
Neuron-Derived Estrogen—A Key Neuromodulator in Synaptic Function and Memory

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
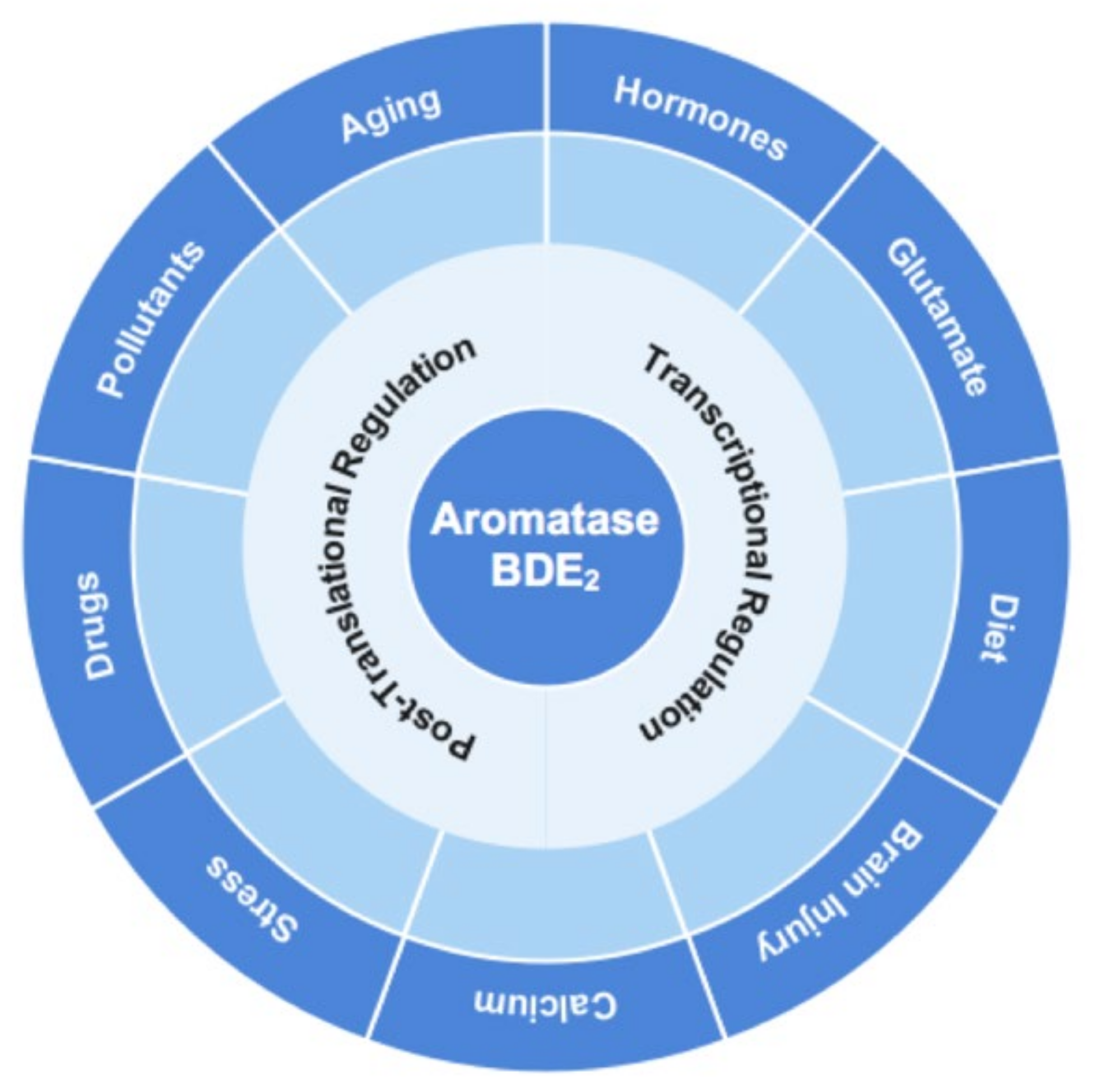
2. Aromatase Localization and Regulation in Neurons
3. NDE2 Is an Important Modulator of Synaptic Plasticity and Cognition
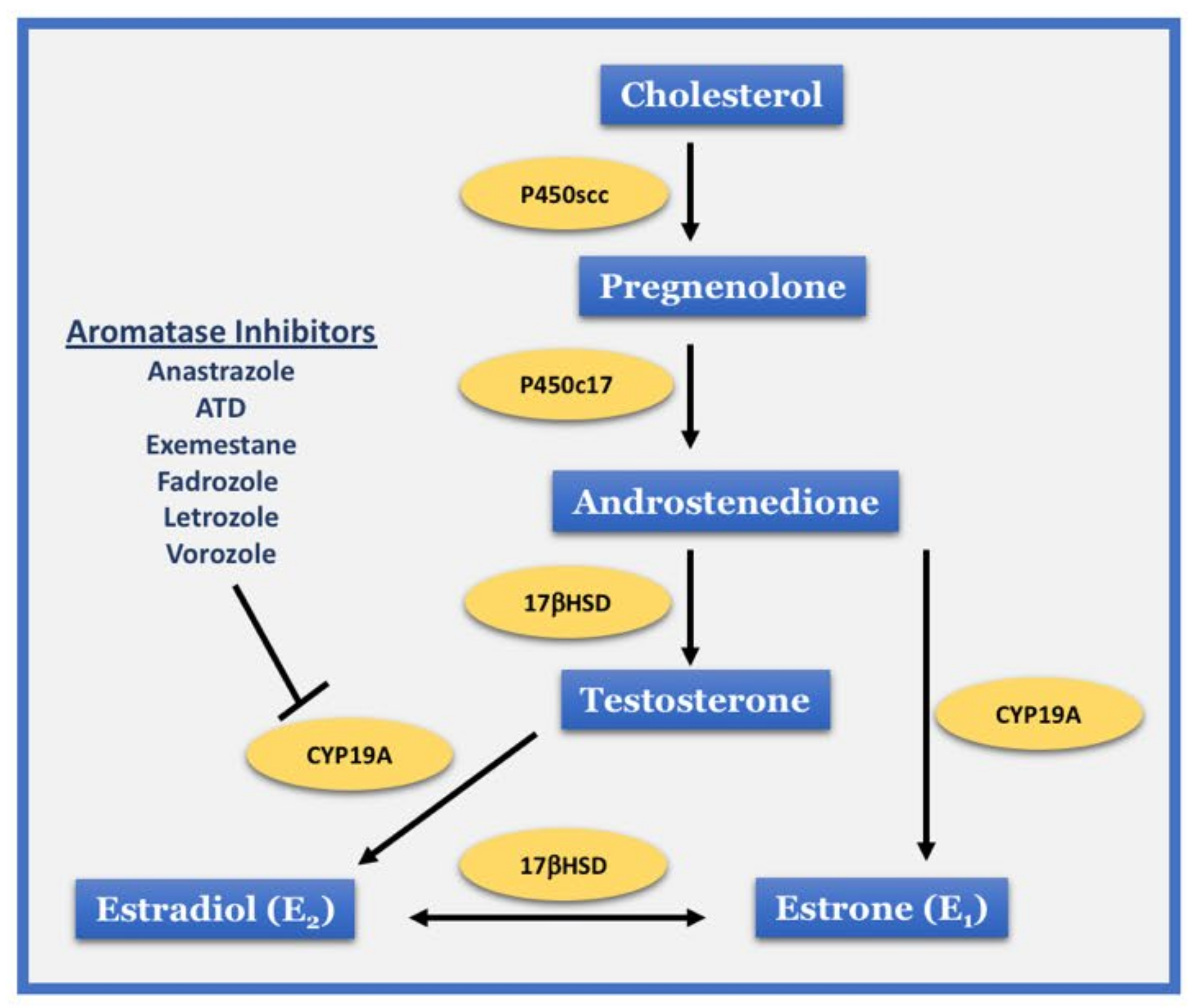
3.1. Evidence from Aromatase Inhibitor Studies
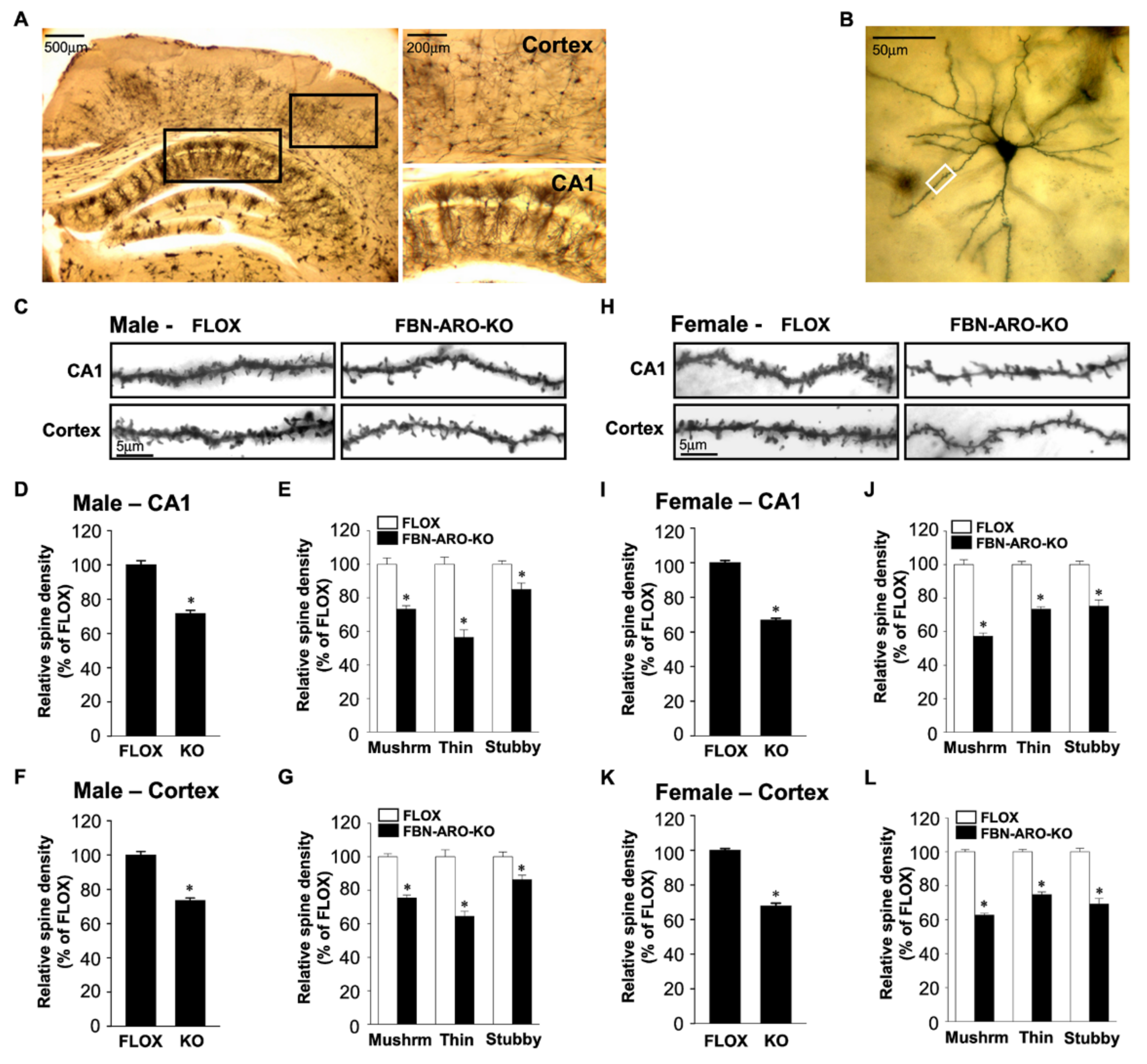
3.2. Evidence from Conditional Knockout Mouse Studies
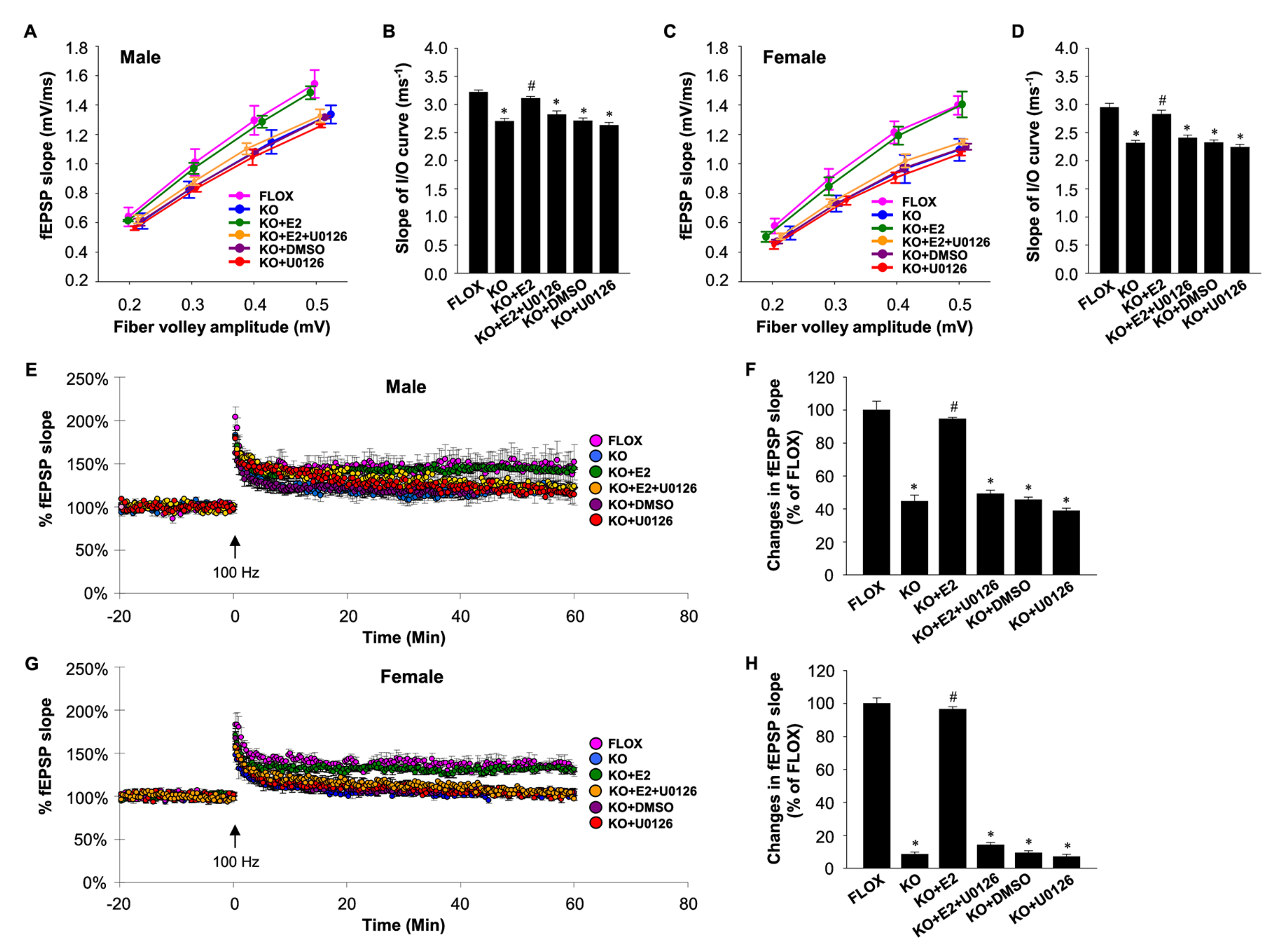
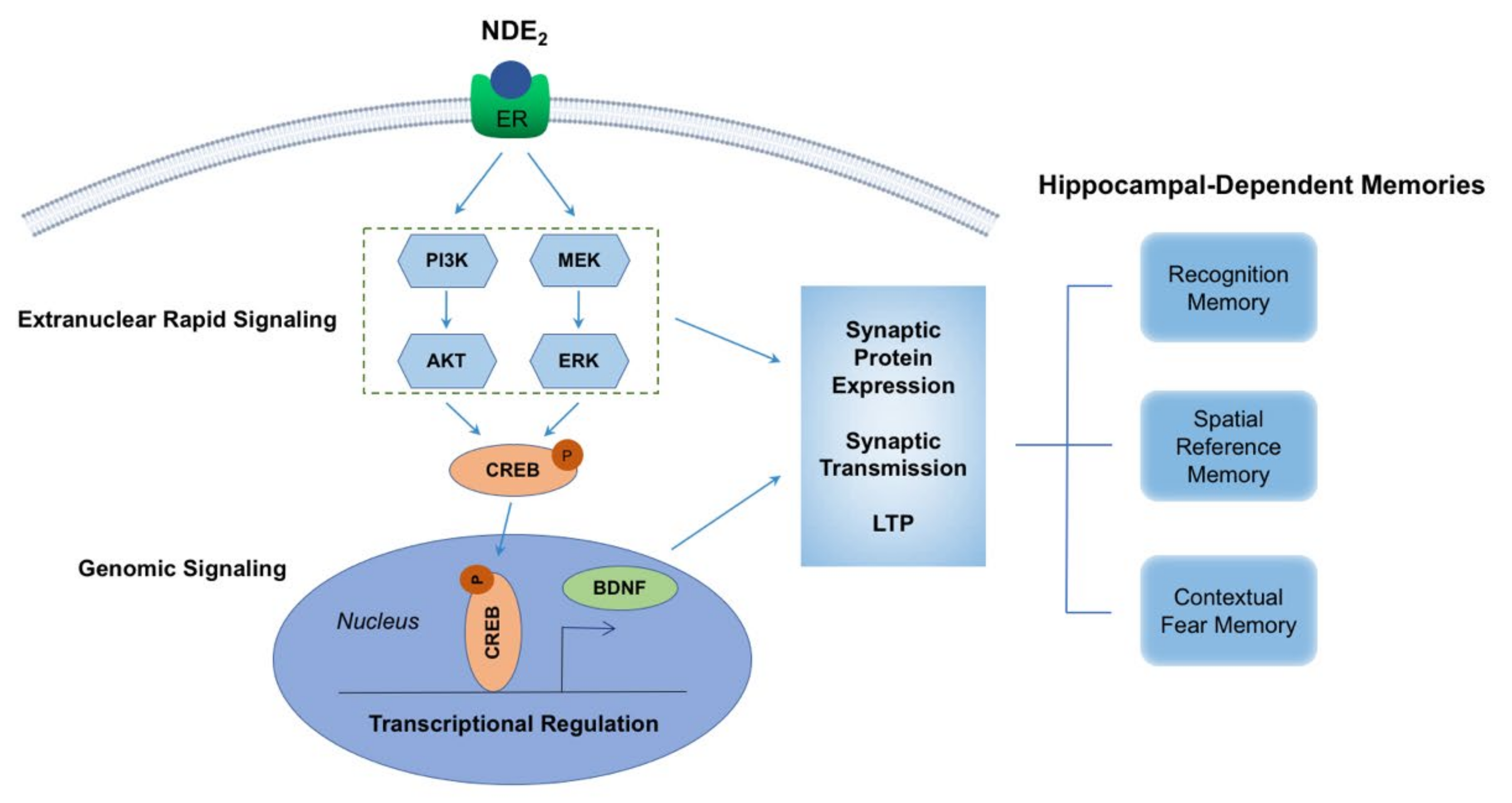
4. Mechanisms Underlying NDE2 Effects on Synaptic Plasticity and Memory
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nelson, L.R.; Bulun, S.E. Estrogen production and action. J. Am. Acad. Dermatol. 2001, 45 (Suppl. 3), S116–S124. [Google Scholar] [CrossRef]
- Spencer, J.L.; Waters, E.M.; Romeo, R.D.; Wood, G.E.; Milner, T.A.; McEwen, B.S. Uncovering the mechanisms of estrogen effects on hippocampal function. Front. Neuroendocrinol. 2008, 29, 219–237. [Google Scholar] [CrossRef] [Green Version]
- Terasawa, E. Neuroestradiol in regulation of GnRH release. Horm. Behav. 2018, 104, 138–145. [Google Scholar] [CrossRef]
- Simpkins, J.W.; Singh, M.; Brock, C.; Etgen, A.M. Neuroprotection and estrogen receptors. Neuroendocrinology 2012, 96, 119–130. [Google Scholar] [CrossRef]
- Brann, D.W.; Dhandapani, K.; Wakade, C.; Mahesh, V.B.; Khan, M.M. Neurotrophic and neuroprotective actions of estrogen: Basic mechanisms and clinical implications. Steroids 2007, 72, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Mahesh, V.B.; Brann, D.W. Neuroendocrine mechanisms underlying the control of gonadotropin secretion by steroids. Steroids 1998, 63, 252–256. [Google Scholar] [CrossRef]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Wang, R.; Li, Y.; Dong, Y.; Zhang, Q.; Liu, J.; O’Connor, J.C.; Xu, J.; et al. Neuron-Derived Estrogen Regulates Synaptic Plasticity and Memory. J. Neurosci. 2019, 39, 2792–2809. [Google Scholar] [CrossRef]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Zhang, Q.; Tang, F.L.; Pratap, U.P.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Neuron-Derived Estrogen Is Critical for Astrocyte Activation and Neuroprotection of the Ischemic Brain. J. Neurosci. 2020, 40, 7355–7374. [Google Scholar] [CrossRef]
- Pedersen, A.L.; Nelson, L.H.; Saldanha, C.J. Centrally Synthesized Estradiol Is a Potent Anti-Inflammatory in the Injured Zebra Finch Brain. Endocrinology 2016, 157, 2041–2051. [Google Scholar] [CrossRef] [Green Version]
- Callard, G.V.; Petro, Z.; Ryan, K.J. Phylogenetic distribution of aromatase and other androgen-converting enzymes in the central nervous system. Endocrinology 1978, 103, 2283–2290. [Google Scholar] [CrossRef]
- Hojo, Y.; Hattori, T.A.; Enami, T.; Furukawa, A.; Suzuki, K.; Ishii, H.T.; Mukai, H.; Morrison, J.H.; Janssen, W.G.; Kominami, S.; et al. Adult male rat hippocampus synthesizes estradiol from pregnenolone by cytochromes P45017alpha and P450 aromatase localized in neurons. Proc. Natl. Acad. Sci. USA 2004, 101, 865–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLusky, N.J.; Naftolin, F.; Goldman-Rakic, P.S. Estrogen formation and binding in the cerebral cortex of the developing rhesus monkey. Proc. Natl. Acad. Sci. USA 1986, 83, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naftolin, F.; Ryan, K.J.; Petro, Z. Aromatization of androstenedione by limbic system tissue from human foetuses. J. Endocrinol. 1971, 51, 795–796. [Google Scholar] [CrossRef]
- Azcoitia, I.; Yague, J.G.; Garcia-Segura, L.M. Estradiol synthesis within the human brain. Neuroscience 2011, 191, 139–147. [Google Scholar] [CrossRef]
- Naftolin, F.; Horvath, T.L.; Jakab, R.L.; Leranth, C.; Harada, N.; Balthazart, J. Aromatase immunoreactivity in axon terminals of the vertebrate brain. An. immunocytochemical study on quail, rat, monkey and human tissues. Neuroendocrinology 1996, 63, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, C.J.; Tuerk, M.J.; Kim, Y.H.; Fernandes, A.O.; Arnold, A.P.; Schlinger, B.A. Distribution and regulation of telencephalic aromatase expression in the zebra finch revealed with a specific antibody. J. Comp. Neurol. 2000, 423, 619–630. [Google Scholar] [CrossRef]
- Stoffel-Wagner, B.; Watzka, M.; Schramm, J.; Bidlingmaier, F.; Klingmuller, D. Expression of CYP19 (aromatase) mRNA in different areas of the human brain. J. Steroid Biochem. Mol. Biol. 1999, 70, 237–241. [Google Scholar] [CrossRef]
- Negri Cesi, P.; Melcangi, R.C.; Celotti, F.; Martini, L. Aromatase activity in cultured brain cells: Difference between neurons and glia. Brain Res. 1992, 589, 327–332. [Google Scholar] [CrossRef]
- Negri Cesi, P.; Melcangi, R.C.; Celotti, F.; Martini, L. Distribution of aromatase activity in cultured neurons and glia cells. J. Steroid Biochem. Mol. Biol. 1993, 44, 637–639. [Google Scholar] [CrossRef]
- Wang, J.; Sareddy, G.R.; Lu, Y.; Pratap, U.P.; Tang, F.; Greene, K.M.; Meyre, P.L.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Astrocyte-Derived Estrogen Regulates Reactive Astrogliosis and is Neuroprotective following Ischemic Brain Injury. J. Neurosci. 2020, 40, 9751–9771. [Google Scholar] [CrossRef]
- Garcia-Segura, L.M.; Wozniak, A.; Azcoitia, I.; Rodriguez, J.R.; Hutchison, R.E.; Hutchison, J.B. Aromatase expression by astrocytes after brain injury: Implications for local estrogen formation in brain repair. Neuroscience 1999, 89, 567–578. [Google Scholar] [CrossRef]
- Pedersen, A.L.; Brownrout, J.L.; Saldanha, C.J. Neuroinflammation and neurosteroidogenesis: Reciprocal modulation during injury to the adult zebra finch brain. Physiol. Behav. 2018, 187, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Biegon, A. In Vivo visualization of aromatase in animals and humans. Front. Neuroendocrinol. 2016, 40, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukai, H.; Tsurugizawa, T.; Ogiue-Ikeda, M.; Murakami, G.; Hojo, Y.; Ishii, H.; Kimoto, T.; Kawato, S. Local neurosteroid production in the hippocampus: Influence on synaptic plasticity of memory. Neuroendocrinology 2006, 84, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Naftolin, F. Brain aromatization of androgens. J. Reprod. Med. 1994, 39, 257–261. [Google Scholar] [PubMed]
- Srivastava, D.P.; Woolfrey, K.M.; Liu, F.; Brandon, N.J.; Penzes, P. Estrogen receptor-b activity modulates synaptic signaling and structure. J. Neurosci. 2010, 30, 13454–13460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, D.; Egbuta, C.; Kanyo, J.E.; Lam, T.T. Phosphorylation of human placental aromatase CYP19A1. Biochem. J. 2019, 476, 3313–3331. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Griswold, J.; Erman, M.; Pangborn, W. Structural basis for androgen specificity and oestrogen synthesis in human aromatase. Nature 2009, 457, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.W.; Shin, I.; Kagawa, N.; Evans, D.B.; Waterman, M.R.; Arteaga, C.L. Aromatase is phosphorylated in situ at serine-118. J. Steroid Biochem. Mol. Biol. 2008, 112, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Hovsepian-Ruby, L. Phosphorylation of Aromatase in the Zebra Finch Brain; University of California: Los Angeles, CA, USA, 2017; p. 19. [Google Scholar]
- Balthazart, J.; Baillien, M.; Ball, G.F. Interactions between kinases and phosphatases in the rapid control of brain aromatase. J. Neuroendocrinol. 2005, 17, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Balthazart, J.; Baillien, M.; Ball, G.F. Phosphorylation processes mediate rapid changes of brain aromatase activity. J. Steroid Biochem. Mol. Biol. 2001, 79, 261–277. [Google Scholar] [CrossRef]
- Balthazart, J.; Baillien, M.; Ball, G.F. Rapid and reversible inhibition of brain aromatase activity. J. Neuroendocrinol. 2001, 13, 63–73. [Google Scholar] [PubMed]
- Fester, L.; Brandt, N.; Windhorst, S.; Prols, F.; Blaute, C.; Rune, G.M. Control. of aromatase in hippocampal neurons. J. Steroid Biochem. Mol. Biol. 2016, 160, 9–14. [Google Scholar] [CrossRef]
- Remage-Healey, L.; Dong, S.; Maidment, N.T.; Schlinger, B.A. Presynaptic control of rapid estrogen fluctuations in the songbird auditory forebrain. J. Neurosci. 2011, 31, 10034–10038. [Google Scholar] [CrossRef] [Green Version]
- Balthazart, J.; Baillien, M.; Ball, G.F. Rapid control of brain aromatase activity by glutamatergic inputs. Endocrinology 2006, 147, 359–366. [Google Scholar] [CrossRef]
- Remage-Healey, L.; Maidment, N.T.; Schlinger, B.A. Forebrain steroid levels fluctuate rapidly during social interactions. Nat. Neurosci. 2008, 11, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.M.; Woolley, C.S. Acute inhibition of neurosteroid estrogen synthesis suppresses status epilepticus in an animal model. Elife 2016, 5, e12917. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Rao, D.; Gibbs, R.B. Effects of Cholinergic Lesions and Cholinesterase Inhibitors on Aromatase and Estrogen Receptor Expression in Different Regions of the Rat Brain. Neuroscience 2018, 384, 203–213. [Google Scholar] [CrossRef]
- Sarachana, T.; Xu, M.; Wu, R.C.; Hu, V.W. Sex. hormones in autism: Androgens and estrogens differentially and reciprocally regulate RORA, a novel candidate gene for autism. PLoS ONE 2011, 6, e17116. [Google Scholar] [CrossRef]
- Honda, S.I.; Harada, N. ARP-1 Regulates the Transcriptional Activity of the Aromatase Gene in the Mouse Brain. Front. Endocrinol. 2020, 11, 306. [Google Scholar] [CrossRef]
- Honda, S.; Kozako, T.; Shimeno, H.; Soeda, S.; Harada, N. LIM-homeodomain transcription factor, Lhx2, is involved in transcriptional control of brain-specific promoter/exon 1f of the mouse aromatase gene. J. Neuroendocrinol. 2012, 24, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Sakurai, H.; Oguro, A.; Tsuji, M.; Vogel, C.F.A.; Yamazaki, T. Retinoid X receptor-mediated neuroprotection via CYP19 upregulation and subsequent increases in estradiol synthesis. J. Steroid Biochem. Mol. Biol. 2019, 193, 105421. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.B.; Wolfe, A.; Cheng, Y.H.; Glidewell-Kenney, C.; Jameson, J.L.; Bulun, S.E. Aromatase promoter I.f is regulated by estrogen receptor alpha (ESR1) in mouse hypothalamic neuronal cell lines. Biol. Reprod. 2009, 81, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Abdelgadir, S.E.; Resko, J.A.; Ojeda, S.R.; Lephart, E.D.; McPhaul, M.J.; Roselli, C.E. Androgens regulate aromatase cytochrome P450 messenger ribonucleic acid in rat brain. Endocrinology 1994, 135, 395–401. [Google Scholar] [CrossRef]
- Harada, N.; Yamada, K.; Foidart, A.; Balthazart, J. Regulation of aromatase cytochrome P-450 (estrogen synthetase) transcripts in the quail brain by testosterone. Brain Res. Mol. Brain Res. 1992, 15, 19–26. [Google Scholar] [CrossRef]
- Roselli, C.E.; Ellinwood, W.E.; Resko, J.A. Regulation of brain aromatase activity in rats. Endocrinology 1984, 114, 192–200. [Google Scholar] [CrossRef]
- Roselli, C.E.; Resko, J.A. Androgens regulate brain aromatase activity in adult male rats through a receptor mechanism. Endocrinology 1984, 114, 2183–2189. [Google Scholar] [CrossRef]
- Vockel, A.; Prove, E.; Balthazart, J. Effects of castration and testosterone treatment on the activity of testosterone-metabolizing enzymes in the brain of male and female zebra finches. J. NeuroBiol. 1990, 21, 808–825. [Google Scholar] [CrossRef]
- Roselli, C.E.; Resko, J.A. Testosterone regulates aromatase activity in discrete brain areas of male rhesus macaques. Biol. Reprod. 1989, 40, 929–934. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.C.; Zhao, H.; Yilmaz, M.B.; Coon, V.J.; Bulun, S.E. Glucocorticoid-induction of hypothalamic aromatase via its brain-specific promoter. Mol. Cell. Endocrinol. 2012, 362, 85–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Hu, P.; Qi, X.R.; Meng, F.T.; Kalsbeek, A.; Zhou, J.N. Acute restraint stress increases intrahypothalamic oestradiol concentrations in conjunction with increased hypothalamic oestrogen receptor beta and aromatase mRNA expression in female rats. J. Neuroendocrinol. 2011, 23, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Marbouti, L.; Zahmatkesh, M.; Riahi, E.; Sadr, S.S. Inhibition of brain 17beta-estradiol synthesis by letrozole induces cognitive decline in male and female rats. NeuroBiol. Learn. Mem. 2020, 175, 107300. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.S.; Black, K.L.; Daniel, J.M. Circulating Estradiol Regulates Brain-Derived Estradiol via Actions at GnRH Receptors to Impact Memory in Ovariectomized Rats. eNeuro 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Prange-Kiel, J.; Dudzinski, D.A.; Prols, F.; Glatzel, M.; Matschke, J.; Rune, G.M. Aromatase Expression in the Hippocampus of AD Patients and 5xFAD Mice. Neural Plast. 2016, 2016, 9802086. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, R.; Faria, A.; Mateus, N.; Calhau, C.; Azevedo, I. Red wine interferes with oestrogen signalling in rat hippocampus. J. Steroid Biochem. Mol. Biol. 2008, 111, 74–79. [Google Scholar] [CrossRef]
- Lephart, E.D.; Adlercreutz, H.; Lund, T.D. Dietary soy phytoestrogen effects on brain structure and aromatase in Long-Evans rats. Neuroreport 2001, 12, 3451–3455. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.S.; Setchell, K.D.; Lephart, E.D. Maternal and perinatal brain aromatase: Effects of dietary soy phytoestrogens. Brain Res. Dev. Brain Res. 2001, 126, 217–221. [Google Scholar] [CrossRef]
- Biegon, A.; Alexoff, D.L.; Kim, S.W.; Logan, J.; Pareto, D.; Schlyer, D.; Wang, G.J.; Fowler, J.S. Aromatase imaging with [N-methyl-11C]vorozole PET in healthy men and women. J. Nucl. Med. 2015, 56, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Chamniansawat, S.; Sawatdiyaphanon, C. Age-Related Memory Impairment Associated With Decreased Endogenous Estradiol in the Hippocampus of Female Rats. Int. J. Toxicol. 2018, 37, 207–215. [Google Scholar] [CrossRef]
- Zhao, Y.; He, L.; Zhang, Y.; Zhao, J.; Liu, Z.; Xing, F.; Liu, M.; Feng, Z.; Li, W.; Zhang, J. Estrogen receptor alpha and beta regulate actin polymerization and spatial memory through an SRC-1/mTORC2-dependent pathway in the hippocampus of female mice. J. Steroid Biochem. Mol. Biol. 2017, 174, 96–113. [Google Scholar] [CrossRef] [PubMed]
- Von Ziegler, N.I.; Schlumpf, M.; Lichtensteiger, W. Prenatal nicotine exposure selectively affects perinatal forebrain aromatase activity and fetal adrenal function in male rats. Brain Res. Dev. Brain Res. 1991, 62, 23–31. [Google Scholar] [CrossRef]
- Biegon, A.; Alia-Klein, N.; Fowler, J.S. Potential contribution of aromatase inhibition to the effects of nicotine and related compounds on the brain. Front. Pharm. 2012, 3, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biegon, A.; Kim, S.W.; Logan, J.; Hooker, J.M.; Muench, L.; Fowler, J.S. Nicotine blocks brain estrogen synthase (aromatase): In Vivo positron emission tomography studies in female baboons. Biol. Psychiatry 2010, 67, 774–777. [Google Scholar] [CrossRef] [Green Version]
- Bogus, K.; Palasz, A.; Suszka-Switek, A.; Worthington, J.J.; Krzystanek, M.; Wiaderkiewicz, R. Chronic Antipsychotic Treatment Modulates Aromatase (CYP19A1) Expression in the Male Rat Brain. J. Mol. Neurosci. 2019, 68, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Bogus, K.; Zarczynska, M.; Palasz, A.; Suszka-Switek, A.; Worthington, J.J.; Krzystanek, M.; Zarczynski, P. Antipsychotics increase steroidogenic enzyme gene expression in the rat brainstem. Mol. Biol. Rep. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Dzyubenko, E.; Juckel, G.; Faissner, A. The antipsychotic drugs olanzapine and haloperidol modify network connectivity and spontaneous activity of neural networks in vitro. Sci. Rep. 2017, 7, 11609. [Google Scholar] [CrossRef] [Green Version]
- Munetsuna, E.; Hattori, M.; Yamazaki, T. Stimulation of estradiol biosynthesis by tributyltin in rat hippocampal slices. Endocr. Res. 2014, 39, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Wong, T.Y.; Wang, Y.; Huang, J.; Leung, L.K. CYP19 expression is induced by 2,3,7,8-tetrachloro-dibenzo-para-dioxin in human glioma cells. Mol. Cell. Endocrinol. 2013, 375, 106–112. [Google Scholar] [CrossRef]
- Chung, E.; Genco, M.C.; Megrelis, L.; Ruderman, J.V. Effects of bisphenol A and triclocarban on brain-specific expression of aromatase in early zebrafish embryos. Proc. Natl. Acad. Sci. USA 2011, 108, 17732–17737. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Segura, L.M.; Veiga, S.; Sierra, A.; Melcangi, R.C. Aromatase: A neuroprotective enzyme. Prog. NeuroBiol. 2003, 71, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, K.A.; Saldanha, C.J. Central aromatization: A dramatic and responsive defense against threat and trauma to the vertebrate brain. Front. Neuroendocrinol. 2020, 56, 100816. [Google Scholar] [CrossRef] [PubMed]
- Kretz, O.; Fester, L.; Wehrenberg, U.; Zhou, L.; Brauckmann, S.; Zhao, S.; Prange-Kiel, J.; Naumann, T.; Jarry, H.; Frotscher, M.; et al. Hippocampal synapses depend on hippocampal estrogen synthesis. J. Neurosci. 2004, 24, 5913–5921. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Fester, L.; Haghshenas, S.; de Vrese, X.; von Hacht, R.; Gloger, S.; Brandt, N.; Bader, M.; Vollmer, G.; Rune, G.M. Oestradiol-induced synapse formation in the female hippocampus: Roles of oestrogen receptor subtypes. J. Neuroendocrinol. 2014, 26, 439–447. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, Y.; Zhang, Y.; He, L.; Qiu, L.; Zhao, J.; Liu, M.; Zhang, J. Letrozole regulates actin cytoskeleton polymerization dynamics in a SRC-1 dependent manner in the hippocampus of mice. J. Steroid Biochem. Mol. Biol. 2017, 167, 86–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.K.; Boridy, S.; McKinney, R.A.; Maysinger, D. Letrozole Potentiates Mitochondrial and Dendritic Spine Impairments Induced by beta Amyloid. J. Aging Res. 2013, 2013, 538979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fester, L.; Prange-Kiel, J.; Zhou, L.; Blittersdorf, B.V.; Bohm, J.; Jarry, H.; Schumacher, M.; Rune, G.M. Estrogen-regulated synaptogenesis in the hippocampus: Sexual dimorphism in vivo but not in vitro. J. Steroid Biochem. Mol. Biol. 2012, 131, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Fester, L.; Haghshenas, S.; de Vrese, X.; von Hacht, R.; Gloger, S.; Brandt, N.; Bader, M.; Vollmer, G.; Rune, G.M. Aromatase inhibitors induce spine synapse loss in the hippocampus of ovariectomized mice. Endocrinology 2010, 151, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Bian, C.; Liu, M.; Zhao, Y.; Sun, T.; Xing, F.; Zhang, J. Orchiectomy and letrozole differentially regulate synaptic plasticity and spatial memory in a manner that is mediated by SRC-1 in the hippocampus of male mice. J. Steroid Biochem. Mol. Biol. 2018, 178, 354–368. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, M.; Tozzi, A.; Calabresi, P.; Pettorossi, V.E.; Grassi, S. Neo-synthesis of estrogenic or androgenic neurosteroids determine whether long-term potentiation or depression is induced in hippocampus of male rat. Front. Cell. Neurosci. 2015, 9, 376. [Google Scholar] [CrossRef] [Green Version]
- Di Mauro, M.; Tozzi, A.; Calabresi, P.; Pettorossi, V.E.; Grassi, S. Different synaptic stimulation patterns influence the local androgenic and estrogenic neurosteroid availability triggering hippocampal synaptic plasticity in the male rat. Eur. J. Neurosci. 2017, 45, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Grassi, S.; Tozzi, A.; Costa, C.; Tantucci, M.; Colcelli, E.; Scarduzio, M.; Calabresi, P.; Pettorossi, V.E. Neural 17beta-estradiol facilitates long-term potentiation in the hippocampal CA1 region. Neuroscience 2011, 192, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Vierk, R.; Brandt, N.; Rune, G.M. Hippocampal estradiol synthesis and its significance for hippocampal synaptic stability in male and female animals. Neuroscience 2014, 274, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, A.; de Iure, A.; Tantucci, M.; Durante, V.; Quiroga-Varela, A.; Giampa, C.; Di Mauro, M.; Mazzocchetti, P.; Costa, C.; Di Filippo, M. Endogenous 17beta-estradiol is required for activity-dependent long-term potentiation in the striatum: Interaction with the dopaminergic system. Front. Cell. Neurosci. 2015, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Dieni, C.V.; Ferraresi, A.; Sullivan, J.A.; Grassi, S.; Pettorossi, V.E.; Panichi, R. Acute inhibition of estradiol synthesis impacts vestibulo-ocular reflex adaptation and cerebellar long-term potentiation in male rats. Brain Struct. Funct. 2018, 223, 837–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.E.; Tierney, M.C.; Wu, W.; Pritchard, K.I.; Rochon, P.A. Endocrine treatment-associated cognitive impairment in breast cancer survivors: Evidence from published studies. Breast Cancer Res. Treat. 2016, 158, 407–420. [Google Scholar] [CrossRef]
- Underwood, E.A.; Rochon, P.A.; Moineddin, R.; Lee, P.E.; Wu, W.; Pritchard, K.I.; Tierney, M.C. Cognitive sequelae of endocrine therapy in women treated for breast cancer: A meta-analysis. Breast Cancer Res. Treat. 2018, 168, 299–310. [Google Scholar] [CrossRef]
- Bayer, J.; Rune, G.; Schultz, H.; Tobia, M.J.; Mebes, I.; Katzler, O.; Sommer, T. The effect of estrogen synthesis inhibition on hippocampal memory. Psychoneuroendocrinology 2015, 56, 213–225. [Google Scholar] [CrossRef]
- Bailey, D.J.; Ma, C.; Soma, K.K.; Saldanha, C.J. Inhibition of hippocampal aromatization impairs spatial memory performance in a male songbird. Endocrinology 2013, 154, 4707–4714. [Google Scholar] [CrossRef] [Green Version]
- Bailey, D.J.; Makeyeva, Y.V.; Paitel, E.R.; Pedersen, A.L.; Hon, A.T.; Gunderson, J.A.; Saldanha, C.J. Hippocampal Aromatization Modulates Spatial Memory and Characteristics of the Synaptic Membrane in the Male Zebra Finch. Endocrinology 2017, 158, 852–859. [Google Scholar] [CrossRef] [Green Version]
- Bailey, D.J.; Saldanha, C.J. The importance of neural aromatization in the acquisition, recall, and integration of song and spatial memories in passerines. Horm. Behav. 2015, 74, 116–124. [Google Scholar] [CrossRef]
- Rosenfeld, C.S.; Shay, D.A.; Vieira-Potter, V.J. Cognitive Effects of Aromatase and Possible Role in Memory Disorders. Front. Endocrinol. 2018, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Zameer, S.; Vohora, D. Effect of aromatase inhibitors on learning and memory and modulation of hippocampal dickkopf-1 and sclerostin in female mice. Pharm. Rep. 2017, 69, 1300–1307. [Google Scholar] [CrossRef]
- Tuscher, J.J.; Szinte, J.S.; Starrett, J.R.; Krentzel, A.A.; Fortress, A.M.; Remage-Healey, L.; Frick, K.M. Inhibition of local estrogen synthesis in the hippocampus impairs hippocampal memory consolidation in ovariectomized female mice. Horm. Behav. 2016, 83, 60–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, C.R.; Graves, K.H.; Parlow, A.F.; Simpson, E.R. Characterization of mice deficient in aromatase (ArKO) because of targeted disruption of the cyp19 gene. Proc. Natl. Acad. Sci. USA 1998, 95, 6965–6970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.; Jones, M.; Simpson, E.; van den Buuse, M. Impaired spatial reference memory in aromatase-deficient (ArKO) mice. NeuroReport 2003, 14, 1979–1982. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Huangfu, X.; Zhao, Y.; Zhang, D.; Zhang, J. Letrozole induces worse hippocampal synaptic and dendritic changes and spatial memory impairment than ovariectomy in adult female mice. Neurosci. Lett. 2019, 706, 61–67. [Google Scholar] [CrossRef]
- Liu, M.; Huangfu, X.; Zhao, Y.; Zhang, D.; Zhang, J. Steroid receptor coactivator-1 mediates letrozole induced downregulation of postsynaptic protein PSD-95 in the hippocampus of adult female rats. J. Steroid Biochem. Mol. Biol. 2015, 154, 168–175. [Google Scholar] [CrossRef]
- Kelleher, R.J., III; Govindarajan, A.; Jung, H.Y.; Kang, H.; Tonegawa, S. Translational control by MAPK signaling in long-term synaptic plasticity and memory. Cell 2004, 116, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Sarbassov, D.D.; Ali, S.M.; Sabatini, D.M. Growing roles for the mTOR pathway. Curr. Opin. Cell Biol. 2005, 17, 596–603. [Google Scholar] [CrossRef]
- Thomas, G.M.; Huganir, R.L. MAPK cascade signalling and synaptic plasticity. Nat. Rev. Neurosci. 2004, 5, 173–183. [Google Scholar] [CrossRef]
- Yoshii, A.; Constantine-Paton, M. BDNF induces transport of PSD-95 to dendrites through PI3K-AKT signaling after NMDA receptor activation. Nat. Neurosci. 2007, 10, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, J.D. The neuronal MAP kinase cascade: A biochemical signal integration system subserving synaptic plasticity and memory. J. Neurochem. 2001, 76, 1–10. [Google Scholar] [CrossRef]
- Levenga, J.; Wong, H.; Milstead, R.A.; Keller, B.N.; LaPlante, L.E.; Hoeffer, C.A. AKT isoforms have distinct hippocampal expression and roles in synaptic plasticity. Elife 2017, 6, e30640. [Google Scholar] [CrossRef] [Green Version]
- Leal, G.; Comprido, D.; Duarte, C.B. BDNF-induced local protein synthesis and synaptic plasticity. Neuropharmacology 2014, 76, 639–656. [Google Scholar] [CrossRef] [Green Version]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for BDNF in learning and memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorski, J.A.; Balogh, S.A.; Wehner, J.M.; Jones, K.R. Learning deficits in forebrain-restricted brain-derived neurotrophic factor mutant mice. Neuroscience 2003, 121, 341–354. [Google Scholar] [CrossRef]
- Mizuno, M.; Yamada, K.; Olariu, A.; Nawa, H.; Nabeshima, T. Involvement of brain-derived neurotrophic factor in spatial memory formation and maintenance in a radial arm maze test in rats. J. Neurosci. 2000, 20, 7116–7121. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brann, D.W.; Lu, Y.; Wang, J.; Sareddy, G.R.; Pratap, U.P.; Zhang, Q.; Tekmal, R.R.; Vadlamudi, R.K. Neuron-Derived Estrogen—A Key Neuromodulator in Synaptic Function and Memory. Int. J. Mol. Sci. 2021, 22, 13242. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413242
Brann DW, Lu Y, Wang J, Sareddy GR, Pratap UP, Zhang Q, Tekmal RR, Vadlamudi RK. Neuron-Derived Estrogen—A Key Neuromodulator in Synaptic Function and Memory. International Journal of Molecular Sciences. 2021; 22(24):13242. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413242
Chicago/Turabian StyleBrann, Darrell W., Yujiao Lu, Jing Wang, Gangadhara R. Sareddy, Uday P. Pratap, Quanguang Zhang, Rajeshwar R. Tekmal, and Ratna K. Vadlamudi. 2021. "Neuron-Derived Estrogen—A Key Neuromodulator in Synaptic Function and Memory" International Journal of Molecular Sciences 22, no. 24: 13242. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413242