
Identification, Characterization and Expression Analysis of Anthocyanin Biosynthesis-related bHLH Genes in Blueberry (Vaccinium corymbosum L.)

 and
and
Abstract
:1. Introduction
2. Results
2.1. The Identified Anthocyanin Biosynthesis-Related Blueberry bHLH Genes
2.2. Physiochemical Properties Analysis of Blueberry bHLH Proteins
2.3. Gene Structrues of VcbHLH Genes and Conserved Conmains and Motifs in Their Encoded Proteins
2.4. Phylogenetic Analysis of Blueberry Anthocyanin Biosynthesis-Related bHLHs
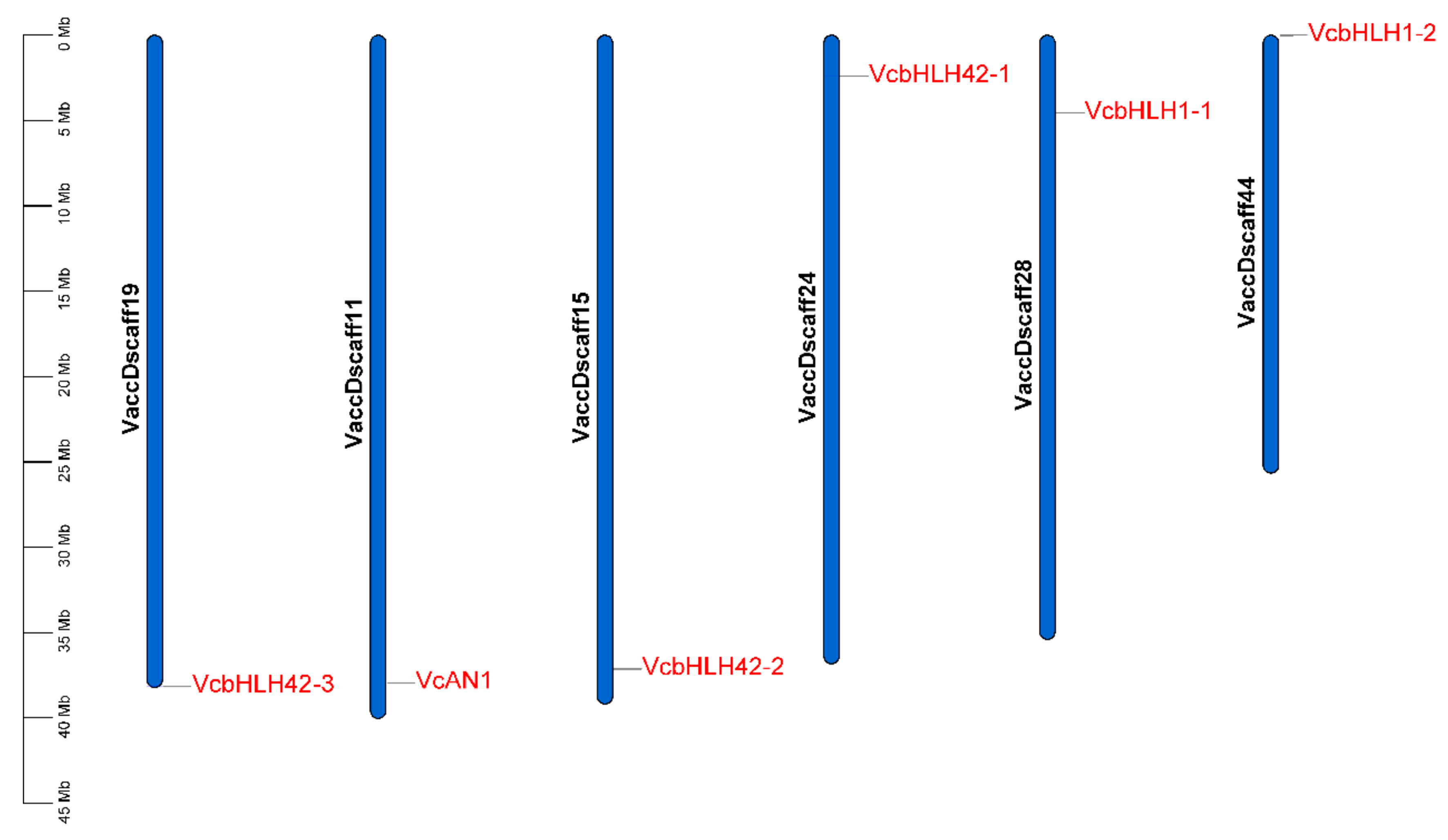
2.5. Chromosomal Location Analysis of Blueberry Anthocyanin Biosynthesis-Related bHLH Genes
2.6. cis-Acting Elements and TFBSs Distribution in Promoters of Blueberry Anthocyanin Biosynthesis-Related bHLH Genes
2.7. Protein-Protein Interaction Analysis of Blueberry bHLH Proteins
2.8. Cloning and Expression Analysis of Blueberry Anthocyanin Biosynthesis-Related bHLH Genes
3. Discussion
3.1. The Identified Blueberry Anthocyanin Biosynthesis-Related bHLHs Contained Conserved bHLH Domains and Key Amino Acids Required for Their DNA Binding Activity and Functions
3.2. Blueberry Anthocyanin Biosynthesis-Related bHLHs Could Be Classified into Two Groups, and the Functions of Each Group Differed from Each Other
3.3. There Are Many Factors Influencing the Expression and Functions of VcbHLH Genes
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of Anthocyanin Biosynthesis-Related Blueberry bHLH Genes
4.3. Bioinformatic Analysis of bHLH Genes and Their Encoded Proteins
4.4. Identification and Analysis of cis-Acting Elements and Transcription Factor Binding Sites in Promoter of Blueberry bHLH Genes
4.5. Gene Cloning and Sequencing of Blueberry bHLH Genes
4.6. Quantitative Real Time PCR Analysis of Blueberry bHLH Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, M.; Li, J.; Zhu, L.; Chang, P.; Li, L.; Zhang, L. Identification and characterization of MYB-bHLH-WD40 regulatory complex members controlling anthocyanidin biosynthesis in blueberry fruits development. Genes 2019, 10, 496. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.C.; Duk, J.Y.; Hee, D.O.; Jong, H.A. Transcriptional regulation of abscisic acid biosynthesis and signal transduction, and anthocyanin biosynthesis in ‘Bluecrop’ highbush blueberry fruit during ripening. PLoS ONE 2019, 14, e220015. [Google Scholar]
- Zhang, L.; Li, W.H.; Jiang, S.M.; Zhu, G.F.; Wang, B.Q.; Li, H.Q. Progress of molecular basis of biosynthesis and transcriptional regulation of anthocyanins. Acta Hortic. Sin. 2008, 6, 909–916. (In Chinese) [Google Scholar]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.C.; Hellens, R.P.; Laing, W.A. MYB transcription factors that colour our fruit. Trends Plant Sci. 2008, 13, 99–102. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, Y.; Li, L.; Meng, H.; Yang, Y.; Dong, Z.; Wang, L.; Wu, G. Genome-wide identification and characterization of bHLH transcription factors related to anthocyanin biosynthesis in red walnut (Juglans regia L.). Front. Genet. 2021, 12, 632509. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Tao, W.; Li, T.; Pan, W.; Chen, X.; Wu, X.; Nie, X.; Cui, L. Genome-wide identification, evolution and expression analysis of basic helix-loop-helix (bHLH) gene family in barley (Hordeum vulgare L.). Curr. Genom. 2020, 21, 621–644. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, S.R.; Habera, L.F.; Dellaporta, S.L.; Wessler, S.R. Lc, a member of the maize R gene family responsible fro tissue-specific anthocyanin production, encodes a protein similar to transcriptional activators and contains the myc-homology region. Proc. Natl. Acad. Sci. USA 1989, 86, 7092–7096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, A.M.; Walbot, V.; Davis, R.W. Arabidopsis and Nicotiana anthocyanin production activated by maize regulators R and C1. Science 1992, 258, 1773–1775. [Google Scholar] [CrossRef]
- Nesi, N.; Debeaujon, I.; Jond, C.; Pelletier, G.; Lepiniec, L. The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in arabidopsis siliques. Plant Cell 2000, 12, 1863–1878. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Li, J.J.; Su, M.Y.; Lin, Z.Y.; Chen, L.; Yang, P.F. A bHLH gene NnTT8 of Nelumbo nucifera regulates anthocyanin biosynthesis. Plant Physiol. Biochem. 2020, 158, 518–523. [Google Scholar] [CrossRef]
- Heng, S.; Wang, L.; Yang, X.; Huang, H.; Chen, G.; Cui, M.; Liu, M.; Lv, Q.; Wan, Z.; Shen, J.; et al. Genetic and comparative transcriptome analysis revealed DEGs involved in the purple leaf formation in Brassica juncea. Front. Genet. 2020, 11, 322. [Google Scholar] [CrossRef]
- Xie, X.B.; Li, S.; Zhang, R.F.; Zhao, J.; Chen, Y.C.; Zhao, Q.; Yao, Y.X.; You, C.X.; Zhang, X.S.; Hao, Y.J. The bHLH transcription factor MdbHLH3 promotes anthocyanin accumulation and fruit colouration in response to low temperature in apples. Plant Cell Environ. 2012, 35, 1884–1897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ye, J.; Liu, C.; Xu, Q.; Long, L.; Deng, X. The citrus CsPH4-Noemi regulatory complex is involved in proanthocyanidin biosynthesis via a positive feedback loop. J. Exp. Bot. 2019, 71, 1306–1321. [Google Scholar] [CrossRef]
- Huang, D.; Yuan, Y.; Tang, Z.; Huang, Y.; Kang, C.; Deng, X.; Xu, Q. Retrotransposon promoter of Ruby1 controls both light- and cold-induced accumulation of anthocyanins in blood orange. Plant Cell Environ. 2019, 42, 1302–1321. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Song, Z.; Zhang, Y.; Li, Z.; Wang, Y.; Liu, X.; Ma, J.; Quan, J.; Wu, X.; Liu, M.; et al. The bHLH transcription factor PPLS1 regulates the color of pulvinus and leaf sheath in foxtail millet (Setaria italica). Theor. Appl. Genet. 2020, 133, 1911–1926. [Google Scholar] [CrossRef]
- Zhao, S.; Xi, X.; Zong, Y.; Li, S.; Li, Y.; Cao, D.; Liu, B. Overexpression of ThMYC4E enhances anthocyanin biosynthesis in common wheat. Int. J. Mol. Sci. 2019, 21, 137. [Google Scholar] [CrossRef] [Green Version]
- Spelt, C.; Quattrocchio, F.; Mol, J.N.; Koes, R. anthocyanin1 of petunia encodes a basic helix-loop-helix protein that directly activates transcription of structural anthocyanin genes. Plant Cell 2000, 12, 1619–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günther, C.S.; Dare, A.P.; McGhie, T.K.; Deng, C.; Lafferty, D.J.; Plunkett, B.J.; Grierson, E.R.P.; Turner, J.L.; Jaakola, L.; Albert, N.W.; et al. Spatiotemporal modulation of flavonoid metabolism in blueberries. Front. Plant Sci. 2020, 11, 545. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Guo, Q.X.; Liu, Y.J.; Liu, H.C.; Wang, F.T.; Jia, C.G. Molecular cloning and functional analysis of a flavanone 3-hydroxylase gene from blueberry. J. Hortic. Sci. Biotechnol. 2017, 92, 57–64. [Google Scholar] [CrossRef]
- Yang, J.F.; Li, B.B.; Shi, W.J.; Gong, Z.Z.; Chen, L.; Hou, Z.X. Transcriptional activation of anthocyanin biosynthesis in developing fruit of blueberries (Vaccinium corymbosum L.) by preharvest and postharvest UV irradiation. J. Agric. Food Chem. 2018, 66, 10931–10942. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.T.; Lim, S.; Lee, J.G.; Lee, E.J. VcBBX, VcMYB21, and VcR2R3MYB transcription factors are involved in UV–B-induced anthocyanin biosynthesis in the peel of harvested blueberry fruit. J. Agric. Food Chem. 2017, 65, 2066–2073. [Google Scholar] [CrossRef]
- Plunkett, B.J.; Espley, R.V.; Dare, A.P.; Warren, B.; Grierson, E.; Cordiner, S.; Turner, J.L.; Allan, A.C.; Albert, N.W.; Davies, K.M.; et al. MYBA from blueberry (Vaccinium Section Cyanococcus) is a subgroup 6 type R2R3MYB transcription factor that activates anthocyanin production. Front. Plant Sci. 2018, 9, 1300. [Google Scholar] [CrossRef]
- Yang, S.; Liu, H.D.; Wang, H.B.; Zhang, H.J.; Liu, F.Z. Molecular cloning and functional characterization of Vc NAC072 reveals its involvement in anthocyanin accumulation in blueberry. Sci. Agric. Sin. 2019, 52, 503–511. (In Chinese) [Google Scholar]
- Die, J.V.; Jones, R.W.; Ogden, E.L.; Ehlenfeldt, M.K.; Rowland, L.J. Characterization and analysis of anthocyanin-related genes in wild-type blueberry and the pink-fruited mutant cultivar ‘Pink Lemonade’: New insights into anthocyanin biosynthesis. Agronomy 2020, 10, 1296. [Google Scholar] [CrossRef]
- Karppinen, K.; Lafferty, D.J.; Albert, N.W.; Mikkola, N.; McGhie, T.; Allan, A.C.; Afzal, B.M.; Häggman, H.; Espley, R.V.; Jaakola, L. MYBA and MYBPA transcription factors co-regulate anthocyanin biosynthesis in blue-coloured berries. New Phytol. 2021, 232, 1350–1367. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Chi, F.M.; Liu, H.D.; Zhang, H.J.; Song, Y. Single-molecule real-time and illumina sequencing to analyze transcriptional regulation of flavonoid synthesis in blueberry. Front. Plant Sci. 2021, 12, 754325. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jin, L.; Pan, X.; Yang, L.; Guo, W. Proteins expression and metabolite profile insight into phenolic biosynthesis during highbush blueberry fruit maturation. Food Chem. 2019, 290, 216–228. [Google Scholar] [CrossRef]
- Gupta, V.; Estrada, A.D.; Blakley, I.; Reid, R.; Patel, K.; Meyer, M.D.; Andersen, S.U.; Brown, A.F.; Lila, M.A.; Loraine, A.E. RNA-Seq analysis and annotation of a draft blueberry genome assembly identifies candidate genes involved in fruit ripening, biosynthesis of bioactive compounds, and stage-specific alternative splicing. Gigascience 2015, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H.; et al. Haplotype-phased genome and evolution of phytonutrient pathways of tetraploid blueberry. Gigascience 2019, 8, giz012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, L.J.; Alkharouf, N.; Darwish, O.; Ogden, E.L.; Polashock, J.J.; Bassil, N.V.; Main, D. Generation and analysis of blueberry transcriptome sequences from leaves, developing fruit, and flower buds from cold acclimation through deacclimation. BMC Plant Biol. 2012, 12, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moglia, A.; Florio, F.E.; Iacopino, S.; Guerrieri, A.; Milani, A.M.; Comino, C.; Barchi, L.; Marengo, A.; Cagliero, C.; Rubiolo, P.; et al. Identification of a new R3 MYB type repressor and functional characterization of the members of the MBW transcriptional complex involved in anthocyanin biosynthesis in eggplant (S. melongena L.). PLoS ONE 2020, 15, e232986. [Google Scholar]
- Wang, L.; Tang, W.; Hu, Y.; Zhang, Y.; Sun, J.; Guo, X.; Lu, H.; Yang, Y.; Fang, C.; Niu, X.; et al. A MYB/bHLH complex regulates tissue-specific anthocyanin biosynthesis in the inner pericarp of red-centered kiwifruit Actinidia chinensis cv. Hongyang. Plant J. 2019, 99, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.G.; Sun, C.H.; Zhang, Q.Y.; Gu, K.D.; Hao, Y.J. The basic helix-loop-helix transcription factor MdbHLH3 modulates leaf senescence in apple via the regulation of dehydratase-enolase-phosphatase complex 1. Hortic. Res. 2020, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Wang, H.; Li, D.; Yu, B.; Hui, Q.; Yan, S.; Huang, Z.; Cui, X.; Cao, B. Identification of candidate HY5-dependent and -independent regulators of anthocyanin biosynthesis in tomato. Plant Cell Physiol. 2019, 60, 643–656. [Google Scholar] [CrossRef]
- Wei, Z.Z.; Hu, K.D.; Zhao, D.L.; Tang, J.; Huang, Z.Q.; Jin, P.; Li, Y.H.; Han, Z.; Hu, L.Y.; Yao, G.F.; et al. MYB44 competitively inhibits the formation of the MYB340-bHLH2-NAC56 complex to regulate anthocyanin biosynthesis in purple-fleshed sweet potato. BMC Plant Biol. 2020, 20, 258. [Google Scholar] [CrossRef]
- Bai, Q.; Duan, B.; Ma, J.; Fen, Y.; Sun, S.; Long, Q.; Lv, J.; Wan, D. Coexpression of PalbHLH1 and PalMYB90 genes from populus alba enhances pathogen resistance in poplar by increasing the flavonoid content. Front. Plant Sci. 2019, 10, 1772. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, C.; Lee, M.M.; Gonzalez, A.; Zhang, F.; Lloyd, A.; Schiefelbein, J. The bHLH genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in the Arabidopsis root. Development 2003, 130, 6431–6439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Li, H.; Wu, J.; Wang, B.; Tian, N.; Liu, J.; Sun, X.; Wu, H.; Huang, Y.; Lu, P.; et al. Genome-wide identification and expression pattern analysis of lipoxygenase gene family in banana. Sci. Rep. 2021, 11, 9948. [Google Scholar] [CrossRef]
- Salih, H.; Tan, L.; Htet, N.W. Genome-wide identification, characterization of bHLH transcription factors in mango. Trop. Plant Biol. 2021, 14, 72–81. [Google Scholar] [CrossRef]
- Carretero-Paulet, L.; Galstyan, A.; Roig-Villanova, I.; Martinez-Garcia, J.F.; Bilbao-Castro, J.R.; Robertson, D.L. Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in arabidopsis, poplar, rice, moss, and algae. Plant Physiol. 2010, 153, 1398–1412. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Sun, L.; Gu, H.; Cheng, D.; Guo, X.; Chen, R.; Wu, Z.; Jiang, J.; Fan, X.; Chen, J. Genome-wide characterization and analysis of bHLH transcription factors related to anthocyanin biosynthesis in spine grapes (Vitis davidii). Sci. Rep. 2021, 11, 6863. [Google Scholar] [CrossRef]
- Bassolino, L.; Buti, M.; Fulvio, F.; Pennesi, A.; Mandolino, G.; Milc, J.; Francia, E.; Paris, R. In silico identification of MYB and bHLH families reveals candidate transcription factors for secondary metabolic pathways in Cannabis sativa L. Plants 2020, 9, 1540. [Google Scholar] [CrossRef]
- Ferré-D’Amaré, A.R.; Pognonec, P.; Roeder, R.G.; Burley, S.K. Structure and function of the b/HLH/Z domain of USF. EMBO J. 1994, 13, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Toumoto, A.; Ihara, K.; Shimizu, M.; Kyogoku, Y.; Ogawa, N.; Oshima, Y.; Hakoshima, T. Crystal structure of PHO4 bHLH domain-DNA complex: Flanking base recognition. EMBO J. 1997, 16, 4689–4697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiferle, C.; Fantini, E.; Bassolino, L.; Povero, G.; Spelt, C.; Buti, S.; Giuliano, G.; Quattrocchio, F.; Koes, R.; Perata, P.; et al. Tomato R2R3-MYB proteins SlANT1 and SlAN2: Same protein activity, different roles. PLoS ONE 2015, 10, e136365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Y.; Zhang, T.; Yu, Y.; Gou, L.; Yang, J.; Xu, J.; Pi, E. Regulatory mechanisms of bHLH transcription factors in plant adaptive responses to various abiotic stresses. Front. Plant Sci. 2021, 12, 677611. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Yu, W.; Gao, Y.; Ni, J.; Yin, L.; Zhang, X.; Li, H.; Wang, D.; Bai, S.; Teng, Y. Light-induced basic/helix-loop-helix 64 enhances anthocyanin biosynthesis and undergoes CONSTITUTIVELY PHOTOMORPHOGENIC 1-mediated degradation in pear. Plant Physiol. 2020, 184, 1684–1701. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yamane, H.; Tao, R. Preharvest long-term exposure to UV-B radiation promotes fruit ripening and modifies stage-specific anthocyanin metabolism in highbush blueberry. Hortic. Res. 2021, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Le Hir, R.; Castelain, M.; Chakraborti, D.; Moritz, T.; Dinant, S.; Bellini, C. AtbHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis thaliana. Physiol. Plant. 2017, 160, 312–327. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.S.; Joo, J.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; Song, S.I.; Cheong, J.J.; Lee, J.S.; Kim, J.K.; Choi, Y.D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef]
- Li, D.; Luo, Z.; Mou, W.; Wang, Y.; Ying, T.; Mao, L. ABA and UV-C effects on quality, antioxidant capacity and anthocyanin contents of strawberry fruit (Fragaria ananassa Duch.). Postharvest Biol. Technol. 2014, 90, 56–62. [Google Scholar] [CrossRef]
- Li, T.; Jia, K.P.; Lian, H.L.; Yang, X.; Li, L.; Yang, H.Q. Jasmonic acid enhancement of anthocyanin accumulation is dependent on phytochrome a signaling pathway under far-red light in Arabidopsis. Biochem. Biophys. Res. Commun. 2014, 454, 78–83. [Google Scholar] [CrossRef]
- Wang, F.; Lin, R.; Feng, J.; Qiu, D.; Chen, W.; Xu, S. Wheat bHLH transcription factor gene, TabHLH060, enhances susceptibility of transgenic Arabidopsis thaliana to Pseudomonas syringae. Physiol. Mol. Plant Pathol. 2015, 90, 123–130. [Google Scholar] [CrossRef]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, M.; Mitra, S.; Muhammad, R.H.; Debnath, B.; Lu, X.; Jian, H.; Qiu, D. Comparative phytochemical profiles and antioxidant enzyme activity analyses of the southern highbush blueberry (Vaccinium corymbosum) at different developmental stages. Molecules 2018, 23, 2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011, 66, 94–116. [Google Scholar] [CrossRef]
- Qiao, F.; Cong, H.; Jiang, X.; Wang, R.; Yin, J.; Qian, D.; Wang, Z.; Nick, P. De Novo characterization of a Cephalotaxus hainanensis transcriptome and genes related to paclitaxel biosynthesis. PLoS ONE 2014, 9, e106900. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Homologous Gene Name | Homologous Gene ID | Identity/% | Reference |
|---|---|---|---|---|---|
| VcAN1 | VaccDscaff11-processed-gene-379.7 | SmelAN1 | SMEL_009g326640.1.01 | 77.12 | [33] |
| AcbHLH42 | MH643775 | 74.42 | [34] | ||
| MdbHLH3 | ADL36597.1 | 65.08 | [14,35] | ||
| Noemi | Cs5g31400 | 63.08 | [15,16] | ||
| SlAN1 | KR076778 | 52.03 | [37] | ||
| IbbHLH2 | itf14g18730.t2 | 46.45 | [11] | ||
| VcbHLH42-1 | VaccDscaff24-augustus-gene-24.28 | AcbHLH42 | MH643775 | 79.10 | [34] |
| AtTT8 | At4G09820 | 73.21 | [13] | ||
| MdbHLH3 | ADL36597.1 | 69.03 | [14,35] | ||
| Noemi | Cs5g31400 | 67.25 | [15,16] | ||
| BjTT8 | BjuB004115 | 58.48 | [38] | ||
| SlAN1 | KR076778 | 54.77 | [37] | ||
| SmelAN1 | SMEL_009g326640.1.01 | 50.14 | [33] | ||
| VcbHLH42-2 | VaccDscaff15-augustus-gene-371.25 | AcbHLH42 | MH643775 | 79.37 | [34] |
| MdbHLH3 | ADL36597.1 | 69.30 | [14,35] | ||
| Noemi | Cs5g31400 | 67.11 | [15,16] | ||
| SlAN1 | KR076778 | 54.77 | [37] | ||
| SmelAN1 | SMEL_009g326640.1.01 | 49.72 | [33] | ||
| VcbHLH42-3 | VaccDscaff19-augustus-gene-381.30 | AcbHLH42 | MH643775 | 79.23 | [34] |
| MdbHLH3 | ADL36597.1 | 69.30 | [14,35] | ||
| Noemi | Cs5g31400 | 67.11 | [15,16] | ||
| SlAN1 | KR076778 | 54.77 | [36] | ||
| SmelAN1 | SMEL_009g326640.1.01 | 49.72 | [33] | ||
| VcbHLH1-1 | VaccDscaff28-augustus-gene-45.27 | PalbHLH1 | PAYT030711.1 | 57.12 | [38] |
| PPLS1 | Seita.7G195400 | 53.85 | [17] | ||
| SmelJAF13 | SMEL_008g319200.1.01 | 49.69 | [33] | ||
| AtEGL3 | At1G63650 | 48.23 | [39] | ||
| AtGL3 | At5G41315 | 47.77 | [39] | ||
| ThMYC4E | KX914905 | 37.54 | [18] | ||
| VcbHLH1-2 | VaccDscaff44-augustus-gene-0.19 | PalbHLH1 | PAYT030711.1 | 55.61 | [38] |
| SmelJAF13 | SMEL_008g319200.1.01 | 48.55 | [33] | ||
| AtEGL3 | At1G63650 | 46.95 | [39] | ||
| AtGL3 | At5G41315 | 46.42 | [39] |
| Protein Name | Gene ID | CDS Length/bp | Protein Size/aa | Molecular Weight/Da | PI | Instability Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| VcAN1 | VaccDscaff11-processed-gene-379.7 | 2328 | 775 | 85,474.08 | 5.55 | 67.34 | −0.595 | Nuclear |
| VcbHLH42-1 | VaccDscaff24-augustus-gene-24.28 | 2190 | 729 | 80,213.15 | 5.46 | 69.43 | −0.618 | Nuclear |
| VcbHLH42-2 | VaccDscaff15-augustus-gene-371.25 | 2190 | 729 | 80,320.22 | 5.45 | 70.96 | −0.64 | Nuclear |
| VcbHLH42-3 | VaccDscaff19-augustus-gene-381.30 | 2190 | 729 | 80,280.2 | 5.49 | 69.96 | −0.629 | Nuclear |
| VcbHLH1-1 | VaccDscaff28-augustus-gene-45.27 | 1803 | 600 | 67,581.48 | 6.25 | 55.99 | −0.388 | Nuclear |
| VcbHLH1-2 | VaccDscaff44-augustus-gene-0.19 | 1860 | 619 | 69,831.17 | 6.16 | 56.26 | −0.358 | Nuclear |
| Target Gene | Primer Name | Primer Sequence | Target Length (bp) | Tm (°C) | Application |
|---|---|---|---|---|---|
| VcAN1 | VcAN1-F | GCAACCCTCTCTCTTTCACT | 2885 | 60.5 | Gene cloning |
| VcAN1-R | CCATTTCATTACCGCAAGC | ||||
| VcbHLH1-1 | VcbHLH1-1-F | GCACAGAATCAATGGCTTC | 2043 | 60 | |
| VcbHLH1-1-R | CGTGTTTGTTGGTTTGGC | ||||
| VcbHLH1-2 | VcbHLH1-2-F | TTCCCTTACCCATCTTCCT | 2415 | 60 | |
| VcbHLH1-2-R | CTGTGTTTGTTGGTTTGGC | ||||
| VcAN1 | VcAN1-qF | CACCCTCCACAGCCTCCGAA | 242 | 59 | qRT-PCR |
| VcAN1-qR | CTATTCCGCCCCGTGTCAGC | ||||
| VcbHLH1-1 | VcbHLH1-1-qF | CTTGGAATGGGGTGATGGGT | 210 | 59 | |
| VcbHLH1-1-qR | CACTCGGTATCGGTCAGGTC | ||||
| VcbHLH1-2 | VcbHLH1-2-qF | AGCTGGAGAGAAGGGTCGAA | 155 | 59 | |
| VcbHLH1-2-qR | GTCCCGTGCTTTCCTCTTGT | ||||
| GAPDH | GAPDH-qF | ACTACCATCCACTCTATCACCG | 116 | 59 | Reference gene |
| GAPDH-qR | AACACCTTACCAACAGCCTTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, F.; Wang, B.; Wu, H.; Wu, J.; Liu, J.; Sun, Y.; Cheng, C.; Qiu, D. Identification, Characterization and Expression Analysis of Anthocyanin Biosynthesis-related bHLH Genes in Blueberry (Vaccinium corymbosum L.). Int. J. Mol. Sci. 2021, 22, 13274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413274
Zhang Y, Liu F, Wang B, Wu H, Wu J, Liu J, Sun Y, Cheng C, Qiu D. Identification, Characterization and Expression Analysis of Anthocyanin Biosynthesis-related bHLH Genes in Blueberry (Vaccinium corymbosum L.). International Journal of Molecular Sciences. 2021; 22(24):13274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413274
Chicago/Turabian StyleZhang, Yongyan, Fan Liu, Bin Wang, Huan Wu, Junwei Wu, Jiapeng Liu, Yueting Sun, Chunzhen Cheng, and Dongliang Qiu. 2021. "Identification, Characterization and Expression Analysis of Anthocyanin Biosynthesis-related bHLH Genes in Blueberry (Vaccinium corymbosum L.)" International Journal of Molecular Sciences 22, no. 24: 13274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413274