
Interleukin-35 Prevents Development of Autoimmune Diabetes Possibly by Maintaining the Phenotype of Regulatory B Cells

 ,
,
Abstract
:1. Introduction
2. Results
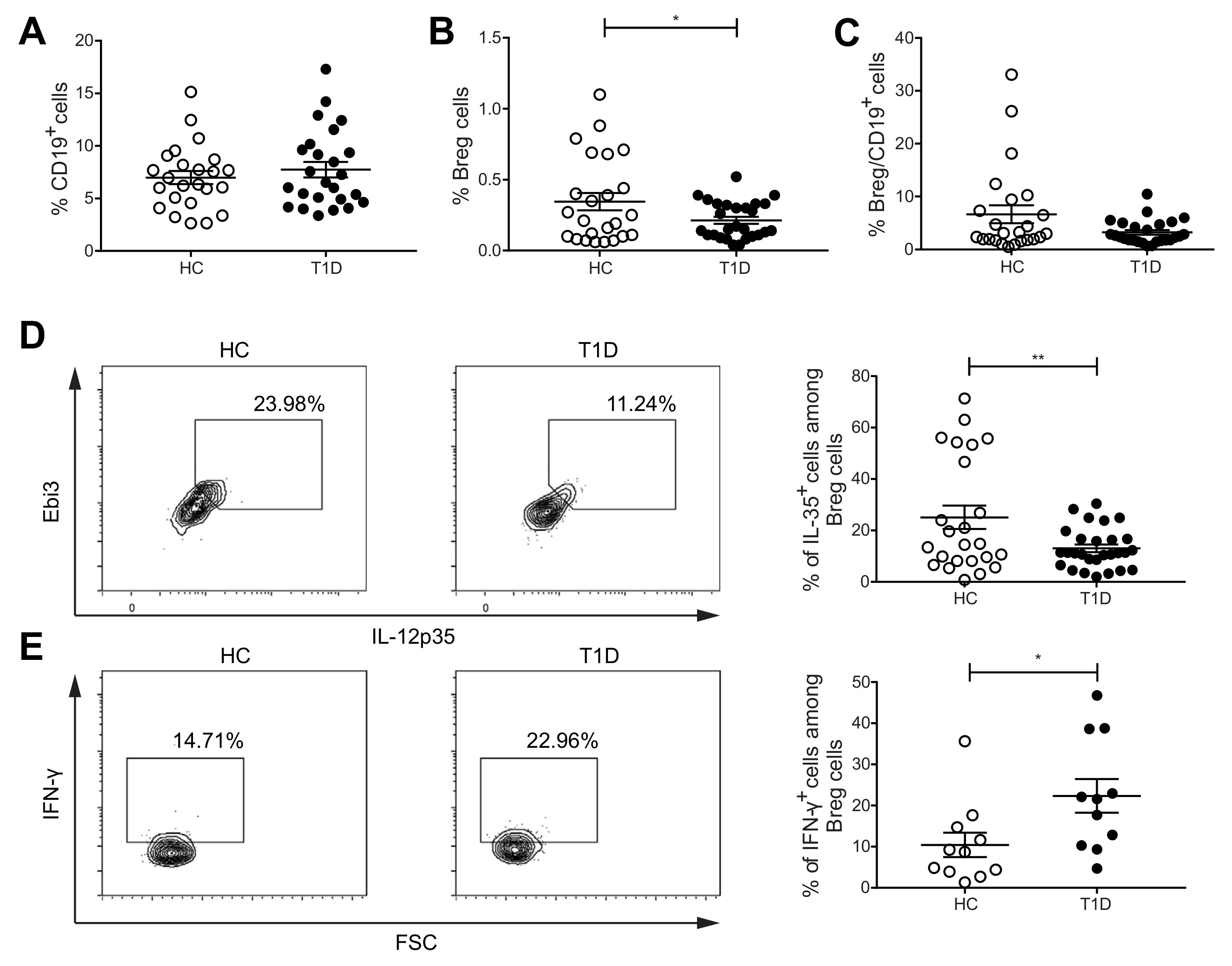
2.1. The Proportions of IL-35+Cells among Breg Cells Are Decreased, but IFN-γ+ Cells among Breg Cells Are Increased in PBMCs of Subjects with T1D
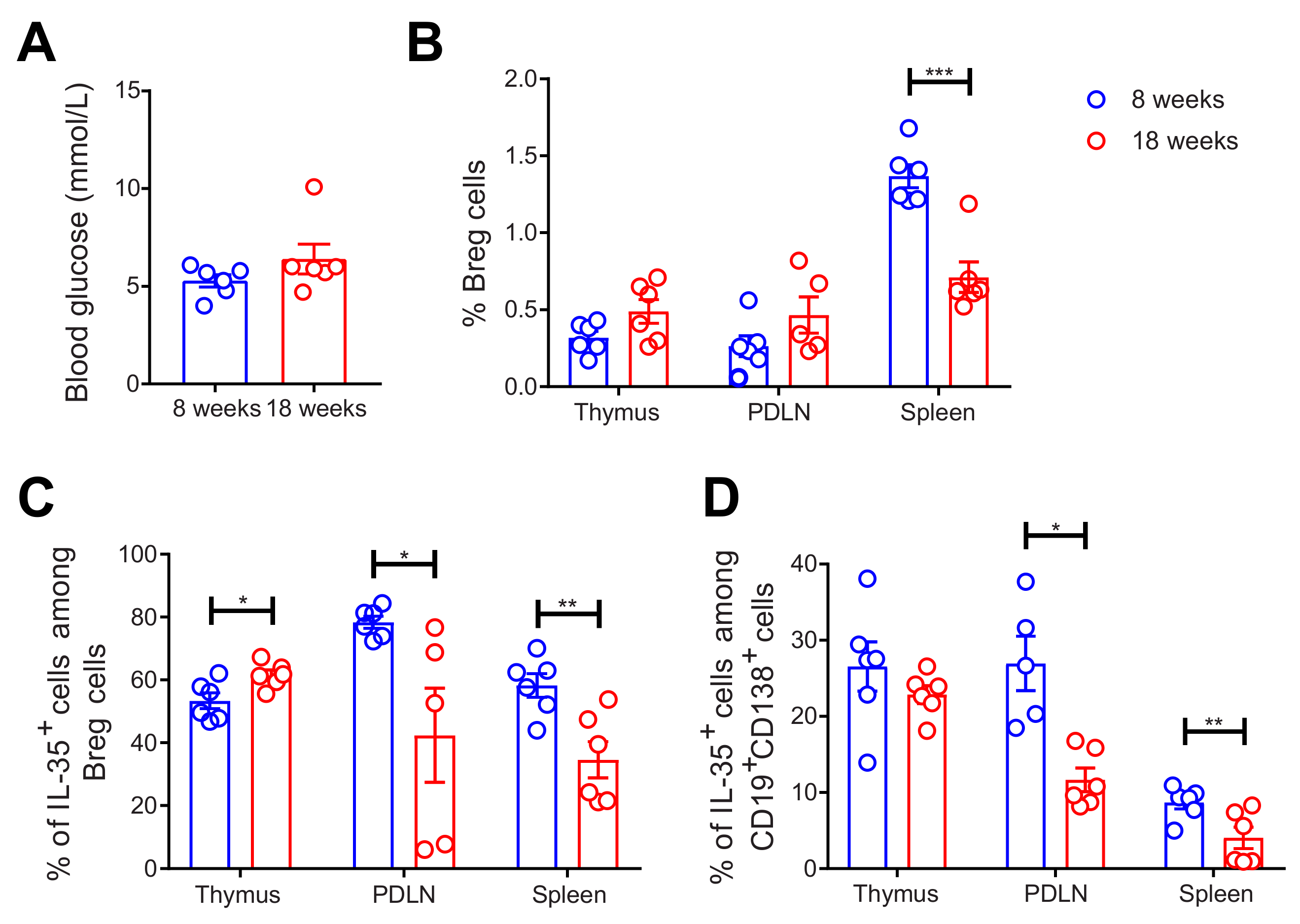
2.2. Decreased Proportions of Breg and IL-35+ Cells among Breg Cells in 18-Week-Old NOD Mice
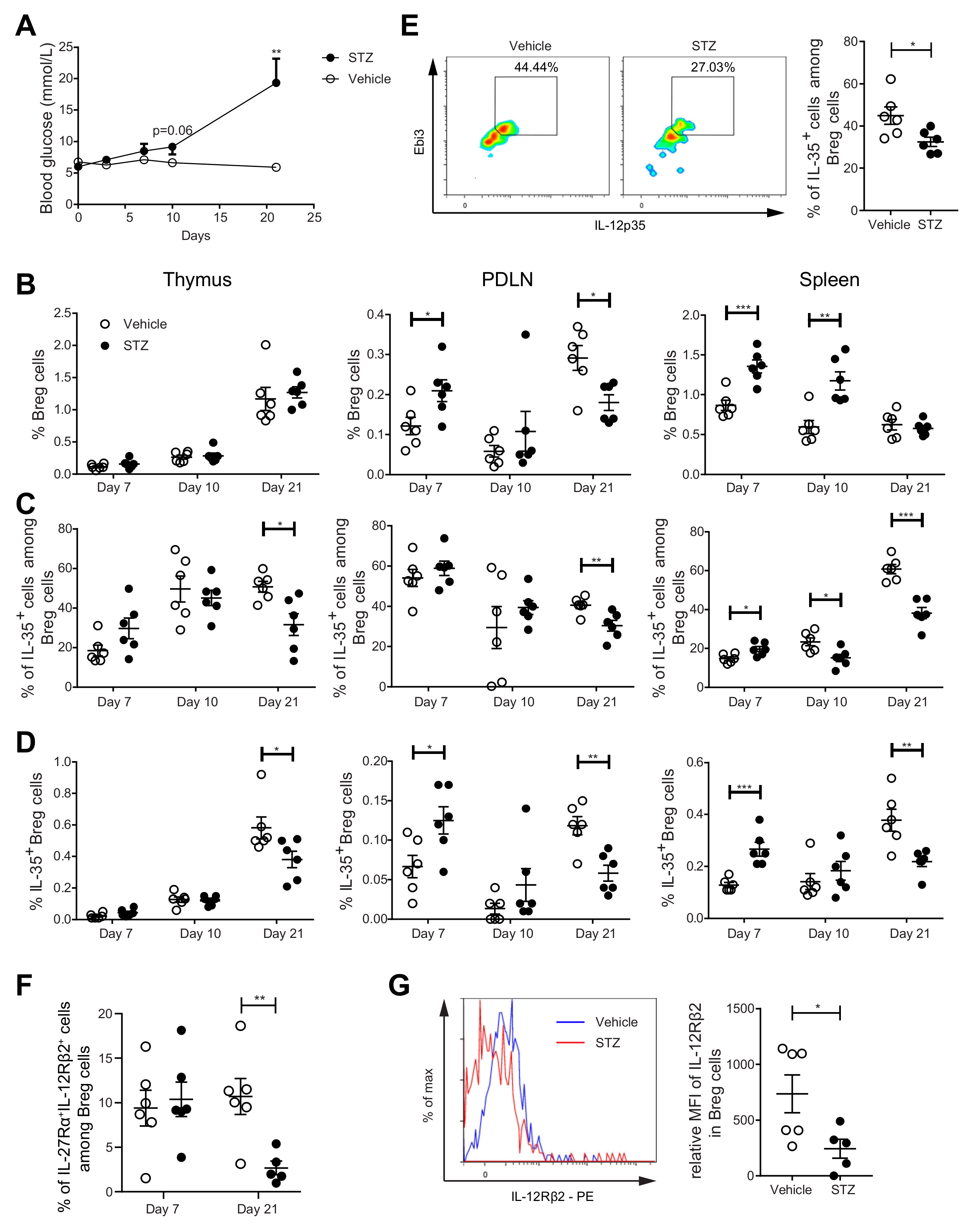
2.3. The Proportions of IL-35+ Breg Cells Are Increased at the Early Stage, but Decreased at the Late Stage, of Experimental T1D Development in MLDSTZ Mice
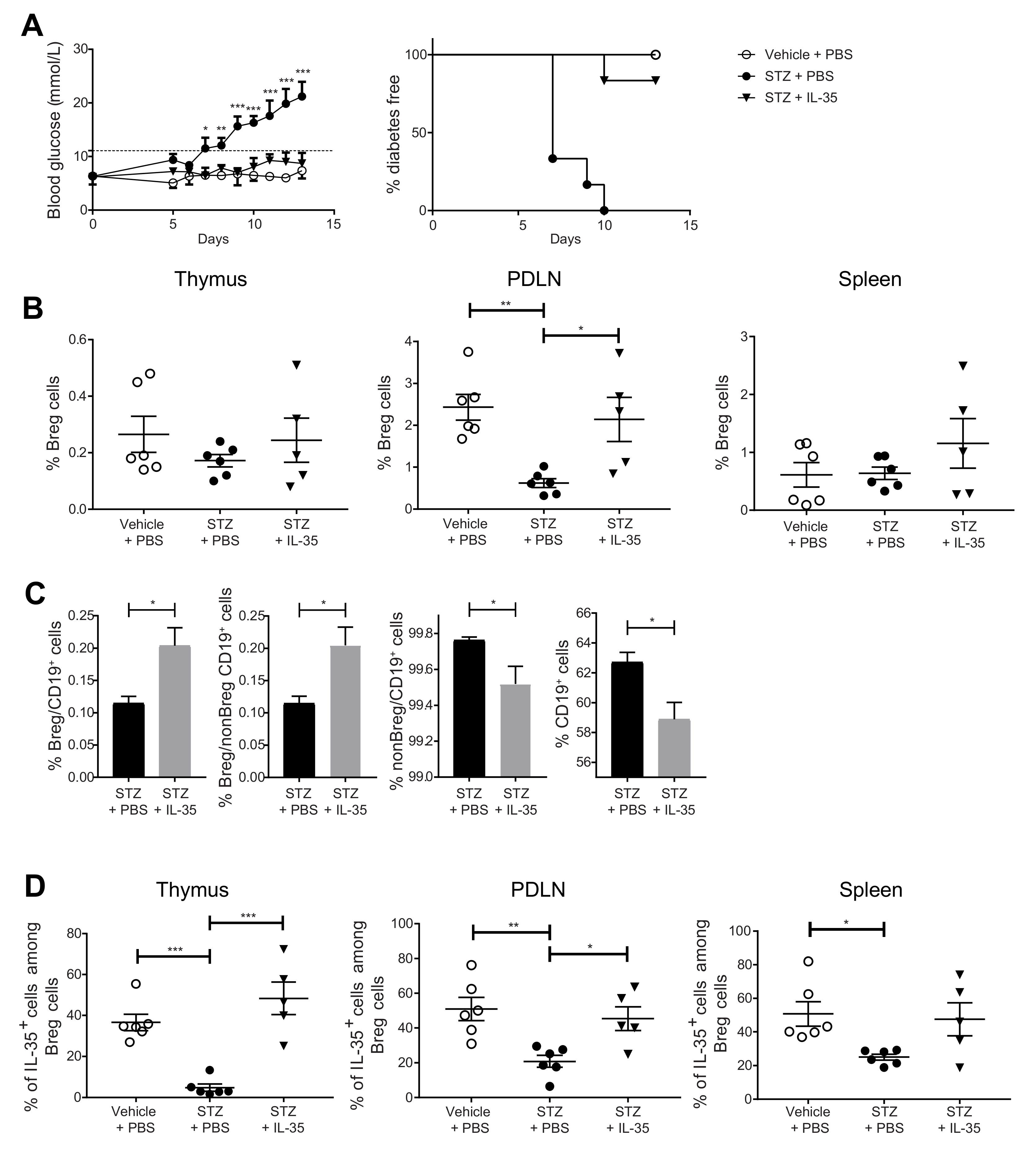
2.4. Systemic Administration of IL-35 Increases the Proportion of IL-35+ Cells among Breg Cells and Prevents the Development of Experimental T1D
2.5. IL-35 Treatment May Prevent the Phenotypic Shift of Breg Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Single Cell Preparation
4.3. Primary Cell Culture, Cell Depletion, and Stimulation for Cytokine Staining
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Patient Recruitment, Peripheral Blood Mononuclear Cells (PBMCs) Preparation
4.6. Monoclonal Antibody Staining and Flow Cytometry
4.7. Breg Cell Sorting
4.8. Quantitative Real-Time-PCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosser, E.C.; Mauri, C. Regulatory B cells: Origin, phenotype, and function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, A.; Mizoguchi, E.; Takedatsu, H.; Blumberg, R.S.; Bhan, A.K. Chronic intestinal inflammatory condition generates IL-10-producing regulatory B cell subset characterized by CD1d upregulation. Immunity 2002, 16, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Fillatreau, S.; Sweenie, C.H.; McGeachy, M.J.; Gray, D.; Anderton, S.M. B cells regulate autoimmunity by provision of IL-10. Nat. Immunol. 2002, 3, 944–950. [Google Scholar] [CrossRef]
- Mauri, C.; Gray, D.; Mushtaq, N.; Londei, M. Prevention of arthritis by interleukin 10-producing B cells. J. Exp. Med. 2003, 197, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Devergne, O.; Birkenbach, M.; Kieff, E. Epstein-Barr virus-induced gene 3 and the p35 subunit of interleukin 12 form a novel heterodimeric hematopoietin. Proc. Natl. Acad. Sci. USA 1997, 94, 12041–12046. [Google Scholar] [CrossRef] [Green Version]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef]
- Niedbala, W.; Wei, X.Q.; Cai, B.; Hueber, A.J.; Leung, B.P.; McInnes, I.B.; Liew, F.Y. IL-35 is a novel cytokine with therapeutic effects against collagen-induced arthritis through the expansion of regulatory T cells and suppression of Th17 cells. Eur. J. Immunol. 2007, 37, 3021–3029. [Google Scholar] [CrossRef]
- Wang, R.X.; Yu, C.R.; Dambuza, I.M.; Mahdi, R.M.; Dolinska, M.B.; Sergeev, Y.V.; Wingfield, P.T.; Kim, S.H.; Egwuagu, C.E. Interleukin-35 induces regulatory B cells that suppress autoimmune disease. Nat. Med. 2014, 20, 633–641. [Google Scholar] [CrossRef]
- Shen, P.; Roch, T.; Lampropoulou, V.; O’Connor, R.A.; Stervbo, U.; Hilgenberg, E.; Ries, S.; Dang, V.D.; Jaimes, Y.; Daridon, C.; et al. IL-35-producing B cells are critical regulators of immunity during autoimmune and infectious diseases. Nature 2014, 507, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Blair, P.A.; Noreña, L.Y.; Flores-Borja, F.; Rawlings, D.J.; Isenberg, D.A.; Ehrenstein, M.R.; Mauri, C. CD19(+)CD24(hi)CD38(hi) B cells exhibit regulatory capacity in healthy individuals but are functionally impaired in systemic Lupus Erythematosus patients. Immunity 2010, 32, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Yanaba, K.; Bouaziz, J.D.; Haas, K.M.; Poe, J.C.; Fujimoto, M.; Tedder, T.F. A regulatory B cell subset with a unique CD1dhiCD5+ phenotype controls T cell-dependent inflammatory responses. Immunity 2008, 28, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Mirlekar, B.; Michaud, D.; Pylayeva-Gupta, Y. IL-35 Detection in B Cells at the mRNA and Protein Level. Methods. Mol. Biol. 2021, 2270, 125–147. [Google Scholar] [CrossRef]
- Choi, J.K.; Egwuagu, C.E. Interleukin 35 Regulatory B Cells. J. Mol. Biol. 2021, 433, 166607. [Google Scholar] [CrossRef]
- Flores-Borja, F.; Bosma, A.; Ng, D.; Reddy, V.; Ehrenstein, M.R.; Isenberg, D.A.; Mauri, C. CD19+ CD24hiCD38hi B cells maintain regulatory T cells while limiting TH1 and TH17 differentiation. Sci. Transl. Med. 2013, 5, 173ra123. [Google Scholar] [CrossRef]
- Oka, A.; Ishihara, S.; Mishima, Y.; Tada, Y.; Kusunoki, R.; Fukuba, N.; Yuki, T.; Kawashima, K.; Matsumoto, S.; Kinoshita, Y. Role of regulatory B cells in chronic intestinal inflammation: Association with pathogenesis of Crohn’s disease. Inflamm. Bowel. Dis. 2014, 20, 315–328. [Google Scholar] [CrossRef]
- Knippenberg, S.; Peelen, E.; Smolders, J.; Thewissen, M.; Menheere, P.; Cohen Tervaert, J.W.; Hupperts, R.; Damoiseaux, J. Reduction in IL-10 producing B cells (Breg) in multiple sclerosis is accompanied by a reduced naïve/memory Breg ratio during a relapse but not in remission. J. Neuroimmunol. 2011, 239, 80–86. [Google Scholar] [CrossRef]
- Hayashi, M.; Yanaba, K.; Umezawa, Y.; Yoshihara, Y.; Kikuchi, S.; Ishiuji, Y.; Saeki, H.; Nakagawa, H. IL-10-producing regulatory B cells are decreased in patients with psoriasis. J. Dermatol. Sci. 2016, 81, 93–100. [Google Scholar] [CrossRef]
- Carter, N.A.; Rosser, E.C.; Mauri, C. Interleukin-10 produced by B cells is crucial for the suppression of Th17/Th1 responses, induction of T regulatory type 1 cells and reduction of collagen-induced arthritis. Arthritis. Res. Ther. 2012, 14, R32. [Google Scholar] [CrossRef] [Green Version]
- Mauri, C.; Menon, M. Human regulatory B cells in health and disease: Therapeutic potential. J. Clin. Investig. 2017, 127, 772–779. [Google Scholar] [CrossRef] [Green Version]
- Sha, S.; Pearson, J.A.; Peng, J.; Hu, Y.; Huang, J.; Xing, Y.; Zhang, L.; Zhu, Y.; Zhao, H.; Wong, F.S.; et al. TLR9 Deficiency in B Cells Promotes Immune Tolerance via Interleukin-10 in a Type 1 Diabetes Mouse Model. Diabetes 2021, 70, 504–515. [Google Scholar] [CrossRef]
- El-Mokhtar, M.A.; Elsherbiny, N.M.; Sayed, D.; Raafat, D.M.; Askar, E.; Hussein, A.; Abdel-Malek, M.A.Y.; Shalaby, A.M. Altered Regulatory B Cell Subsets in Children with Type 1 Diabetes Mellitus. J. Immunol. Res. 2020, 2020, 8935694. [Google Scholar] [CrossRef]
- Kleffel, S.; Vergani, A.; Tezza, S.; Ben Nasr, M.; Niewczas, M.A.; Wong, S.; Bassi, R.; D’Addio, F.; Schatton, T.; Abdi, R.; et al. Interleukin-10+ regulatory B cells arise within antigen-experienced CD40+ B cells to maintain tolerance to islet autoantigens. Diabetes 2015, 64, 158–171. [Google Scholar] [CrossRef] [Green Version]
- Marwaha, A.K.; Crome, S.Q.; Panagiotopoulos, C.; Berg, K.B.; Qin, H.; Ouyang, Q.; Xu, L.; Priatel, J.J.; Levings, M.K.; Tan, R. Cutting edge: Increased IL-17-secreting T cells in children with new-onset type 1 diabetes. J. Immunol. 2010, 185, 3814–3818. [Google Scholar] [CrossRef]
- McClymont, S.A.; Putnam, A.L.; Lee, M.R.; Esensten, J.H.; Liu, W.; Hulme, M.A.; Hoffmüller, U.; Baron, U.; Olek, S.; Bluestone, J.A.; et al. Plasticity of human regulatory T cells in healthy subjects and patients with type 1 diabetes. J. Immunol. 2011, 186, 3918–3926. [Google Scholar] [CrossRef]
- Singh, K.; Kadesjö, E.; Lindroos, J.; Hjort, M.; Lundberg, M.; Espes, D.; Carlsson, P.O.; Sandler, S.; Thorvaldson, L. Interleukin-35 administration counteracts established murine type 1 diabetes--possible involvement of regulatory T cells. Sci. Rep. 2015, 5, 12633. [Google Scholar] [CrossRef] [Green Version]
- Putnam, A.L.; Brusko, T.M.; Lee, M.R.; Liu, W.; Szot, G.L.; Ghosh, T.; Atkinson, M.A.; Bluestone, J.A. Expansion of human regulatory T-cells from patients with type 1 diabetes. Diabetes 2009, 58, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Espes, D.; Singh, K.; Sandler, S.; Carlsson, P.O. Increased Interleukin-35 Levels in Patients With Type 1 Diabetes With Remaining C-Peptide. Diabetes Care 2017, 40, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Pinto, A.I.; Smith, J.; Kissack, M.R.; Hogg, K.G.; Green, E.A. Thymic B Cell-Mediated Attack of Thymic Stroma Precedes Type 1 Diabetes Development. Front. Immunol. 2018, 9, 1281. [Google Scholar] [CrossRef]
- Chen, Y.G.; Mathews, C.E.; Driver, J.P. The Role of NOD Mice in Type 1 Diabetes Research: Lessons from the Past and Recommendations for the Future. Front. Endocrinol. 2018, 9, 51. [Google Scholar] [CrossRef]
- Luo, Z.; Soläng, C.; Mejia-Cordova, M.; Thorvaldson, L.; Blixt, M.; Sandler, S.; Singh, K. Kinetics of immune cell responses in the multiple low-dose streptozotocin mouse model of type 1 diabetes. FASEB Bioadv. 2019, 1, 538–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, G.P.; Ettinger, R.; Shirota, Y.; Yarboro, C.H.; Illei, G.G.; Lipsky, P.E. Identification and characterization of circulating human transitional B cells. Blood 2005, 105, 4390–4398. [Google Scholar] [CrossRef] [PubMed]
- Schloot, N.C.; Hanifi-Moghaddam, P.; Goebel, C.; Shatavi, S.V.; Flohé, S.; Kolb, H.; Rothe, H. Serum IFN-gamma and IL-10 levels are associated with disease progression in non-obese diabetic mice. Diabetes Metab. Res. Rev. 2002, 18, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Li, L.; Garland, A.; He, Q.; Wang, H.; Katz, J.D.; Tisch, R.; Wang, B. IFN-γ receptor deficiency prevents diabetes induction by diabetogenic CD4+, but not CD8+, T cells. Eur. J. Immunol. 2012, 42, 2010–2018. [Google Scholar] [CrossRef]
- El-Sheikh, A.; Suarez-Pinzon, W.L.; Power, R.F.; Rabinovitch, A. Both CD4(+)and CD8(+)T cells are required for IFN-gamma gene expression in pancreatic islets and autoimmune diabetes development in biobreeding rats. J. Autoimmun. 1999, 12, 109–119. [Google Scholar] [CrossRef]
- Wang, B.; André, I.; Gonzalez, A.; Katz, J.D.; Aguet, M.; Benoist, C.; Mathis, D. Interferon-gamma impacts at multiple points during the progression of autoimmune diabetes. Proc. Natl. Acad. Sci. USA 1997, 94, 13844–13849. [Google Scholar] [CrossRef] [Green Version]
- Arif, S.; Tree, T.I.; Astill, T.P.; Tremble, J.M.; Bishop, A.J.; Dayan, C.M.; Roep, B.O.; Peakman, M. Autoreactive T cell responses show proinflammatory polarization in diabetes but a regulatory phenotype in health. J. Clin. Investig. 2004, 113, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Norihisa, Y.; Benjamin, D.; Kantor, R.R.; Young, H.A. Interferon-gamma gene expression in human B-cell lines: Induction by interleukin-2, protein kinase C activators, and possible effect of hypomethylation on gene regulation. Blood 1992, 80, 724–732. [Google Scholar] [CrossRef] [Green Version]
- Dayton, M.A.; Knobloch, T.J.; Benjamin, D. Human B cell lines express the interferon gamma gene. Cytokine 1992, 4, 454–460. [Google Scholar] [CrossRef]
- Harris, D.P.; Haynes, L.; Sayles, P.C.; Duso, D.K.; Eaton, S.M.; Lepak, N.M.; Johnson, L.L.; Swain, S.L.; Lund, F.E. Reciprocal regulation of polarized cytokine production by effector B and T cells. Nat. Immunol. 2000, 1, 475–482. [Google Scholar] [CrossRef]
- Jackson, S.W.; Jacobs, H.M.; Arkatkar, T.; Dam, E.M.; Scharping, N.E.; Kolhatkar, N.S.; Hou, B.; Buckner, J.H.; Rawlings, D.J. B cell IFN-γ receptor signaling promotes autoimmune germinal centers via cell-intrinsic induction of BCL-6. J. Exp. Med. 2016, 213, 733–750. [Google Scholar] [CrossRef] [Green Version]
- Chodisetti, S.B.; Fike, A.J.; Domeier, P.P.; Choi, N.M.; Soni, C.; Rahman, Z.S.M. TLR7 Negatively Regulates B10 Cells Predominantly in an IFNγ Signaling Dependent Manner. Front. Immunol. 2020, 11, 1632. [Google Scholar] [CrossRef]
- Tai, N.; Wong, F.S.; Wen, L. TLR9 deficiency promotes CD73 expression in T cells and diabetes protection in nonobese diabetic mice. J. Immunol. 2013, 191, 2926–2937. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lee, A.S.; Shameli, A.; Geng, X.; Finegood, D.; Santamaria, P.; Dutz, J.P. TLR9 blockade inhibits activation of diabetogenic CD8+ T cells and delays autoimmune diabetes. J. Immunol. 2010, 184, 5645–5653. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Peng, J.; Tai, N.; Pearson, J.A.; Hu, C.; Guo, J.; Hou, L.; Zhao, H.; Wong, F.S.; Wen, L. Toll-like receptor 9 negatively regulates pancreatic islet beta cell growth and function in a mouse model of type 1 diabetes. Diabetologia 2018, 61, 2333–2343. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, F.; Johnson, M.C.; Li, C.; Samulski, R.J.; Wang, B.; Tisch, R. β-cell-specific IL-35 therapy suppresses ongoing autoimmune diabetes in NOD mice. Eur. J. Immunol. 2017, 47, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Bettini, M.; Castellaw, A.H.; Lennon, G.P.; Burton, A.R.; Vignali, D.A. Prevention of autoimmune diabetes by ectopic pancreatic β-cell expression of interleukin-35. Diabetes 2012, 61, 1519–1526. [Google Scholar] [CrossRef] [Green Version]
- Pescovitz, M.D.; Greenbaum, C.J.; Krause-Steinrauf, H.; Becker, D.J.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Marks, J.B.; McGee, P.F.; Moran, A.M.; et al. Rituximab, B-lymphocyte depletion, and preservation of beta-cell function. N. Engl. J. Med. 2009, 361, 2143–2152. [Google Scholar] [CrossRef]
- Bloem, S.J.; Roep, B.O. The elusive role of B lymphocytes and islet autoantibodies in (human) type 1 diabetes. Diabetologia 2017, 60, 1185–1189. [Google Scholar] [CrossRef] [Green Version]
- Like, A.A.; Rossini, A.A. Streptozotocin-induced pancreatic insulitis: New model of diabetes mellitus. Science 1976, 193, 415–417. [Google Scholar] [CrossRef]
- Luo, Z.; Thorvaldson, L.; Blixt, M.; Singh, K. Determination of Regulatory T Cell Subsets in Murine Thymus, Pancreatic Draining Lymph Node and Spleen Using Flow Cytometry. JoVE 2019, 144, e58848. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Takaki, S.; Nagasaki, M.; Otsu, M.; Yamashita, H.; Sugita, J.; Yoshimura, K.; Eto, K.; Komuro, I.; et al. Adipose Natural Regulatory B Cells Negatively Control Adipose Tissue Inflammation. Cell Metab. 2013, 18, 759–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Martinell, M.; Luo, Z.; Espes, D.; Stålhammar, J.; Sandler, S.; Carlsson, P.O. Cellular immunological changes in patients with LADA are a mixture of those seen in patients with type 1 and type 2 diabetes. Clin. Exp. Immunol. 2019, 197, 64–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 24) | T1D (n = 29) | |
|---|---|---|
| Age (years) | 42.4 ± 3.1 | 44.0 ± 3.1 |
| Male gender (n, %) | 13 (54.2) | 16 (55.2) |
| BMI (kg/m2) | 27.0 ± 1.3 | 25.6 ± 0.7 |
| Age at onset (years) | n.a. | 22.8 ± 2.5 |
| Disease duration (years) | n.a. | 21.2 ± 3.1 |
| HbA1c (mmol/mol) | 33.8 ± 0.6 | 59.2 ± 2.2 *** |
| HbA1c (%) | 5.2 ± 0.05 | 7.6 ± 0.2 *** |
| fC-peptide (nmol/L) | 0.9 ± 0.09 | 0.07 ± 0.03 *** |
| fp-glucose (mmol/L) | 5.7 ± 0.1 | 10.6 ± 0.7 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Z.; Lundin, S.; Mejia-Cordova, M.; Hassani, I.; Blixt, M.; Hjelmqvist, D.; Lau, J.; Espes, D.; Carlsson, P.-O.; Sandler, S.; et al. Interleukin-35 Prevents Development of Autoimmune Diabetes Possibly by Maintaining the Phenotype of Regulatory B Cells. Int. J. Mol. Sci. 2021, 22, 12988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312988
Luo Z, Lundin S, Mejia-Cordova M, Hassani I, Blixt M, Hjelmqvist D, Lau J, Espes D, Carlsson P-O, Sandler S, et al. Interleukin-35 Prevents Development of Autoimmune Diabetes Possibly by Maintaining the Phenotype of Regulatory B Cells. International Journal of Molecular Sciences. 2021; 22(23):12988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312988
Chicago/Turabian StyleLuo, Zhengkang, Sara Lundin, Mariela Mejia-Cordova, Imane Hassani, Martin Blixt, Daisy Hjelmqvist, Joey Lau, Daniel Espes, Per-Ola Carlsson, Stellan Sandler, and et al. 2021. "Interleukin-35 Prevents Development of Autoimmune Diabetes Possibly by Maintaining the Phenotype of Regulatory B Cells" International Journal of Molecular Sciences 22, no. 23: 12988. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312988