
Tryptophan Pathway Abnormalities in a Murine Model of Hereditary Glaucoma

 , , , ,
, , , ,  and
and 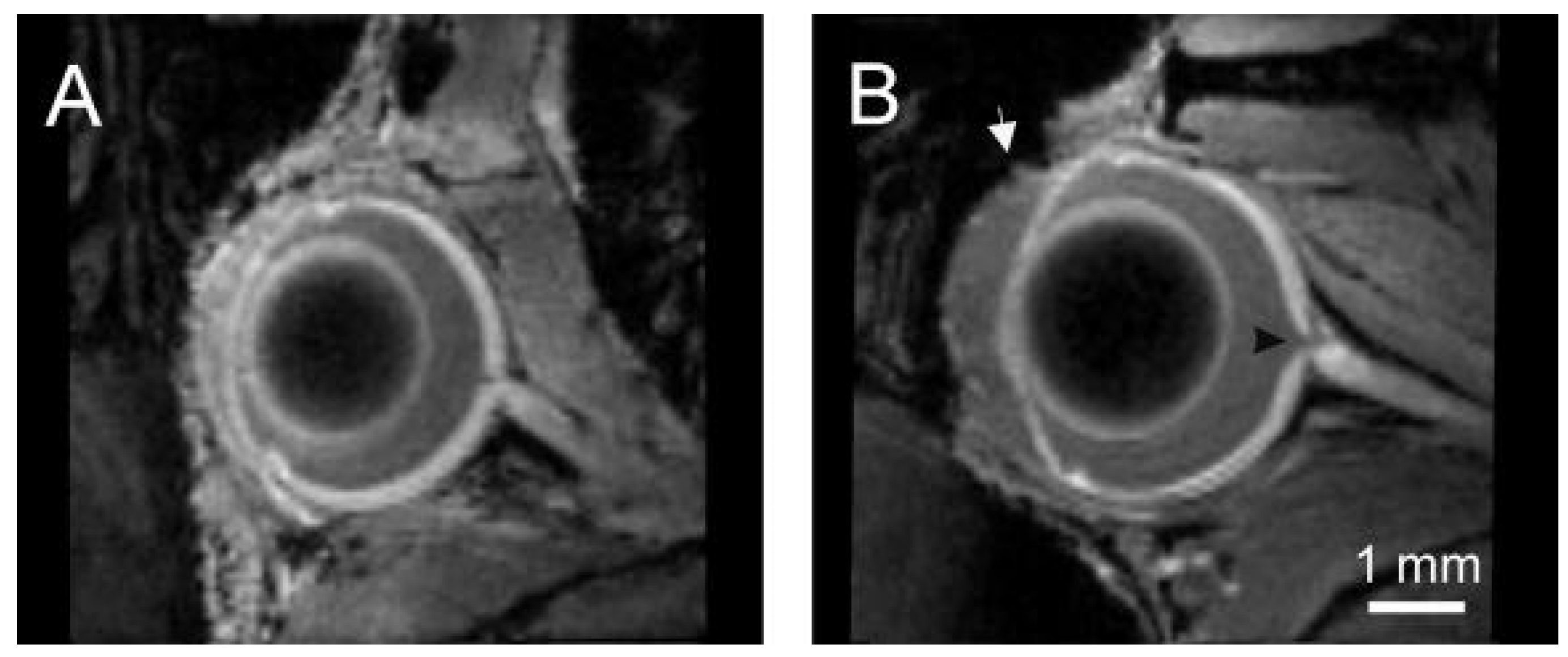{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Retina
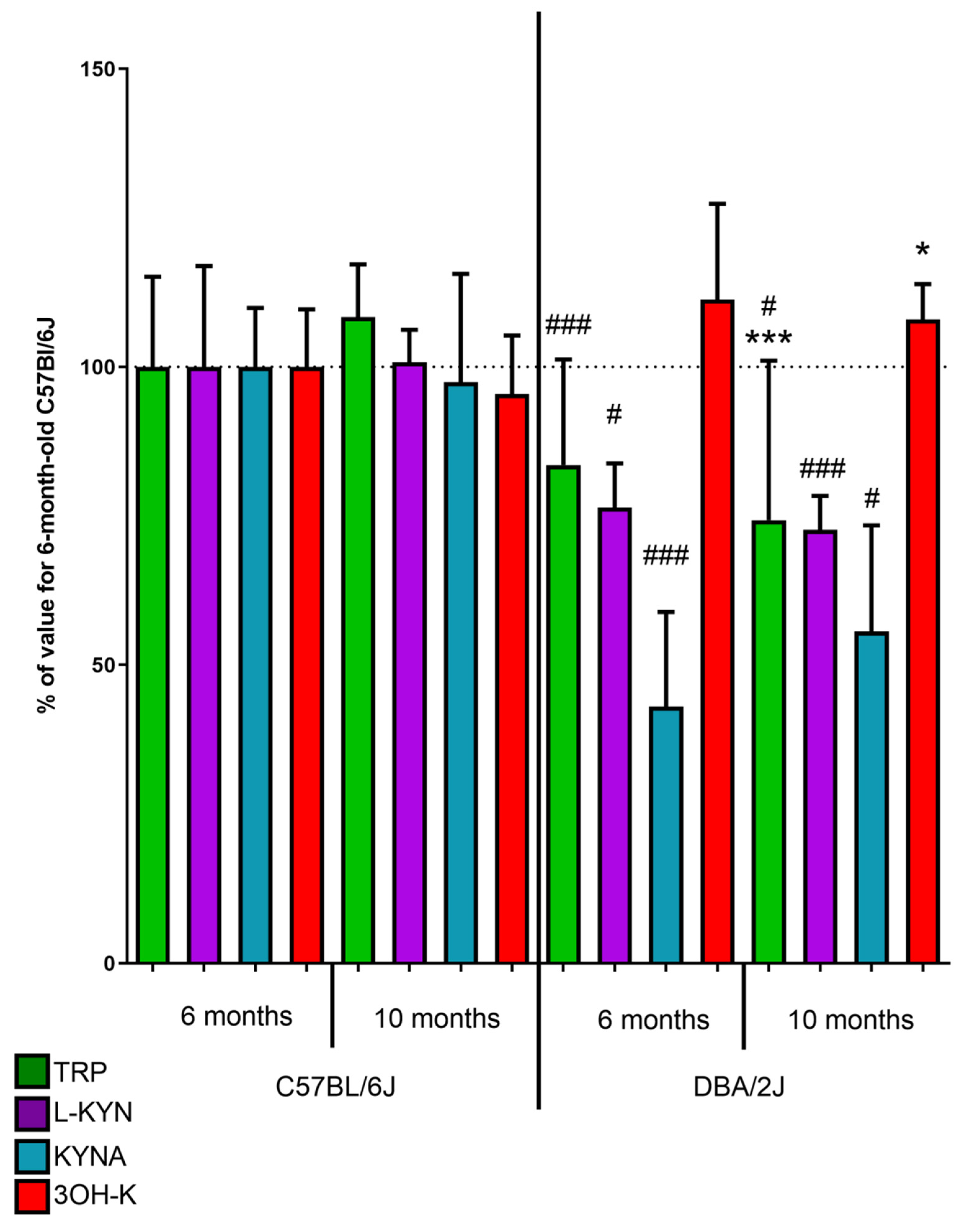
2.1.1. Kynurenic Acid
2.1.2. 3-Hydroxykynurenine
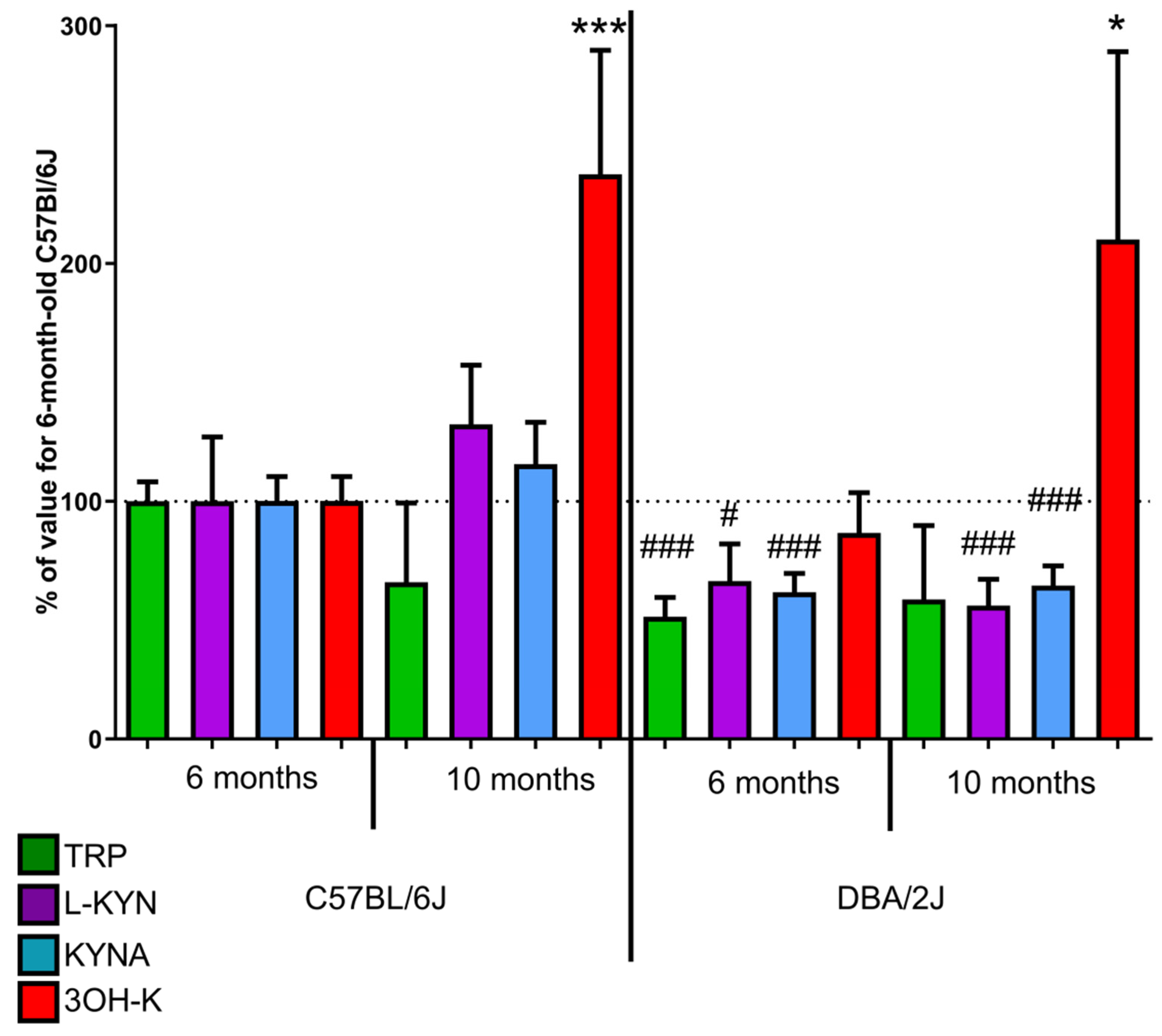
2.1.3. Tryptophan
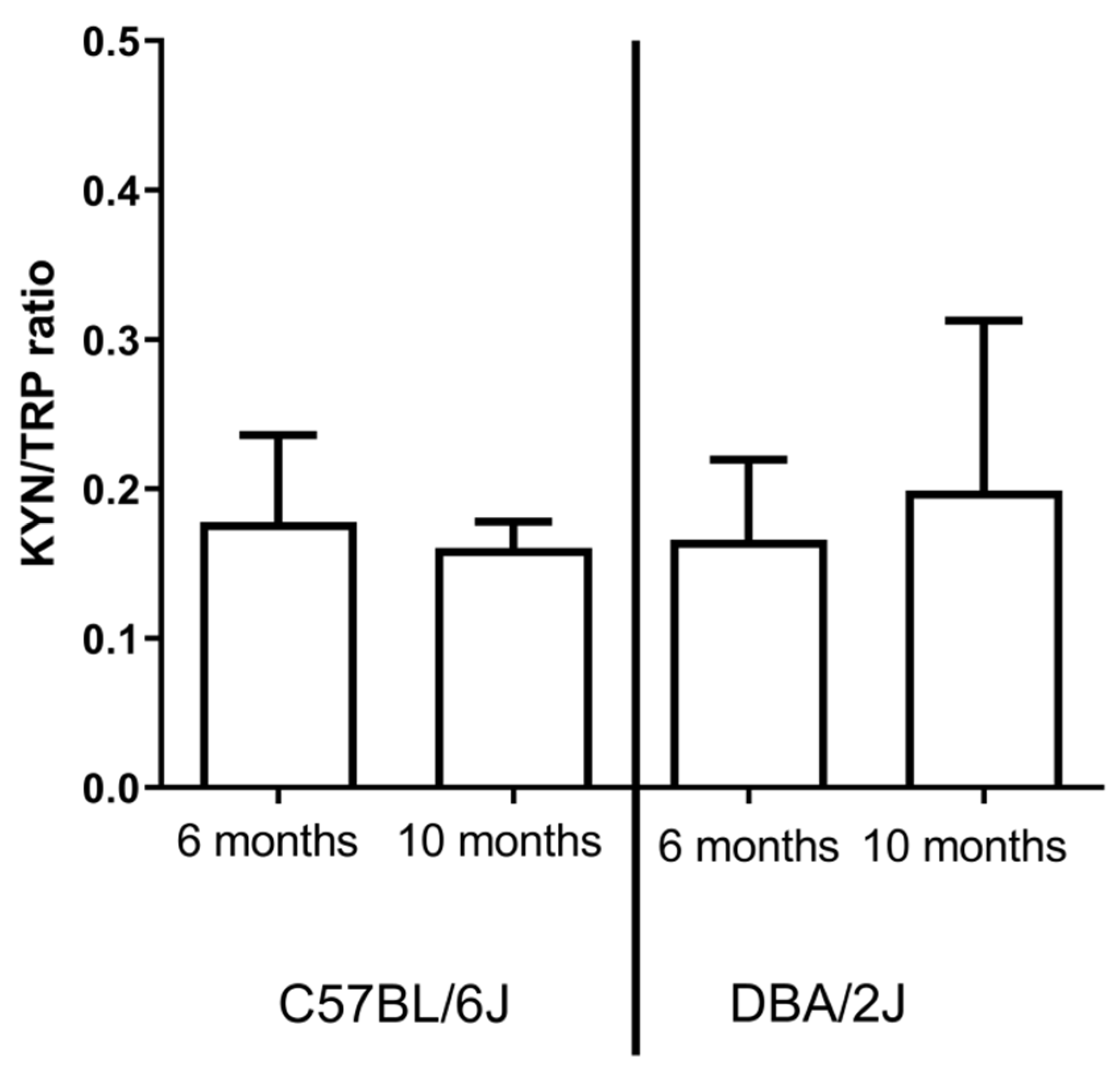
2.1.4. L-Kynurenine
2.2. Serum
2.2.1. Kynurenic acid
2.2.2. 3-Hydroxykynurenine
2.2.3. Tryptophan
2.2.4. L-Kynurenine
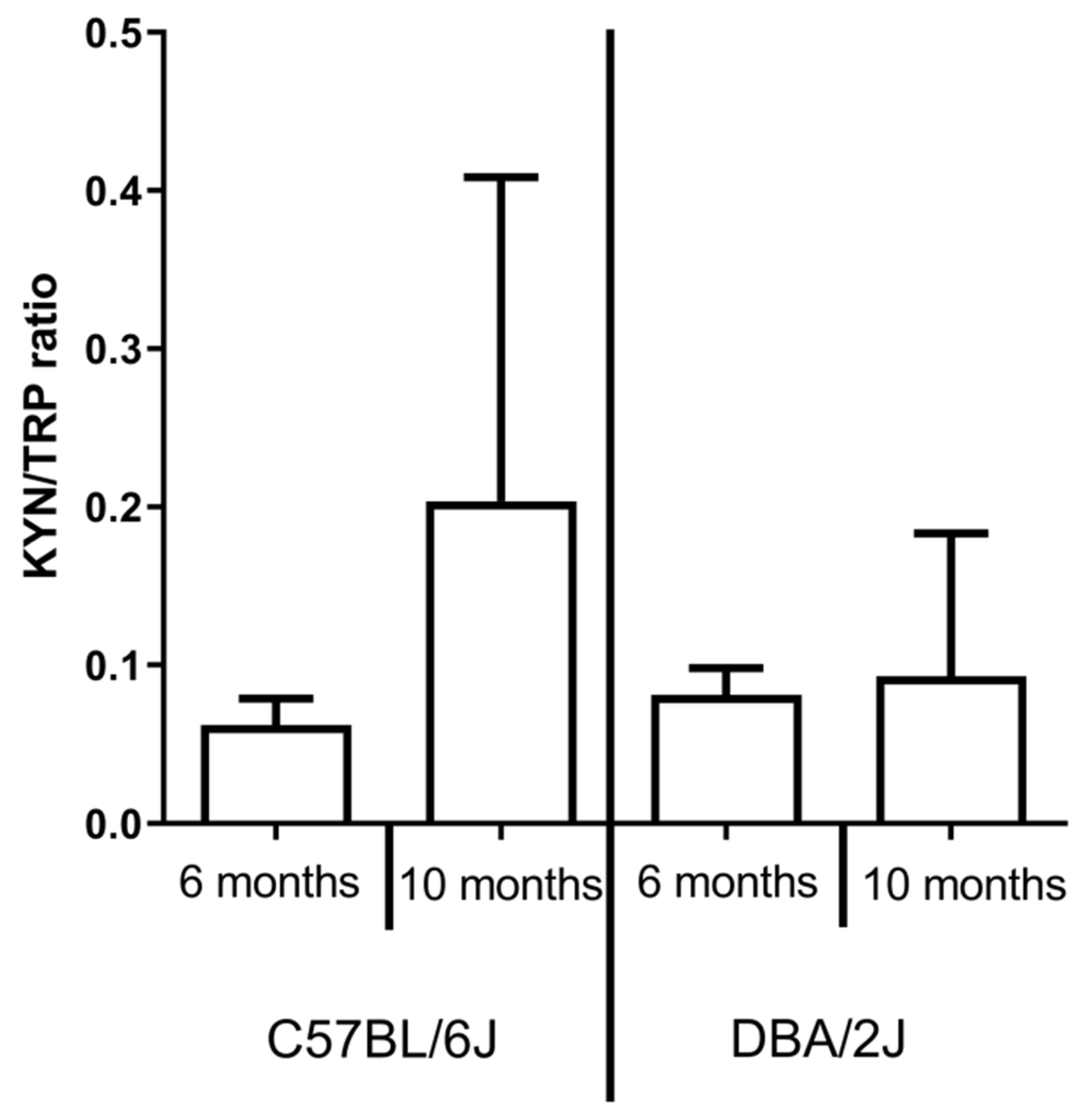
2.3. IDO activity
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Sample Collection
4.3. Determination of Tryptophan and its Metabolites
4.4. Visualization of the Neural Retina and Ocular Morphology with Magnetic Resonance Imaging
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tham, Y.-C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.-Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Posarelli, C.; Ortenzio, P.; Ferreras, A.; Toro, M.D.; Passani, A.; Loiudice, P.; Oddone, F.; Casini, G.; Figus, M. Twenty-Four-Hour Contact Lens Sensor Monitoring of Aqueous Humor Dynamics in Surgically or Medically Treated Glaucoma Patients. J. Ophthalmol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posarelli, C.; Toro, M.D.; Rejdak, R.; Żarnowski, T.; Pożarowska, D.; Longo, A.; Miccoli, M.; Nardi, M.; Figus, M. Safety and Efficacy of Second Ahmed Valve Implant in Refractory Glaucoma. J. Clin. Med. 2020, 9, 2039. [Google Scholar] [CrossRef] [PubMed]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Longo, A.; Avitabile, T.; Uva, M.G.; Bonfiglio, V.; Russo, A.; Toro, M.D.; Faro, S.; Reibaldi, M. Morphology of the optic nerve head in glaucomatous eyes with visual field defects in superior or inferior hemifield. Eur. J. Ophthalmol. 2018, 28, 175–181. [Google Scholar] [CrossRef]
- Kasi, A.; Faiq, M.A.; Chan, K.C. In vivo imaging of structural, metabolic and functional brain changes in glaucoma. Neural Regen. Res. 2019, 14, 446–449. [Google Scholar] [CrossRef]
- Lawlor, M.; Danesh-Meyer, H.; Levin, L.A.; Davagnanam, I.; De Vita, E.; Plant, G.T. Glaucoma and the brain: Trans-synaptic degeneration, structural change, and implications for neuroprotection. Surv. Ophthalmol. 2018, 63, 296–306. [Google Scholar] [CrossRef]
- Davis, B.M.; Crawley, L.; Pahlitzsch, M.; Javaid, F.; Cordeiro, M.F. Glaucoma: The retina and beyond. Acta Neuropathol. 2016, 132, 807–826. [Google Scholar] [CrossRef] [Green Version]
- Guillemin, G.J. Quinolinic acid: Neurotoxicity. FEBS J. 2012, 279, 1356–1365. [Google Scholar] [CrossRef]
- Ferreira, F.S.; Biasibetti-Brendler, H.; Pierozan, P.; Schmitz, F.; Bertó, C.G.; Prezzi, C.A.; Manfredini, V.; Wyse, A.T.S. Kynurenic acid restores Nrf2 levels and prevents quinolinic acid-induced toxicity in rat striatal slices. Mol. Neurobiol. 2018, 55, 8538–8549. [Google Scholar] [CrossRef]
- Pierozan, P.; Biasibetti-Brendler, H.; Schmitz, F.; Ferreira, F.; Pessoa-Pureur, R.; Wyse, A.T.S. Kynurenic acid prevents cytoskeletal disorganization induced by quinolinic acid in mixed cultures of Rat Striatum. Mol. Neurobiol. 2017, 55, 5111–5124. [Google Scholar] [CrossRef] [PubMed]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Rejdak, R.; Żarnowski, T.; Turski, W.A.; Kocki, T.; Zagorski, Z.; Zrenner, E.; Schuettauf, F. Alterations of kynurenic acid content in the retina in response to retinal ganglion cell damage. Vis. Res. 2003, 43, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Fiedorowicz, M.; Choragiewicz, T.; Thaler, S.; Schuettauf, F.; Nowakowska, D.; Wojtunik, K.; Reibaldi, M.; Avitabile, T.; Kocki, T.; Turski, W.A.; et al. Tryptophan and kynurenine pathway metabolites in animal models of retinal and optic nerve damage: Different dynamics of changes. Front. Physiol. 2019, 10, 1254. [Google Scholar] [CrossRef]
- Nahomi, R.B.; Nam, M.-H.; Rankenberg, J.; Rakete, S.; Houck, J.A.; Johnson, G.C.; Stankowska, D.L.; Pantcheva, M.B.; MacLean, P.S.; Nagaraj, R.H. Kynurenic acid protects against ischemia/reperfusion-induced retinal ganglion cell death in mice. Int. J. Mol. Sci. 2020, 21, 1795. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; van der Merwe, Y.; Sims, J.; Parra, C.; Ho, L.C.; Schuman, J.S.; Wollstein, G.; Lathrop, K.L.; Chan, K.C. Age-related changes in eye, brain and visuomotor behavior in the DBA/2J mouse model of chronic glaucoma. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Crosbie, D.E.; Keaney, J.; Tam, L.C.S.; Stamer, W.D.; Campbell, M.; Humphries, P. Age-related changes in eye morphology and aqueous humor dynamics in DBA/2J mice using contrast-enhanced ocular MRI. Magn. Reson. Imaging 2019, 59, 10–16. [Google Scholar] [CrossRef]
- Turner, A.J.; Wall, R.V.; Gupta, V.; Klistorner, A.; Graham, S.L. DBA/2J mouse model for experimental glaucoma: Pitfalls and problems. Clin. Exp. Ophthalmol. 2017, 45, 911–922. [Google Scholar] [CrossRef]
- Cheng, H.; Nair, G.; Walker, T.A.; Kim, M.K.; Pardue, M.T.; Thulé, P.M.; Olson, D.E.; Duong, T.Q. Structural and functional MRI reveals multiple retinal layers. Proc. Natl. Acad. Sci. USA 2006, 103, 17525–17530. [Google Scholar] [CrossRef] [Green Version]
- Porciatti, V.; Chou, T.-H.; Feuer, W.J. C57BL/6J, DBA/2J, and DBA/2J.Gpnmb+ mice have different visual signal processing in the inner retina. Mol. Vis. 2010, 16, 2939–2947. [Google Scholar]
- Schuettauf, F.; Rejdak, R.; Walski, M.; Frontczak-Baniewicz, M.; Voelker, M.; Blatsios, G.; Shinoda, K.; Zagorski, Z.; Zrenner, E.; Grieb, P. Retinal neurodegeneration in the DBA/2J mouse?a model for ocular hypertension. Acta Neuropathol. 2004, 107, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Fiedorowicz, M.; Orzel, J.; Kossowski, B.; Welniak-Kaminska, M.; Choragiewicz, T.; Swiatkiewicz, M.; Rejdak, R.; Bogorodzki, P.; Grieb, P. Anterograde transport in axons of the retinal ganglion cells and its relationship to the intraocular pressure during aging in mice with hereditary pigmentary glaucoma. Curr. Eye Res. 2017, 43, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Dengler-Crish, C.M.; Smith, M.A.; Inman, D.M.; Wilson, G.N.; Young, J.W.; Crish, S.D. Anterograde transport blockade precedes deficits in retrograde transport in the visual projection of the DBA/2J mouse model of glaucoma. Front. Neurosci. 2014, 8, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rejdak, R.; Kohler, K.; Kocki, T.; Shenk, Y.; Turski, W.A.; Okuno, E.; Lehaci, C.; Zagorski, Z.; Zrenner, E.; Schuettauf, F. Age-dependent decrease of retinal kynurenate and kynurenine aminotransferases in DBA/2J mice, a model of ocular hypertension. Vis. Res. 2004, 44, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Schuettauf, F.; Thaler, S.; Bolz, S.; Fries, J.; Kalbacher, H.; Mankowska, A.; Zurakowski, D.; Zrenner, E.; Rejdak, R. Alterations of amino acids and glutamate transport in the DBA/2J mouse retina; possible clues to degeneration. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 1157–1168. [Google Scholar] [CrossRef]
- John, S.W.; Smith, R.S.; Savinova, O.V.; Hawes, N.L.; Chang, B.; Turnbull, D.; Davisson, M.; Roderick, T.H.; Heckenlively, J.R. Essential iris atrophy, pigment dispersion, and glaucoma in DBA/2J mice. Investig. Ophthalmol. Vis. Sci. 1998, 39, 951–962. [Google Scholar]
- Fiedorowicz, M.; Wełniak-Kamińska, M.; Świątkiewicz, M.; Orzeł, J.; Chorągiewicz, T.; Toro, M.D.; Rejdak, R.; Bogorodzki, P.; Grieb, P. Changes of ocular dimensions as a marker of disease progression in a murine model of pigmentary glaucoma. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- De Lara, M.J.P.; Santano, C.; Guzman-Aranguez, A.; Valiente-Soriano, F.J.; Avilés-Trigueros, M.; Vidal-Sanz, M.; de la Villa, P.; Pintor, J. Assessment of inner retina dysfunction and progressive ganglion cell loss in a mouse model of glaucoma. Exp. Eye Res. 2014, 122, 40–49. [Google Scholar] [CrossRef]
- Scholz, M.; Buder, T.; Seeber, S.; Adamek, E.; Becker, C.-M.; Lütjen-Drecoll, E. Dependency of intraocular pressure elevation and glaucomatous changes in DBA/2J and DBA/2J-Rj Mice. Investig. Opthalmol. Vis. Sci. 2008, 49, 613–621. [Google Scholar] [CrossRef]
- Libby, R.T.; Anderson, M.G.; Pang, I.-H.; Robinson, Z.H.; Savinova, O.V.; Cosma, I.M.; Snow, A.; Wilson, L.A.; Smith, R.S.; Clark, A.F.; et al. Inherited glaucoma in DBA/2J mice: Pertinent disease features for studying the neurodegeneration. Vis. Neurosci. 2005, 22, 637–648. [Google Scholar] [CrossRef]
- Chou, T.-H.; Kocaoglu, O.P.; Borja, D.; Ruggeri, M.; Uhlhorn, S.R.; Manns, F.; Porciatti, V. Postnatal elongation of eye size in dba/2j mice compared with C57BL/6J Mice: In vivo analysis with whole-eye oct. Investig. Opthalmolg. Vis. Sci. 2011, 52, 3604–3612. [Google Scholar] [CrossRef]
- Danias, J.; Lee, K.C.; Zamora, M.-F.; Chen, B.; Shen, F.; Filippopoulos, T.; Su, Y.; Goldblum, D.; Podos, S.M.; Mittag, T. Quantitative analysis of retinal ganglion cell (RGC) loss in aging DBA/2NNia glaucomatous mice: Comparison with RGC loss in aging C57/BL6 mice. Investig. Opthalmol. Vis. Sci. 2003, 44, 5151–5162. [Google Scholar] [CrossRef] [Green Version]
- Grillo, S.L.; Montgomery, C.L.; Johnson, H.M.; Koulen, P. Quantification of changes in visual function during disease development in a mouse model of pigmentary glaucoma. J. Glaucoma 2018, 27, 828–841. [Google Scholar] [CrossRef]
- Wilson, G.N.; Smith, M.A.; Inman, D.M.; Dengler-Crish, C.M.; Crish, S.D. Early cytoskeletal protein modifications precede overt structural degeneration in the DBA/2J mouse model of Glaucoma. Front. Neurosci. 2016, 10, 494. [Google Scholar] [CrossRef]
- Wong, A.A.; Brown, R.E. Age-related changes in visual acuity, learning and memory in C57BL/6J and DBA/2J mice. Neurobiol. Aging 2007, 28, 1577–1593. [Google Scholar] [CrossRef]
- Cone, F.E.; Steinhart, M.R.; Oglesby, E.N.; Kalesnykas, G.; Pease, M.E.; Quigley, H.A. The effects of anesthesia, mouse strain and age on intraocular pressure and an improved murine model of experimental glaucoma. Exp. Eye Res. 2012, 99, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.S.; Roderick, T.H.; Sundberg, J.P. Microphthalmia and associated abnormalities in inbred black mice. Lab. Anim. Sci. 1994, 44, 551–560. [Google Scholar]
- Fukui, S.; Schwarcz, R.; Rapoport, S.I.; Takada, Y.; Smith, Q.R. Blood-brain barrier transport of kynurenines: Implications for brain synthesis and metabolism. J. Neurochem. 1991, 56, 2007–2017. [Google Scholar] [CrossRef]
- Fernandez-Sanchez, L.; Perez de Sevilla Muller, L.; Brecha, N.; Cuenca, N. Morphological and functional study of retinal astrocytes in DBA/2J Mice. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5096. [Google Scholar]
- Inman, D.; Buckingham, B.P.; Sappington, R.M.; Calkins, D.J.; Horner, P.J. Retinal vessel volume and pericyte number decrease over time in the DBA/2 mouse model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1320. [Google Scholar]
- Hu, P.; Hunt, N.H.; Arfuso, F.; Shaw, L.C.; Uddin, M.N.; Zhu, M.; Devasahayam, R.; Adamson, S.J.; Benson, V.L.; Chan-Ling, T.; et al. Increased indoleamine 2,3-dioxygenase and quinolinic acid expression in microglia and müller cells of diabetic human and rodent retina. Investig. Opthalmol. Vis. Sci. 2017, 58, 5043–5055. [Google Scholar] [CrossRef] [Green Version]
- Colín-González, A.L.; Maldonado, P.D.; Santamaría, A. 3-Hydroxykynurenine: An intriguing molecule exerting dual actions in the Central Nervous System. Neurotoxicology 2013, 34, 189–204. [Google Scholar] [CrossRef]
- Parrott, J.M.; O’Connor, J.C. Kynurenine 3-Monooxygenase: An influential mediator of neuropathology. Front. Psychiatry 2015, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Bosco, A.; Romero, C.O.; Breen, K.T.; Chagovetz, A.A.; Steele, M.R.; Ambati, B.K.; Vetter, M.L. Neurodegeneration severity can be predicted from early microglia alterations monitored in vivo in a mouse model of chronic glaucoma. Dis. Model. Mech. 2015, 8, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Suthakar, K.; Ryugo, D.K. Descending projections from the inferior colliculus to medial olivocochlear efferents: Mice with normal hearing, early onset hearing loss, and congenital deafness. Hear. Res. 2017, 343, 34–49. [Google Scholar] [CrossRef]
- Faingold, C.L.; Randall, M.; Mhaskar, Y.; Uteshev, V.V. Differences in serotonin receptor expression in the brainstem may explain the differential ability of a serotonin agonist to block seizure-induced sudden death in DBA/2 vs. DBA/1 mice. Brain Res. 2011, 1418, 104–110. [Google Scholar] [CrossRef]
- Matsushima, Y.; Imai, T.; Watanabe, O.; Kawahara, H.; Ohne, M.; Takai, H. Spontaneous calcified tongue lesions in DBA Mice. Exp. Anim. 1984, 33, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Wirthgen, E.; Hoeflich, A.; Rebl, A.; Günther, J. Kynurenic Acid: The janus-faced role of an immunomodulatory tryptophan metabolite and its link to pathological conditions. Front. Immunol. 2018, 8, 1957. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, K.R.; Castellano-Gonzalez, G.; Guillemin, G.J.; Lovejoy, D.B. Major developments in the design of inhibitors along the kynurenine pathway. Curr. Med. Chem. 2017, 24, 2471–2495. [Google Scholar] [CrossRef] [Green Version]
- Harper, M.M.; Woll, A.W.; Evans, L.P.; Delcau, M.; Akurathi, A.; Hedberg-Buenz, A.; Soukup, D.A.; Boehme, N.; Hefti, M.M.; Dutca, L.M.; et al. Blast preconditioning protects retinal ganglion cells and reveals targets for prevention of neurodegeneration following blast-mediated traumatic brian injury. Investig. Opthalmol. Vis. Sci. 2019, 60, 4159–4170. [Google Scholar] [CrossRef] [Green Version]
- Borisova, M.A.; Snytnikova, O.A.; Litvinova, E.A.; Achasova, K.M.; Babochkina, T.I.; Pindyurin, A.V.; Tsentalovich, Y.P.; Kozhevnikova, E.N. Fucose ameliorates tryptophan metabolism and behavioral abnormalities in a mouse model of chronic colitis. Nutrients 2020, 12, 445. [Google Scholar] [CrossRef] [Green Version]
- Cseh, E.K.; Veres, G.; Szentirmai, M.; Nánási, N.; Szatmári, I.; Fülöp, F.; Vécsei, L.; Zádori, D. HPLC method for the assessment of tryptophan metabolism utilizing separate internal standard for each detector. Anal. Biochem. 2019, 574, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, J.; Kamata, S.; Miura, A.; Nagata, T.; Kainuma, R.; Ishii, I. Differential adaptive responses to 1- or 2-day fasting in various mouse tissues revealed by quantitative PCR analysis. FEBS Open Bio 2015, 5, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Gao, P.; Zhu, D. Optimization of Zn2+-containing mobile phase for simultaneous determination of kynurenine, kynurenic acid and tryptophan in human plasma by high performance liquid chromatography. J. Chromatogr. B 2010, 878, 603–608. [Google Scholar] [CrossRef]
- Heyes, M.P.; Quearry, B.J. Quantification of 3-hydroxykynurenine in brain by high-performance liquid chromatography and electrochemical detection. J. Chromatogr. B Biomed. Sci. Appl. 1988, 428, 340–344. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiedorowicz, M.; Choragiewicz, T.; Turski, W.A.; Kocki, T.; Nowakowska, D.; Wertejuk, K.; Kamińska, A.; Avitabile, T.; Wełniak-Kaminska, M.; Grieb, P.; et al. Tryptophan Pathway Abnormalities in a Murine Model of Hereditary Glaucoma. Int. J. Mol. Sci. 2021, 22, 1039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031039
Fiedorowicz M, Choragiewicz T, Turski WA, Kocki T, Nowakowska D, Wertejuk K, Kamińska A, Avitabile T, Wełniak-Kaminska M, Grieb P, et al. Tryptophan Pathway Abnormalities in a Murine Model of Hereditary Glaucoma. International Journal of Molecular Sciences. 2021; 22(3):1039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031039
Chicago/Turabian StyleFiedorowicz, Michal, Tomasz Choragiewicz, Waldemar A. Turski, Tomasz Kocki, Dominika Nowakowska, Kamila Wertejuk, Agnieszka Kamińska, Teresio Avitabile, Marlena Wełniak-Kaminska, Pawel Grieb, and et al. 2021. "Tryptophan Pathway Abnormalities in a Murine Model of Hereditary Glaucoma" International Journal of Molecular Sciences 22, no. 3: 1039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031039