
Protective Effects of Glutamine and Leucine Supplementation on Sepsis-Induced Skeletal Muscle Injuries

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Body Weight (BW) Changes and Gastrocnemius (GA) Muscle Weights
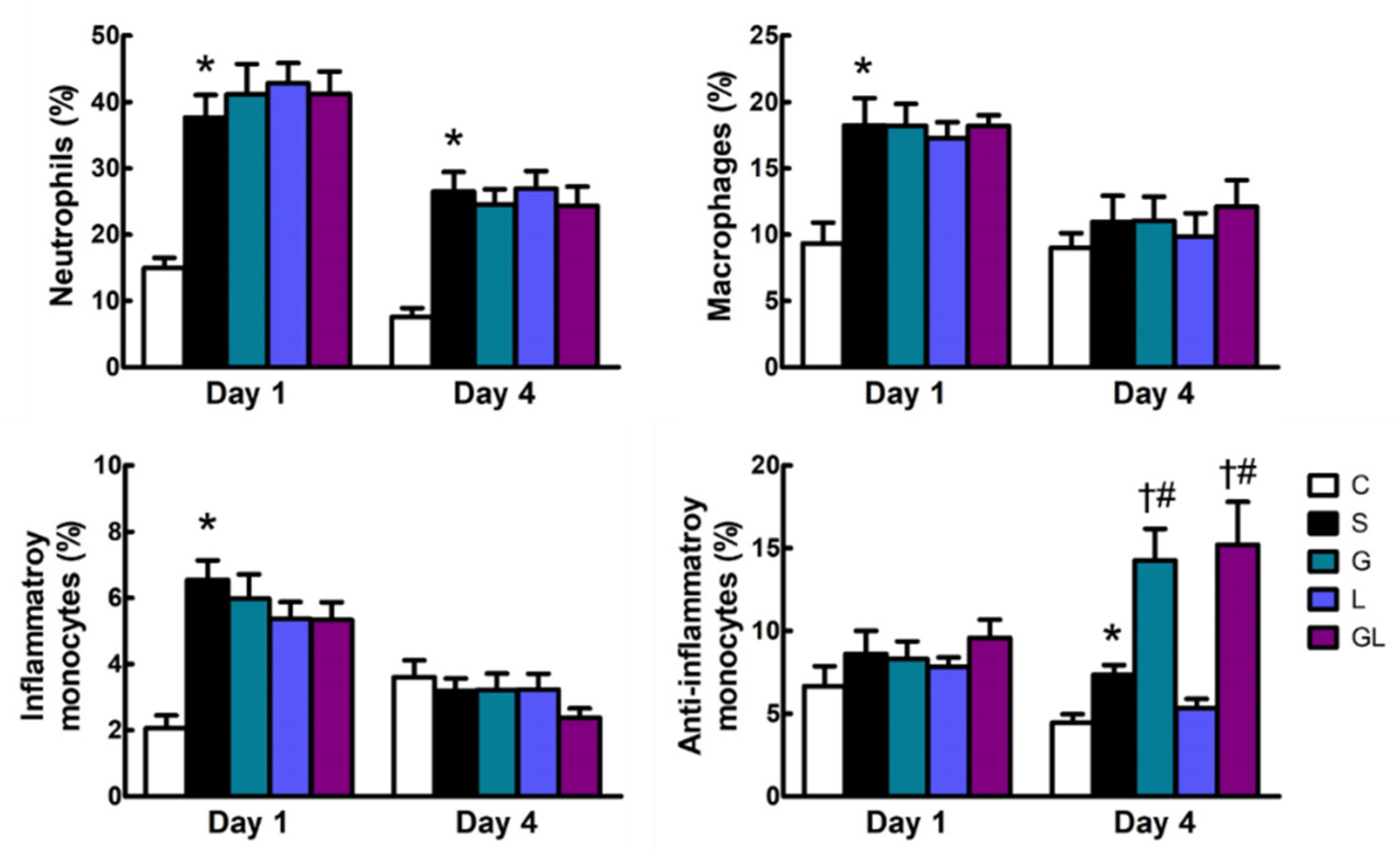
2.2. Leukocyte Populations in the Blood
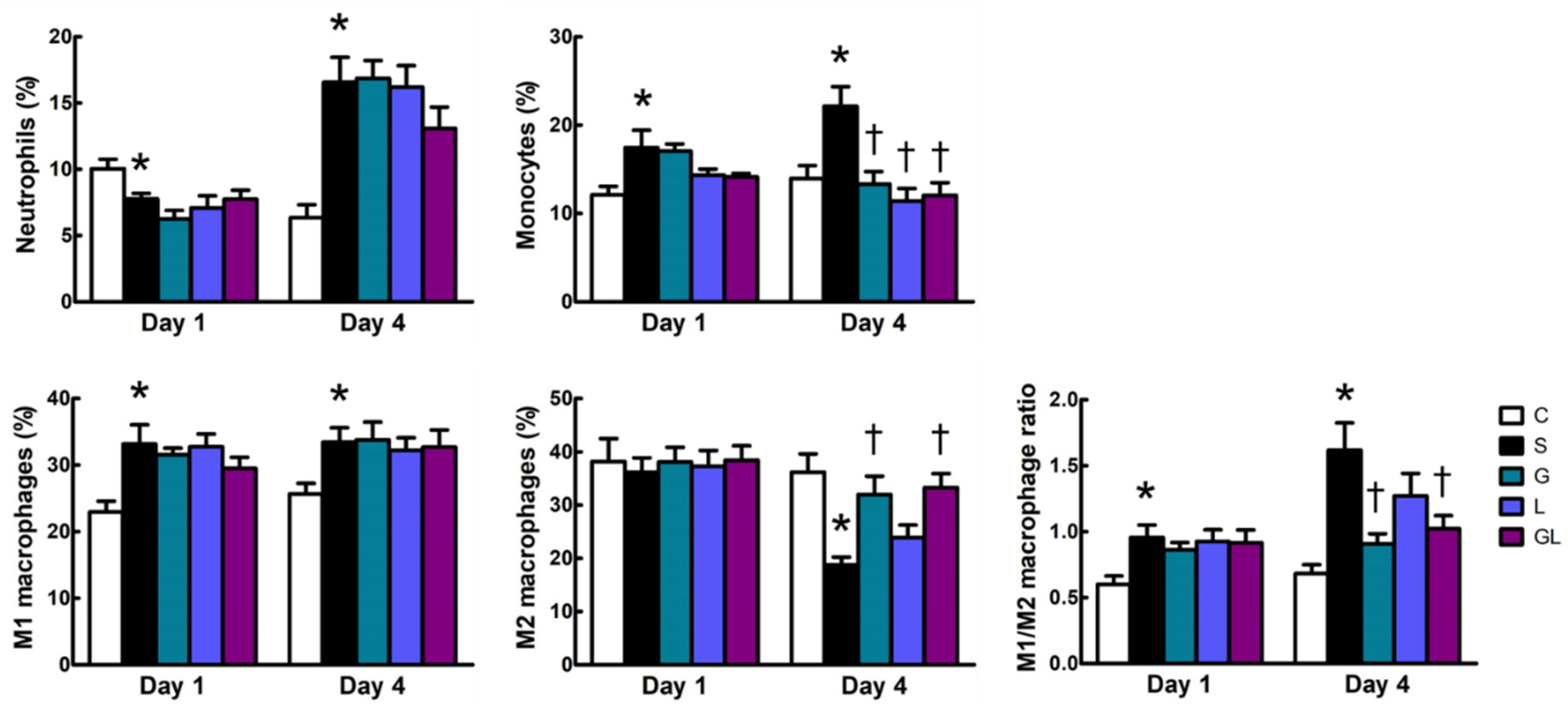
2.3. Leukocyte Populations in Muscles
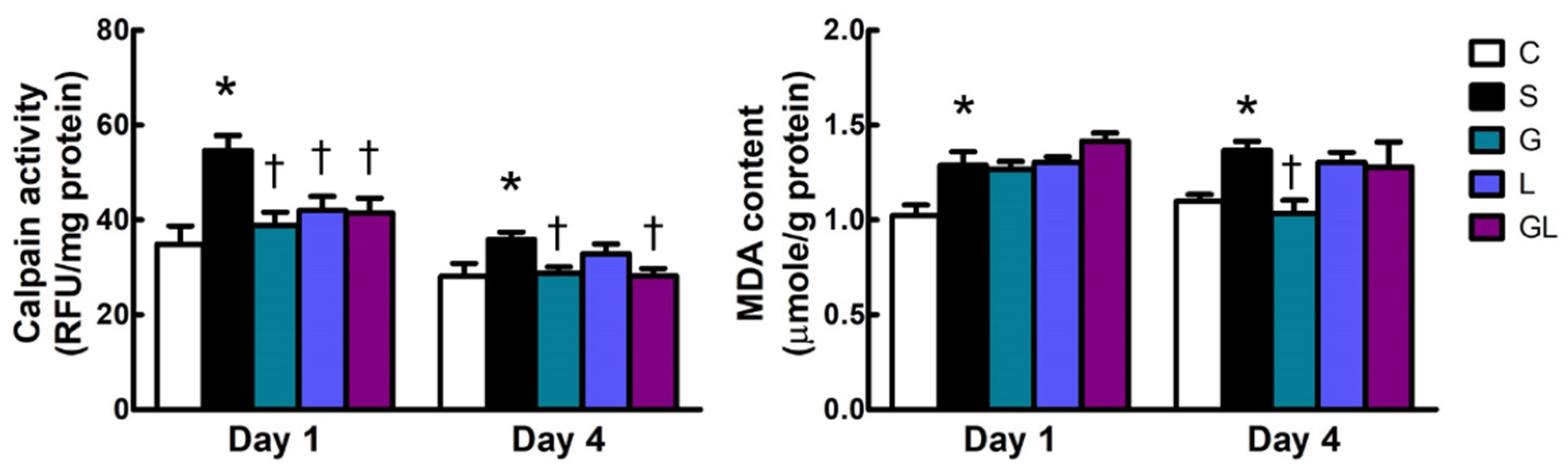
2.4. Calpain Activity and Malondialdehyde (MDA) Content in Muscle Tissues
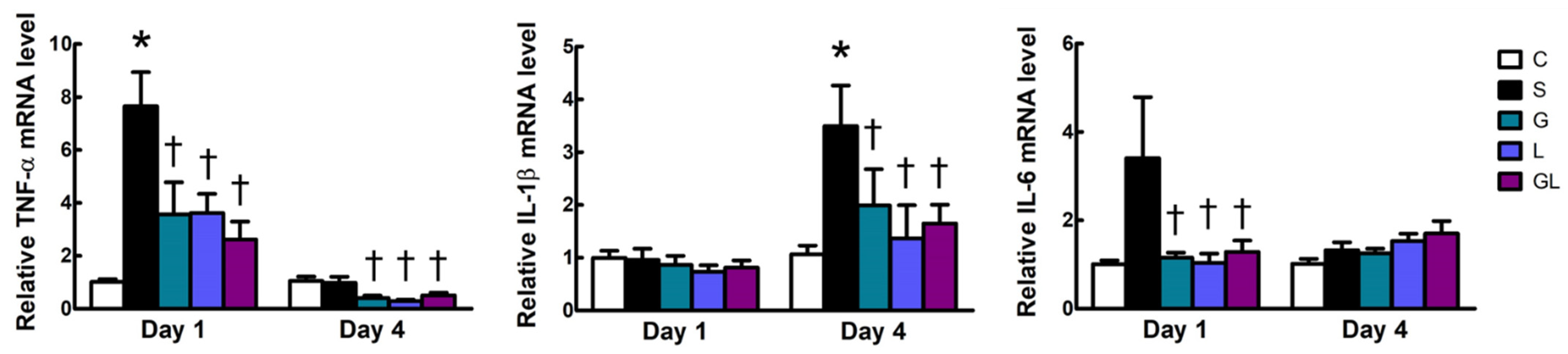
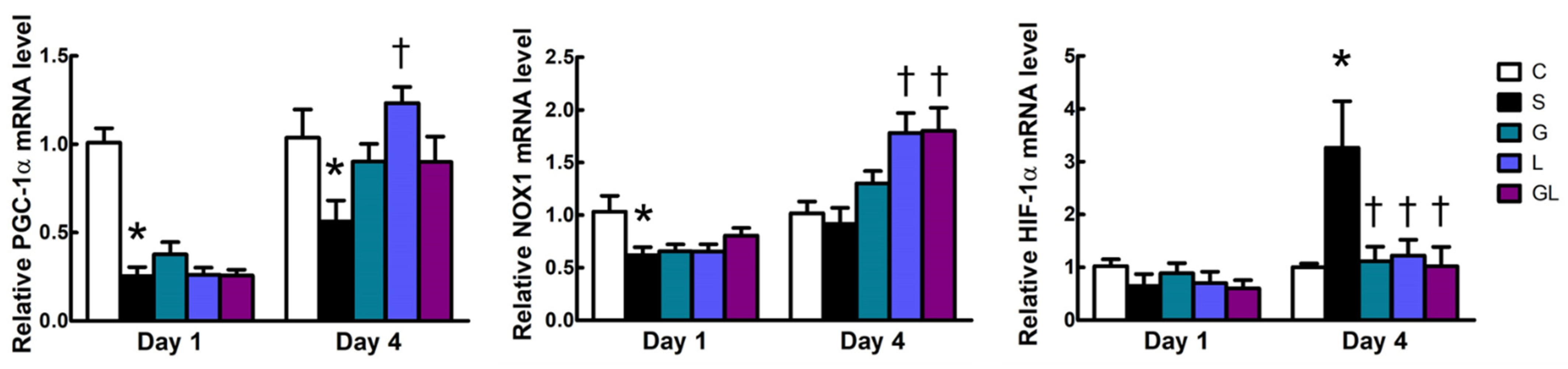
2.5. Muscle mRNA Expressions of Inflammatory and Mitochondrion-Related Genes
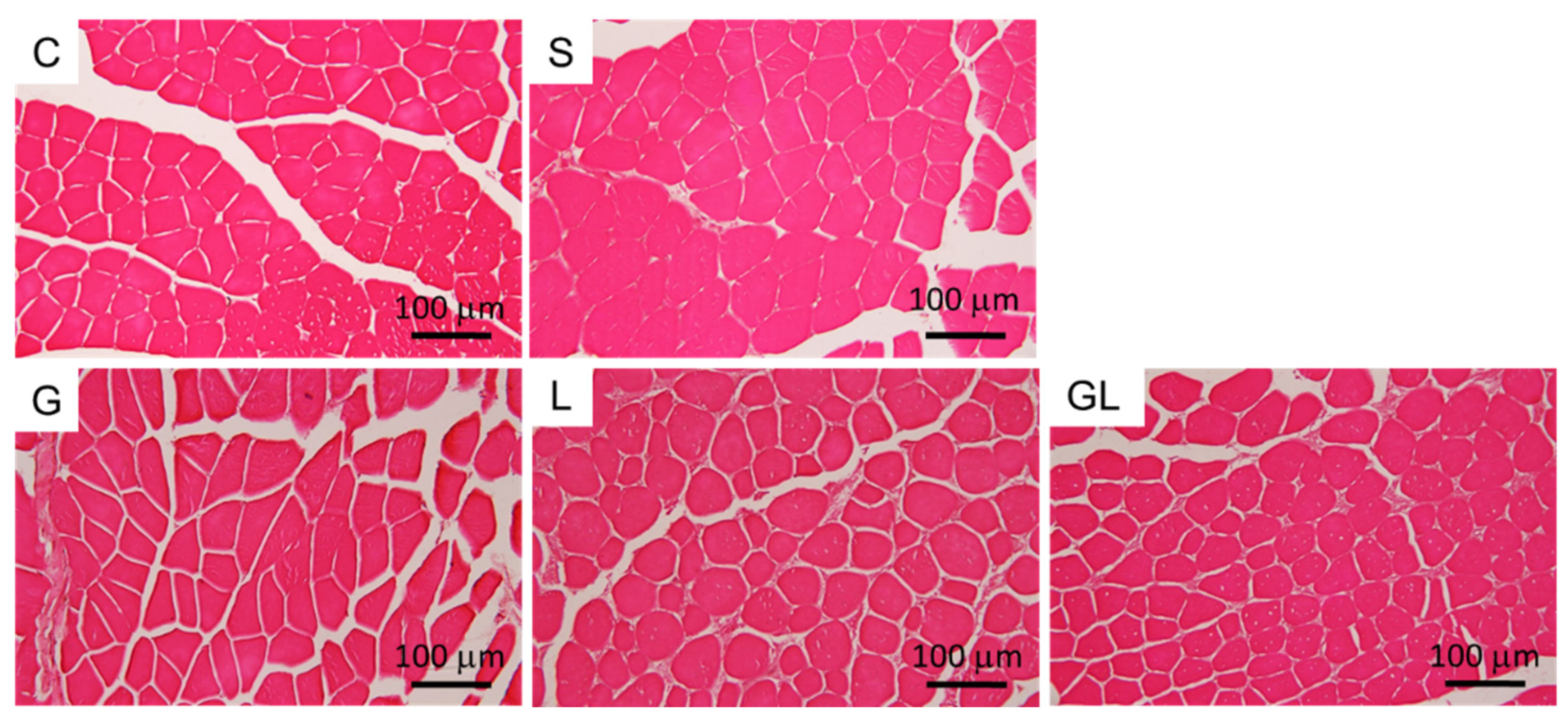
2.6. Muscle Histology
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Protocols
4.3. Distribution of Circulating Leukocytes
4.4. Leukocyte Populations in Muscles
4.5. Calpain Activity and MDA Concentration
4.6. Real-Time Polymerase Chain Reaction (PCR)
4.7. Histology
4.8. Statistical Analysis
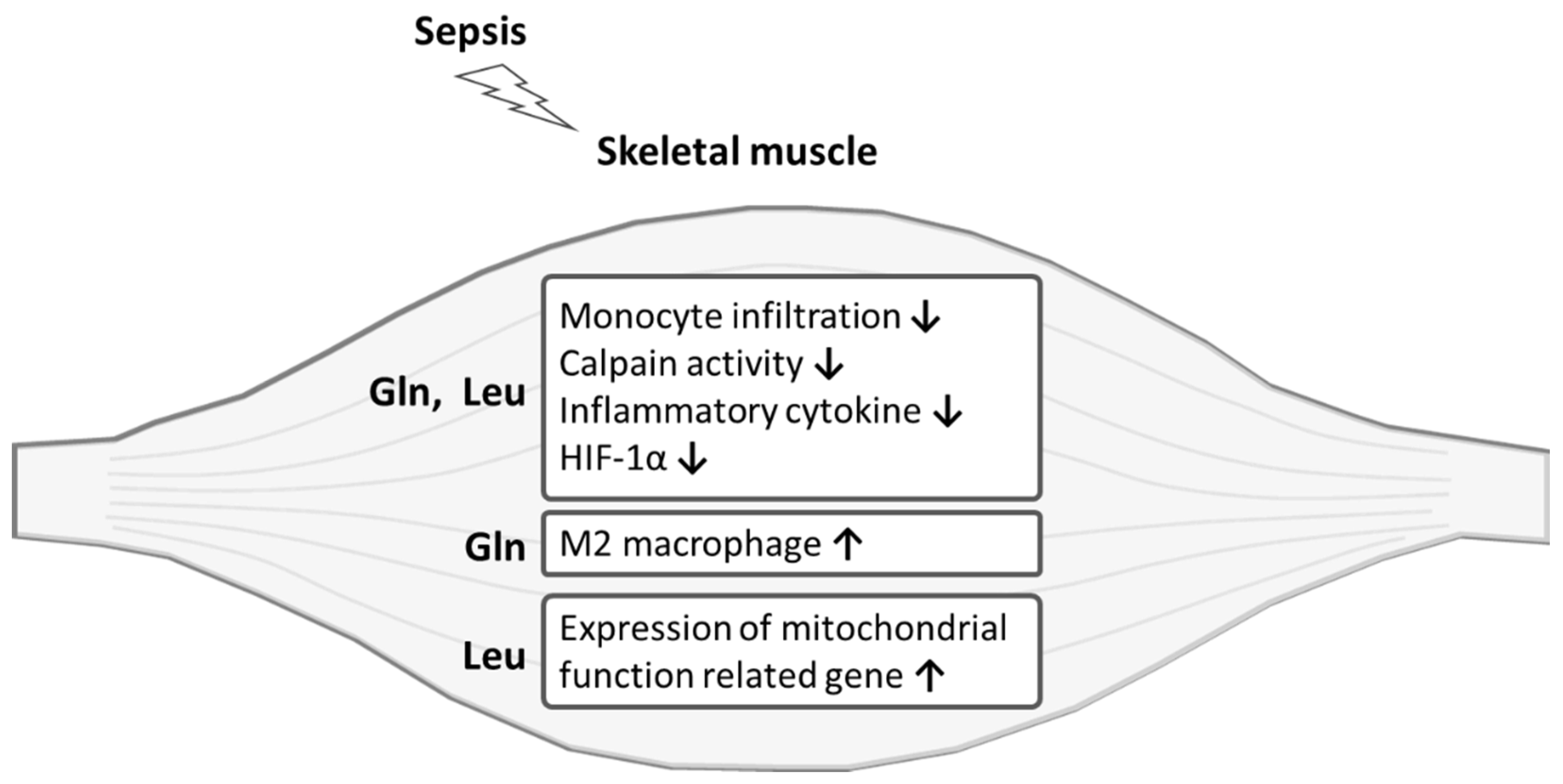
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Hermans, G.; De Jonghe, B.; Bruyninckx, F.; Berghe, G.V. Clinical review: Critical illness polyneuropathy and myopathy. Crit. Care 2008, 12, 238. [Google Scholar] [CrossRef] [Green Version]
- Batt, J.; Herridge, M.S.; dos Santos, C.C. From skeletal muscle weakness to functional outcomes following critical illness: A Translational Biology Perspective. Thorax 2019, 74, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Harrison, T.B.; Rich, M.M. Mechanisms of Neuromuscular Dysfunction in Critical Illness. Crit. Care Clin. 2008, 24, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, L.H. Critical illness polyneuropathy and myopathy: Clinical features, risk factors and prognosis. Eur. J. Neurol. 2006, 13, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, O.; Reid, M.B.; Berghe, G.V.D.; Vanhorebeek, I.; Hermans, G.; Rich, M.M.; Larsson, L. The Sick and the Weak: Neuropathies/Myopathies in the Critically Ill. Physiol. Rev. 2015, 95, 1025–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, L.A.; Supinski, G.S. Sepsis-induced myopathy. Crit. Care Med. 2009, 37, S354–S367. [Google Scholar] [CrossRef] [Green Version]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and immune function, supplementation and clinical translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [Green Version]
- Roth, E. Nonnutritive Effects of Glutamine. J. Nutr. 2008, 138, 2025S–2031S. [Google Scholar] [CrossRef]
- Wischmeyer, P.E. Glutamine: Mode of action in critical illness. Crit. Care Med. 2007, 35, S541–S544. [Google Scholar] [CrossRef] [PubMed]
- Mok, E.; Violante, C.E.-D.; Daubrosse, C.; Gottrand, F.; Rigal, O.; Fontan, J.-E.; Cuisset, J.-M.; Guilhot, J.; Hankard, R. Oral glutamine and amino acid supplementation inhibit whole-body protein degradation in children with Duchenne muscular dystrophy. Am. J. Clin. Nutr. 2006, 83, 823–828. [Google Scholar] [CrossRef]
- Pai, M.-H.; Lei, C.-S.; Su, S.-T.; Yeh, S.-L.; Hou, Y.-C. Effects of dietary glutamine supplementation on immune cell polarization and muscle regeneration in diabetic mice with limb ischemia. Eur. J. Nutr. 2020, 59, 921–933. [Google Scholar] [CrossRef]
- Van Loon, L.J. Leucine as a pharmaconutrient in health and disease. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 71–77. [Google Scholar] [CrossRef]
- Ham, D.J.; Caldow, M.K.; Lynch, G.S.; Koopman, R. Leucine as a treatment for muscle wasting: A critical review. Clin. Nutr. 2014, 33, 937–945. [Google Scholar] [CrossRef]
- Lang, C.H.; Frost, R.A. Differential effect of sepsis on ability of leucine and IGF-I to stimulate muscle translation initiation. Am. J. Physiol. Metab. 2004, 287, E721–E730. [Google Scholar] [CrossRef] [PubMed]
- Hasselgren, P.-O.; James, J.H.; Warner, B.W.; Hummel, R.P.; Fischer, J.E. Protein Synthesis and Degradation in Skeletal Muscle From Septic Rats. Arch. Surg. 1988, 123, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Chen, D.; Yu, B.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; He, J. Leucine Protects Against Skeletal Muscle Atrophy in Lipopolysaccharide-Challenged Rats. J. Med. Food 2017, 20, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Holecek, M. Muscle wasting in animal models of severe illness. Int. J. Exp. Pathol. 2012, 93, 157–171. [Google Scholar] [CrossRef]
- Li, T.; Xiao, G.; Tan, S.; Shi, X.; Yin, L.; Tan, C.; Gu, J.; Liu, Y.; Deng, H.; Liu, K.; et al. HSF1 Attenuates LPS-Induced Acute Lung Injury in Mice by Suppressing Macrophage Infiltration. Oxidative Med. Cell. Longev. 2020, 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [Green Version]
- Rigamonti, E.; Zordan, P.; Sciorati, C.; Rovere-Querini, P.; Brunelli, S. Macrophage Plasticity in Skeletal Muscle Repair. BioMed Res. Int. 2014, 2014, 560629. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Smith, I.J.; Lecker, S.H.; Hasselgren, P.-O. Calpain activity and muscle wasting in sepsis. Am. J. Physiol. Metab. 2008, 295, E762–E771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocheteau, P.; Chatre, L.; Briand, D.; Mebarki, M.; Jouvion, G.; Bardon, J.; Crochemore, C.; Serrani, P.; Lecci, P.P.; Latil, M.; et al. Sepsis induces long-term metabolic and mitochondrial muscle stem cell dysfunction amenable by mesenchymal stem cell therapy. Nat. Commun. 2015, 6, 10145. [Google Scholar] [CrossRef] [Green Version]
- Arulkumaran, N.; Deutschman, C.S.; Pinsky, M.R.; Zuckerbraun, B.; Schumacker, P.T.; Gomez, H.; Gomez, A.; Murray, P.; Kellum, J.A. Mitochondrial Function in Sepsis. Shock 2016, 45, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Hypoxia-inducible factor 1: Regulator of mitochondrial metabolism and mediator of ischemic preconditioning. Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1813, 1263–1268. [Google Scholar] [CrossRef] [Green Version]
- Arnold, R.S.; Shi, J.; Murad, E.; Whalen, A.M.; Sun, C.Q.; Polavarapu, R.; Parthasarathy, S.; Petros, J.A.; Lambeth, J.D. Hydrogen peroxide mediates the cell growth and transformation caused by the mitogenic oxidase Nox1. Proc. Natl. Acad. Sci. USA 2001, 98, 5550–5555. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Bae, I.H.; Bae, Y.S.; Um, H.-D. Link between Mitochondria and NADPH Oxidase 1 Isozyme for the Sustained Production of Reactive Oxygen Species and Cell Death. J. Biol. Chem. 2006, 281, 36228–36235. [Google Scholar] [CrossRef]
- Handschin, C.; Spiegelman, B.M. The role of exercise and PGC1α in inflammation and chronic disease. Nature 2008, 454, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.; Kny, M.; Riediger, F.; Busch, K.; Schmidt, S.; Luft, F.C.; Slevogt, H.; Fielitz, J. Deletion of Nlrp3 protects from inflammation-induced skeletal muscle atrophy. Intensiv. Care Med. Exp. 2017, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Han, W.; Wang, C.; Sui, D.; Bian, J.; Bo, L.; Deng, X. Upregulation of Heme Oxygenase-1 by Hemin Alleviates Sepsis-Induced Muscle Wasting in Mice. Oxidative Med. Cell. Longev. 2018, 2018, 8927104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.; Tacke, R.; Hedrick, C.C.; Hanna, R.N. Nonclassical Patrolling Monocyte Function in the Vasculature. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1306–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, L.; Henry, A.; Poron, F.; Baba-Amer, Y.; Van Rooijen, N.; Plonquet, A.; Gherardi, R.K.; Chazaud, B. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J. Exp. Med. 2007, 204, 1057–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Huang, D.; Saederup, N.; Charo, I.F.; Ransohoff, R.M.; Zhou, L. Macrophages recruited via CCR2 produce insulin-like growth factor-1 to repair acute skeletal muscle injury. FASEB J. 2010, 25, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Dias, I.H.K.; Milic, I.; Heiss, C.; Ademowo, O.S.; Polidori, M.C.; Devitt, A.; Griffiths, H.R. Inflammation, lipid (Per)oxidation, and Redox Regulation. Antioxid. Redox Signal. 2020, 33, 166–190. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and Inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozakowska, M.; Pietraszek-Gremplewicz, K.; Jozkowicz, A.; Dulak, J. The role of oxidative stress in skeletal muscle injury and regeneration: Focus on antioxidant enzymes. J. Muscle Res. Cell Motil. 2015, 36, 377–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, E.; Sestili, P. Reactive Oxygen Species in Skeletal Muscle Signaling. J. Signal Transduct. 2012, 2012, 982794. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, R.A.; Garcia-Smith, R.; Gannon, N.P.; Bisoffi, M.; Trujillo, K.A.; Conn, C.A. Leucine treatment enhances oxidative capacity through complete carbohydrate oxidation and increased mitochondrial density in skeletal muscle cells. Amino Acids 2013, 45, 901–911. [Google Scholar] [CrossRef]
- Anthony, J.C.; Yoshizawa, F.; Anthony, T.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Leucine Stimulates Translation Initiation in Skeletal Muscle of Postabsorptive Rats via a Rapamycin-Sensitive Pathway. J. Nutr. 2000, 130, 2413–2419. [Google Scholar] [CrossRef]
- Liu, Z.; Jahn, L.A.; Wei, L.; Long, W.; Barrett, E.J. Amino Acids Stimulate Translation Initiation and Protein Synthesis through an Akt-Independent Pathway in Human Skeletal Muscle. J. Clin. Endocrinol. Metab. 2002, 87, 5553–5558. [Google Scholar] [CrossRef] [Green Version]
- Kimball, S.R.; Shantz, L.M.; Horetsky, R.L.; Jefferson, L.S. Leucine regulates translation of specific mRNAs in L6 myoblasts through mTOR-mediated changes in availability of eIF4E and phosphorylation of ribosomal protein S6. J. Biol. Chem. 1999, 274, 11647–11652. [Google Scholar] [CrossRef] [Green Version]
- Dardevet, D.; Sornet, C.; Balage, M.; Grizard, J. Stimulation of In Vitro Rat Muscle Protein Synthesis by Leucine Decreases with Age. J. Nutr. 2000, 130, 2630–2635. [Google Scholar] [CrossRef] [PubMed]
- Rittirsch, D.; Huber-Lang, M.S.; A Flierl, M.; A Ward, P. Immunodesign of experimental sepsis by cecal ligation and puncture. Nat. Protoc. 2008, 4, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.-M.; Pai, M.-H.; Yeh, C.-L.; Hou, Y.-C.; Yeh, S.-L. Glutamine administration ameliorates sepsis-induced kidney injury by downregulating the high-mobility group box protein-1-mediated pathway in mice. Am. J. Physiol. Physiol. 2012, 302, F150–F158. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.-M.; Hsiung, Y.-C.; Pai, M.-H.; Yeh, S.-L. Glutamine administration in early or late septic phase downregulates lymphocyte PD-1/PD-L1 expression and the inflammatory response in mice with polymicrobial sepsis. J. Parenter. Enter. Nutr. 2018, 42, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Mok, E.; Constantin, B.; Favreau, F.; Neveux, N.; Magaud, C.; Delwail, A.; Hankard, R. L-Glutamine Administration Reduces Oxidized Glutathione and MAP Kinase Signaling in Dystrophic Muscle of mdx Mice. Pediatr. Res. 2008, 63, 268–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poncet, N.; Mitchell, F.E.; Ibrahim, A.F.M.; McGuire, V.A.; English, G.; Arthur, S.; Shi, Y.-B.; Taylor, P.M. The Catalytic Subunit of the System L1 Amino Acid Transporter (Slc7a5) Facilitates Nutrient Signalling in Mouse Skeletal Muscle. PLoS ONE 2014, 9, e89547. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | S | G | L | GL | |
|---|---|---|---|---|---|
| BW change (%) | −2.73 ± 0.19 | −15.61 ± 0.95 * | −15.19 ± 0.95 | −14.05 ± 0.99 | −15.35 ± 0.88 |
| GA muscle weight (% of BW) | 1.18 ± 0.01 | 1.06 ± 0.03 * | 1.12 ± 0.02 | 1.16 ± 0.02 † | 1.14 ± 0.02 |
| Gene | Primer Sequences (5′ to 3′) | Accession Number |
|---|---|---|
| TNF-α | F: CCCTCACACTCAGATCATCTTCT R: GCTACGACGTGGGCTACAG | NM_013693.3 |
| IL-1β | F: TGCCACCTTTTGACAGTGATG R: ATGTGCTGCTGCGAGATTT | NM_008361.4 |
| IL-6 | F: GGGACTGATGCTGGTGACAA R: ACAGGTCTGTTGGGAGTGGT | NM_001314054.1 |
| PGC-1α | F: CTGCGGGATGATGGAGACAG R: TCGTTCGACCTGCGTAAAGT | NM_008904.2 |
| NOX1 | F: GGAGTGGCATCCCTTCACTC R: GGCATTGGTGAGTGCTGTTG | NM_172203.2 |
| HIF-1α | F: CTGTTATGAGGCTCACCATCAG R: CAGTCCATCTGTGCCTTCATC | NM_001313919.1 |
| HPRT | F: AGCCTAAGATGAGCGCAAGT R: TTACTAGGCAGATGGCCACA | NM_013556.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.-C.; Pai, M.-H.; Wu, J.-M.; Yang, P.-J.; Lee, P.-C.; Chen, K.-Y.; Yeh, S.-L.; Lin, M.-T. Protective Effects of Glutamine and Leucine Supplementation on Sepsis-Induced Skeletal Muscle Injuries. Int. J. Mol. Sci. 2021, 22, 13003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313003
Hou Y-C, Pai M-H, Wu J-M, Yang P-J, Lee P-C, Chen K-Y, Yeh S-L, Lin M-T. Protective Effects of Glutamine and Leucine Supplementation on Sepsis-Induced Skeletal Muscle Injuries. International Journal of Molecular Sciences. 2021; 22(23):13003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313003
Chicago/Turabian StyleHou, Yu-Chen, Man-Hui Pai, Jin-Ming Wu, Po-Jen Yang, Po-Chu Lee, Kuen-Yuan Chen, Sung-Ling Yeh, and Ming-Tsan Lin. 2021. "Protective Effects of Glutamine and Leucine Supplementation on Sepsis-Induced Skeletal Muscle Injuries" International Journal of Molecular Sciences 22, no. 23: 13003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313003