
Delayed Administration of Angiotensin Receptor (AT2R) Agonist C21 Improves Survival and Preserves Sensorimotor Outcomes in Female Diabetic Rats Post-Stroke through Modulation of Microglial Activation

 ,
,
Abstract
:1. Introduction
2. Results
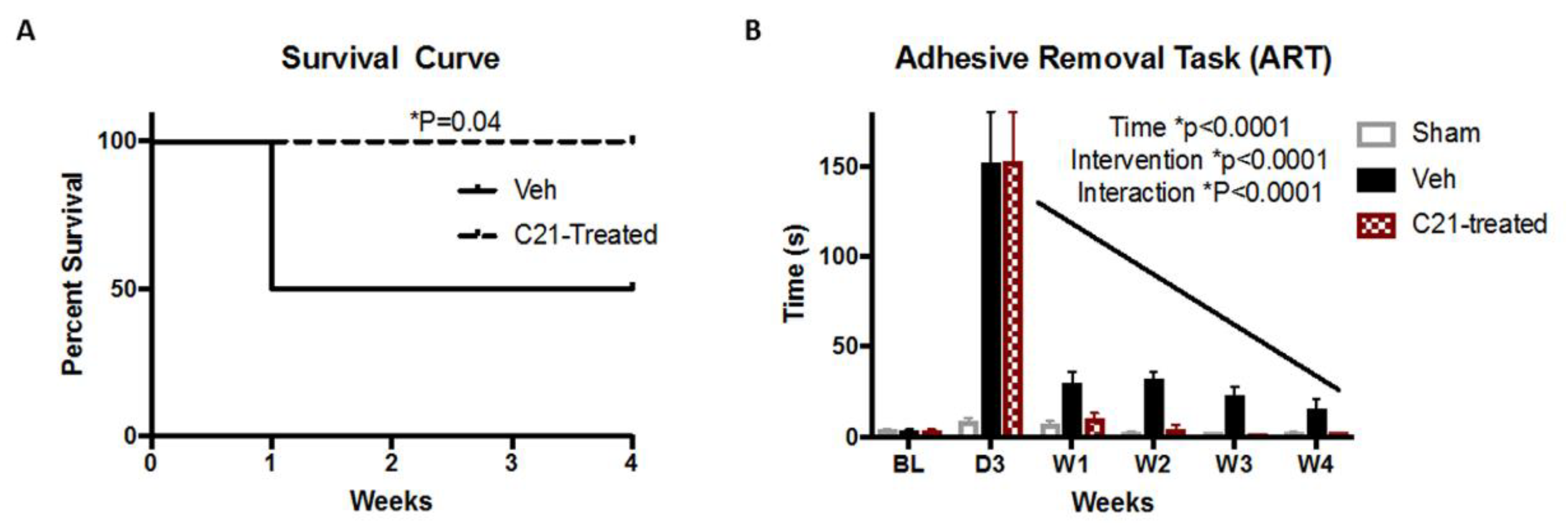
2.1. Delayed Administration of C21 Improves Survival and Sensorimotor Outcomes after Stroke
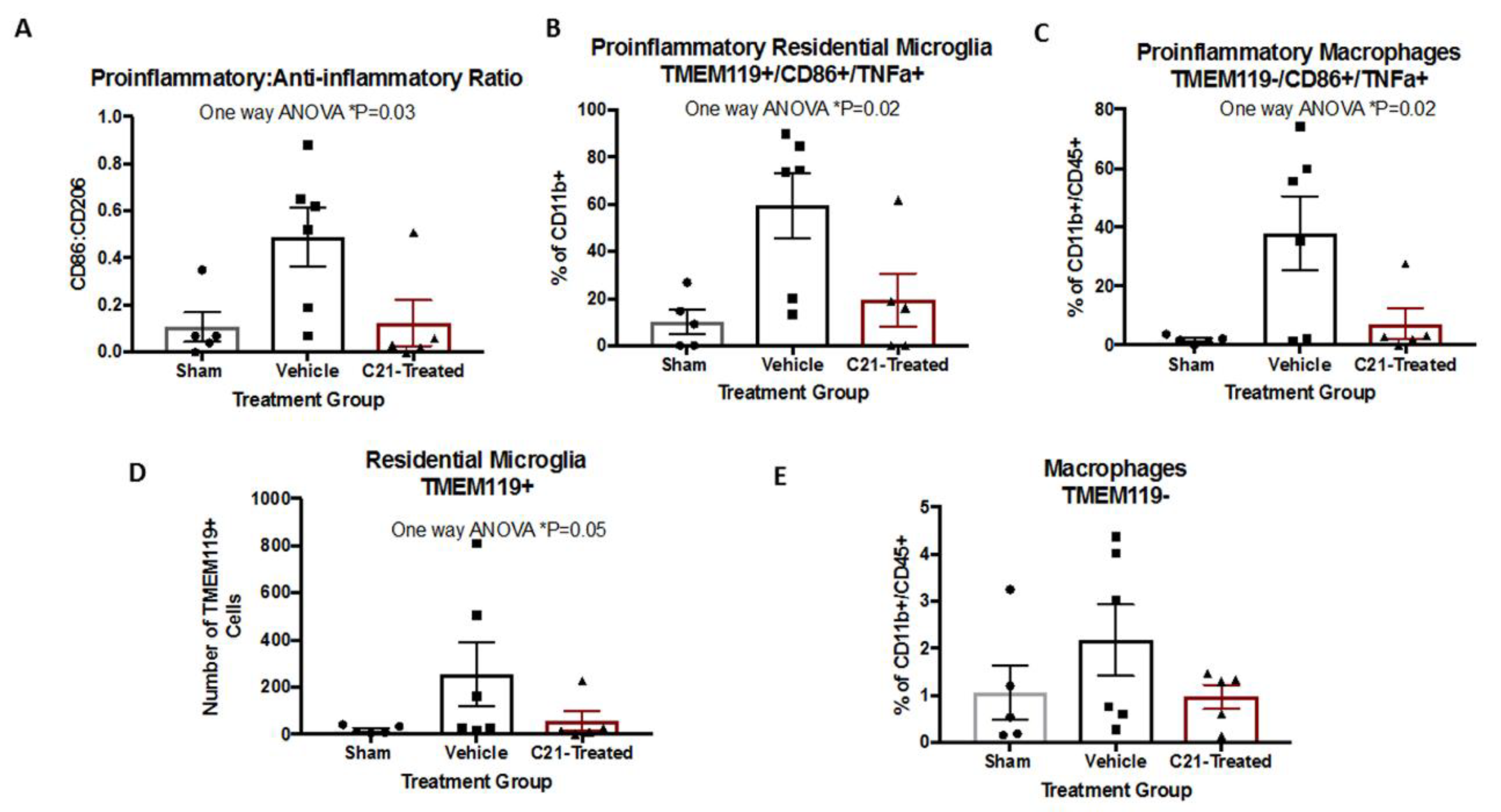
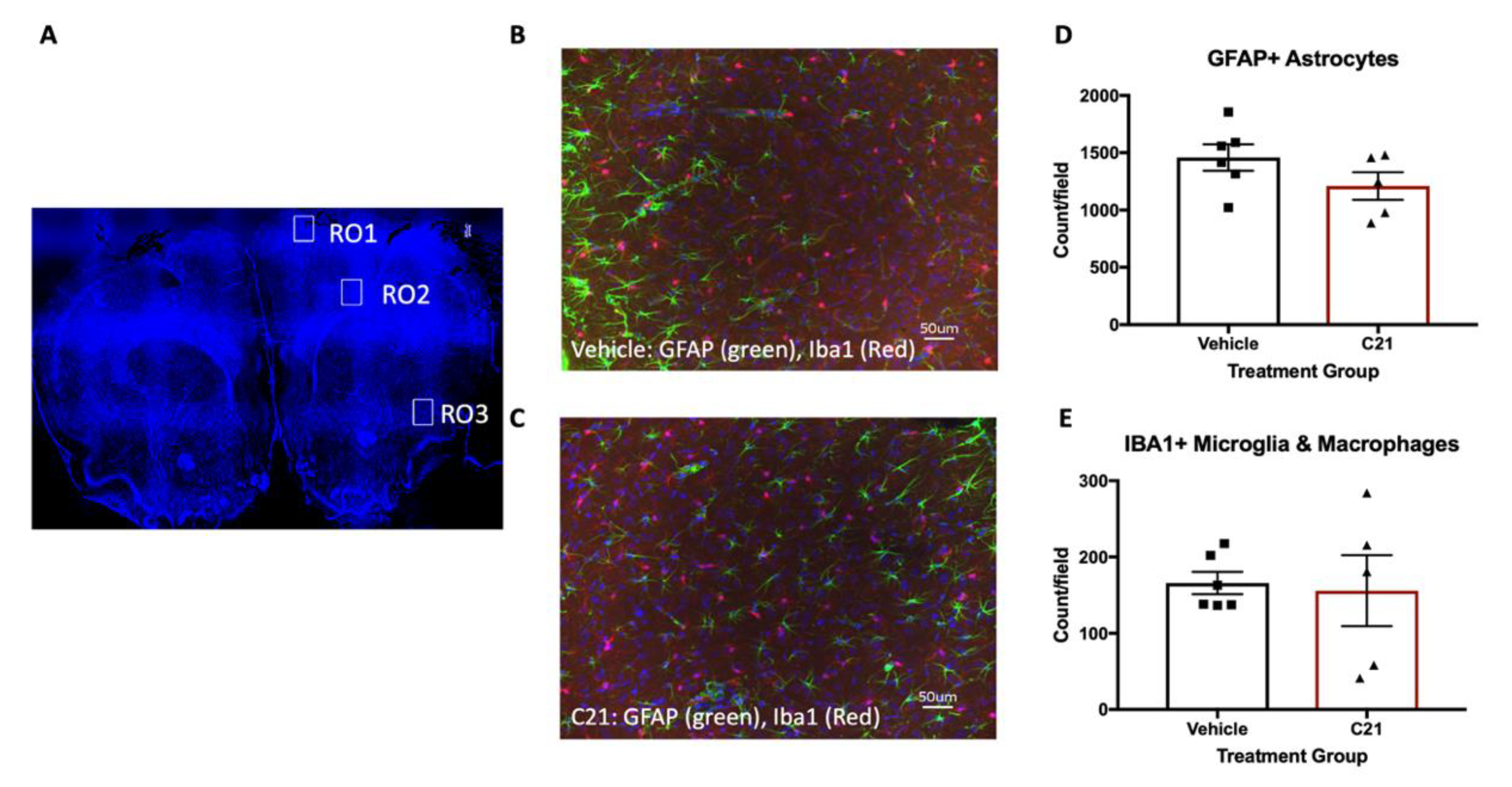
2.2. Delayed Administration of C21 Decreases Proinflammatory Microglia/Macrophages after Stroke
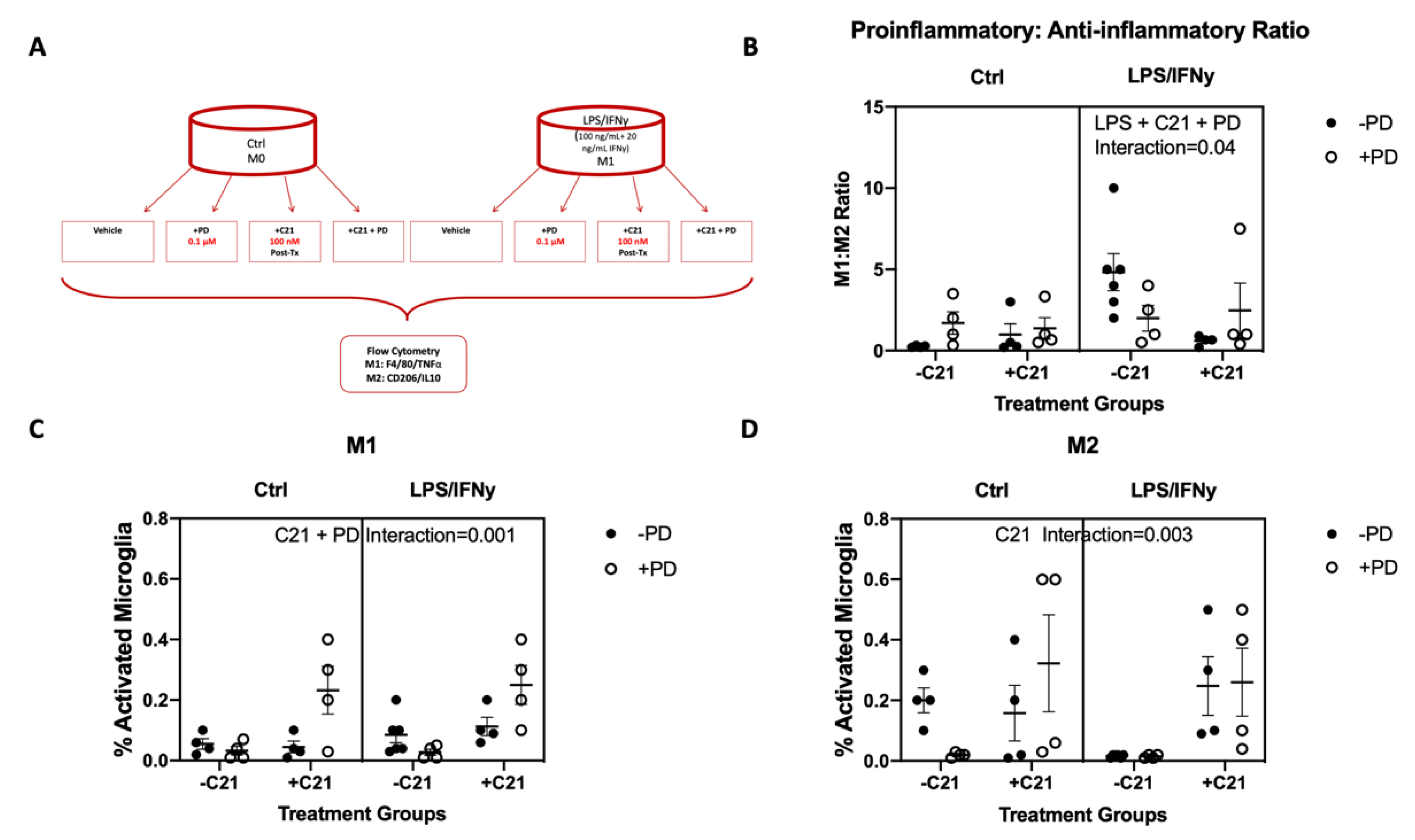
2.3. C21 Treatment Induces an M2 Phenotype and Lowers the M1:M2 Ratio In Vitro in the Microglial Cell Line
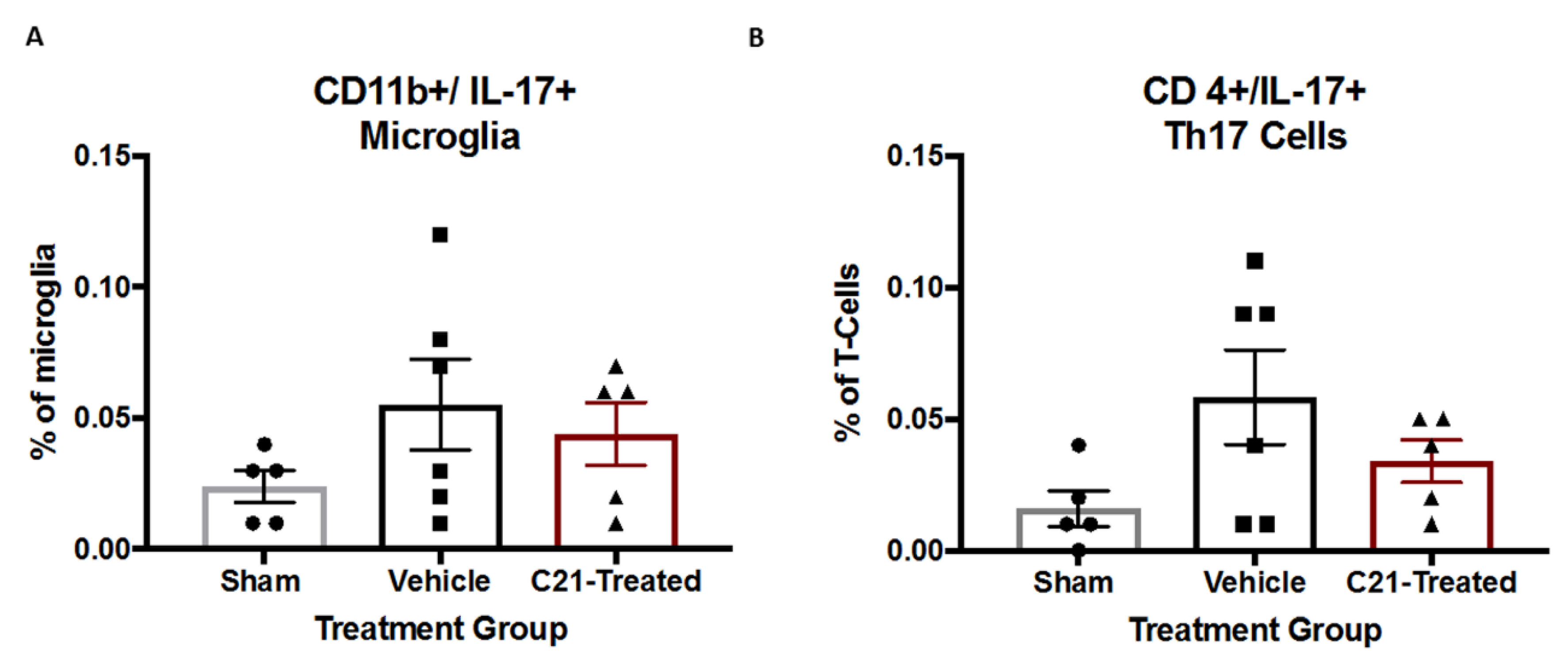
2.4. Stroke Did Not Significantly Alter IL-17 Levels from Microglia
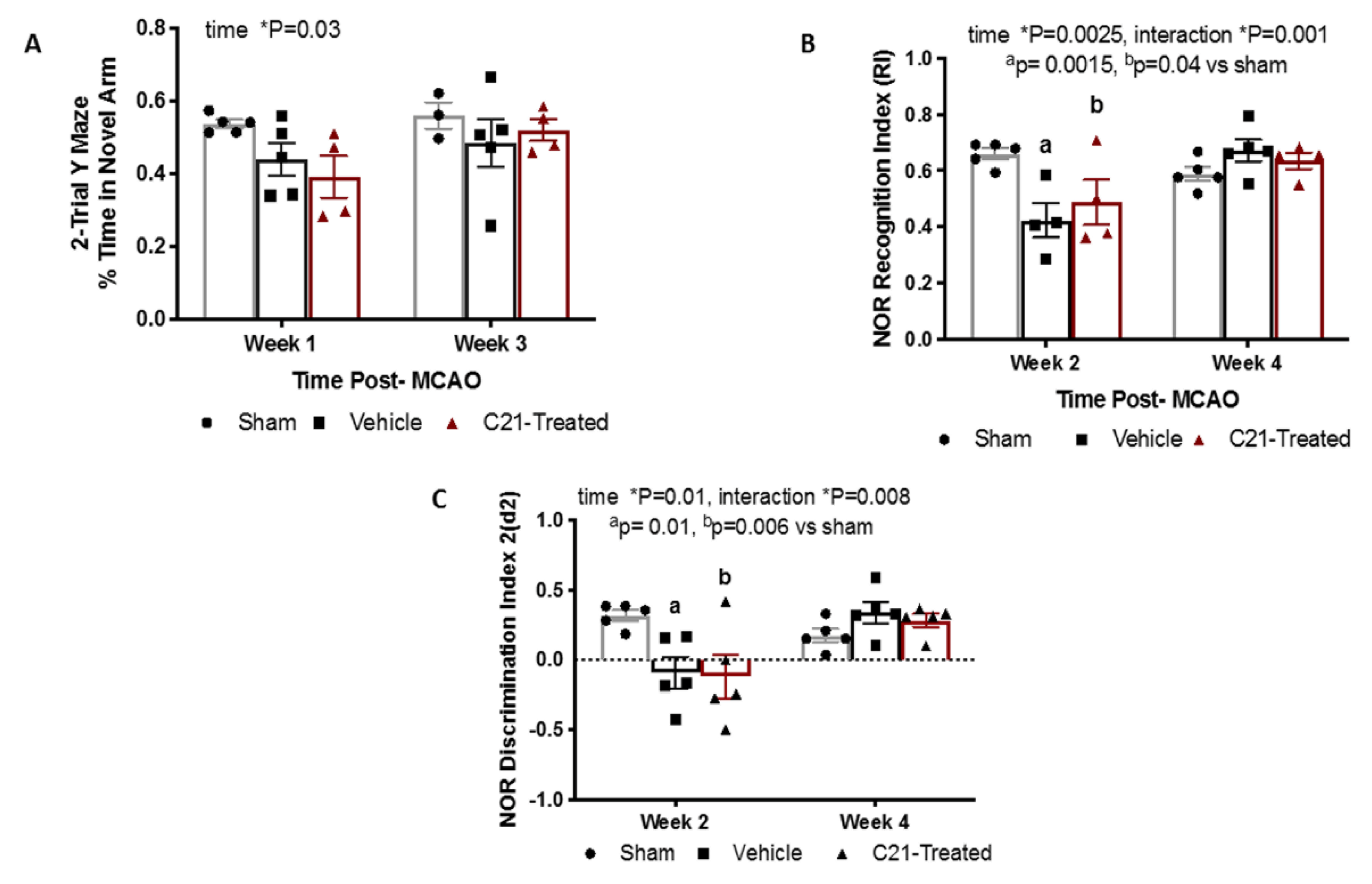
2.5. At a Dose of 0.03 mg/Kg/Day Delayed Administration of C21 Does Not Significantly Alter Cognition in Female Rats
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. Middle Cerebral Artery Occlusion (MCAO) Surgery
4.3. Treatment and Behavioral Assessments
Dose and Timing Justification
4.4. Blinding and Randomization
4.5. Cycle Synchronization
4.6. Assessment of Sensorimotor Function
4.7. Assessment of Cognitive Function
4.8. Euthanasia, Specimen Collection and Molecular Techniques
4.9. Flow Cytometry
4.10. Myelin Debris Removal and Microglial Isolation
4.11. Cellular Staining
4.12. Imaging and Analysis
4.13. Immunohistochemistry (IHC)
4.14. Cell Culture
4.15. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AT2R | Angiotensin II type 2 receptor |
| BDNF | Brain-derived neurotrophic factor |
| TH17 | T helper 17 |
| MCAO | Middle cerebral artery occlusion |
| ART | Adhesive removal test |
References
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart disease and stroke statistics-2019 update: A report from the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef]
- Khoury, J.; Kleindorfer, D.; Alwell, K.; Moomaw, C.J.; Woo, D.; Adeoye, O.; Flaherty, M.L.; Khatri, P.; Ferioli, S.; Broderick, J.P.; et al. Diabetes mellitus: A risk factor for ischemic stroke in a large biracial population. Stroke 2013, 44, 1500–1504. [Google Scholar] [CrossRef] [Green Version]
- Peters, S.A.; Huxley, R.R.; Woodward, M. Diabetes as a risk factor for stroke in women compared with men: A systematic review and meta-analysis of 64 cohorts, including 775,385 individuals and 12,539 strokes. Lancet 2014, 383, 1973–1980. [Google Scholar] [CrossRef]
- Jackson, L.; Dumanli, S.; Johnson, M.H.; Fagan, S.C.; Ergul, A. Microglia knockdown reduces inflammation and preserves cognition in diabetic animals after experimental stroke. J. Neuroinflamm. 2020, 17, 1–14. [Google Scholar] [CrossRef]
- Jackson, L.; Dong, G.; Althomali, W.; Sayed, M.A.; Eldahshan, W.; Baban, B.; Johnson, M.H.; Filosa, J.; Fagan, S.C.; Ergul, A. Delayed administration of angiotensin II type 2 receptor (AT2R) agonist compound 21 prevents the development of post-stroke cognitive impairment in diabetes through the modulation of microglia polarization. Transl. Stroke Res. 2019, 11, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.; Li, W.; Abdul, Y.; Dong, G.; Baban, B.; Ergul, A. Diabetic stroke promotes a sexually dimorphic expansion of t cells. NeuroMolecular Med. 2019, 21, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.; Eldahshan, W.; Fagan, S.C.; Ergul, A. Within the brain: The renin angiotensin system. Int. J. Mol. Sci. 2018, 19, 876. [Google Scholar] [CrossRef] [Green Version]
- Eldahshan, W.; Ishrat, T.; Pillai, B.; Sayed, M.A.; Alwhaibi, A.; Fouda, A.Y.; Ergul, A.; Fagan, S.C. Angiotensin Ii type 2 receptor stimulation with compound 21 improves neurological function after stroke in female rats: A pilot study. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H1192–H1201. [Google Scholar] [PubMed]
- Ahmed, H.A.; Ishrat, T.; Pillai, B.; Fouda, A.Y.; Sayed, M.A.; Eldahshan, W.; Waller, J.L.; Ergul, A.; Fagan, S.C. Ras modulation prevents progressive cognitive impairment after experimental stroke: A randomized, blinded preclinical trial. J. Neuroinflamm. 2018, 15, 229. [Google Scholar] [CrossRef] [PubMed]
- Alhusban, A.; Fouda, A.Y.; Bindu, P.; Ishrat, T.; Soliman, S.; Fagan, S.C. Compound 21 is pro-angiogenic in the brain and results in sustained recovery after ischemic stroke. J. Hypertens. 2015, 33, 170–180. [Google Scholar] [PubMed]
- White, M.C.; Fleeman, R.; Arnold, A.C. Sex differences in the metabolic effects of the renin-angiotensin system. Biol. Sex Differ. 2019, 10, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.; Baessler, A.; Schunkert, H. Renin angiotensin system and gender differences in the cardiovascular system. Cardiovasc. Res. 2002, 53, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Schwengel, K.; Namsolleck, P.; Lucht, K.; Clausen, B.H.; Lambertsen, K.L.; Valero-Esquitino, V.; Thöne-Reineke, C.; Müller, S.; Widdop, R.E.; Denton, K.M.; et al. Angiotensin AT2-receptor stimulation improves survival and neurological outcome after experimental stroke in mice. J. Mol. Med. 2016, 94, 957–966. [Google Scholar] [CrossRef]
- Miao, H.; Li, R.; Han, C.; Lu, X.; Zhang, H. Minocycline promotes posthemorrhagic neurogenesis via M2 microglia polarization via upregulation of the Trkb/Bdnf pathway in rats. J. Neurophysiol. 2018, 120, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Underwood, E.L.; Thompson, L.T. A high-fat diet causes impairment in hippocampal memory and sex-dependent alterations in peripheral metabolism. Neural Plast. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ward, R.; Valenzuela, J.P.; Dong, G.; Fagan, S.C.; Ergul, A. Diabetes worsens functional outcomes in young female rats: Comparison of stroke models, tissue plasminogen activator effects, and sexes. Transl. Stroke Res. 2017, 8, 429–439. [Google Scholar] [CrossRef]
- Ward, R.; Valenzuela, J.P.; Li, W.; Dong, G.; Fagan, S.C.; Ergul, A. Poststroke cognitive impairment and hippocampal neurovascular remodeling: The impact of diabetes and sex. Am. J. Physiol. Circ. Physiol. 2018, 315, H1402–H1413. [Google Scholar] [CrossRef]
- Rewell, S.; Churilov, L.; Sidon, T.K.; Aleksoska, E.; Cox, S.F.; MacLeod, M.; Howells, D.W. Evolution of ischemic damage and behavioural deficit over 6 months after MCAo in the rat: Selecting the optimal outcomes and statistical power for multi-centre preclinical trials. PLoS ONE 2017, 12, e0171688. [Google Scholar] [CrossRef]
- Lansberg, M.; O’Brien, M.W.; Tong, D.C.; Moseley, M.E.; Albers, G.W. Evolution of cerebral infarct volume assessed by diffusion-weighted magnetic resonance imaging. Arch. Neurol. 2001, 58, 613–617. [Google Scholar] [CrossRef] [Green Version]
- Cordeira, J.; Kolluru, S.S.; Rosenblatt, H.; Kry, J.; Strecker, R.E.; McCarley, R.W. Learning and memory are impaired in the object recognition task during metestrus/diestrus and after sleep deprivation. Behav. Brain Res. 2018, 339, 124–129. [Google Scholar] [CrossRef]
- Whitten, W.K. Modification of the oestrous cycle of the mouse by external stimuli associated with the male. J. Endocrinol. 1956, 13, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Whitten, W.K. Modification of the oestrous cycle of the mouse by external stimuli associated with the male; changes in the oestrous cycle determined by vaginal smears. J. Endocrinol. 1958, 17, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Mora, S.; Dussaubat, N.; Díaz-Véliz, G. Effects of the estrous cycle and ovarian hormones on behavioral indices of anxiety in female rats. Psychoneuroendocrinology 1996, 21, 609–620. [Google Scholar] [CrossRef]
- Marcondes, F.K.; Miguel, K.J.; Melo, L.L.; Spadari-Bratfisch, R.C. Estrous cycle influences the response of female rats in the elevated plus-maze test. Physiol. Behav. 2001, 74, 435–440. [Google Scholar]
- Meziane, H.; Ouagazzal, A.M.; Aubert, L.; Wietrzych, M.; Krezel, W. Estrous cycle effects on behavior of C57bl/6j and Balb/Cbyj female mice: Implications for phenotyping strategies. Genes Brain Behav. 2007, 6, 192–200. [Google Scholar] [CrossRef]
- Valero-Esquitino, V.; Lucht, K.; Namsolleck, P.; Monnet-Tschudi, F.; Stubbe, T.; Lucht, F.; Liu, M.; Ebner, F.; Brandt, C.; Danyel, L.A.; et al. Direct angiotensin type 2 receptor (At2r) stimulation attenuates t-cell and microglia activation and prevents demyelination in experimental autoimmune encephalomyelitis in mice. Clin Sci 2015, 128, 95–109. [Google Scholar] [CrossRef]
- Jing, F.; Mogi, M.; Sakata, A.; Iwanami, J.; Tsukuda, K.; Ohshima, K.; Min, L.J.; Steckelings, U.M.; Unger, T.; Dahlof, B.; et al. Direct stimulation of angiotensin ii type 2 receptor enhances spatial memory. J. Cereb. Blood Flow Metab. 2012, 32, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Sakata, A.; Mogi, M.; Iwanami, J.; Tsukuda, K.; Min, L.J.; Fujita, T.; Iwai, M.; Ito, M.; Horiuchi, M. Sex-different effect of angiotensin ii type 2 receptor on ischemic brain injury and cognitive function. Brain Res. 2009, 1300, 14–23. [Google Scholar] [CrossRef]
- Stratoulias, V.; Venero, J.L.; Tremblay, M.E.; Joseph, B. Microglial subtypes: Diversity within the microglial community. EMBO J. 2019, 38, e101997. [Google Scholar] [CrossRef]
- Tan, Y.L.; Yuan, Y.; Tian, L. Microglial regional heterogeneity and its role in the brain. Mol. Psychiatry 2020, 25, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Dubbelaar, M.L.; Kracht, L.; Eggen, B.J.L.; Boddeke, E. The kaleidoscope of microglial phenotypes. Front. Immunol. 2018, 9, 1753. [Google Scholar] [CrossRef] [PubMed]
- Hurn, P.D.; Subramanian, S.; Parker, S.M.; Afentoulis, M.E.; Kaler, L.J.; Vandenbark, A.A.; Offner, H. T- and B-cell-deficient mice with experimental stroke have reduced lesion size and inflammation. J. Cereb. Blood Flow Metab. 2007, 27, 1798–1805. [Google Scholar] [PubMed]
- Becker, K.J. Modulation of the postischemic immune response to improve stroke outcome. Stroke 2010, 41, S75–S78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taleb, S.; Tedgui, A.; Mallat, Z. Il-17 and Th17 cells in atherosclerosis: Subtle and contextual roles. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Gelderblom, M.; Weymar, A.; Bernreuther, C.; Velden, J.; Arunachalam, P.; Steinbach, K.; Orthey, E.; Arumugam, T.V.; Leypoldt, F.; Simova, O.; et al. Neutralization of the Il-17 axis diminishes neutrophil invasion and protects from ischemic stroke. Blood 2012, 120, 3793–3802. [Google Scholar] [CrossRef] [Green Version]
- Alhusban, A.; Kozak, A.; Ergul, A.; Fagan, S.C. At1 receptor antagonism is proangiogenic in the brain: Bdnf a novel mediator. J. Pharmacol. Exp. Ther. 2013, 344, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.; Somanath, P.R.; Kozak, A.; Goc, A.; El-Remessy, A.B.; Ergul, A.; Johnson, M.H.; Alhusban, A.; Soliman, S.; Fagan, S.C. Vascular protection by angiotensin receptor antagonism involves differential vegf expression in both hemispheres after experimental stroke. PLoS ONE 2011, 6, e24551. [Google Scholar] [CrossRef] [Green Version]
- Fouda, A.Y.; Alhusban, A.; Ishrat, T.; Pillai, B.; Eldahshan, W.; Waller, J.L.; Ergul, A.; Fagan, S.C. Brain-derived neurotrophic factor knockdown blocks the angiogenic and protective effects of angiotensin modulation after experimental stroke. Mol. Neurobiol. 2017, 54, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Ishitsuka, K.; Ago, T.; Arimura, K.; Nakamura, K.; Tokami, H.; Makihara, N.; Kuroda, J.; Kamouchi, M.; Kitazono, T. Neurotrophin production in brain pericytes during hypoxia: A role of pericytes for neuroprotection. Microvasc. Res. 2012, 83, 352–359. [Google Scholar] [CrossRef]
- Nakajima, K.; Honda, S.; Tohyama, Y.; Imai, Y.; Kohsaka, S.; Kurihara, T. Neurotrophin secretion from cultured microglia. J. Neurosci. Res. 2001, 65, 322–331. [Google Scholar] [CrossRef]
- Nakajima, K.; Tohyama, Y.; Kohsaka, S.; Kurihara, T. Ceramide activates microglia to enhance the production/secretion of brain-derived neurotrophic factor (bdnf) without induction of deleterious factors in vitro. J. Neurochem. 2002, 80, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Holt, L.M.; Olsen, M.L. Novel applications of magnetic cell sorting to analyze cell-type specific gene and protein expression in the central nervous system. PLoS ONE 2016, 11, e0150290. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Types | CD11b | CD45 | TMEM119 | CD86 | TNFα | CD206 | IL-17 | CD4 |
|---|---|---|---|---|---|---|---|---|
| M1(CD86+/TNFα+) | + | +low | N/A | + | N/A | N/A | ||
| M2(CD206+/IL10+) | + | +low | N/A | - | N/A | + | ||
| Residential Microglia (TMEM119+) | + | + | + | N/A | N/A | N/A | ||
| Infiltrating Macrophages | + | + | - | N/A | N/A | N/A | ||
| M1 macrophages | + | + | - | + | + | N/A | ||
| Inactivated microglia | + | +low | N/A | - | N/A | - | ||
| IL17+ Microglia | + | +low | N/A | N/A | N/A | N/A | + | N/A |
| Th17 | N/A | N/A | N/A | N/A | N/A | N/A | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson-Cowan, L.; Eldahshan, W.; Dumanli, S.; Dong, G.; Jamil, S.; Abdul, Y.; Althomali, W.; Baban, B.; Fagan, S.C.; Ergul, A. Delayed Administration of Angiotensin Receptor (AT2R) Agonist C21 Improves Survival and Preserves Sensorimotor Outcomes in Female Diabetic Rats Post-Stroke through Modulation of Microglial Activation. Int. J. Mol. Sci. 2021, 22, 1356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031356
Jackson-Cowan L, Eldahshan W, Dumanli S, Dong G, Jamil S, Abdul Y, Althomali W, Baban B, Fagan SC, Ergul A. Delayed Administration of Angiotensin Receptor (AT2R) Agonist C21 Improves Survival and Preserves Sensorimotor Outcomes in Female Diabetic Rats Post-Stroke through Modulation of Microglial Activation. International Journal of Molecular Sciences. 2021; 22(3):1356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031356
Chicago/Turabian StyleJackson-Cowan, LaDonya, Wael Eldahshan, Selin Dumanli, Guangkuo Dong, Sarah Jamil, Yasir Abdul, Waleed Althomali, Babak Baban, Susan C. Fagan, and Adviye Ergul. 2021. "Delayed Administration of Angiotensin Receptor (AT2R) Agonist C21 Improves Survival and Preserves Sensorimotor Outcomes in Female Diabetic Rats Post-Stroke through Modulation of Microglial Activation" International Journal of Molecular Sciences 22, no. 3: 1356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031356