
The Role of Oxidative Stress and Autophagy in Blue-Light-Induced Damage to the Retinal Pigment Epithelium in Zebrafish In Vitro and In Vivo

 ,
,  , and
, and
Abstract
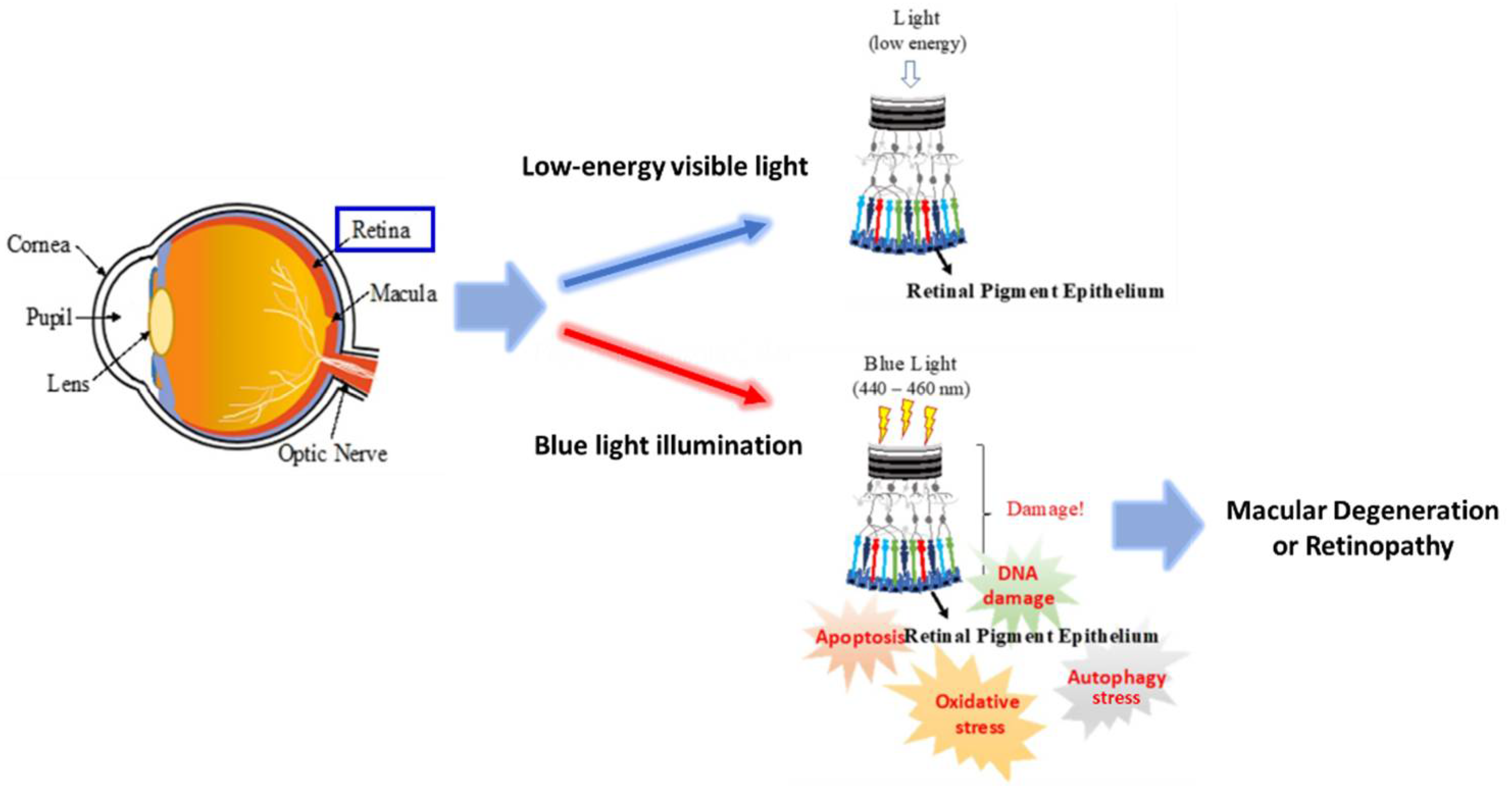
:1. Introduction
2. Results
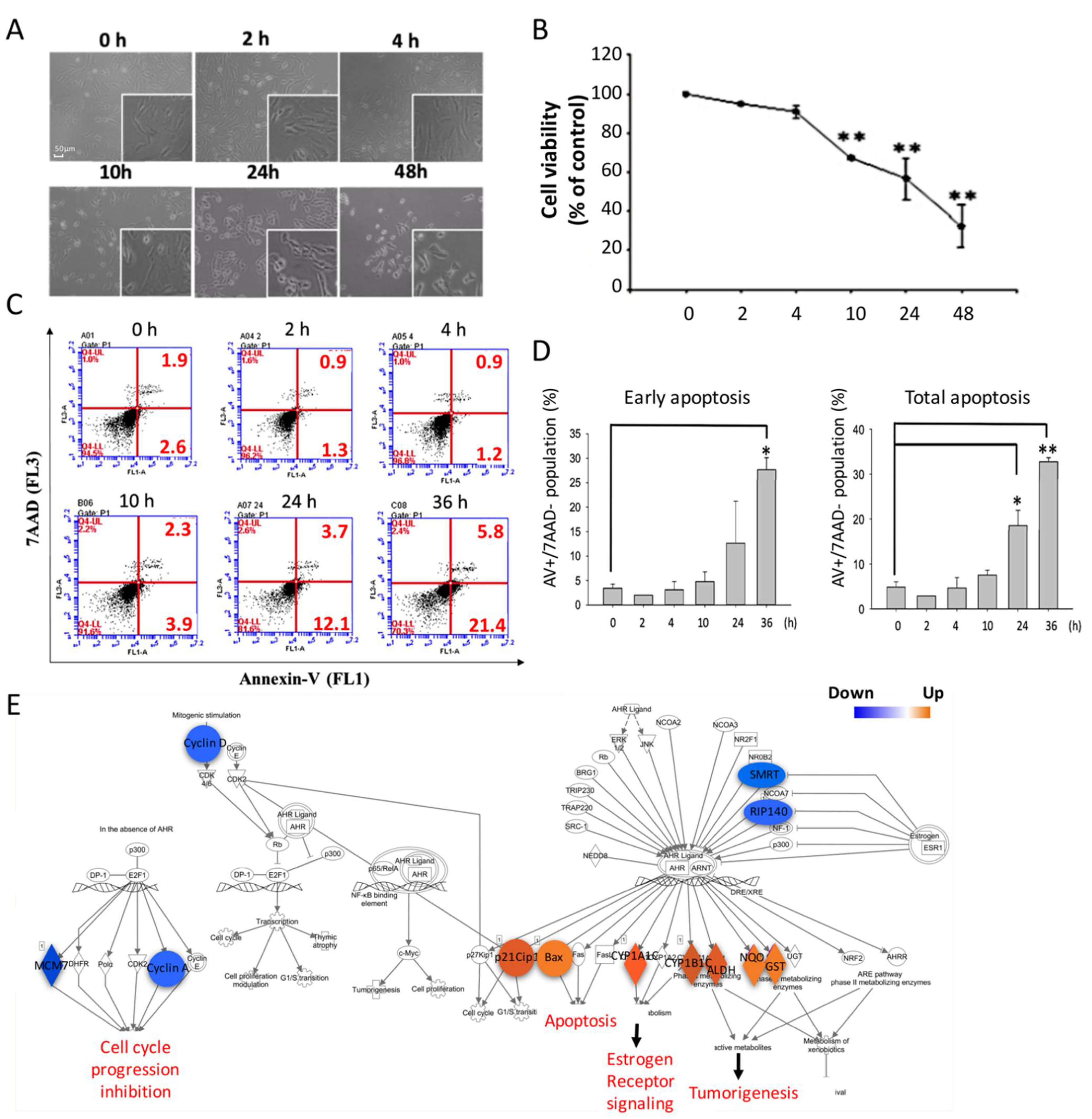
2.1. Blue Light Exposure Induced ARPE-19 Cell Death by Activating Apoptosis
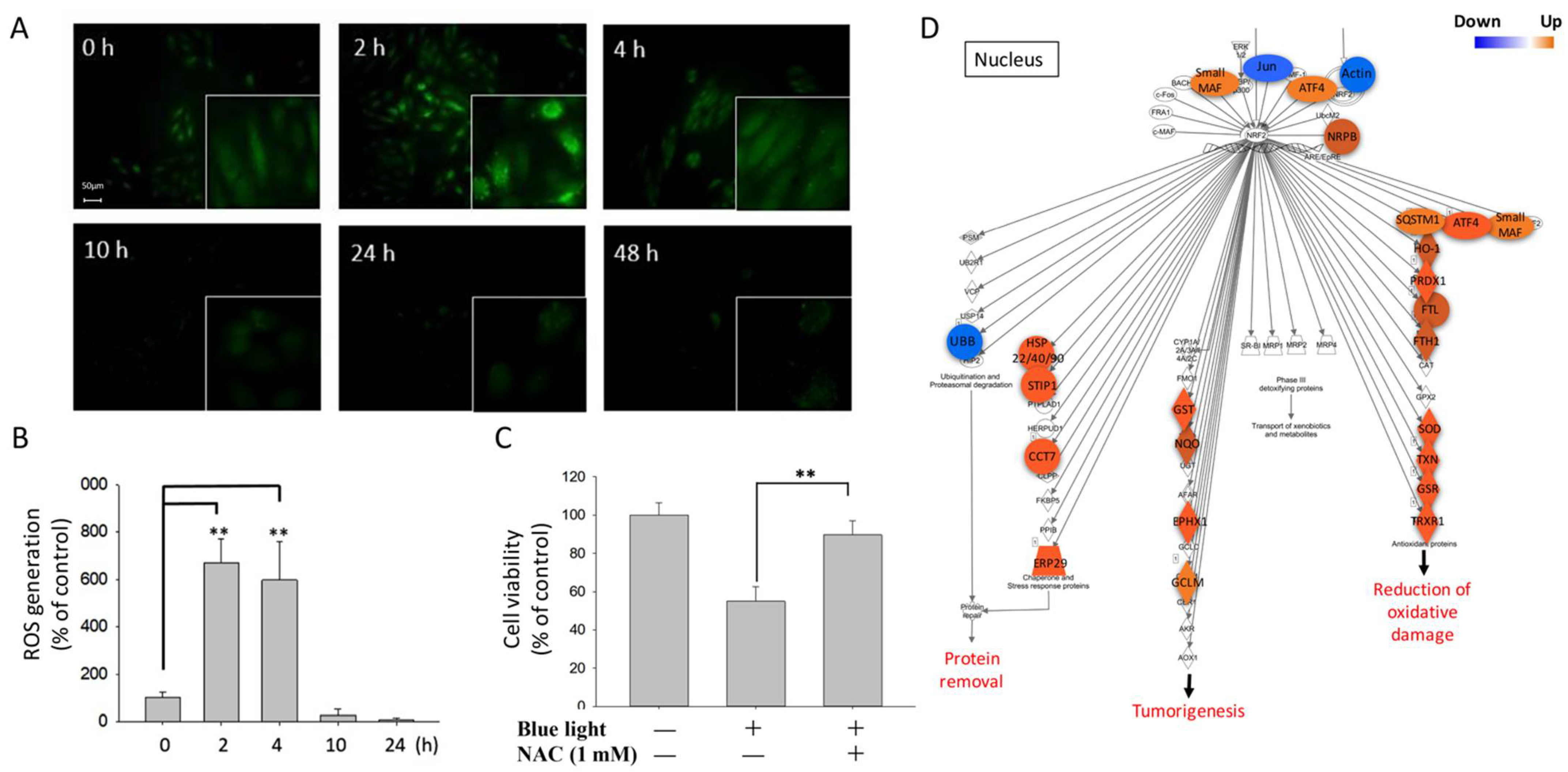
2.2. Oxidative Stress Generation Induced by Blue Light Exposure in ARPE-19 Cells
2.3. DNA Damage Triggered by Blue Light Stimulation
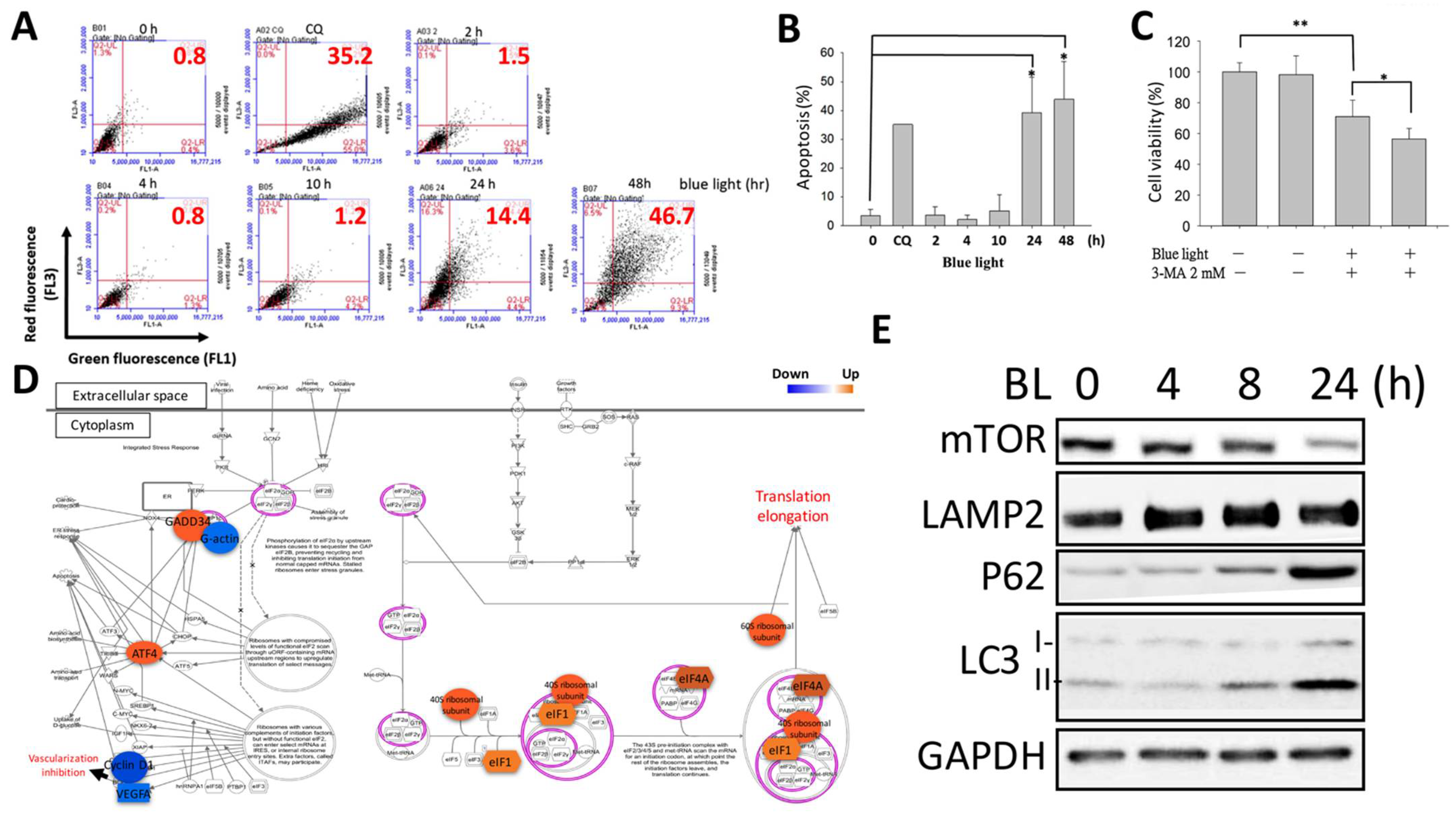
2.4. Cellular Autophagy Activation in ARPE-19 Cells Due to Blue Light Exposure
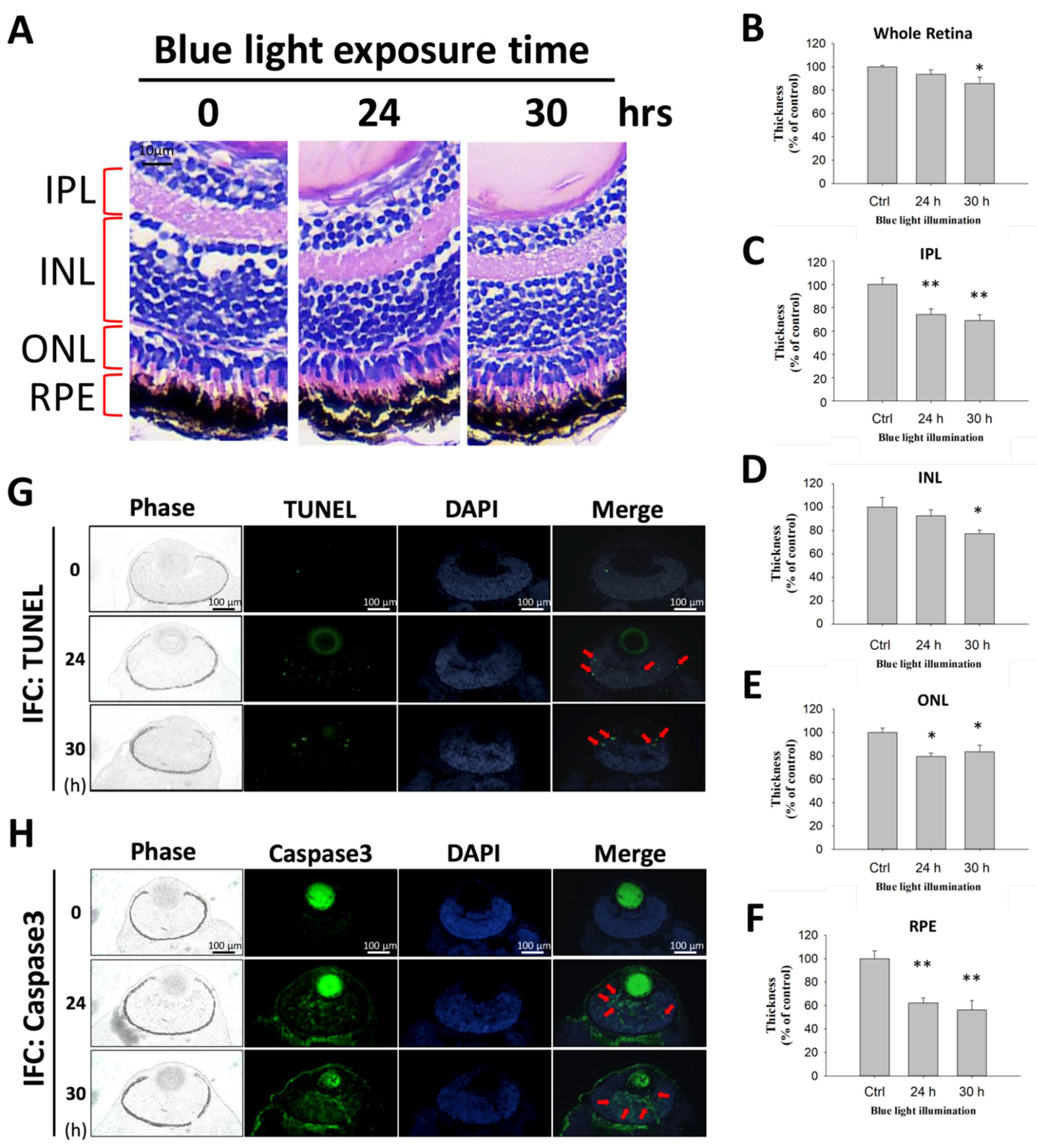
2.5. The Long-Term Effect of Blue Light Exposure on the Retina of Zebrafish Larvae
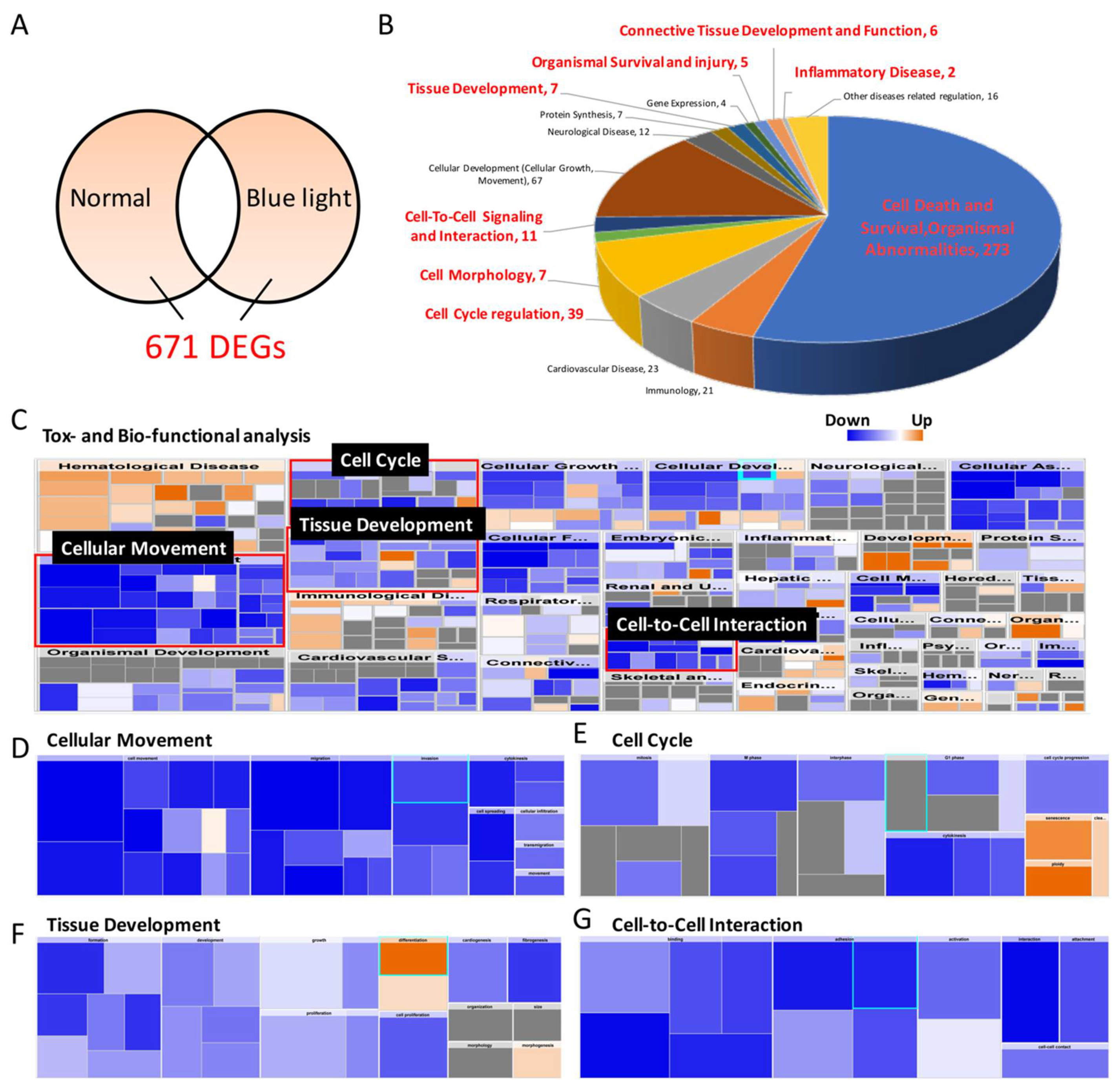
2.6. RNA Sequencing to Determine Potential Signal Transduction Involved in Blue-Light-Induced Damage
3. Discussion
3.1. General Discussion
3.2. Blue-Light-Induced Apoptosis
3.3. Blue-Light-Induced Oxidative Stress
3.4. Blue-Light-Induced DNA Damage
3.5. Blue-Light-Induced Autophagy
3.6. Animal Model
3.7. NGS Data for Potential Mechanism Prediction
4. Materials and Methods
4.1. Culture
4.2. Assessment of Cell Viability
4.3. Assessment of Cell Viability
4.4. Determination of Apoptosis by an Annexin V/7AAD Assay
4.5. Determination of Intracellular ROS
4.6. Assessment of DNA Damage
4.7. Acridine Orange Staining for the Assessment of Autophagy
4.8. RNA Extraction and Analysis
4.9. Library Preparation and Sequencing
4.10. Biological Pathway Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Powell, S.R.; Wang, P.; Divald, A.; Teichberg, S.; Haridas, V.; McCloskey, T.W.; Davies, K.J.; Katzeff, H. Aggregates of oxidized proteins (lipofuscin) induce apoptosis through proteasome inhibition and dysregulation of pro-apoptotic proteins. Free Radic. Biol. Med. 2005, 38, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Sun, T.; Jiang, Y.; Wu, L.; Cai, X.; Sun, X.; Sun, X. Photooxidative damage in retinal pigment epithelial cells via grp78 and the protective role of grape skin polyphenols. Food Chem. Toxicol. 2014, 74, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Brandstetter, C.; Mohr, L.K.; Latz, E.; Holz, F.G.; Krohne, T.U. Light induces nlrp3 inflammasome activation in retinal pigment epithelial cells via lipofuscin-mediated photooxidative damage. J. Mol. Med. 2015, 93, 905–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, S.E.; Sundelin, S.P.; Wihlmark, U.; Brunk, U.T. Aging of cultured retinal pigment epithelial cells: Oxidative reactions, lipofuscin formation and blue light damage. Doc. Ophthalmol. 2003, 106, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on rpe degeneration in non-neovascular amd. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Xu, X.R.; Yu, H.T.; Yang, Y.; Hang, L.; Yang, X.W.; Ding, S.H. Quercetin phospholipid complex significantly protects against oxidative injury in arpe-19 cells associated with activation of nrf2 pathway. Eur. J. Pharmacol. 2016, 770, 1–8. [Google Scholar] [CrossRef]
- Woo, J.M.; Shin, D.Y.; Lee, S.J.; Joe, Y.; Zheng, M.; Yim, J.H.; Callaway, Z.; Chung, H.T. Curcumin protects retinal pigment epithelial cells against oxidative stress via induction of heme oxygenase-1 expression and reduction of reactive oxygen. Mol. Vis. 2012, 18, 901–908. [Google Scholar]
- Young, R.W. Solar radiation and age-related macular degeneration. Surv. Ophthalmol. 1988, 32, 252–269. [Google Scholar] [CrossRef]
- Shen, C.J.; Cheng, Y.M.; Wang, C.L. Lncrna pvt1 epigenetically silences mir-195 and modulates emt and chemoresistance in cervical cancer cells. J. Drug Target. 2017, 25, 637–644. [Google Scholar] [CrossRef]
- Vila, N.; Siblini, A.; Esposito, E.; Bravo-Filho, V.; Zoroquiain, P.; Aldrees, S.; Logan, P.; Arias, L.; Burnier, M.N. Blue-light filtering alters angiogenic signaling in human retinal pigmented epithelial cells culture model. BMC Ophthalmol. 2017, 17, 198. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Kuse, Y.; Tsuruma, K.; Shimazawa, M.; Hara, H. The involvement of the oxidative stress in murine blue led light-induced retinal damage model. Biol. Pharm. Bull. 2017, 40, 1219–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, T.; Omoto, M.; Noda, K.; Ebinuma, M.; Kubota, S.; Koizumi, H.; Yoshida, S.; Ozawa, Y.; Shimmura, S.; Ishida, S.; et al. Retinal phototoxicity in a novel murine model of intraocular lens implantation. Mol. Vis. 2009, 15, 2751–2761. [Google Scholar] [PubMed]
- Tanito, M.; Kaidzu, S.; Anderson, R.E. Protective effects of soft acrylic yellow filter against blue light-induced retinal damage in rats. Exp. Eye Res. 2006, 83, 1493–1504. [Google Scholar] [CrossRef]
- Boulton, M.; Rozanowska, M.; Rozanowski, B.; Wess, T. The photoreactivity of ocular lipofuscin. Photochem. Photobiol. Sci. 2004, 3, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Wielgus, A.R.; Chignell, C.F.; Ceger, P.; Roberts, J.E. Comparison of a2e cytotoxicity and phototoxicity with all-trans-retinal in human retinal pigment epithelial cells. Photochem. Photobiol. 2010, 86, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Nordberg, J.; Arner, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Narimatsu, T.; Negishi, K.; Miyake, S.; Hirasawa, M.; Osada, H.; Kurihara, T.; Tsubota, K.; Ozawa, Y. Blue light-induced inflammatory marker expression in the retinal pigment epithelium-choroid of mice and the protective effect of a yellow intraocular lens material in vivo. Exp. Eye Res. 2015, 132, 48–51. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Gottlieb, E.; Brooks, D.G.; Murphy, M.P.; Dunaief, J.L. Mitochondria-derived reactive oxygen species mediate blue light-induced death of retinal pigment epithelial cells. Photochem. Photobiol. 2004, 79, 470–475. [Google Scholar] [CrossRef]
- Wielgus, A.R.; Collier, R.J.; Martin, E.; Lih, F.B.; Tomer, K.B.; Chignell, C.F.; Roberts, J.E. Blue light induced a2e oxidation in rat eyes—Experimental animal model of dry amd. Photochem. Photobiol. Sci. 2010, 9, 1505–1512. [Google Scholar] [CrossRef]
- Wu, J.; Seregard, S.; Spangberg, B.; Oskarsson, M.; Chen, E. Blue light induced apoptosis in rat retina. Eye 1999, 13 Pt 4, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Seko, Y.; Pang, J.; Tokoro, T.; Ichinose, S.; Mochizuki, M. Blue light-induced apoptosis in cultured retinal pigment epithelium cells of the rat. Graefes. Arch. Clin. Exp. Ophthalmol. 2001, 239, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P.; Kierulf, P.; Gaustad, P.; Skulberg, A.; Bruun, J.N.; Halvorsen, S.; Sorensen, E. Plasma endotoxin as a predictor of multiple organ failure and death in systemic meningococcal disease. J. Infect. Dis. 1989, 159, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Sorrell, M.; Berman, Z. Functional interplay between atm/atr-mediated DNA damage response and DNA repair pathways in oxidative stress. Cell Mol. Life Sci. 2014, 71, 3951–3967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaarniranta, K.; Tokarz, P.; Koskela, A.; Paterno, J.; Blasiak, J. Autophagy regulates death of retinal pigment epithelium cells in age-related macular degeneration. Cell Biol. Toxicol. 2017, 33, 113–128. [Google Scholar] [PubMed] [Green Version]
- Boya, P.; Esteban-Martinez, L.; Serrano-Puebla, A.; Gomez-Sintes, R.; Villarejo-Zori, B. Autophagy in the eye: Development, degeneration, and aging. Prog. Retin. Eye Res. 2016, 55, 206–245. [Google Scholar] [CrossRef]
- Chen, Y.; Sawada, O.; Kohno, H.; Le, Y.Z.; Subauste, C.; Maeda, T.; Maeda, A. Autophagy protects the retina from light-induced degeneration. J. Biol. Chem. 2013, 288, 7506–7518. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Hu, Q.; Li, L.; Tang, X.; Zou, J.; Huang, L.; Li, X. Protective effects of autophagy against blue light-induced retinal degeneration in aged mice. Sci. China Life Sci. 2019, 62, 244–256. [Google Scholar] [CrossRef]
- Chou, H.L.; Lin, Y.H.; Liu, W.; Wu, C.Y.; Li, R.N.; Huang, H.W.; Chou, C.H.; Chiou, S.J.; Chiu, C.C. Combination therapy of chloroquine and c(2)-ceramide enhances cytotoxicity in lung cancer h460 and h1299 cells. Cancers 2019, 11, 370. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Predicted Activation State | p-Value | Activation z-Score | Involved Molecules |
|---|---|---|---|---|
| Cellular Movement | Decreased | 9.36 × 10−47 | −4.498 | ACTB, ACTN1, ACTN4, ACTR2, ADAM19, ADAM9, ADAMTS5, AHNAK, AKAP12, AKT1S1, ALCAM, ALDH1A3, ALDOA, ANGPTL2, ANXA2, ANXA3, ARHGAP11A, ARL4C, ARPC2, ASPH, ASPM, AXL, BAX, BTG2, CA9, CALD1, CALR, CAP1, CAST, CCBE1, CCN1, CCN2, CCNA2, CCND1, CD274, CD44, CD63, CDH11, CDH4, CDK1, CDKN1A, CENPF, CFL1, CLIC4, CNN2, COL11A1, COL4A1,COL4A2, CRMP1, CRY1, CSF1, CTSL, CYP1A1, CYP1B1, DAG1, DCBLD2, DDRGK1, DEK, DIAPH1, DIO2, DKK1, DNAJA1, DNAJB4, DSP, DUSP1, ECT2, EDN1, EFNB2, EGFR, EGLN1, EMP1, ENAH, ENO1, ENPP2, EPPK1, ERRFI1, ETS1, EZR, F2R, FASN, FAT1, FAT3, FBN1, FERMT2, FGF1, FGF5, FKBP4, FLNA, FLNB, FLNC, FLRT2, FN1, FOSL2, FOXF2, FOXQ1, FSTL1, GADD45A, GAPDH, GAS5, GDF5, GIPC1, GNAI2, GNG12, GPC1, GPI, GTSE1, HIF1A, HMGA2, HMGB1, HMGB2, HMOX1, HSP90AA1, HSPA8, HSPB1, HSPD1, HSPG2, ID1, ID3, IGF2R, IGFBP3, IGFBP5, IGFBP7, IL7R, INHBA, INPPL1, IRS2, ITGA3, ITGA4, ITGA5, ITGAV, ITGB1, ITGB3, JUN, JUNB, KDM3A, KIF14, KIF20B, KIFC1, KLF2, KLF6, KPNA2, KRT18, KRT8, LCP1, LDHA, LIMA1, LMNB1,LMNB2, LMO4,LMO7, LOX, LOXL2, LRP1, LTBP2, LTBP3, LTBR, MAP1B, MAP2, MAP4, MBOAT7, MDM2, MIR100HG, MSN, MYADM, MYH10,MYH9, MYL12A, NAV1, NDST1, NEDD9, NEU1, NGF, NINJ1, NKX3-1, NOG, NPPB, NQO1, NR3C2, NREP, NRP1, OSGIN1, P4HB, PALLD, PCOLCE2, PDGFB, PDK1, PHLPP1, PLXNB2, PODXL, PPIF, PPP1R15A, PRDX1, PTEN, PTPRB, PTPRF, PTPRJ, RACGAP1, RELN, RPL13A, RPS19, RUNX2, SEMA3B, SEMA7A, SERPINB2, SERPINE1, SERPINE2, SGK1, SLC16A3, SLC3A2, SLC48A1, SLC7A11, SLC7A5, SLIT3, SOD1, SORT1, SOX9, SPARC, SPHK1, SQSTM1, TAGLN2, TFPI, TGFB2, TGFBR1, THBS1, TIMP3, TKT, TLN1, TMEM30A, TMPO, TMSB10/TMSB4X, TNFRSF10B, TNIK, TNS1, TPI1, TPM1, TPM2, TPM3, TPT1, TSPAN14, TUBA1C, TUBB, TXN, TXNRD1, UCHL1, VASP, VCL, VEGFA, VIM, VSIR, WBP2, WWC1, YBX1, ZYX |
| Cell Cycle | Decreased | 1.49 × 10−23 | −2.301 | AKAP12, ANLN, BIRC5, BUB1B, CAP1, CCNB1, CDC20, CDK1, CDKN1A, CENPE, CEP55, CFL1, CIT, CKAP2, CSF1, DIAPH3, ECT2, FLNA, GADD45A, GIPC1, GNAI2, GPC1, HIPK2, HSPB1, INCENP, ITGB1, KIF14, KIF20A, KIF20B, KIF23, KIFC1, MCM7, MDM2, MYH10, NDC80, NEK6, NEK7, NUSAP1, PLK1, PRC1, RACGAP1, SPART, SPDL1, TACC3, TM4SF1, TOP2A, UBE2S, YBX1 |
| Tissue Development | Decreased | 7.58 × 10−16 | −1.174 | ANXA2, AXL, BMPER, CA9, CALR, CCN1, CCND1, CD44, CDKN1A, CLIC4, COL4A2, CSF1, DAG1, DKK1, ECT2, EDN1, EFNB2, EGFR, F2R, FAT1, FGF1, FLNB, FN1, FOSL2, FRAS1, GPC1, HEG1, HIF1A, HIPK2, HMGB1, HMGB2, HMOX1, HSPG2, ID1, ID3, IGFBP3, ITGA4, ITGB1, ITGB3, JUN, KRT18, KRT8, LOXL2, LTBR, MYDGF, NGF, NKX3-1, NRP1, ODC1, PALLD, PDGFB, PLOD3, PTEN, PTPRJ, RUNX2, SERPINE1, SOX9, SPARC, TGFBR1, THBS1, TIMP3, VCL, VEGFA |
| Cell-Cell Signaling and Interaction | Decreased | 7.45 × 10-15 | −1.439 | ADAM9, AHNAK, ANXA2, AXL, BAMBI, BAX, BHLHE40, BMPER, BNIP3, CALR, CCN1, CCN2, CCND1, CD274, CD44, CDKN1A, CFL1, CRY1, CSF1, CTSL, DKK1, DUSP1, EDN1, EFNB2, EGFR, ETS1, F2R, FBN1, FERMT2, FGF1, FLNA, FN1, GADD45A, GAS5, GNAI2, HMGB1, HSPB8, HSPD1, HSPG2, HSPH1, ID1, ID3, ITGA3, ITGA4, ITGA5, ITGAV, ITGB1, ITGB3, KLF2, KRT18, KRT8, LOX, LRP1, LTBR, MGST1, MT-CO1, MT-ND2, MT-ND3, MT-ND4, MT-ND5, NBR1, NCOR2, NGF, NPPB, ODC1, PDGFB, PFKFB3, PHLPP1, PLXNB2, PRDX1, PRKDC, PTEN, PTPRJ, RAB3B, SERPINE2, SOD1, TAB3, TACC3, TGFB2, TGFBR1, THBS1, TLN1, TNFRSF10B, TOP2A, TPT1, VASP, VEGFA, VIM, VSIR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, K.-C.; Hsu, Y.-T.; Liu, W.; Huang, H.-L.; Chen, L.-Y.; He, C.-X.; Sheu, S.-J.; Chen, K.-J.; Lee, P.-Y.; Lin, Y.-H.; et al. The Role of Oxidative Stress and Autophagy in Blue-Light-Induced Damage to the Retinal Pigment Epithelium in Zebrafish In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 1338. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031338
Cheng K-C, Hsu Y-T, Liu W, Huang H-L, Chen L-Y, He C-X, Sheu S-J, Chen K-J, Lee P-Y, Lin Y-H, et al. The Role of Oxidative Stress and Autophagy in Blue-Light-Induced Damage to the Retinal Pigment Epithelium in Zebrafish In Vitro and In Vivo. International Journal of Molecular Sciences. 2021; 22(3):1338. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031338
Chicago/Turabian StyleCheng, Kai-Chun, Yun-Tzu Hsu, Wangta Liu, Huey-Lan Huang, Liang-Yu Chen, Chen-Xi He, Shwu-Jiuan Sheu, Kuo-Jen Chen, Po-Yen Lee, Yi-Hsiung Lin, and et al. 2021. "The Role of Oxidative Stress and Autophagy in Blue-Light-Induced Damage to the Retinal Pigment Epithelium in Zebrafish In Vitro and In Vivo" International Journal of Molecular Sciences 22, no. 3: 1338. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031338