummer, W.; Koepsell, H. Polyspecific cation transporters mediate luminal release of acetylcholine from bronchial epithelium. Am. J. Respir. Cell Mol. Biol. 2005, 33, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, T.N.; Elmquist, W.F.; Fairbanks, C.A. OCT2 and MATE1 provide bidirectional agmatine transport. Mol. Pharm. 2011, 8, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Pereira, J.N.; Tadjerpisheh, S.; Abu Abed, M.; Saadatmand, A.R.; Weksler, B.; Romero, I.A.; Couraud, P.O.; Brockmöller, J.; Tzvetkov, M.V. The poorly membrane permeable antipsychotic drugs amisulpride and sulpiride are substrates of the organic cation transporters from the SLC22 family. AAPS J. 2014, 16, 1247–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorboulev, V.; Ulzheimer, J.C.; Akhoundova, A.; Ulzheimer-Teuber, I.; Karbach, U.; Quester, S.; Baumann, C.; Lang, F.; Busch, A.E.; Koepsell, H. Cloning and characterization of two human polyspecific organic cation transporters. DNA Cell Biol. 1997, 16, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Tahara, H.; Kusuhara, H.; Endou, H.; Koepsell, H.; Imaoka, T.; Fuse, E.; Sugiyama, Y. A species difference in the transport activities of H2 receptor antagonists by rat and human renal organic anion and cation transporters. J. Pharmacol. Exp. Ther. 2005, 315, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Zolk, O.; Solbach, T.F.; König, J.; Fromm, M.F. Functional characterization of the human organic cation transporter 2 variant p.270Ala>Ser. Drug Metab. Dispos. 2009, 37, 1312–1318. [Google Scholar] [CrossRef] [Green Version]
- Amphoux, A.; Vialou, V.; Drescher, E.; Brüss, M.; Mannoury La Cour, C.; Rochat, C.; Millan, M.J.; Giros, B.; Bönisch, H.; Gautron, S. Differential pharmacological in vitro properties of organic cation transporters and regional distribution in rat brain. Neuropharmacology 2006, 50, 941–952. [Google Scholar] [CrossRef]
- Chen, J.; Brockmöller, J.; Seitz, T.; König, J.; Chen, X.; Tzvetkov, M.V. Tropane alkaloids as substrates and inhibitors of human organic cation transporters of the SLC22 (OCT) and the SLC47 (MATE) families. Biol. Chem. 2017, 398, 237–249. [Google Scholar] [CrossRef]
- Jung, N.; Lehmann, C.; Rubbert, A.; Knispel, M.; Hartmann, P.; van Lunzen, J.; Stellbrink, H.J.; Faetkenheuer, G.; Taubert, D. Relevance of the organic cation transporters 1 and 2 for antiretroviral drug therapy in human immunodeficiency virus infection. Drug Metab. Dispos. 2008, 36, 1616–1623. [Google Scholar] [CrossRef]
- Kimura, N.; Masuda, S.; Tanihara, Y.; Ueo, H.; Okuda, M.; Katsura, T.; Inui, K. Metformin is a superior substrate for renal organic cation transporter OCT2 rather than hepatic OCT1. Drug Metab. Pharmacokinet. 2005, 20, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Elsby, R.; Chidlaw, S.; Outteridge, S.; Pickering, S.; Radcliffe, A.; Sullivan, R.; Jones, H.; Butler, P. Mechanistic in vitro studies confirm that inhibition of the renal apical efflux transporter multidrug and toxin extrusion (MATE) 1, and not altered absorption, underlies the increased metformin exposure observed in clinical interactions with cimetidine, trimethoprim or pyrimethamine. Pharmacol. Res. Perspect. 2017, 5, e00357. [Google Scholar] [CrossRef]
- Lee, W.K.; Reichold, M.; Edemir, B.; Ciarimboli, G.; Warth, R.; Koepsell, H.; Thévenod, F. Organic cation transporters OCT1, 2, and 3 mediate high-affinity transport of the mutagenic vital dye ethidium in the kidney proximal tubule. Am. J. Physiol. Ren. Physiol. 2009, 296, F1504–F1513. [Google Scholar] [CrossRef] [Green Version]
- Belzer, M.; Morales, M.; Jagadish, B.; Mash, E.A.; Wright, S.H. Substrate-dependent ligand inhibition of the human organic cation transporter OCT2. J. Pharmacol. Exp. Ther. 2013, 346, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Minematsu, T.; Iwai, M.; Umehara, K.; Usui, T.; Kamimura, H. Characterization of human organic cation transporter 1 (OCT1/SLC22A1)- and OCT2 (SLC22A2)-mediated transport of 1-(2-methoxyethyl)-2-methyl-4,9-dioxo-3-(pyrazin-2-ylmethyl)- 4,9-dihydro-1H-naphtho[2,3-d]imidazolium bromide (YM155 monobromide), a novel small molecule survivin suppressant. Drug Metab. Dispos. 2010, 38, 1–4. [Google Scholar] [CrossRef]
- Barendt, W.M.; Wright, S.H. The human organic cation transporter (hOCT2) recognizes the degree of substrate ionization. J. Biol. Chem. 2002, 277, 22491–22496. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Shu, Y.; Liang, X.; Chen, E.C.; Yee, S.W.; Zur, A.A.; Li, S.; Xu, L.; Keshari, K.R.; Lin, M.J.; et al. OCT1 is a high-capacity thiamine transporter that regulates hepatic steatosis and is a target of metformin. Proc. Natl. Acad. Sci. USA 2014, 111, 9983–9988. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Ikawa, T.; Takeshita, H.; Kanno, S.; Nagai, T.; Takada, M.; Mukai, T.; Wempe, M.F. Human organic cation transporter 2 (hOCT2): Inhibitor studies using S2-hOCT2 cells. Toxicology 2013, 310, 98–103. [Google Scholar] [CrossRef]
- Sugano, K.; Kansy, M.; Artursson, P.; Avdeef, A.; Bendels, S.; Di, L.; Ecker, G.F.; Faller, B.; Fischer, H.; Gerebtzoff, G.; et al. Coexistence of passive and carrier-mediated processes in drug transport. Nat. Rev. Drug Discov. 2010, 9, 597–614. [Google Scholar] [CrossRef]
- Sayyed, K.; Camillerapp, C.; Le Vée, M.; Bruyère, A.; Nies, A.T.; Abdel-Razzak, Z.; Fardel, O. Inhibition of organic cation transporter (OCT) activities by carcinogenic heterocyclic aromatic amines. Toxicol. In Vitro 2019, 54, 10–22. [Google Scholar] [CrossRef]
- Sandoval, P.J.; Zorn, K.M.; Clark, A.M.; Ekins, S.; Wright, S.H. Assessment of Substrate-Dependent Ligand Interactions at the Organic Cation Transporter OCT2 Using Six Model Substrates. Mol. Pharmacol. 2018, 94, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Kimura, N.; Masuda, S.; Katsura, T.; Inui, K. Transport of guanidine compounds by human organic cation transporters, hOCT1 and hOCT2. Biochem. Pharmacol. 2009, 77, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Bakhiya, A.; Bahn, A.; Burckhardt, G.; Wolff, N. Human organic anion transporter 3 (hOAT3) can operate as an exchanger and mediate secretory urate flux. Cell. Physiol. Biochem. 2003, 13, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Cant, J.P. Glucose transporter in bovine mammary epithelial cells is an asymmetric carrier that exhibits cooperativity and trans-stimulation. Am. J. Physiol. Cell Physiol. 2003, 285, C1226–C1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, H.C.; Colas, C.; Finke, K.; Springer, S.; Stoner, L.; Zur, A.A.; Venteicher, B.; Campbell, J.; Hall, C.; Flint, A.; et al. Reevaluating the Substrate Specificity of the L-Type Amino Acid Transporter (LAT1). J. Med. Chem. 2018, 61, 7358–7373. [Google Scholar] [CrossRef] [PubMed]
- Fardel, O.; Le Vee, M.; Jouan, E.; Denizot, C.; Parmentier, Y. Nature and uses of fluorescent dyes for drug transporter studies. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1233–1251. [Google Scholar] [CrossRef]
- Cetinkaya, I.; Ciarimboli, G.; Yalçinkaya, G.; Mehrens, T.; Velic, A.; Hirsch, J.R.; Gorboulev, V.; Koepsell, H.; Schlatter, E. Regulation of human organic cation transporter hOCT2 by PKA, PI3K, and calmodulin-dependent kinases. Am. J. Physiol. Ren. Physiol. 2003, 284, F293–F302. [Google Scholar] [CrossRef] [Green Version]
- Koepp, T.N.; Tokaj, A.; Nedvetsky, P.I.; Conchon Costa, A.C.; Snieder, B.; Schröter, R.; Ciarimboli, G. Properties of Transport Mediated by the Human Organic Cation Transporter 2 Studied in a Polarized Three-Dimensional Epithelial Cell Culture Model. Int. J. Mol. Sci. 2021, 22, 9658. [Google Scholar] [CrossRef]
- Jouan, E.; Le Vee, M.; Denizot, C.; Da Violante, G.; Fardel, O. The mitochondrial fluorescent dye rhodamine 123 is a high-affinity substrate for organic cation transporters (OCTs) 1 and 2. Fundam. Clin. Pharmacol. 2014, 28, 65–77. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chedik, L.; Bruyere, A.; Le Vee, M.; Stieger, B.; Denizot, C.; Parmentier, Y.; Potin, S.; Fardel, O. Inhibition of Human Drug Transporter Activities by the Pyrethroid Pesticides Allethrin and Tetramethrin. PLoS ONE 2017, 12, e0169480. [Google Scholar] [CrossRef] [Green Version]
- de Winter, J.C.F. Using the Student’s t-test with extremely small sample sizes. Pract. Assess. Res. Eval. 2013, 18, 10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OCT2 Substrate | Nature | KM (µM) | Reference |
|---|---|---|---|
| Acetylcholine | Endogenous substance (neurotransmitter) | 117 | [34] |
| Agmatine | Endogenous substance (metabolite) | 1400–1840 | [16,35] |
| Amisulpride | Exogenous substance (antiemetic drug) | 168 | [36] |
| Choline | Endogenous substance (neurotransmitter) | 210 | [37] |
| Cimetidine | Exogenous substance (antihistamine drug) | 73 | [38] |
| Dopamine | Endogenous substance (neurotransmitter) | 932–1400 | [39,40] |
| Epinephrine | Endogenous substance (biogenic amine) | 420 | [40] |
| Ipratropium | Exogenous substance (anticholinergic drug) | 0.3 | [41] |
| Lamivudine | Exogenous substance (antiretroviral drug) | 248 | [42] |
| Metformin | Exogenous substance (antidiabetic drug) | 990–3171 | [39,43,44] |
| MPP+ | Exogenous substance (laboratory reagent) | 3.1–20 | [39,45,46] |
| Sepantronium | Exogenous substance (anticancer drug) | 2.7 | [47] |
| Serotonin | Endogenous substance (neurotransmitter) | 290 | [40] |
| TEA | Exogenous substance (laboratory reagent) | 28.5–76 | [37,48] |
| Thiamine | Endogenous substance (vitamin precursor) | 750 | [49] |
| Molecular Descriptor | Value Mean | p-Value (t-Test) | |||
|---|---|---|---|---|---|
| OCT2 Substrates | |||||
| Block | Name | Description | Trans-Stimulating | Non-Trans-Stimulating | |
| CI | Mi | Mean first ionization potential (scaled on Carbon atom) | 1.17 | 1.14 | 0.0004 |
| CI | Mp | Mean atomic polarizability (scaled on Carbon atom) | 0.56 | 0.64 | 0.0007 |
| MP | ALOGP | Ghose-Crippen octanol-water partition coeff. (logP) | −1.02 | 0.61 | 0.0012 |
| CI | C% | Percentage of C atoms | 23.10 | 35.93 | 0.0015 |
| CI | Mv | Mean atomic van der Waals volume (scaled on Carbon atom) | 0.51 | 0.60 | 0.0030 |
| MP | Uc | Unsaturation count | 0.90 | 2.82 | 0.0054 |
| CI | GD | Graph density | 0.23 | 0.15 | 0.0061 |
| CI | nAB | Number of aromatic bonds | 0 | 5.73 | 0.0073 |
| MP | BLTF96 | Verhaar Fish base-line toxicity from MLOGP (mmol/L) | 0.04 | −1.83 | 0.0079 |
| MP | MLOGP | Moriguchi octanol-water partition coeff. (logP) | −1.68 | 0.52 | 0.0079 |
| MP | BLTD48 | Verhaar Daphnia base-line toxicity from MLOGP (mmol/L) | 0.28 | −1.81 | 0.0080 |
| MP | BLTA96 | Verhaar Algae base-line toxicity from MLOGP (mmol/L) | 0.45 | −1.74 | 0.0080 |
| CI | nBM | Number of multiple bonds | 1 | 7.36 | 0.0094 |
| CI | nCsp2 | Number of sp2 hybridized Carbon atoms | 1 | 6.82 | 0.0120 |
| CI | SCBO | Sum of conventional bond orders (H-depleted) | 8.75 | 22.23 | 0.0123 |
| MP | Ui | Unsaturation index | 0.90 | 2.30 | 0.0130 |
| CI | nC | Number of Carbon atoms | 5.25 | 12.18 | 0.0131 |
| CI | nBO | Number of non-H bonds | 7.75 | 17.64 | 0.0161 |
| CI | H% | Percentage of H atoms | 61.03 | 50.29 | 0.0188 |
| CD | RNCG | Relative negative charge | 0.48 | 0.25 | 0.0195 |
| CI | nSK | Number of non-H atoms | 8.75 | 16.82 | 0.0225 |
| CI | MW | Molecular weight | 127.5 | 238.7 | 0.0254 |
| CI | Sv | Sum of atomic van der Waals volumes (scaled on carbon atom) | 11.49 | 20.28 | 0.0295 |
| CI | Sp | Sum of atomic polarizabilities (scaled on carbon atom) | 12.54 | 21.65 | 0.0325 |
| CI | AMW | Average molecular weight | 5.68 | 7.05 | 0.0350 |
| MP | Vx | McGowan volume | 188.5 | 309.8 | 0.0366 |
| MP | VvdwMG | van der Waals volume from McGowan volume | 80.62 | 130.3 | 0.0366 |
| MP | AMR | Ghose–Crippen molar refractivity | 35.26 | 62.75 | 0.0383 |
| MP | VvdwZAZ | van der Waals volume from Zhao–Abraham–Zissimos equation | 134.2 | 225 | 0.0395 |
| MP | PDI | Packing density index | 0.82 | 0.91 | 0.0418 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lefèvre, C.R.; Le Vée, M.; Gaubert, S.; Jouan, E.; Bruyere, A.; Moreau, C.; Fardel, O. Substrate-Dependent Trans-Stimulation of Organic Cation Transporter 2 Activity. Int. J. Mol. Sci. 2021, 22, 12926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312926
Lefèvre CR, Le Vée M, Gaubert S, Jouan E, Bruyere A, Moreau C, Fardel O. Substrate-Dependent Trans-Stimulation of Organic Cation Transporter 2 Activity. International Journal of Molecular Sciences. 2021; 22(23):12926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312926
Chicago/Turabian StyleLefèvre, Charles R., Marc Le Vée, Sophie Gaubert, Elodie Jouan, Arnaud Bruyere, Caroline Moreau, and Olivier Fardel. 2021. "Substrate-Dependent Trans-Stimulation of Organic Cation Transporter 2 Activity" International Journal of Molecular Sciences 22, no. 23: 12926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312926