
Primary Cilia Structure Is Prolonged in Enteric Neurons of 5xFAD Alzheimer’s Disease Model Mice

 , ,
, , 
Abstract
:1. Introduction
2. Results
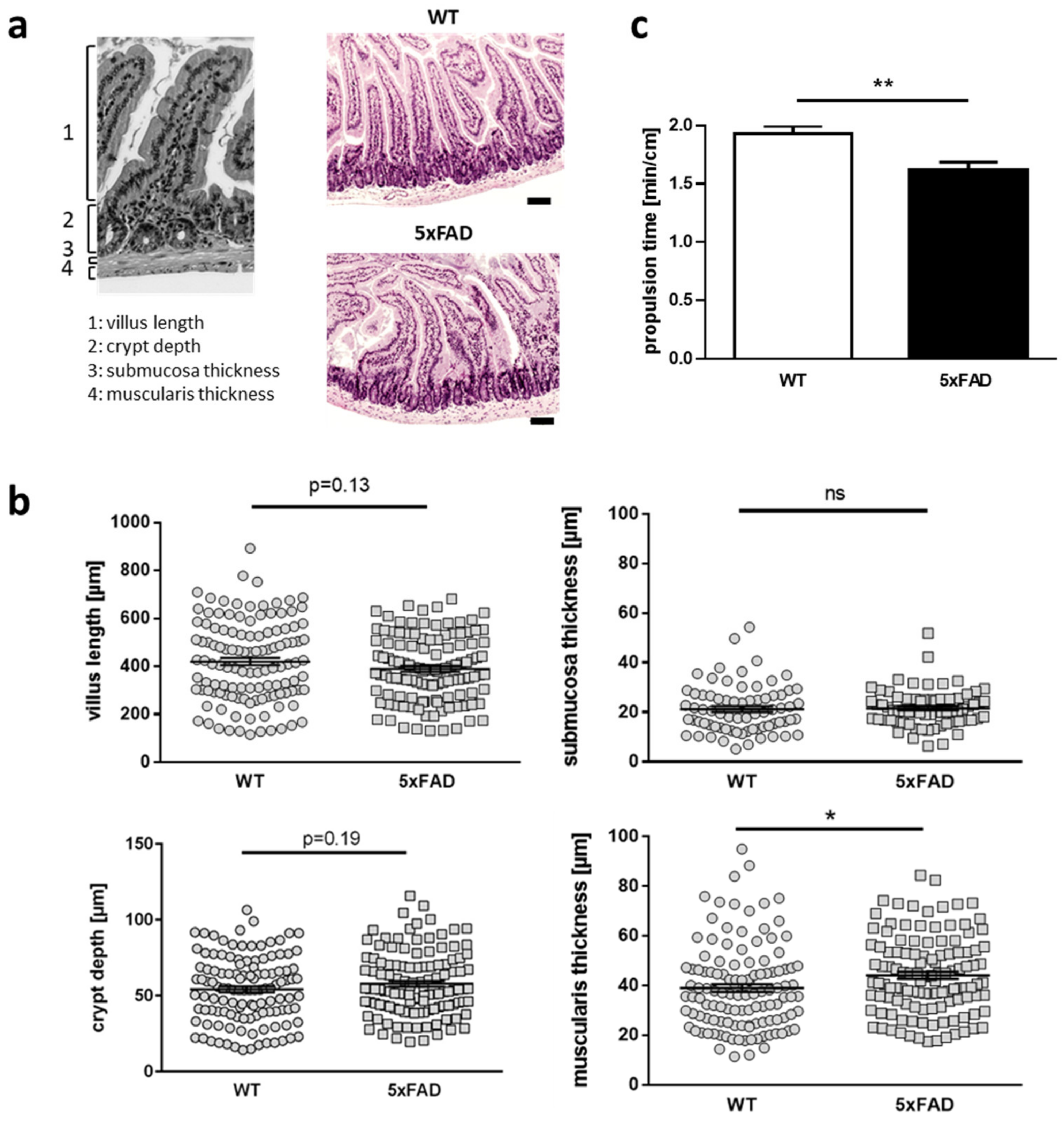
2.1. Differences in Gut Anatomy and Function Driven by FAD Mutations in a Mouse Model of Alzheimer’s Disease
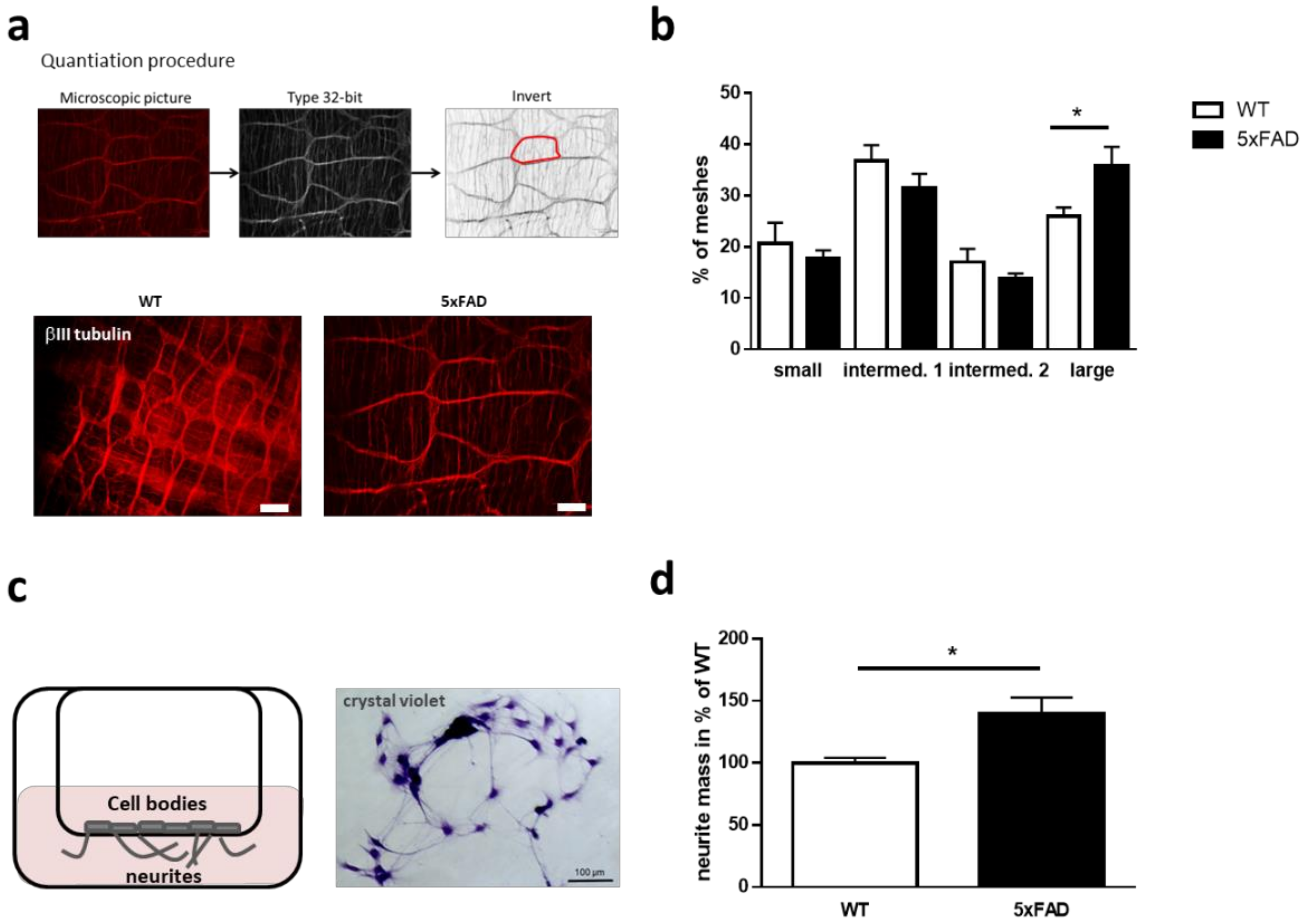
2.2. Induced Neurite Extension of Enteric Neurons Derived from 5xFAD Mice
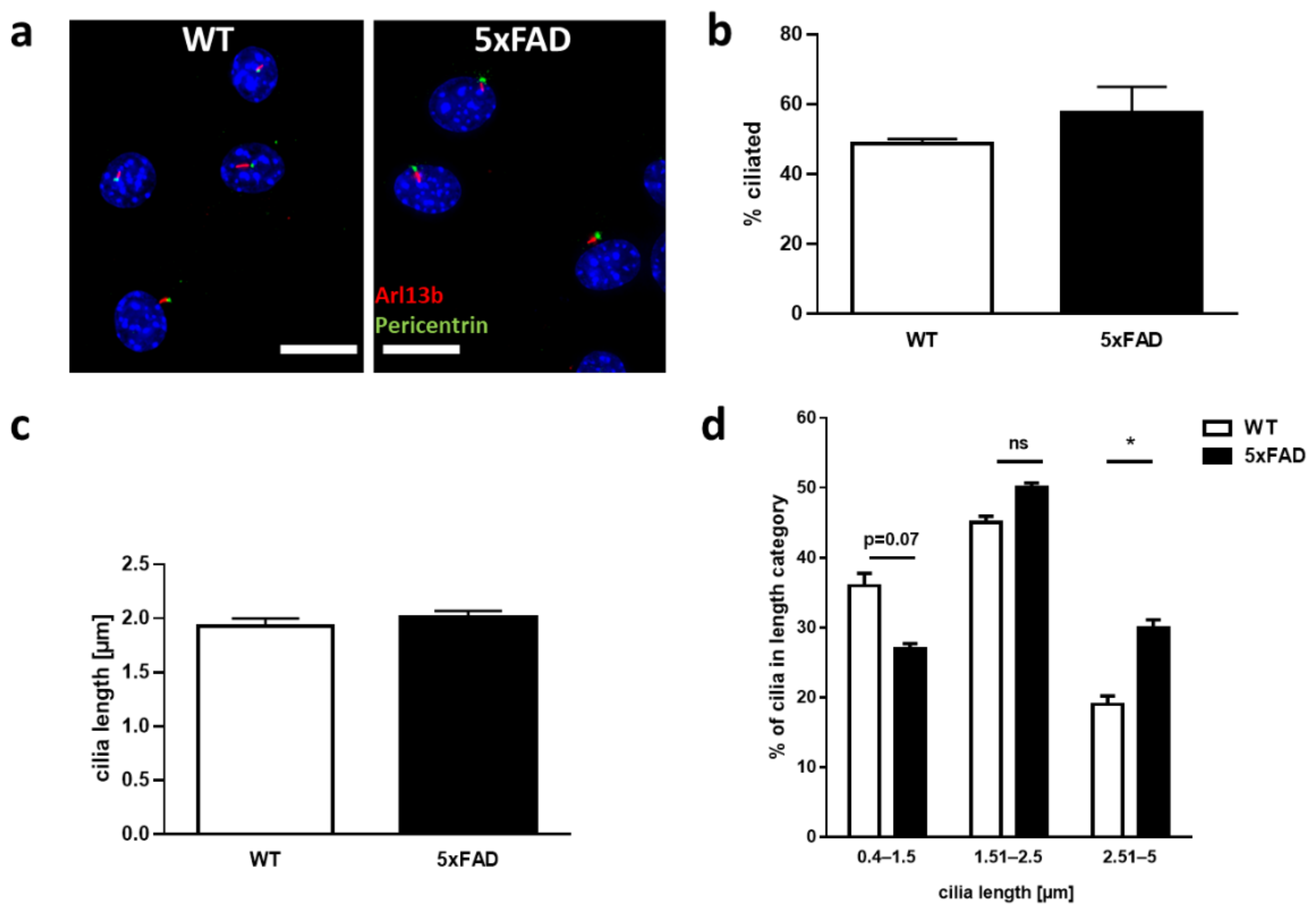
2.3. Characterization of Primary Cilia in Primary Enteric Neuron Culture of 5xFAD Mice
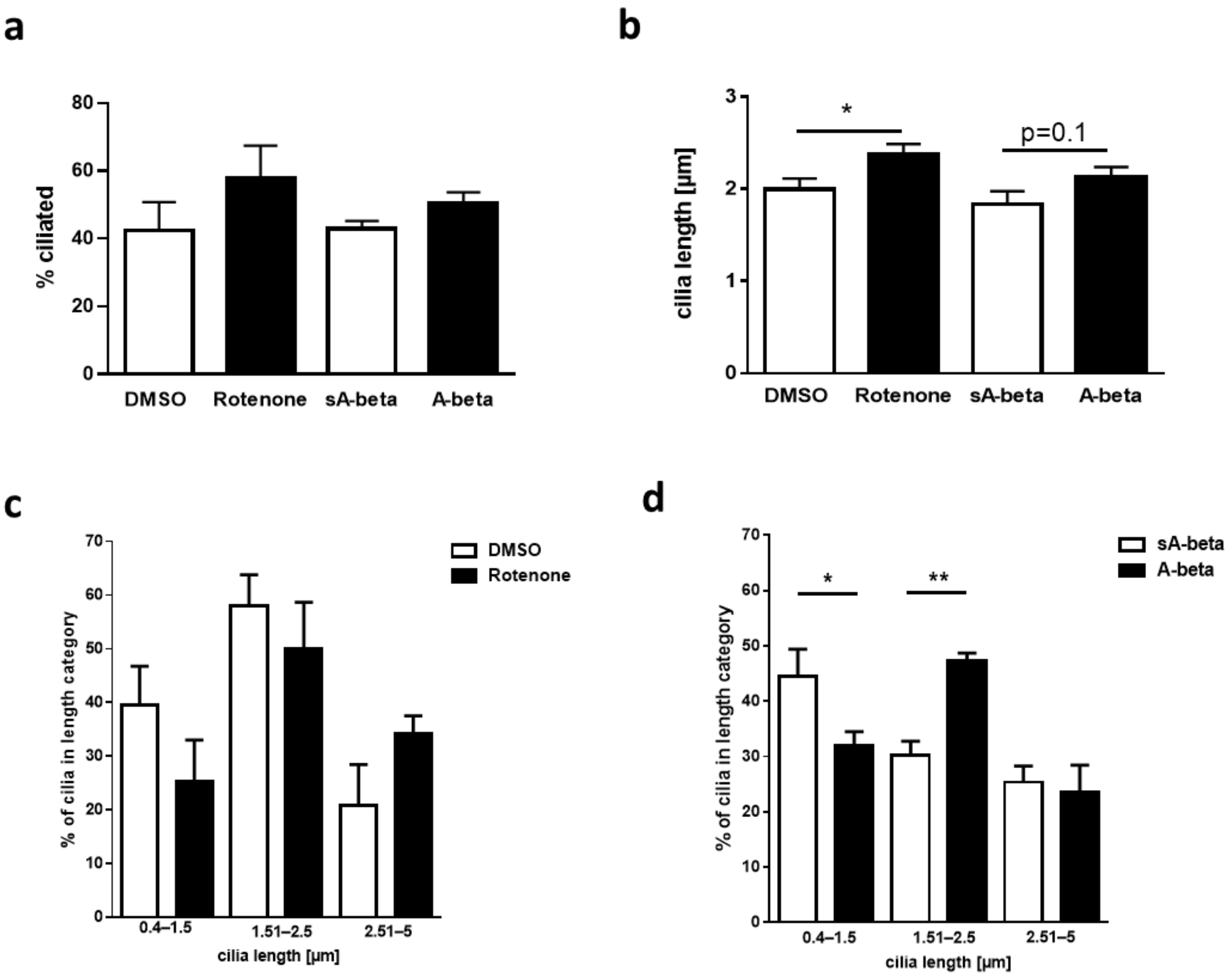
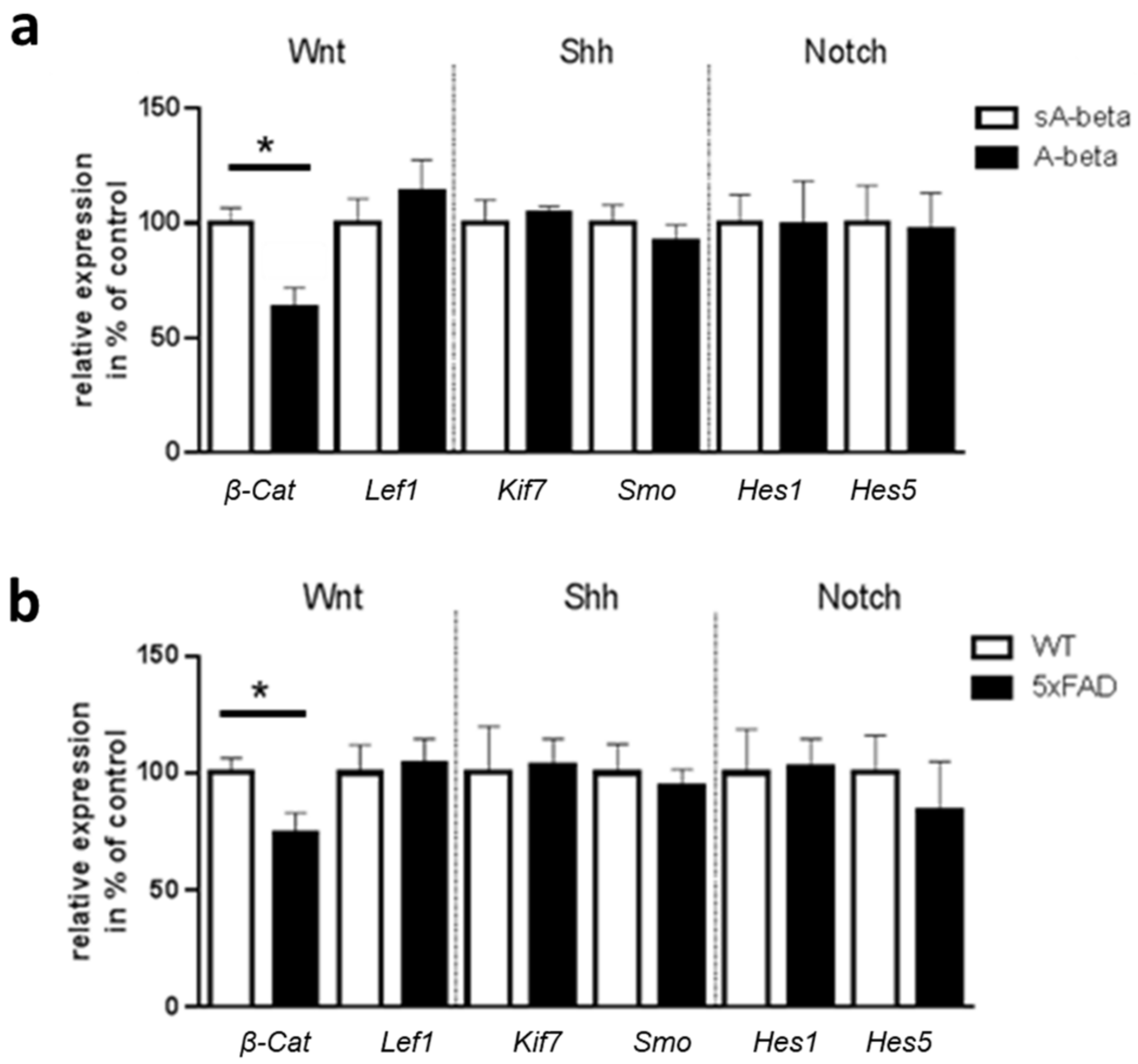
2.4. Effects of Acute Administration of A-beta Peptide on Primary Cilia of Murine Enteric Neurons and on Ciliary Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. HE Staining and Duodenum Morphological Analysis
4.3. Ex Vivo Gut Motility Measurement
4.4. Primary Culture of Enteric Neurons
4.5. βIII Tubulin Staining of LMMPs
4.6. RNA Preparation and qPCR
4.7. Neurite Mass Quantitation
4.8. Immunofluorescence
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Beales, P.; Jackson, P.K. Cilia—The prodigal organelle. Cilia 2012, 1, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef]
- Kenny, T.D.; Beales, P.L. Ciliopathies: A reference for clinicians. Clin. Kidney J. 2013, 7, 92. [Google Scholar] [CrossRef]
- Estrada-Cuzcano, A.; Roepman, R.; Cremers, F.P.; den Hollander, A.I.; Mans, D.A. Non-syndromic retinal ciliopathies: Translating gene discovery into therapy. Hum. Mol. Genet. 2012, 21, R111–R124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnicka-Turek, O.; Steimle, J.D.; Huang, W.; Felker, L.; Kamp, A.; Kweon, J.; Peterson, M.; Reeves, R.H.; Maslen, C.L.; Gruber, P.J.; et al. Cilia gene mutations cause atrioventricular septal defects by multiple mechanisms. Hum. Mol. Genet. 2016, 25, 3011–3028. [Google Scholar] [CrossRef] [PubMed]
- Grochowsky, A.; Gunay-Aygun, M. Clinical characteristics of individual organ system disease in non-motile ciliopathies. Transl. Sci. Rare Dis. 2019, 4, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suciu, S.K.; Caspary, T. Cilia, neural development and disease. Semin. Cell Dev. Biol. 2021, 110, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Armato, U.; Chakravarthy, B.; Pacchiana, R.; Whitfield, J.F. Alzheimer’s disease: An update of the roles of receptors, astrocytes and primary cilia. Int. J. Mol. Med. 2013, 31, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, B.; Zhang, Y. Serotonin 5-HT6 receptors affect cognition in a mouse model of Alzheimer’s disease by regulating cilia function. Alzheimers Res. Ther. 2017, 9, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorobyeva, A.G.; Saunders, A.J. Amyloid-beta interrupts canonical Sonic hedgehog signaling by distorting primary cilia structure. Cilia 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, L.H.; Schreiber, H.L.t.; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Luesma, M.J.; Cantarero, I.; Castiella, T.; Soriano, M.; Garcia-Verdugo, J.M.; Junquera, C. Enteric neurons show a primary cilium. J Cell Mol. Med. 2013, 17, 147–153. [Google Scholar] [CrossRef]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef]
- Spencer, N.J.; Hu, H. Enteric nervous system: Sensory transduction, neural circuits and gastrointestinal motility. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Brandscheid, C.; Schuck, F.; Reinhardt, S.; Schafer, K.H.; Pietrzik, C.U.; Grimm, M.; Hartmann, T.; Schwiertz, A.; Endres, K. Altered Gut Microbiome Composition and Tryptic Activity of the 5xFAD Alzheimer’s Mouse Model. J. Alzheimers Dis. 2017, 56, 775–788. [Google Scholar] [CrossRef]
- Chen, C.; Ahn, E.H.; Kang, S.S.; Liu, X.; Alam, A.; Ye, K. Gut dysbiosis contributes to amyloid pathology, associated with C/EBPβ/AEP signaling activation in Alzheimer’s disease mouse model. Sci. Adv. 2020, 6, eaba0466. [Google Scholar] [CrossRef] [PubMed]
- Stoye, N.M.; Dos Santos Guilherme, M.; Endres, K. Alzheimer’s disease in the gut-Major changes in the gut of 5xFAD model mice with ApoA1 as potential key player. FASEB J. 2020, 34, 11883–11899. [Google Scholar] [CrossRef]
- Harach, T.; Marungruang, N.; Duthilleul, N.; Cheatham, V.; Mc Coy, K.D.; Frisoni, G.; Neher, J.J.; Fak, F.; Jucker, M.; Lasser, T.; et al. Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci. Rep. 2017, 7, 41802. [Google Scholar] [CrossRef]
- Chakravarthy, B.; Gaudet, C.; Menard, M.; Atkinson, T.; Chiarini, A.; Dal Pra, I.; Whitfield, J. The p75 neurotrophin receptor is localized to primary cilia in adult murine hippocampal dentate gyrus granule cells. Biochem. Biophys. Res. Commun. 2010, 401, 458–462. [Google Scholar] [CrossRef]
- Chen, G.F.; Xu, T.H.; Yan, Y.; Zhou, Y.R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [PubMed]
- Wheway, G.; Nazlamova, L.; Hancock, J.T. Signaling through the Primary Cilium. Front Cell Dev. Biol. 2018, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, Y.M.; Borcherding, D.; Kanthasamy, A.; Kim, H.J.; Willette, A.A.; Jergens, A.; Allenspach, K.; Mochel, J.P. The Gut–brain Axis in Neurodegenerative Diseases and Relevance of the Canine Model: A Review. Front. Aging Neurosci. 2019, 11, 130. [Google Scholar] [CrossRef]
- Gries, M.; Christmann, A.; Schulte, S.; Weyland, M.; Rommel, S.; Martin, M.; Baller, M.; Roth, R.; Schmitteckert, S.; Unger, M.; et al. Parkinson mice show functional and molecular changes in the gut long before motoric disease onset. Mol. Neurodegener. 2021, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Yagi, H.; Uemura, M.T.; Hatanaka, Y.; Yamakado, H.; Takahashi, R. Inoculation of α-synuclein preformed fibrils into the mouse gastrointestinal tract induces Lewy body-like aggregates in the brainstem via the vagus nerve. Mol. Neurodegener. 2018, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, K.; Mulak, A. Brain–gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Gleeson, J.G. The role of primary cilia in neuronal function. Neurobiol. Dis. 2010, 38, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenke, E.; Bourke, B.; Knaus, U. Crohn’s Strictures-Moving Away from the Knife. Front. Pediatr. 2017, 5, 141. [Google Scholar] [CrossRef] [Green Version]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement. 2018, 4, 575–590. [Google Scholar] [CrossRef]
- Huizinga, J.D.; Lammers, W.J. Gut peristalsis is governed by a multitude of cooperating mechanisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1–G8. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Rademakers, G.; Vaes, N.; Schonkeren, S.; Koch, A.; Sharkey, K.A.; Melotte, V. The role of enteric neurons in the development and progression of colorectal cancer. Biochim. Biophys. Acta Rev. Cancer. 2017, 1868, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Butler Tjaden, N.E.; Trainor, P.A. The developmental etiology and pathogenesis of Hirschsprung disease. Transl. Res. 2013, 162, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Semar, S.; Klotz, M.; Letiembre, M.; Van Ginneken, C.; Braun, A.; Jost, V.; Bischof, M.; Lammers, W.J.; Liu, Y.; Fassbender, K.; et al. Changes of the enteric nervous system in amyloid-beta protein precursor transgenic mice correlate with disease progression. J. Alzheimers Dis. 2013, 36, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Mazi, A.R.; Arzuman, A.S.; Gurel, B.; Sahin, B.; Tuzuner, M.B.; Ozansoy, M.; Baykal, A.T. Neonatal Neurodegeneration in Alzheimer’s Disease Transgenic Mouse Model. J. Alzheimers Dis. Rep. 2018, 2, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Milward, E.A.; Papadopoulos, R.; Fuller, S.J.; Moir, R.D.; Small, D.; Beyreuther, K.; Masters, C.L. The amyloid protein precursor of Alzheimer’s disease is a mediator of the effects of nerve growth factor on neurite outgrowth. Neuron 1992, 9, 129–137. [Google Scholar] [CrossRef]
- Kumamoto, N.; Gu, Y.; Wang, J.; Janoschka, S.; Takemaru, K.; Levine, J.; Ge, S. A role for primary cilia in glutamatergic synaptic integration of adult-born neurons. Nat. Neurosci. 2012, 15, 399–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, V.; Lim, D.A.; Dahmane, N.; Sanchez, P.; Brionne, T.C.; Herzberg, C.D.; Gitton, Y.; Carleton, A.; Alvarez-Buylla, A.; Ruizi Altaba, A. Sonic hedgehog controls stem cell behavior in the postnatal and adult brain. Development 2005, 132, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breunig, J.J.; Sarkisian, M.R.; Arellano, J.I.; Morozov, Y.M.; Ayoub, A.E.; Sojitra, S.; Wang, B.; Flavell, R.A.; Rakic, P.; Town, T. Primary cilia regulate hippocampal neurogenesis by mediating sonic hedgehog signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 13127–13132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarthy, B.; Gaudet, C.; Menard, M.; Brown, L.; Atkinson, T.; Laferla, F.M.; Ito, S.; Armato, U.; Dal Pra, I.; Whitfield, J. Reduction of the immunostainable length of the hippocampal dentate granule cells’ primary cilia in 3xAD-transgenic mice producing human Abeta(1-42) and tau. Biochem. Biophys. Res. Commun. 2012, 427, 218–222. [Google Scholar] [CrossRef]
- Miyoshi, K.; Kasahara, K.; Miyazaki, I.; Asanuma, M. Factors that influence primary cilium length. Acta Med. Okayama 2011, 65, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, S.R.; Kretschmer, V.; Brucker, L.; Schneider, S.; Volz, A.K.; Oancea-Castillo, L.D.R.; May-Simera, H.L. Bardet-Biedl Syndrome proteins regulate cilia disassembly during tissue maturation. Cell. Mol. Life. Sci. 2019, 76, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Kyun, M.L.; Kim, S.O.; Lee, H.G.; Hwang, J.A.; Hwang, J.; Soung, N.K.; Cha-Molstad, H.; Lee, S.; Kwon, Y.T.; Kim, B.Y.; et al. Wnt3a Stimulation Promotes Primary Ciliogenesis through beta-Catenin Phosphorylation-Induced Reorganization of Centriolar Satellites. Cell Rep. 2020, 30, 1447–1462.e1445. [Google Scholar] [CrossRef]
- Jia, L.; Piña-Crespo, J.; Li, Y. Restoring Wnt/β-catenin signaling is a promising therapeutic strategy for Alzheimer’s disease. Mol. Brain 2019, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Einstein, E.B.; Patterson, C.A.; Hon, B.J.; Regan, K.A.; Reddi, J.; Melnikoff, D.E.; Mateer, M.J.; Schulz, S.; Johnson, B.N.; Tallent, M.K. Somatostatin signaling in neuronal cilia is critical for object recognition memory. J. Neurosci. 2010, 30, 4306–4314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guennewig, B.; Lim, J.; Marshall, L.; McCorkindale, A.N.; Paasila, P.J.; Patrick, E.; Kril, J.J.; Halliday, G.M.; Cooper, A.A.; Sutherland, G.T. Defining early changes in Alzheimer’s disease from RNA sequencing of brain regions differentially affected by pathology. Sci. Rep. 2021, 11, 4865. [Google Scholar] [CrossRef]
- Gonkowski, S.; Rytel, L. Somatostatin as an Active Substance in the Mammalian Enteric Nervous System. Int. J. Mol. Sci. 2019, 20, 4461. [Google Scholar] [CrossRef] [Green Version]
- Tobin, J.L.; Di Franco, M.; Eichers, E.; May-Simera, H.; Garcia, M.; Yan, J.; Quinlan, R.; Justice, M.J.; Hennekam, R.C.; Briscoe, J.; et al. Inhibition of neural crest migration underlies craniofacial dysmorphology and Hirschsprung’s disease in Bardet–Biedl syndrome. Proc. Natl. Acad. Sci. USA 2008, 105, 6714–6719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, J.M.; Brooks, E.M.; Mawe, G.M. Gastrointestinal Motility Monitor (GIMM). J. Vis. Exp. 2010, 46, 2435. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.H.; Ngwainmbi, J.; Grider, J.R.; Dewey, W.L.; Akbarali, H.I. An in-vitro preparation of isolated enteric neurons and glia from the myenteric plexus of the adult mouse. J. Vis. Exp. 2013, 78, 505688. [Google Scholar] [CrossRef]
- Smit, M.; Leng, J.; Klemke, R.L. Assay for neurite outgrowth quantification. Biotechniques 2003, 35, 254–256. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | 5′-3′ |
|---|---|---|
| β-Catenin | forward | ACTGCTGGGACTCTG |
| reverse | TGATGGCGTAGAACAG | |
| Lef1 | forward | GTCCCTTTCTCCACCCATC |
| reverse | AAGTGCTCGTCGCTGTAG | |
| Kif7 | forward | CTGGAGAAGGACTAGGTCG |
| reverse | TTTCCAGGCAGACGCTTCTC | |
| Smo | forward | GCTGCCACTTCTATGACTTCT |
| reverse | GCCGATTCTTGATCTCACAGT | |
| Hes1 | forward | CGAGCCTGTTGGGGAAATAC |
| reverse | GGTAGGTCATGGCGTTGATC | |
| Hes5 | forward | CTGGTGCAGGCTCTTGG |
| reverse | AAACAAGTACCGTGGCGGTGGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, V.T.T.; Brücker, L.; Volz, A.-K.; Baumgärtner, J.C.; dos Santos Guilherme, M.; Valeri, F.; May-Simera, H.; Endres, K. Primary Cilia Structure Is Prolonged in Enteric Neurons of 5xFAD Alzheimer’s Disease Model Mice. Int. J. Mol. Sci. 2021, 22, 13564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413564
Nguyen VTT, Brücker L, Volz A-K, Baumgärtner JC, dos Santos Guilherme M, Valeri F, May-Simera H, Endres K. Primary Cilia Structure Is Prolonged in Enteric Neurons of 5xFAD Alzheimer’s Disease Model Mice. International Journal of Molecular Sciences. 2021; 22(24):13564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413564
Chicago/Turabian StyleNguyen, Vu Thu Thuy, Lena Brücker, Ann-Kathrin Volz, Julia C. Baumgärtner, Malena dos Santos Guilherme, Francesco Valeri, Helen May-Simera, and Kristina Endres. 2021. "Primary Cilia Structure Is Prolonged in Enteric Neurons of 5xFAD Alzheimer’s Disease Model Mice" International Journal of Molecular Sciences 22, no. 24: 13564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413564