
Drug Reaction with Eosinophilia and Systemic Symptoms: A Complex Interplay between Drug, T Cells, and Herpesviridae

Abstract
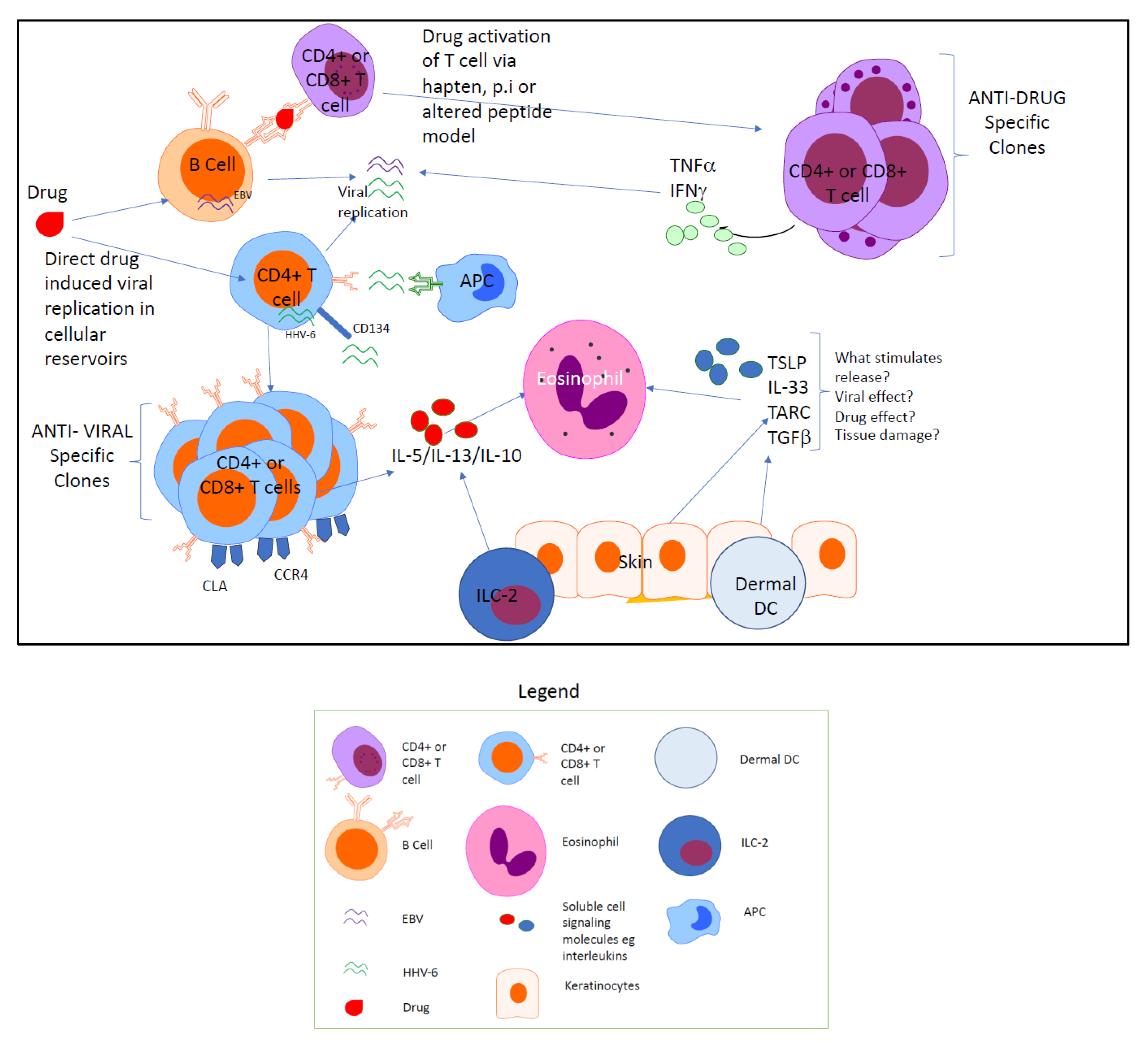
:1. Introduction
2. Models of T Cell-Mediated Hypersensitivity
3. T Cell Repertoire Ontogeny and the Role of Human Herpesviridae (HHV)
4. HHV Replication and DRESS
5. T Cell Effectors Are Context-Specific and Correspond to the Stage of DRESS
6. Conclusions
- (1)
- Generation of specific anti-drug effector T cells via recognition through MHC and TCR interactions;
- (2)
- Direct drug-mediated HHV viral reactivation and replication in host reservoir cells generating anti-viral cytotoxic T cells.
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Hakkarainen, K.M.; Hedna, K.; Petzold, M.; Hägg, S. Percentage of patients with preventable adverse drug reactions and preventability of adverse drug reactions—A meta-analysis. PLoS ONE 2012, 7, e33236. [Google Scholar] [CrossRef] [PubMed]
- Kongkaew, C.; Noyce, P.R.; Ashcroft, D.M. Hospital admissions associated with adverse drug reactions: A systematic review of prospective observational studies. Ann. Pharmacother. 2008, 42, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Lazarou, J.; Pomeranz, B.H.; Corey, P.N. Incidence of Adverse Drug Reactions in Hospitalized PatientsA Meta-analysis of Prospective Studies. JAMA 1998, 279, 1200–1205. [Google Scholar] [CrossRef]
- Pirmohamed, M.; James, S.; Meakin, S.; Green, C.; Scott, A.K.; Walley, T.J.; Farrar, K.; Park, B.K.; Breckenridge, A.M. Adverse drug reactions as cause of admission to hospital: Prospective analysis of 18 820 patients. BMJ 2004, 329, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Suh, D.C.; Woodall, B.S.; Shin, S.K.; Hermes-De Santis, E.R. Clinical and economic impact of adverse drug reactions in hospitalized patients. Ann. Pharmacother. 2000, 34, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.M. Textbook of Adverse Drug Reactions; Oxford University Press: Oxford, UK, 1977. [Google Scholar]
- Edwards, I.R.; Aronson, J.K. Adverse drug reactions: Definitions, diagnosis, and management. Lancet 2000, 356, 1255–1259. [Google Scholar] [CrossRef]
- Pavlos, R.; Mallal, S.; Phillips, E. HLA and pharmacogenetics of drug hypersensitivity. Pharmacogenomics 2012, 13, 1285–1306. [Google Scholar] [CrossRef] [Green Version]
- Tennis, P.; Stern, R.S. Risk of serious cutaneous disorders after initiation of use of phenytoin, carbamazepine, or sodium valproate: A record linkage study. Neurology 1997, 49, 542–546. [Google Scholar] [CrossRef]
- Guberman, A.H.; Besag, F.M.; Brodie, M.J.; Dooley, J.M.; Duchowny, M.S.; Pellock, J.M.; Richens, A.; Stern, R.S.; Trevathan, E. Lamotrigine-associated rash: Risk/benefit considerations in adults and children. Epilepsia 1999, 40, 985–991. [Google Scholar] [CrossRef]
- Kardaun, S.H.; Mockenhaupt, M.; Roujeau, J.C. Comments on: DRESS syndrome. J. Am. Acad. Dermatol. 2014, 71, 1000. [Google Scholar] [CrossRef]
- Picard, D.; Janela, B.; Descamps, V.; D’Incan, M.; Courville, P.; Jacquot, S.; Rogez, S.; Mardivirin, L.; Moins-Teisserenc, H.; Toubert, A.; et al. Drug reaction with eosinophilia and systemic symptoms (DRESS): A multiorgan antiviral T cell response. Sci. Transl. Med. 2010, 2, 46ra62. [Google Scholar] [CrossRef] [PubMed]
- Shiohara, T.; Iijima, M.; Ikezawa, Z.; Hashimoto, K. The diagnosis of a DRESS syndrome has been sufficiently established on the basis of typical clinical features and viral reactivations. Br. J. Dermatol. 2007, 156, 1083–1084. [Google Scholar] [CrossRef] [PubMed]
- Naisbitt, D.J.; Gordon, S.F.; Pirmohamed, M.; Burkhart, C.; Cribb, A.E.; Pichler, W.J.; Park, B.K. Antigenicity and immunogenicity of sulphamethoxazole: Demonstration of metabolism-dependent haptenation and T-cell proliferation in vivo. Br. J. Pharmacol. 2001, 133, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.K.; Naisbitt, D.J.; Gordon, S.F.; Kitteringham, N.R.; Pirmohamed, M. Metabolic activation in drug allergies. Toxicology 2001, 158, 11–23. [Google Scholar] [CrossRef]
- Pichler, W.; Yawalkar, N.; Schmid, S.; Helbling, A. Pathogenesis of drug-induced exanthems. Allergy 2002, 57, 884–893. [Google Scholar] [CrossRef]
- Ariza, A.; Mayorga, C.; Fernandez, T.D.; Barbero, N.; Martín-Serrano, A.; Pérez-Sala, D.; Sánchez-Gómez, F.J.; Blanca, M.; Torres, M.J.; Montanez, M.I. Hypersensitivity reactions to β-lactams: Relevance of hapten-protein conjugates. J. Investig. Allergol. Clin. Immunol. 2015, 25, 12–25. [Google Scholar]
- El-Ghaiesh, S.; Monshi, M.M.; Whitaker, P.; Jenkins, R.; Meng, X.; Farrell, J.; Elsheikh, A.; Peckham, D.; French, N.; Pirmohamed, M.; et al. Characterization of the antigen specificity of T-cell clones from piperacillin-hypersensitive patients with cystic fibrosis. J. Pharmacol. Exp. Ther. 2012, 341, 597–610. [Google Scholar] [CrossRef] [Green Version]
- Pichler, W.J. The p-i Concept: Pharmacological Interaction of Drugs With Immune Receptors. World Allergy Organ J. 2008, 1, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Pichler, W.; Watkins, S. Interaction of Small Molecules with Specific Immune Receptors: The p-i Concept and its Consequences. Curr. Immunol. Rev. 2014, 10, 7–18. [Google Scholar] [CrossRef]
- Schnyder, B.; Mauri-Hellweg, D.; Zanni, M.; Bettens, F.; Pichler, W.J. Direct, MHC-dependent presentation of the drug sulfamethoxazole to human alphabeta T cell clones. J. Clin. Investig. 1997, 100, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Illing, P.T.; Vivian, J.P.; Dudek, N.L.; Kostenko, L.; Chen, Z.; Bharadwaj, M.; Miles, J.J.; Kjer-Nielsen, L.; Gras, S.; Williamson, N.A.; et al. Immune self-reactivity triggered by drug-modified HLA-peptide repertoire. Nature 2012, 486, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Ostrov, D.A.; Grant, B.J.; Pompeu, Y.A.; Sidney, J.; Harndahl, M.; Southwood, S.; Oseroff, C.; Lu, S.; Jakoncic, J.; de Oliveira, C.A.; et al. Drug hypersensitivity caused by alteration of the MHC-presented self-peptide repertoire. Proc. Natl. Acad. Sci. USA 2012, 109, 9959–9964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutrell, A.G.; Hernandez, J.E.; Fleming, J.W.; Edwards, M.T.; Moore, M.A.; Brothers, C.H.; Scott, T.R. Updated clinical risk factor analysis of suspected hypersensitivity reactions to abacavir. Ann. Pharmacother. 2004, 38, 2171–2172. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.; Ward, K.M.; Stern, J.J. A near-fatal hypersensitivity reaction to abacavir: Case report and literature review. AIDS Read. 2001, 11, 222–226. [Google Scholar] [PubMed]
- Adam, J.; Eriksson, K.K.; Schnyder, B.; Fontana, S.; Pichler, W.J.; Yerly, D. Avidity determines T-cell reactivity in abacavir hypersensitivity. Eur. J. Immunol. 2012, 42, 1706–1716. [Google Scholar] [CrossRef]
- Adam, J.; Wuillemin, N.; Watkins, S.; Jamin, H.; Eriksson, K.K.; Villiger, P.; Fontana, S.; Pichler, W.J.; Yerly, D. Abacavir induced T cell reactivity from drug naïve individuals shares features of allo-immune responses. PLoS ONE 2014, 9, e95339. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.; Riley, B.T.; Kass, I.; Buckle, A.M.; Borg, N.A. The Role of Conformational Dynamics in Abacavir-Induced Hypersensitivity Syndrome. Sci. Rep. 2019, 9, 10523. [Google Scholar] [CrossRef]
- Hetherington, S.; Hughes, A.R.; Mosteller, M.; Shortino, D.; Baker, K.L.; Spreen, W.; Lai, E.; Davies, K.; Handley, A.; Dow, D.J.; et al. Genetic variations in HLA-B region and hypersensitivity reactions to abacavir. Lancet 2002, 359, 1121–1122. [Google Scholar] [CrossRef]
- Mallal, S.; Nolan, D.; Witt, C.; Masel, G.; Martin, A.M.; Moore, C.; Sayer, D.; Castley, A.; Mamotte, C.; Maxwell, D.; et al. Association between presence of HLA-B*5701, HLA-DR7, and HLA-DQ3 and hypersensitivity to HIV-1 reverse-transcriptase inhibitor abacavir. Lancet 2002, 359, 727–732. [Google Scholar] [CrossRef]
- Saag, M.; Balu, R.; Phillips, E.; Brachman, P.; Martorell, C.; Burman, W.; Stancil, B.; Mosteller, M.; Brothers, C.; Wannamaker, P.; et al. High Sensitivity of Human Leukocyte Antigen-B*5701 as a Marker for Immunologically Confirmed Abacavir Hypersensitivity in White and Black Patients. Clin. Infect. Dis. 2008, 46, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, J.; Zhao, L.-M.; Peng, W.; Shen, G.-Q.; Xue, L.; Zheng, X.-X.; He, X.-J.; Gong, C.-Y.; Miao, L.-Y. Strong association between HLA-B*1502 and carbamazepine-induced Stevens-Johnson syndrome and toxic epidermal necrolysis in mainland Han Chinese patients. Eur. J. Clin. Pharmacol. 2011, 67, 885. [Google Scholar] [CrossRef] [PubMed]
- Konvinse, K.C.; Trubiano, J.A.; Pavlos, R.; James, I.; Shaffer, C.M.; Bejan, C.A.; Schutte, R.J.; Ostrov, D.A.; Pilkinton, M.A.; Rosenbach, M.; et al. HLA-A*32:01 is strongly associated with vancomycin-induced drug reaction with eosinophilia and systemic symptoms. J. Allergy Clin. Immunol. 2019, 144, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshi, O.; Taisei, M.; Amara, Y.; Atsushi, T.; Michiaki, K.; Yuji, S.; Zenro, I.; Masafumi, I.; Tetsuo, S.; Koji, H.; et al. Genome-wide association study identifies HLA-A*3101 allele as a genetic risk factor for carbamazepine-induced cutaneous adverse drug reactions in Japanese population. Human Mol. Genet. 2011, 20, 1034–1041. [Google Scholar] [CrossRef]
- Agrawal, B. Heterologous Immunity: Role in Natural and Vaccine-Induced Resistance to Infections. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, R.M.; Selin, L.K. No one is naive: The significance of heterologous T-cell immunity. Nat. Rev. Immunol. 2002, 2, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Welsh, R.M.; Che, J.W.; Brehm, M.A.; Selin, L.K. Heterologous immunity between viruses. Immunol. Rev. 2010, 235, 244–266. [Google Scholar] [CrossRef] [Green Version]
- White, K.D.; Chung, W.H.; Hung, S.I.; Mallal, S.; Phillips, E.J. Evolving models of the immunopathogenesis of T cell-mediated drug allergy: The role of host, pathogens, and drug response. J. Allergy Clin. Immunol. 2015, 136, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Amir, A.L.; D’Orsogna, L.J.A.; Roelen, D.L.; van Loenen, M.M.; Hagedoorn, R.S.; de Boer, R.; van der Hoorn, M.A.W.G.; Kester, M.G.D.; Doxiadis, I.I.N.; Falkenburg, J.H.F.; et al. Allo-HLA reactivity of virus-specific memory T cells is common. Blood 2010, 115, 3146–3157. [Google Scholar] [CrossRef] [Green Version]
- D’Orsogna, L.J.A.; Roelen, D.L.; van der Meer-Prins, E.M.W.; van der Pol, P.; Franke-van Dijk, M.E.; Eikmans, M.; Anholts, J.; Rossjohn, J.; McCluskey, J.; Mulder, A.; et al. Tissue Specificity of Cross-Reactive Allogeneic Responses by EBV EBNA3A-Specific Memory T Cells. Transplantation 2011, 91, 494–500. [Google Scholar] [CrossRef]
- Deckers, J.G.M.; Boonstra, J.G.; Van der Kooij, S.W.; Daha, M.R.; Van der Woude, F.J. Tissue-specific Characteristics of Cytotoxic Grafit-infiltrating T Cells during Renal Allograft Rejection1. Transplantation 1997, 64, 178–181. [Google Scholar] [CrossRef]
- Almeida, C.A.; van Miert, P.; O’Driscoll, K.; Zoet, Y.M.; Chopra, A.; Witt, C.; John, M.; Claas, F.H.J.; D’Orsogna, L.J.A. Virus-specific T-cell clonotypes might contribute to drug hypersensitivity reactions through heterologous immunity. J. Allergy Clin. Immunol. 2019, 144, 608–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, A.; Lucas, M.; Strhyn, A.; Keane, N.M.; McKinnon, E.; Pavlos, R.; Moran, E.M.; Meyer-Pannwitt, V.; Gaudieri, S.; D’Orsogna, L.; et al. Abacavir-reactive memory T cells are present in drug naïve individuals. PLoS ONE 2015, 10, e0117160. [Google Scholar] [CrossRef] [Green Version]
- Sylwester, A.W.; Mitchell, B.L.; Edgar, J.B.; Taormina, C.; Pelte, C.; Ruchti, F.; Sleath, P.R.; Grabstein, K.H.; Hosken, N.A.; Kern, F.; et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J. Exp. Med. 2005, 202, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Komanduri, K.V.; Donahoe, S.M.; Moretto, W.J.; Schmidt, D.K.; Gillespie, G.; Ogg, G.S.; Roederer, M.; Nixon, D.F.; McCune, J.M. Direct measurement of CD4+ and CD8+ T-cell responses to CMV in HIV-1-infected subjects. Virology 2001, 279, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, P.; Khan, N. CD8(+) T-cell immunity to cytomegalovirus. Hum. Immunol. 2004, 65, 456–464. [Google Scholar] [CrossRef]
- Colf, L.; Bankovich, A.; Hanick, N.; Bowerman, N.; Jones, L.; Kranz, D.; Garcia, K. How a Single T Cell Receptor Recognizes Both Self and Foreign MHC. Cell 2007, 129, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Seishima, M.; Yamanaka, S.; Fujisawa, T.; Tohyama, M.; Hashimoto, K. Reactivation of human herpesvirus (HHV) family members other than HHV-6 in drug-induced hypersensitivity syndrome. Br. J. Dermatol. 2006, 155, 344–349. [Google Scholar] [CrossRef]
- Tohyama, M.; Hashimoto, K.; Yasukawa, M.; Kimura, H.; Horikawa, T.; Nakajima, K.; Urano, Y.; Matsumoto, K.; Iijima, M.; Shear, N.H. Association of human herpesvirus 6 reactivation with the flaring and severity of drug-induced hypersensitivity syndrome. Br. J. Dermatol. 2007, 157, 934–940. [Google Scholar] [CrossRef]
- Kano, Y.; Hiraharas, K.; Sakuma, K.; Shiohara, T. Several herpesviruses can reactivate in a severe drug-induced multiorgan reaction in the same sequential order as in graft-versus-host disease. Br. J. Dermatol. 2006, 155, 301–306. [Google Scholar] [CrossRef]
- Shiohara, T.; Inaoka, M.; Kano, Y. Drug-induced hypersensitivity syndrome (DIHS): A reaction induced by a complex interplay among herpesviruses and antiviral and antidrug immune responses. Allergol. Int. 2006, 55, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Descamps, V.; Valance, A.; Edlinger, C.; Fillet, A.M.; Grossin, M.; Lebrun-Vignes, B.; Belaich, S.; Crickx, B. Association of human herpesvirus 6 infection with drug reaction with eosinophilia and systemic symptoms. Arch. Dermatol. 2001, 137, 301–304. [Google Scholar] [PubMed]
- Niu, J.; Jia, Q.; Ni, Q.; Yang, Y.; Chen, G.; Yang, X.; Zhai, Z.; Yu, H.; Guan, P.; Lin, R.; et al. Association of CD8(+) T lymphocyte repertoire spreading with the severity of DRESS syndrome. Sci. Rep. 2015, 5, 9913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salahuddin, S.Z.; Ablashi, D.V.; Markham, P.D.; Josephs, S.F.; Sturzenegger, S.; Kaplan, M.; Halligan, G.; Biberfeld, P.; Wong-Staal, F.; Kramarsky, B.; et al. Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders. Science 1986, 234, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, F.; Nakamura, Y.; Ommori, R.; Miyashita, K.; Iioka, H.; Miyashita, N.; Nishikawa, M.; Himuro, Y.; Ogawa, K.; Asada, H. Predominant Contribution of CD4 T Cells to Human Herpesvirus 6 (HHV-6) Load in the Peripheral Blood of Patients with Drug-induced Hypersensitivity Syndrome and Persistent HHV-6 Infection. Acta Derm. Venereol. 2018, 98, 146–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prusty, B.K.; Gulve, N.; Chowdhury, S.R.; Schuster, M.; Strempel, S.; Descamps, V.; Rudel, T. HHV-6 encoded small non-coding RNAs define an intermediate and early stage in viral reactivation. npj Genom. Med. 2018, 3, 25. [Google Scholar] [CrossRef]
- Takahashi, R.; Kano, Y.; Yamazaki, Y.; Kimishima, M.; Mizukawa, Y.; Shiohara, T. Defective regulatory T cells in patients with severe drug eruptions: Timing of the dysfunction is associated with the pathological phenotype and outcome. J. Immunol. 2009, 182, 8071–8079. [Google Scholar] [CrossRef] [Green Version]
- Beutler, A.S.; Li, S.; Nicol, R.; Walsh, M.J. Carbamazepine is an inhibitor of histone deacetylases. Life Sci. 2005, 76, 3107–3115. [Google Scholar] [CrossRef]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J. Biol. Chem. 2001, 276, 36734–36741. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kobayashi, T.; Voisin, B.; Jo, J.-H.; Sakamoto, K.; Jin, S.-P.; Kelly, M.; Pasieka, H.B.; Naff, J.L.; Meyerle, J.H.; et al. Targeted therapy guided by single-cell transcriptomic analysis in drug-induced hypersensitivity syndrome: A case report. Nat. Med. 2020, 26, 236–243. [Google Scholar] [CrossRef]
- Miyagawa, F.; Nakamura, Y.; Miyashita, K.; Iioka, H.; Himuro, Y.; Ogawa, K.; Nishimura, C.; Nishikawa, M.; Mitsui, Y.; Ito, Y.; et al. Preferential expression of CD134, an HHV-6 cellular receptor, on CD4T cells in drug-induced hypersensitivity syndrome (DIHS)/drug reaction with eosinophilia and systemic symptoms (DRESS). J. Dermatol. Sci. 2016, 83, 151–154. [Google Scholar] [CrossRef]
- Watanabe, H. Recent Advances in Drug-Induced Hypersensitivity Syndrome/Drug Reaction with Eosinophilia and Systemic Symptoms. J. Immunol. Res. 2018, 2018, 5163129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, M.; Shimamoto, T.; Isegawa, Y.; Yamanishi, K. The R3 Region, One of Three Major Repetitive Regions of Human Herpesvirus 6, Is a Strong Enhancer of Immediate-Early Gene U95. J. Virol. 2001, 75, 10149–10160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummel, M.; Abecassis, M.M. A model for reactivation of CMV from latency. J. Clin. Virol. 2002, 25, S123–S136. [Google Scholar] [CrossRef]
- Uno, H.; Kabashima, K.; Tohyama, M.; Watanabe, T.; Hashimoto, K.; Iijima, M.; Sueki, H.; Watanabe, H. TNF-α as a useful predictor of human herpesvirus-6 reactivation and indicator of the disease process in drug-induced hypersensitivity syndrome (DIHS)/drug reaction with eosinophilia and systemic symptoms (DRESS). J. Dermatol. Sci. 2014, 74, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Kamijima, M.; Wang, H.; Yamanoshita, O.; Ito, Y.; Xia, L.; Yanagiba, Y.; Chen, C.; Okamura, A.; Huang, Z.; Qiu, X.; et al. Occupational trichloroethylene hypersensitivity syndrome: Human herpesvirus 6 reactivation and rash phenotypes. J. Dermatol. Sci. 2013, 72, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Ushigome, Y.; Mizukawa, Y.; Kimishima, M.; Yamazaki, Y.; Takahashi, R.; Kano, Y.; Shiohara, T. Monocytes are involved in the balance between regulatory T cells and Th17 cells in severe drug eruptions. Clin. Exp. Allergy 2018, 48, 1453–1463. [Google Scholar] [CrossRef]
- Shiohara, T.; Kano, Y. Drug reaction with eosinophilia and systemic symptoms (DRESS): Incidence, pathogenesis and management. Expert Opin. Drug Saf. 2017, 16, 139–147. [Google Scholar] [CrossRef]
- Yazicioglu, M.; Elmas, R.; Turgut, B.; Genchallac, T. The association between DRESS and the diminished numbers of peripheral B lymphocytes and natural killer cells. Pediatr. Allergy Immunol. 2012, 23, 289–296. [Google Scholar] [CrossRef]
- Kano, Y.; Seishima, M.; Shiohara, T. Hypogammaglobulinemia as an early sign of drug-induced hypersensitivity syndrome. J. Am. Acad. Dermatol. 2006, 55, 727–728. [Google Scholar] [CrossRef]
- Kano, Y.; Inaoka, M.; Shiohara, T. Association between anticonvulsant hypersensitivity syndrome and human herpesvirus 6 reactivation and hypogammaglobulinemia. Arch. Dermatol. 2004, 140, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Omenetti, S.; Pizarro, T.T. The Treg/Th17 Axis: A Dynamic Balance Regulated by the Gut Microbiome. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, A.K.; Su, Z.; Gauvin, D.M.; Paulsboe, S.E.; Duggan, R.; Lasko, L.M.; Honore, P.; Kort, M.E.; McGaraughty, S.P.; Scott, V.E.; et al. IL-23 induces regulatory T cell plasticity with implications for inflammatory skin diseases. Sci. Rep. 2019, 9, 17675. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Morito, H.; Hasegawa, A.; Daikoku, N.; Miyagawa, F.; Okazaki, A.; Fukumoto, T.; Kobayashi, N.; Kasai, T.; Watanabe, H.; et al. Identification of thymus and activation-regulated chemokine (TARC/CCL17) as a potential marker for early indication of disease and prediction of disease activity in drug-induced hypersensitivity syndrome (DIHS)/drug rash with eosinophilia and systemic symptoms (DRESS). J. Dermatol. Sci. 2013, 69, 38–43. [Google Scholar] [CrossRef]
- Sallusto, F.; Lanzavecchia, A.; Mackay, C.R. Chemokines and chemokine receptors in T-cell priming and Th1/Th2-mediated responses. Immunol. Today 1998, 19, 568–574. [Google Scholar] [CrossRef]
- Iellem, A.; Mariani, M.; Lang, R.; Recalde, H.; Panina-Bordignon, P.; Sinigaglia, F.; D’Ambrosio, D. Unique chemotactic response profile and specific expression of chemokine receptors CCR4 and CCR8 by CD4(+)CD25(+) regulatory T cells. J. Exp. Med. 2001, 194, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Komatsu-Fujii, T.; Kaneko, S.; Chinuki, Y.; Suyama, Y.; Ohta, M.; Niihara, H.; Morita, E. Serum TARC levels are strongly correlated with blood eosinophil count in patients with drug eruptions. Allergol. Int. 2017, 66, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Nakamura-Nishimura, Y.; Miyagawa, F.; Miyashita, K.; Ommori, R.; Azukizawa, H.; Asada, H. Serum thymus and activation-regulated chemokine is associated with the severity of drug reaction with eosinophilia and systemic symptoms/drug-induced hypersensitivity syndrome. Br. J. Dermatol. 2018, 178, 1430–1432. [Google Scholar] [CrossRef]
- Miyagawa, F.; Hasegawa, A.; Imoto, K.; Ogawa, K.; Kobayashi, N.; Ito, K.; Fujita, H.; Aihara, M.; Watanabe, H.; Sueki, H.; et al. Differential expression profile of Th1/Th2-associated chemokines characterizes Stevens-Johnson syndrome/toxic epidermal necrolysis (SJS/TEN) and drug-induced hypersensitivity syndrome/drug reaction with eosinophilia and systemic symptoms (DIHS/DRESS) as distinct entities. Eur. J. Dermatol. 2015, 25, 87–89. [Google Scholar] [CrossRef]
- Komatsu-Fujii, T.; Chinuki, Y.; Niihara, H.; Hayashida, K.; Ohta, M.; Okazaki, R.; Kaneko, S.; Morita, E. The thymus and activation-regulated chemokine (TARC) level in serum at an early stage of a drug eruption is a prognostic biomarker of severity of systemic inflammation. Allergol. Int. 2018, 67, 90–95. [Google Scholar] [CrossRef]
- Teraki, Y.; Fukuda, T. Skin-Homing IL-13-Producing T Cells Expand in the Circulation of Patients with Drug Rash with Eosinophilia and Systemic Symptoms. Dermatology 2017, 233, 242–249. [Google Scholar] [CrossRef]
- Tsai, Y.G.; Liou, J.H.; Hung, S.I.; Chen, C.B.; Chiu, T.M.; Wang, C.W.; Chung, W.H. Increased Type 2 Innate Lymphoid Cells in Patients with Drug Reaction with Eosinophilia and Systemic Symptoms Syndrome. J. Investig. Dermatol. 2019, 139, 1722–1731. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.Y.; Kim, J.; Ham, J.; Cho, S.H.; Kang, H.R.; Kim, H.Y. Altered T cell and monocyte subsets in prolonged immune reconstitution inflammatory syndrome related with DRESS (drug reaction with eosinophilia and systemic symptoms). Asia Pac. Allergy 2020, 10, e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ange, N.; Alley, S.; Fernando, S.L.; Coyle, L.; Yun, J. Drug Reaction with Eosinophilia and Systemic Symptoms (DRESS) syndrome successfully treated with mepolizumab. J. Allergy Clin. Immunol. Pract. 2018, 6, 1059–1060. [Google Scholar] [CrossRef] [PubMed]
- Damsky, W.E.; Vesely, M.D.; Lee, A.I.; Choi, J.; Meyer, A.C.; Chen, M.; Ahmad, T.; King, B. Drug-induced hypersensitivity syndrome with myocardial involvement treated with tofacitinib. JAAD Case Rep. 2019, 5, 1018–1026. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, Y.; Azukizawa, H.; Asada, H.; Niihara, H.; Morita, E.; Yamauchi, T.; Mizukawa, Y.; Kusakabe, Y.; Numazawa, S.; Izumi, M.; et al. Drug-induced hypersensitivity syndrome/drug reaction with eosinophilia and systemic symptoms due to lamotrigine differs from that due to other drugs. J. Dermatol. 2019, 46, 226–233. [Google Scholar] [CrossRef]
- Hammond, C.L.; Roztocil, E.; Phipps, R.P.; Feldon, S.E.; Woeller, C.F. Proton pump inhibitors attenuate myofibroblast formation associated with thyroid eye disease through the aryl hydrocarbon receptor. PLoS ONE 2019, 14, e0222779. [Google Scholar] [CrossRef]
- Rochman, M.; Xie, Y.M.; Mack, L.; Caldwell, J.M.; Klingler, A.M.; Osswald, G.A.; Azouz, N.P.; Rothenberg, M.E. Broad Transcriptional Response of the Human Esophageal Epithelium to Proton Pump Inhibitors. J. Allergy Clin. Immunol. 2020, in press. [Google Scholar] [CrossRef]


{kind=link}
| 1 | Hospitalisation |
| 2 | Skin eruption |
| 3 | Fever >38 °C |
| 4 | Lymphadenopathy at least 2 sites |
| 5 | Involvement of at least one internal organ |
| 6a | Lymphocytosis (>4 × 103/μL) or lymphopenia (<1.5 × 103/μL) |
| 6b | Eosinophilia >10% or >700/μL |
| 6c | Thrombocytopaenia (<120 × 103/μL) |
| 1 | Maculopapular rash developing >3 weeks after starting with a limited number of drugs |
| 2 | Prolonged clinical symptoms 2 weeks after discontinuation of the causative drug |
| 3 | Fever >38 °C |
| 4 | Liver abnormalities (alanine aminotransferase >100 U/L) |
| 5 | Leucocyte abnormalities (at least one present) |
| a | Leucocytosis (>11 × 109/L) |
| b | Atypical lymphocytosis (>5%) |
| c | Eosinophilia (>1.5 × 109/L) |
| 6 | Lymphadenopathy |
| 7 | HHV-6 reactivation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganeshanandan, L.; Lucas, M. Drug Reaction with Eosinophilia and Systemic Symptoms: A Complex Interplay between Drug, T Cells, and Herpesviridae. Int. J. Mol. Sci. 2021, 22, 1127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031127
Ganeshanandan L, Lucas M. Drug Reaction with Eosinophilia and Systemic Symptoms: A Complex Interplay between Drug, T Cells, and Herpesviridae. International Journal of Molecular Sciences. 2021; 22(3):1127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031127
Chicago/Turabian StyleGaneshanandan, Luckshman, and Michaela Lucas. 2021. "Drug Reaction with Eosinophilia and Systemic Symptoms: A Complex Interplay between Drug, T Cells, and Herpesviridae" International Journal of Molecular Sciences 22, no. 3: 1127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031127