
Reduced Immunosenescence of Peripheral Blood T Cells in Parkinson’s Disease with CMV Infection Background

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Alterations in the Ratio of Peripheral T Cells and NK Cells between PD Patients and HD Group in the Context of CMV Persistence
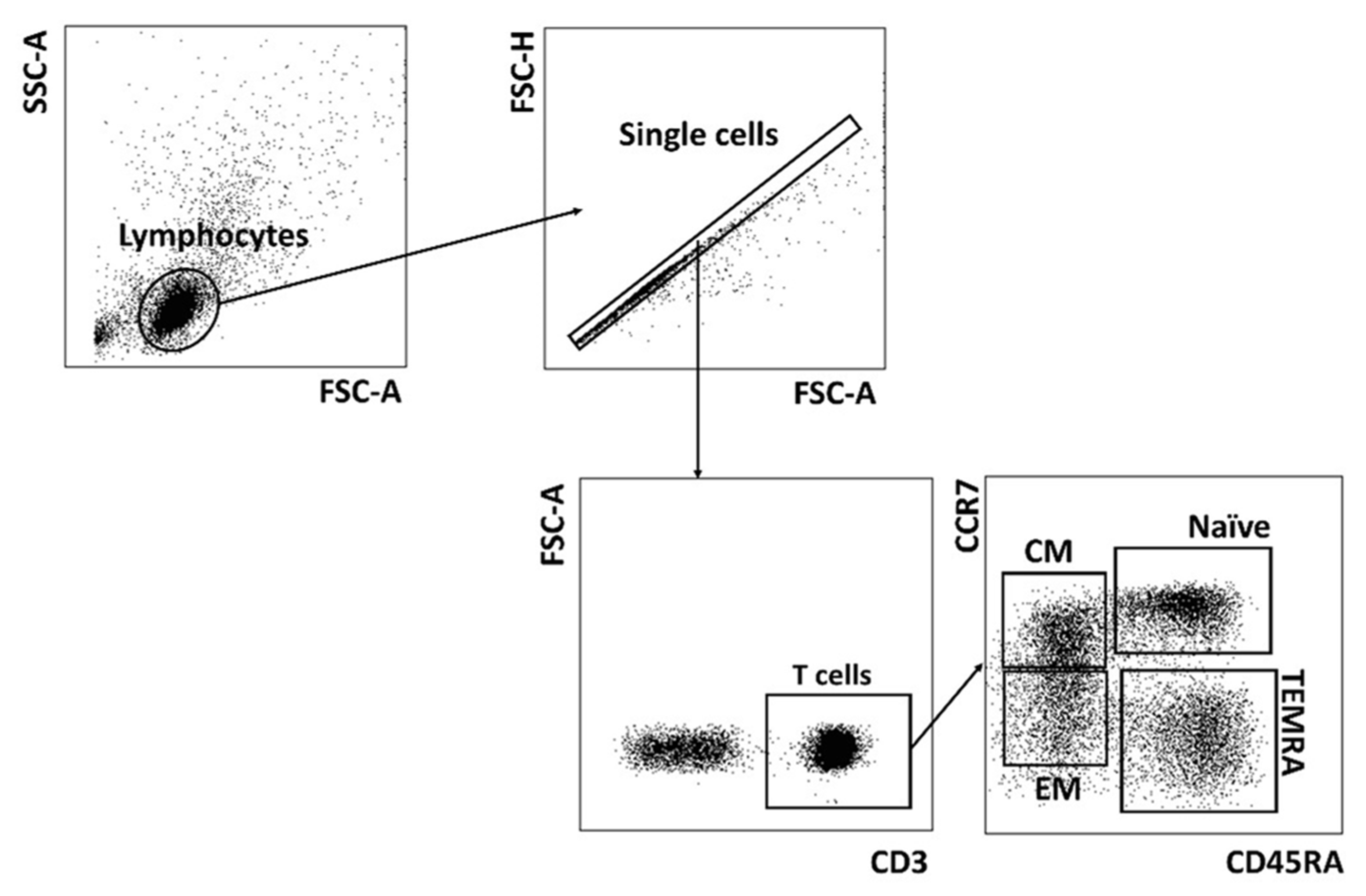
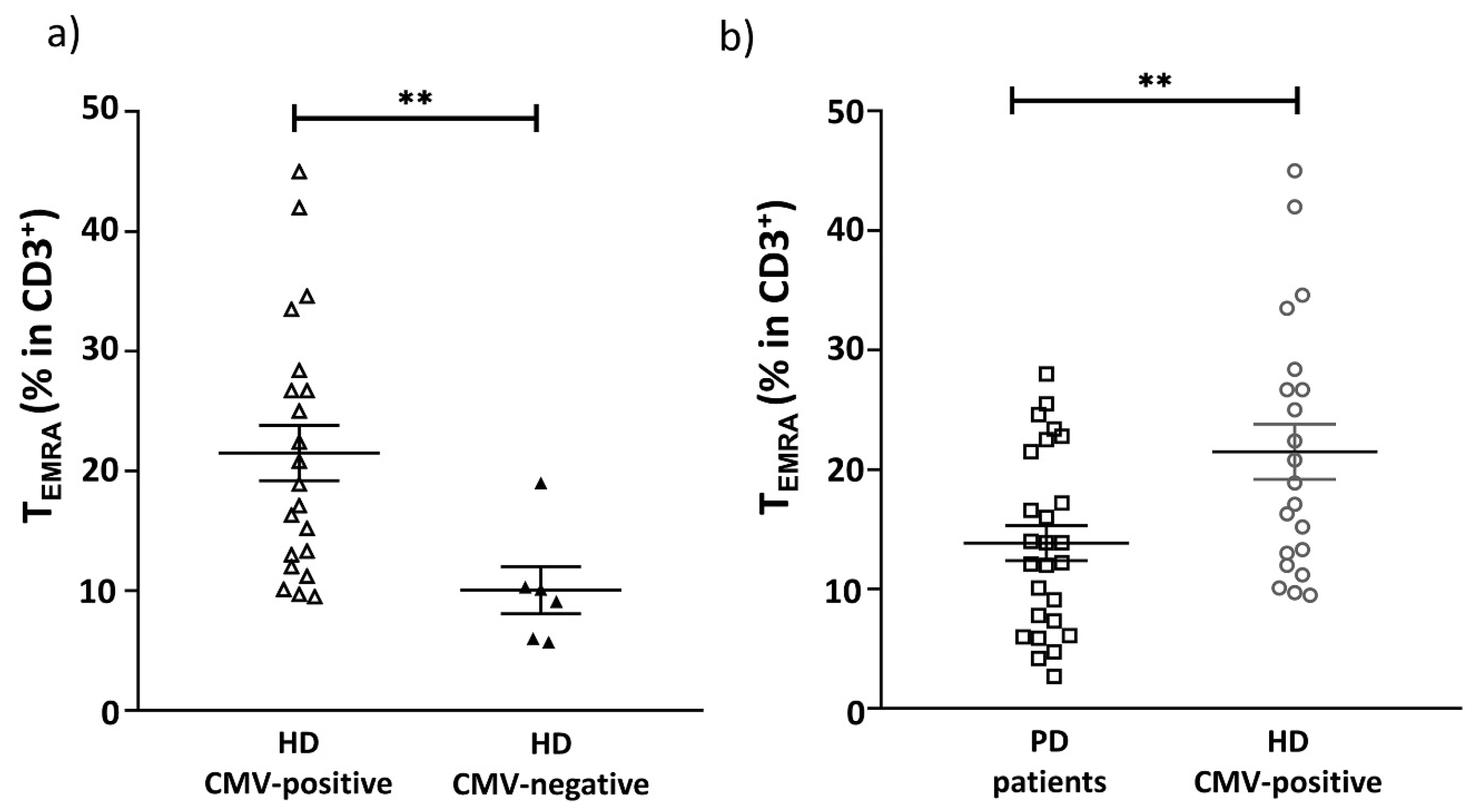
2.2. CMV Infection Accompanies the Alterations in the Repertoire of Differentiated Cells in PD Patients
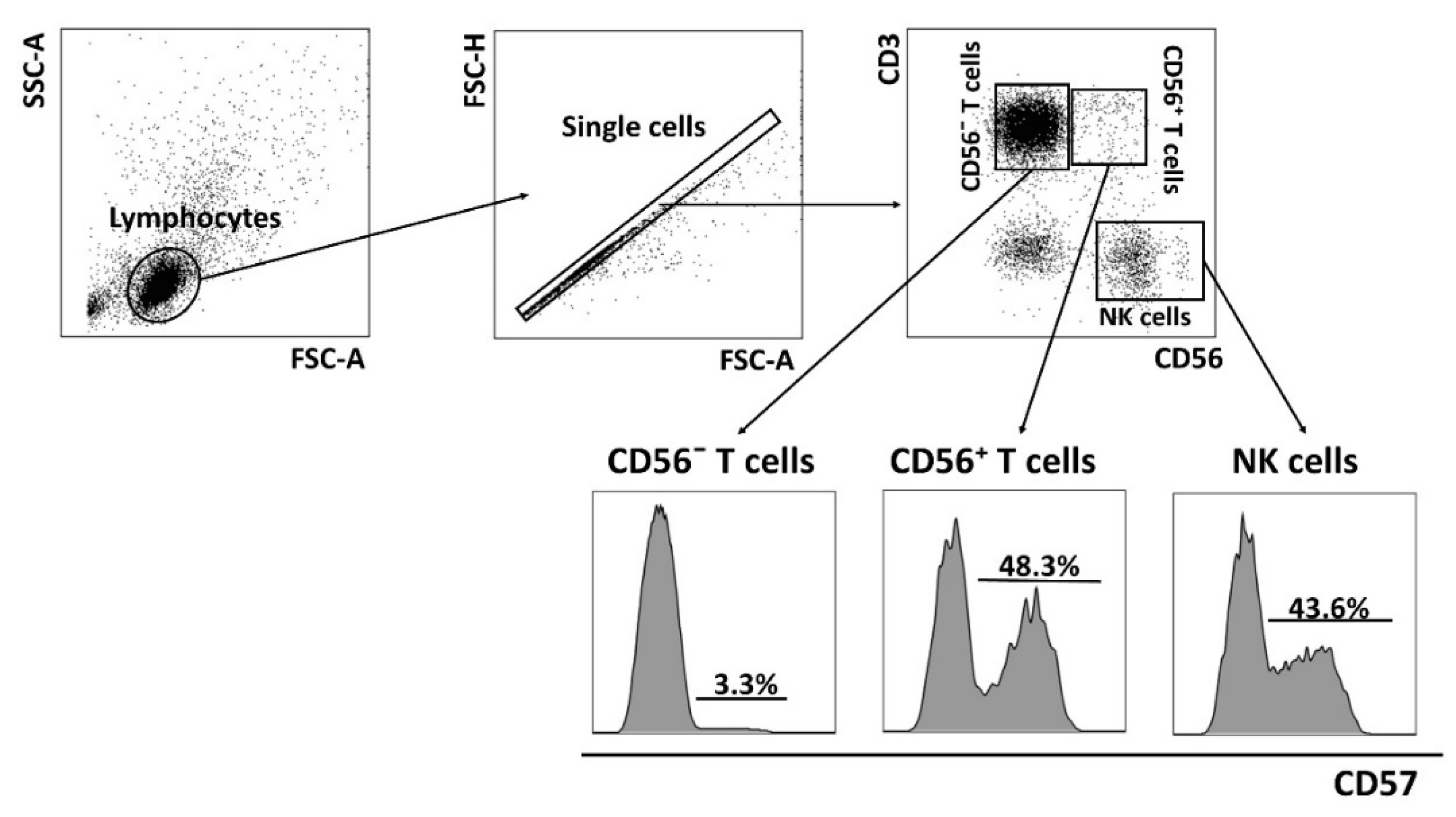
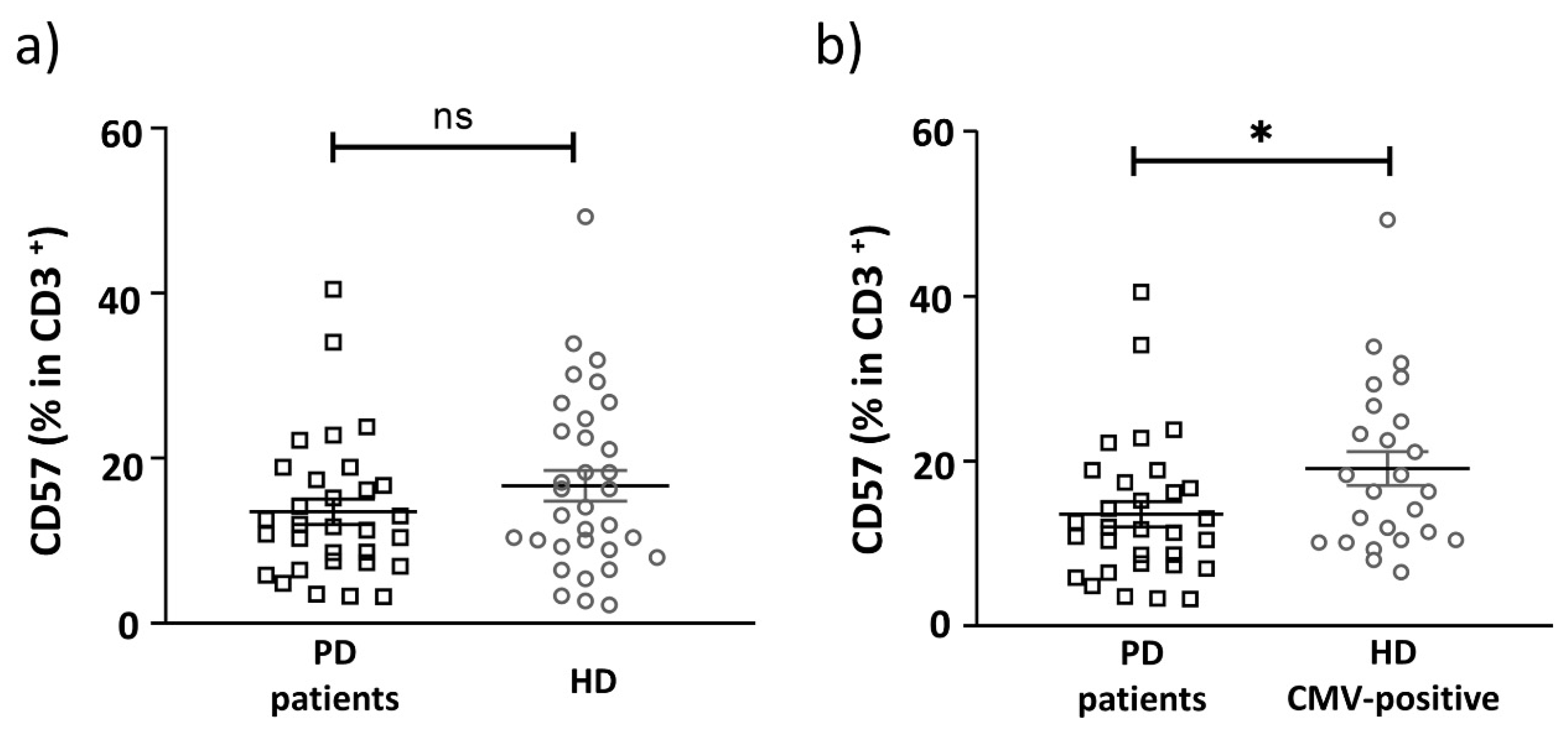
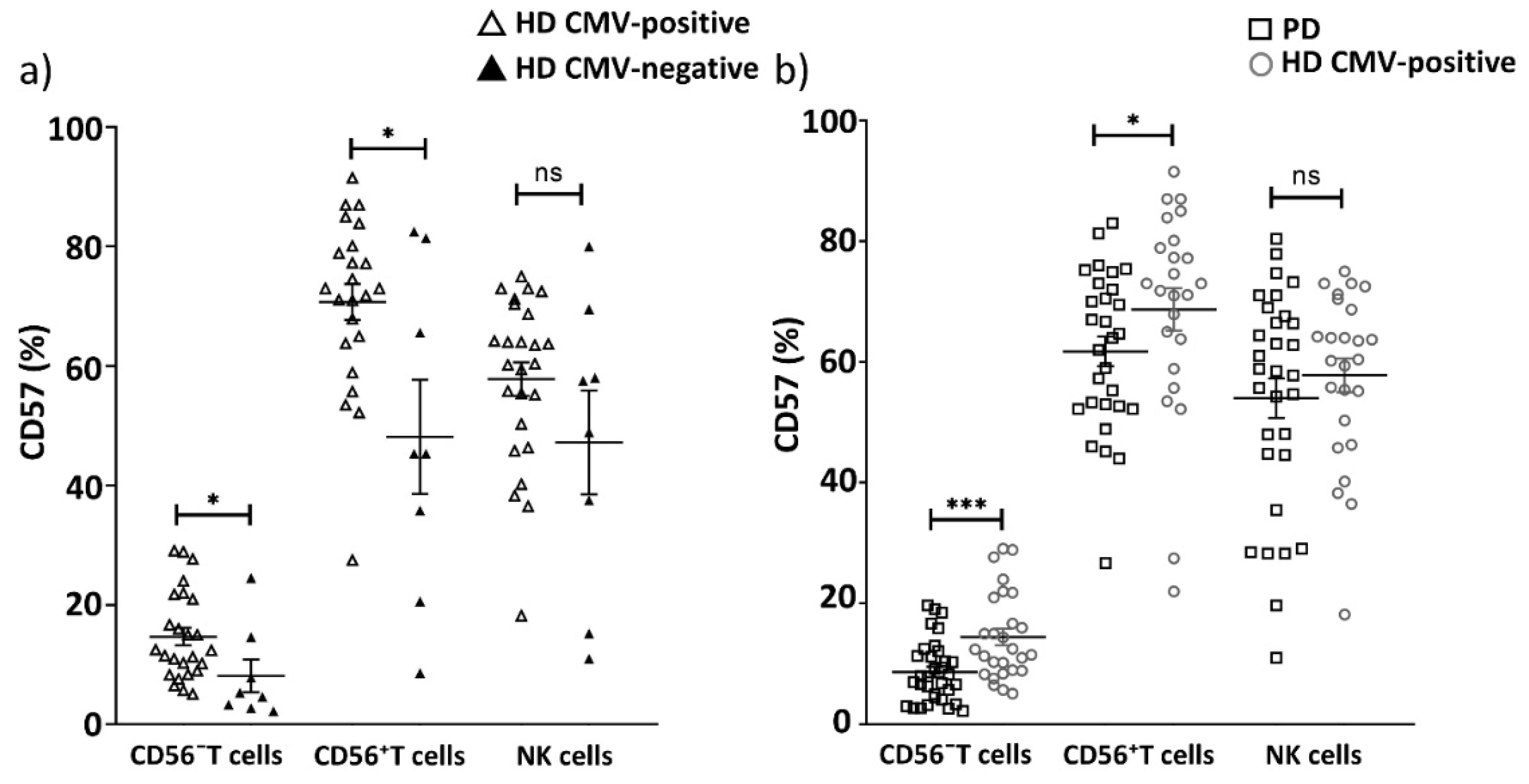
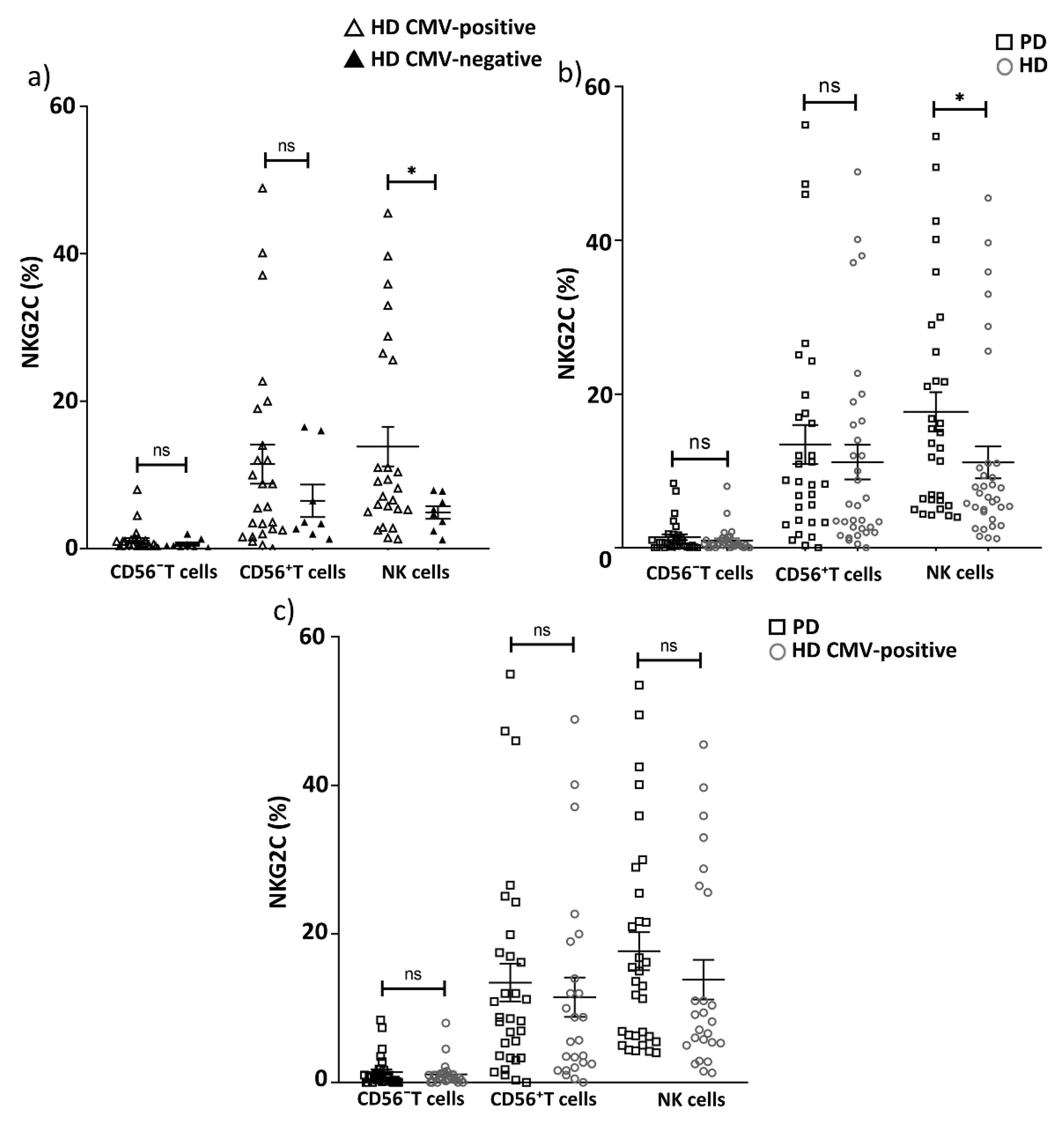
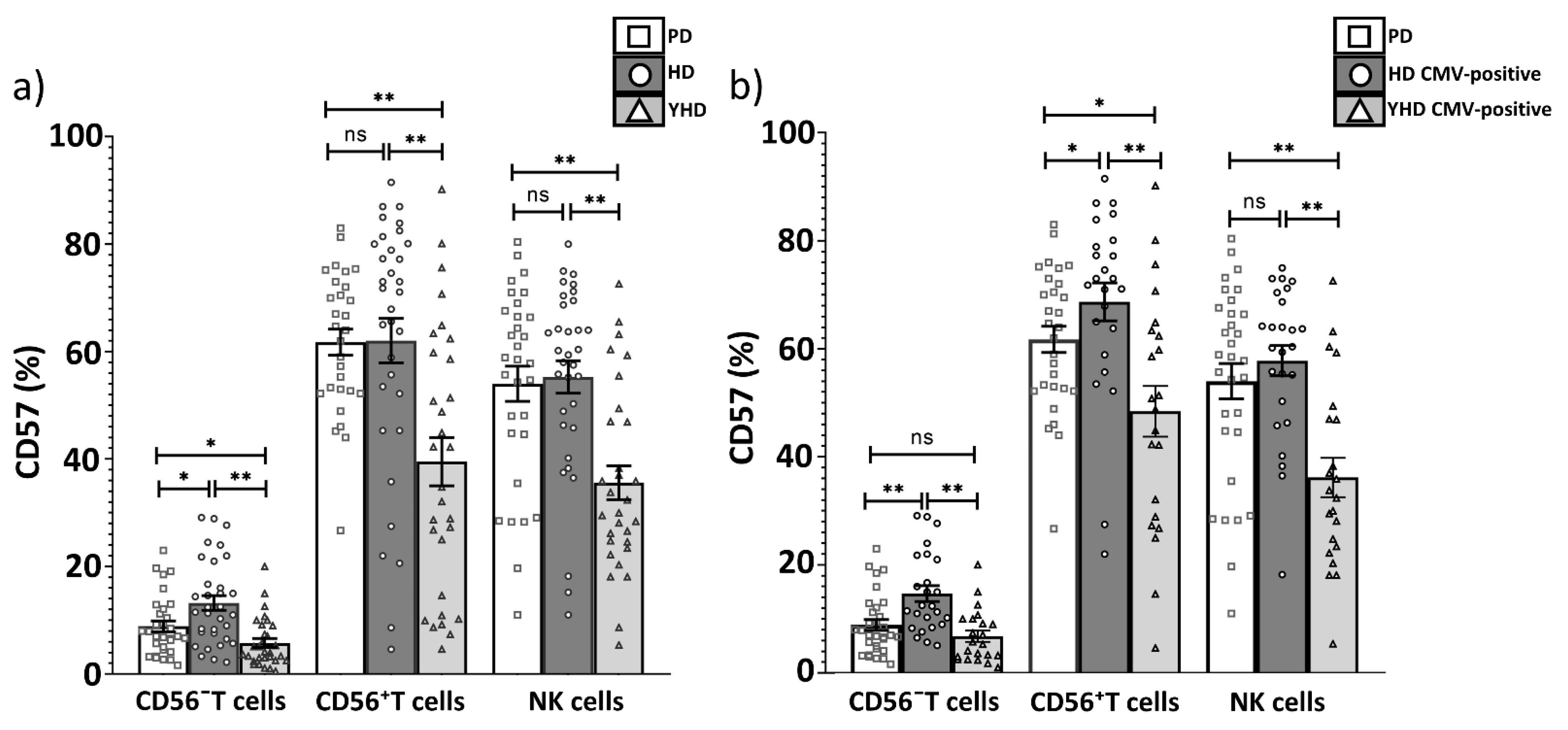
2.3. Replicative Senescence of CD56− and CD56+ T Cells Evaluated by CD57 and NKG2C Expression in PD Patients and CMV-Positive HD Cohorts
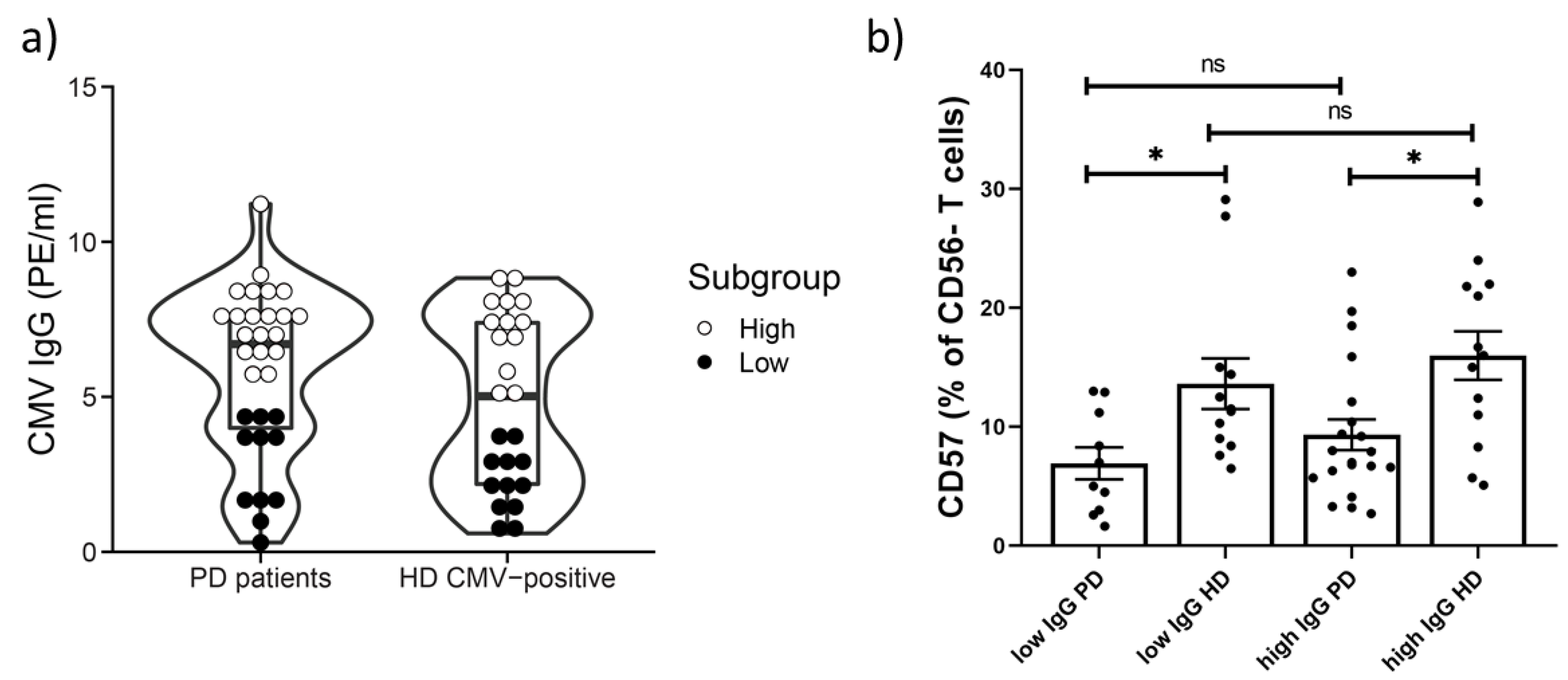
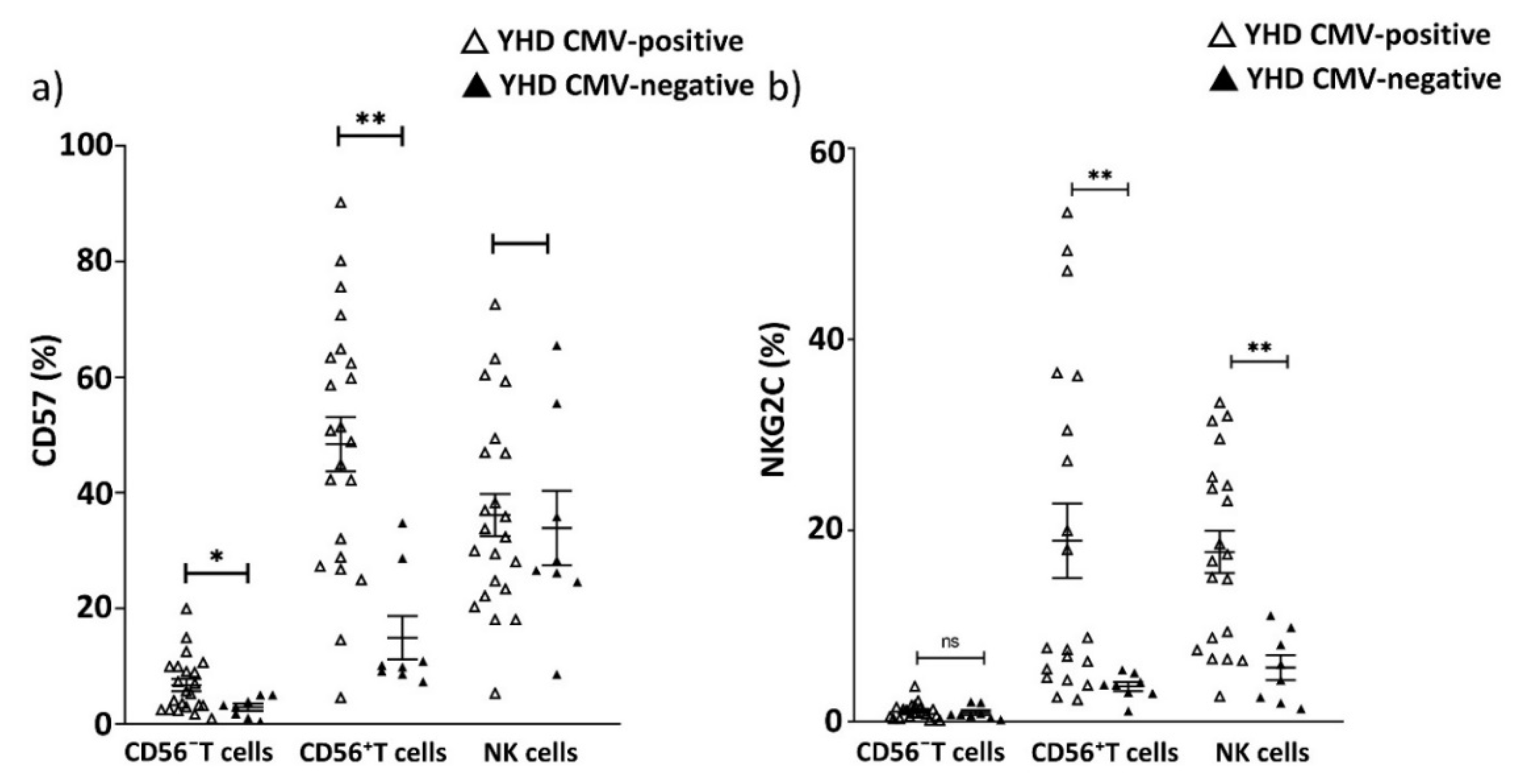
2.4. Age-Related Increase in CD57+CD56− T Cells Subset Not Detected in PD Patients
3. Discussion
4. Materials and Methods
4.1. Participants and Ethics Statement
4.2. Isolation of PBMC from Peripheral Donor Blood and the Sample Preparation
4.3. Antibodies for Flow Cytometry
4.4. Flow Cytometry and Data Analysis
4.5. CMV Serology Status
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Tysnes, O.B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural Transm. 2017, 124, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-J.; Lin, K.-L.; Chen, S.-D.; Liou, C.-W.; Chuang, Y.-C.; Lin, H.-Y.; Lin, T.-K. The Overcrowded Crossroads: Mitochondria, Alpha-Synuclein, and the Endo-Lysosomal System Interaction in Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illarioshkin, S.N.; Shadrina, M.I.; Slominsky, P.A.; Bespalova, E.V.; Zagorovskaya, T.B.; Bagyeva, G.K.; Markova, E.D.; Limborska, S.A.; Ivanova-Smolenskaya, I.A. A common leucine-rich repeat kinase 2 gene mutation in familial and sporadic Parkinson’s disease in Russia. Eur. J. Neurol. 2007, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Prigione, A.; Piazza, F.; Brighina, L.; Begni, B.; Galbussera, A.; DiFrancesco, J.C.; Andreoni, S.; Piolti, R.; Ferrarese, C. Alpha-synuclein nitration and autophagy response are induced in peripheral blood cells from patients with Parkinson disease. Neurosci. Lett. 2010, 477, 6–10. [Google Scholar] [CrossRef]
- Wu, G.; Wang, X.; Feng, X.; Zhang, A.; Li, J.; Gu, K.; Huang, J.; Pang, S.; Dong, H.; Gao, H.; et al. Altered expression of autophagic genes in the peripheral leukocytes of patients with sporadic Parkinson’s disease. Brain Res. 2011, 1394, 105–111. [Google Scholar] [CrossRef]
- Botelho, J.; Lyra, P.; Proença, L.; Godinho, C.; Mendes, J.J.; Machado, V. Relationship between Blood and Standard Biochemistry Levels with Periodontitis in Parkinson’s Disease Patients: Data from the NHANES 2011–2012. J. Pers. Med. 2020, 10, 69. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.M.; Kotzbauer, P.T.; Uryu, K.; Leight, S.; Trojanowski, J.Q.; Lee, V.M.Y. Neuroinflammation and oxidation/nitration of alpha-synuclein linked to dopaminergic neurodegeneration. J. Neurosci. 2008, 28, 7687–7698. [Google Scholar] [CrossRef]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar] [CrossRef]
- Doty, K.R.; Guillot-Sestier, M.V.; Town, T. The role of the immune system in neurodegenerative disorders: Adaptive or maladaptive? Brain Res. 2015, 1617, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal death and implications for therapeutic intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, L.M.; Toulouse, A.; Connor, T.J.; Nolan, Y.M. Contributions of central and systemic inflammation to the pathophysiology of Parkinson’s disease. Neuropharmacology 2012, 62, 2154–2168. [Google Scholar] [CrossRef] [Green Version]
- García-Domínguez, I.; Veselá, K.; García-Revilla, J.; Carrillo-Jiménez, A.; Roca-Ceballos, M.A.; Santiago, M.; de Pablos, R.M.; Venero, J.L. Peripheral Inflammation Enhances Microglia Response and Nigral Dopaminergic Cell Death in an in vivo MPTP Model of Parkinson’s Disease. Front. Cell. Neurosci. 2018, 12, 398. [Google Scholar] [CrossRef] [PubMed]
- Brochard, V.; Combadière, B.; Prigent, A.; Laouar, Y.; Perrin, A.; Beray-Berthat, V.; Bonduelle, O.; Alvarez-Fischer, D.; Callebert, J.; Launay, J.M.; et al. Infiltration of CD4+ lymphocytes into the brain contributes to neurodegeneration in a mouse model of Parkinson disease. J. Clin. Investig. 2009, 119, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Liu, Y.; Zhang, J.; Sun, L.; Liu, Z.; Zhang, S.; Wang, B.; Su, D.; Su, Z. Roles of inflammation response in microglia cell through Toll-like receptors 2/interleukin-23/interleukin-17 pathway in cerebral ischemia/reperfusion injury. Neuroscience 2011, 176, 162–172. [Google Scholar] [CrossRef]
- Perry, V.H. Innate inflammation in Parkinson’s disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Goncharova, L.B.; Tarakanov, A.O. Molecular networks of brain and immunity. Brain Res. Rev. 2007, 55, 155–166. [Google Scholar] [CrossRef]
- Ferrari, C.C.; Tarelli, R. Parkinson’s disease and systemic inflammation. Park. Dis. 2011, 2011, 436813. [Google Scholar] [CrossRef] [Green Version]
- Rentzos, M.; Nikolaou, C.; Andreadou, E.; Paraskevas, G.P.; Rombos, A.; Zoga, M.; Tsoutsou, A.; Boufidou, F.; Kapaki, E.; Vassilopoulos, D. Circulating interleukin-15 and RANTES chemokine in Parkinson’s disease. Acta Neurol. Scand. 2007, 116, 374–379. [Google Scholar] [CrossRef]
- Nagai, Y.; Ueno, S.; Saeki, Y.; Soga, F.; Hirano, M.; Yanagihara, T. Decrease of the D3 dopamine receptor mRNA expression in lymphocytes from patients with Parkinson’s disease. Neurology 1996, 46, 791–795. [Google Scholar] [CrossRef]
- Migliore, L.; Petrozzi, L.; Lucetti, C.; Gambaccini, G.; Bernardini, S.; Scarpato, R.; Trippi, F.; Barale, R.; Frenzilli, G.; Rodilla, V.; et al. Oxidative damage and cytogenetic analysis in leukocytes of Parkinson’s disease patients. Neurology 2002, 58, 1809–1815. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.; Kuroiwa, A.; Uitti, R.J.; Wszolek, Z.K.; Yamada, T. Alterations of T-lymphocyte populations in Parkinson disease. Park. Relat. Disord. 2005, 11, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qi, B.; Xu, W.; Ma, B.; Li, L.; Chen, Q.; Qian, W.; Liu, X.; Qu, H. Clinical correlation of peripheral CD4+-cell sub-sets, their imbalance and Parkinson’s disease. Mol. Med. Rep. 2015, 12, 6105–6111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams-Gray, C.H.; Wijeyekoon, R.S.; Scott, K.M.; Hayat, S.; Barker, R.A.; Jones, J.L. Abnormalities of age-related T cell senescence in Parkinson’s disease. J. Neuroinflam. 2018, 15, 166. [Google Scholar] [CrossRef] [PubMed]
- Ravenholt, R.T.; Foege, W.H. 1918 influenza, encephalitis lethargica, parkinsonism. Lancet 1982, 2, 860–864. [Google Scholar] [CrossRef]
- Peatfield, R.C. Basal ganglia damage and subcortical dementia after possible insidious Coxsackie virus encephalitis. Acta Neurol. Scand. 1987, 76, 340–345. [Google Scholar] [CrossRef]
- Hemling, N.; Röyttä, M.; Rinne, J.; Pöllänen, P.; Broberg, E.; Tapio, V.; Vahlberg, T.; Hukkanen, V. Herpesviruses in brains in Alzheimer’s and Parkinson’s diseases. Ann. Neurol. 2003, 54, 267–271. [Google Scholar] [CrossRef]
- Bu, X.L.; Yao, X.Q.; Jiao, S.S.; Zeng, F.; Liu, Y.H.; Xiang, Y.; Liang, C.R.; Wang, Q.H.; Wang, X.; Cao, H.Y.; et al. A study on the association between infectious burden and Alzheimer’s disease. Eur. J. Neurol. 2015, 22, 1519–1525. [Google Scholar] [CrossRef]
- Deleidi, M.; Isacson, O. Viral and Inflammatory Triggers of Neurodegenerative Diseases. Sci. Transl. Med. 2012, 4, 121ps3. [Google Scholar] [CrossRef] [Green Version]
- Ludlow, M.; Kortekaas, J.; Herden, C.; Hoffmann, B.; Tappe, D.; Trebst, C.; Griffin, D.E.; Brindle, H.E.; Solomon, T.; Brown, A.S.; et al. Neurotropic virus infections as the cause of immediate and delayed neuropathology. Acta Neuropathol. 2015, 131, 159–184. [Google Scholar] [CrossRef] [Green Version]
- Fülöp, T.; Larbi, A.; Pawelec, G. Human T Cell Aging and the Impact of Persistent Viral Infections. Front. Immunol. 2013, 4, 271. [Google Scholar] [CrossRef] [Green Version]
- Pawelec, G.; Larbi, A.; Derhovanessian, E. Senescence of the Human Immune System. J. Comp. Pathol. 2010, 142, S39–S44. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Larbi, A. Markers of T Cell Senescence in Humans. Int. J. Mol. Sci. 2017, 18, 1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennifer, P.C.; Rita, B.E. T Cell Replicative Senescence in Human Aging. Curr. Pharm. Des. 2013, 19, 1680–1698. [Google Scholar] [CrossRef]
- Weltevrede, M.; Eilers, R.; de Melker, H.E.; van Baarle, D. Cytomegalovirus persistence and T-cell immunosenescence in people aged fifty and older: A systematic review. Exp. Gerontol. 2016, 77, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Grutza, R.; Moskorz, W.; Senff, T.; Bäcker, E.; Lindemann, M.; Zimmermann, A.; Uhrberg, M.; Lang, P.A.; Timm, J.; Cosmovici, C. NKG2C pos NK Cells Regulate the Expansion of Cytomegalovirus-Specific CD8 T Cells. J. Immunol. 2020, 204, 2910–2917. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, E.I.; Zvyagin, I.V.; Streltsova, M.A.; Mikelov, A.I.; Erokhina, S.A.; Telford, W.G.; Sapozhnikov, A.M.; Lebedev, Y.B. Surface NKG2C Identifies Differentiated αβT-Cell Clones Expanded in Peripheral Blood. Front. Immunol. 2021, 11, 613882. [Google Scholar] [CrossRef]
- Bayard, C.; Lepetitcorps, H.; Roux, A.; Larsen, M.; Fastenackels, S.; Salle, V.; Vieillard, V.; Marchant, A.; Stern, M.; Boddaert, J.; et al. Coordinated expansion of both memory T cells and NK cells in response to CMV infection in humans. Eur. J. Immunol. 2016, 46, 1168–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, B.I.; Akbar, A.N. Convergence of innate and adaptive immunity during human aging. Front. Immunol. 2016, 7, 445. [Google Scholar] [CrossRef] [Green Version]
- Van Acker, H.H.; Capsomidis, A.; Smits, E.L.; Van Tendeloo, V.F. CD56 in the immune system: More than a marker for cytotoxicity? Front. Immunol. 2017, 8, 892. [Google Scholar] [CrossRef]
- Focosi, D.; Bestagno, M.; Burrone, O.; Petrini, M. CD57 + T lymphocytes and functional immune deficiency. J. Leukoc. Biol. 2010, 87, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Pita-Lopez, M.L.; Gayoso, I.; DelaRosa, O.; Casado, J.G.; Alonso, C.; Muñoz-Gomariz, E.; Tarazona, R.; Solana, R. Effect of ageing on CMV-specific CD8 T cells from CMV seropositive healthy donors. Immun. Ageing 2009, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vergès, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a unique CD57+NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.; Boltz, D.A.; Webster, R.G.; Smeyne, R.J. Viral parkinsonism. Biochim. Biophys. Acta-Mol. Basis Dis. 2009, 1792, 714–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schetters, S.T.T.; Gomez-Nicola, D.; Garcia-Vallejo, J.J.; Van Kooyk, Y. Neuroinflammation: Microglia and T Cells Get Ready to Tango. Front. Immunol. 2018, 8, 1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankowski, R.; Mader, S.; Valdés-Ferrer, S.I. Systemic inflammation and the brain: Novel roles of genetic, molecular, and environmental cues as drivers of neurodegeneration. Front. Cell. Neurosci. 2015, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bachari, S.; Naish, J.H.; Parker, G.J.M.; Emsley, H.C.A.; Parkes, L.M. Blood–Brain Barrier Leakage Is Increased in Parkinson’s Disease. Front. Physiol. 2020, 11, 1636. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.; Kartikasari, A.E.R.; Gorry, P.R.; Flanagan, K.L.; Plebanski, M. Potential Impact of Human Cytomegalovirus Infection on Immunity to Ovarian Tumours and Cancer Progression. Biomedicines 2021, 9, 351. [Google Scholar] [CrossRef]
- Pourgheysari, B.; Khan, N.; Best, D.; Bruton, R.; Nayak, L.; Moss, P.A.H. The Cytomegalovirus-Specific CD4 + T-Cell Response Expands with Age and Markedly Alters the CD4 + T-Cell Repertoire. J. Virol. 2007, 81, 7759–7765. [Google Scholar] [CrossRef] [Green Version]
- Mahnke, Y.D.; Brodie, T.M.; Sallusto, F.; Roederer, M.; Lugli, E. The who’s who of T-cell differentiation: Human memory T-cell subsets. Eur. J. Immunol. 2013, 43, 2797–2809. [Google Scholar] [CrossRef]
- Hassouneh, F.; Goldeck, D.; Pera, A.; van Heemst, D.; Slagboom, P.E.; Pawelec, G.; Solana, R. Functional Changes of T-Cell Subsets with Age and CMV Infection. Int. J. Mol. Sci. 2021, 22, 9973. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, E.; Streltsova, M.; Kanevskiy, L.M.; Erokhina, S.A.; Telford, W.G. Identification of Human Memory-Like NK Cells. Curr. Protoc. Cytom. 2017, 79, 9–50. [Google Scholar] [CrossRef] [PubMed]
- Gumá, M.; Angulo, A.; Vilches, C.; Gómez-Lozano, N.; Malats, N.; López-Botet, M. Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 2004, 104, 3664–3671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Priol, Y.; Puthier, D.; Lécureuil, C.; Combadière, C.; Debré, P.; Nguyen, C.; Combadière, B. High Cytotoxic and Specific Migratory Potencies of Senescent CD8+CD57+ Cells in HIV-Infected and Uninfected Individuals. J. Immunol. 2006, 177, 5145–5154. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Middel, J.; Jansen, G.; Verhoef, J.; Nottet, H. Enhanced expression of fractalkine in HIV-1 associated dementia. J. Neuroimmunol. 2001, 115, 168–175. [Google Scholar] [CrossRef]
- Garretti, F.; Agalliu, D.; Arlehamn, C.S.L.; Sette, A.; Sulzer, D. Autoimmmunity in parkinson’s disease: The role of α:-synuclein-specific T cells. Front. Immunol. 2019, 10, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira Braga, F.A.; Hertoghs, K.M.L.; van Lier, R.A.W.; van Gisbergen, K.P.J.M. Molecular characterization of HCMV-specific immune responses: Parallels between CD8+ T cells, CD4+ T cells, and NK cells. Eur. J. Immunol. 2015, 45, 2433–2445. [Google Scholar] [CrossRef]
- Kared, H.; Martelli, S.; Ng, T.P.; Pender, S.L.F.; Larbi, A. CD57 in human natural killer cells and T-lymphocytes. Cancer Immunol. Immunother. 2016, 65, 441–452. [Google Scholar] [CrossRef]
- Streltsova, M.A.; Erokhina, S.A.; Kanevskiy, L.M.; Lee, D.A.; Telford, W.G.; Sapozhnikov, A.M.; Kovalenko, E.I. Analysis of NK cell clones obtained using interleukin-2 and gene-modified K562 cells revealed the ability of ‘senescent’ NK cells to lose CD57 expression and start expressing NKG2A. PLoS ONE 2018, 13, e0208469. [Google Scholar] [CrossRef]
- Lindestam Arlehamn, C.S.; Dhanwani, R.; Pham, J.; Kuan, R.; Frazier, A.; Rezende Dutra, J.; Phillips, E.; Mallal, S.; Roederer, M.; Marder, K.S.; et al. α-Synuclein-specific T cell reactivity is associated with preclinical and early Parkinson’s disease. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-W.; Yang, S.-Y.; Yang, C.-C.; Chang, C.-W.; Wu, Y.-R. Plasma and Serum Alpha-Synuclein as a Biomarker of Diagnosis in Patients With Parkinson’s Disease. Front. Neurol. 2020, 10, 1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, A.J.; Ben-Shlomo, Y.; Daniel, S.E.; Lees, A.J. What features improve the accuracy of clinical diagnosis in Parkinson’s disease: A clinicopathologic study. Neurology 1992, 42, 1142–1146. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PD Patients n = 31 | HD n = 33 | |
|---|---|---|
| Age (Median) | 59 ± 11.6 | 56 ± 11 |
| Male (n) | 13 | 12 |
| MDS-UPDRS motor score (mean) | 25 ± 8.7 | |
| Hoehn and Yahr (mean) | 2.7 ± 0.6 | |
| Duration of PD (mean years) | 7 ± 0.8 | |
| CMV IgG (% positive) | 100 (31/31) | 76 (25/33) |
| Subset (% of Lymphocytes) | PD Patients n = 31 ① | HD n = 33 ② | CMV-Positive HD n = 25 ③ | p-Value | |
|---|---|---|---|---|---|
| ① vs. ② | ① vs. ③ | ||||
| CD3+ | 70.7 ± 2.1 | 70.6 ± 1.8 | 71.5 ± 2.1 | 0.9 | 0.7 |
| CD3−CD56+ | 13.9 ± 1.7 | 12.1 ± 1.3 | 11.8 ± 1.5 | 0.5 | 0.4 |
| CD3+CD4+CD8− | 66.6 ± 1.9 | 63.5 ± 1.9 | 63.6 ± 2.4 | 0.2 | 0.4 |
| CD3+CD4−CD8+ | 25.5 ± 1.5 | 27.9 ± 1.8 | 28.76 ± 2.3 | 0.3 | 0.3 |
| Subset (% of T Cells) | PD Patients n = 27 ① | HD n = 27 ② | CMV-Positive HD n = 21 ③ | p-Value | |
|---|---|---|---|---|---|
| ① vs. ② | ① vs. ③ | ||||
| CCR7+CD45RA+ (Naïve) | 40.1 ± 2.8 | 37.3 ± 2.7 | 34.5 ± 3.1 | 0.5 | 0.1 |
| CCR7+CD45RA− (TCM) | 29.8 ± 1.9 | 28.3 ± 1.9 | 28.4 ± 1.9 | 0.6 | 0.6 |
| CCR7−CD45RA− (TEM) | 15.0 ± 1.7 | 15.3 ± 1.1 | 15.4 ± 1.3 | 0.5 | 0.8 |
| CCR7−CD45RA+ (TEMRA) | 13.85 ± 1.8 | 18.95 ± 2. | 21.50 ± 2.3 | 0.06 | 0.005 |
| Subsets | PD Patients n = 31 ① | HD n = 33 ② | CMV-Positive HD n = 25 ③ | p-Value | |
|---|---|---|---|---|---|
| ① vs. ② | ① vs. ③ | ||||
| CD56− T cells | 52.4 ± 2.2 | 52.9 ± 2.4 | 53.63 ± 2.7 | 0.9 | 0.6 |
| CD56+ T cells | 3.9 ± 0.5 | 4.1 ± 0.6 | 4.7 ± 0.7 | 0.9 | 0.6 |
| CD57 (% of CD3+ cells) | 13.52 ± 1.5 | 16.7 ± 1.9 | 19.1 ± 2.1 | 0.2 | 0.03 |
| CD57 (% of CD56+ T cells) | 61.8 ± 2.4 | 62.1 ± 4.1 | 68.7 ± 3.5 | 0.3 | 0.03 |
| CD57 (% of NK) | 54.0 ± 3.2 | 55.2 ± 3.0 | 57.8 ± 2.8 | 0.8 | 0.5 |
| NKG2C (% of CD56− T cells) | 1.3 ± 0.4 | 0.9 ± 0.3 | 1.1 ± 0.3 | 0.3 | 0.6 |
| NKG2C (%of CD56+ T cells) | 13.5 ± 2.5 | 11.6 ± 2.3 | 11.5 ± 2.5 | 0.5 | 0.6 |
| NKG2C (% of NK cells) | 17.7 ± 2.6 | 11.1 ± 2.1 | 13.8 ± 2.7 | 0.05 | 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vavilova, J.D.; Boyko, A.A.; Ponomareva, N.V.; Fokin, V.F.; Fedotova, E.Y.; Streltsova, M.A.; Kust, S.A.; Grechikhina, M.V.; Bril, E.V.; Zimnyakova, O.S.; et al. Reduced Immunosenescence of Peripheral Blood T Cells in Parkinson’s Disease with CMV Infection Background. Int. J. Mol. Sci. 2021, 22, 13119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313119
Vavilova JD, Boyko AA, Ponomareva NV, Fokin VF, Fedotova EY, Streltsova MA, Kust SA, Grechikhina MV, Bril EV, Zimnyakova OS, et al. Reduced Immunosenescence of Peripheral Blood T Cells in Parkinson’s Disease with CMV Infection Background. International Journal of Molecular Sciences. 2021; 22(23):13119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313119
Chicago/Turabian StyleVavilova, Julia D., Anna A. Boyko, Natalya V. Ponomareva, Vitaly F. Fokin, Ekaterina Y. Fedotova, Maria A. Streltsova, Sofya A. Kust, Maria V. Grechikhina, Ekaterina V. Bril, Olga S. Zimnyakova, and et al. 2021. "Reduced Immunosenescence of Peripheral Blood T Cells in Parkinson’s Disease with CMV Infection Background" International Journal of Molecular Sciences 22, no. 23: 13119. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313119