
Effects of Pyrroloquinoline Quinone on Lipid Metabolism and Anti-Oxidative Capacity in a High-Fat-Diet Metabolic Dysfunction-Associated Fatty Liver Disease Chick Model


Abstract
:1. Introduction
2. Results
2.1. Effects of PQQ.Na2 on Performance and Egg Quality of Laying Hens
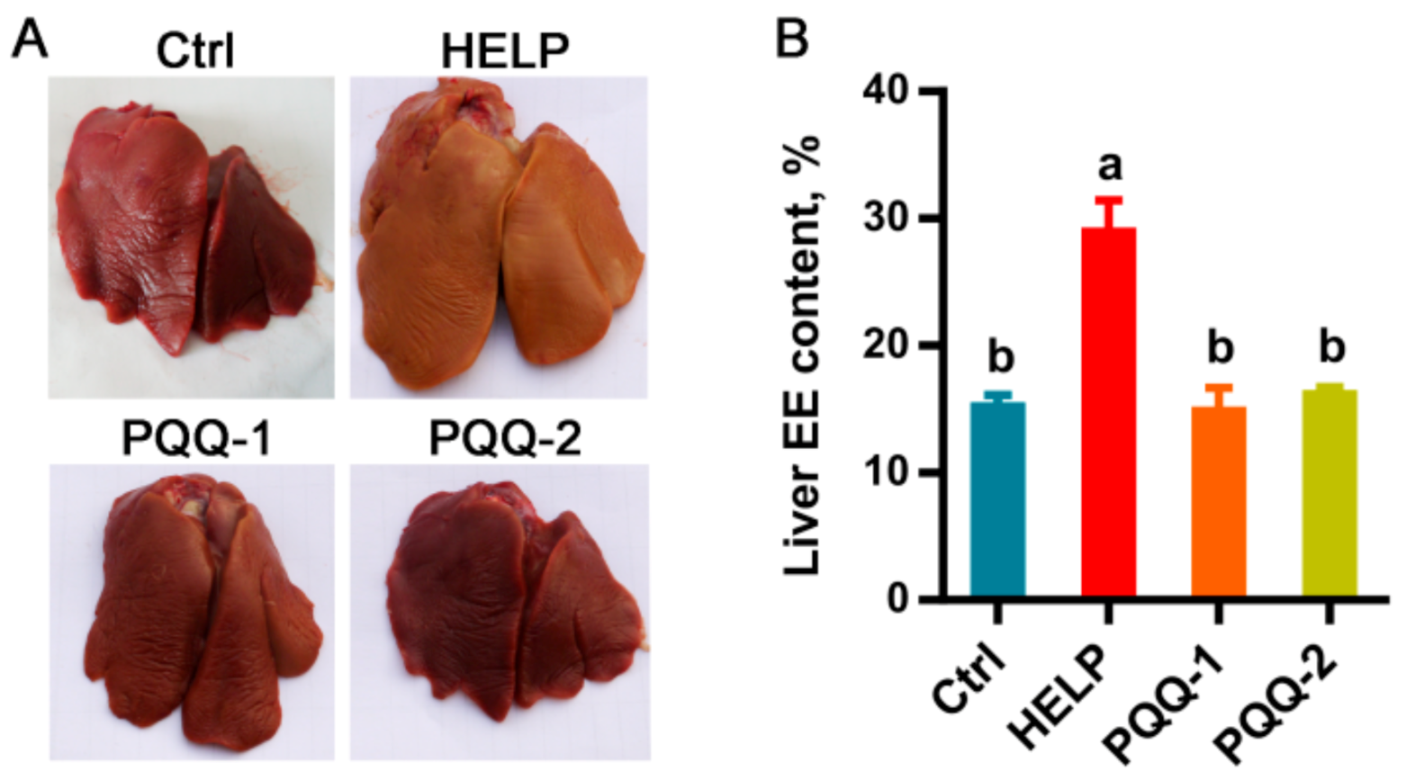
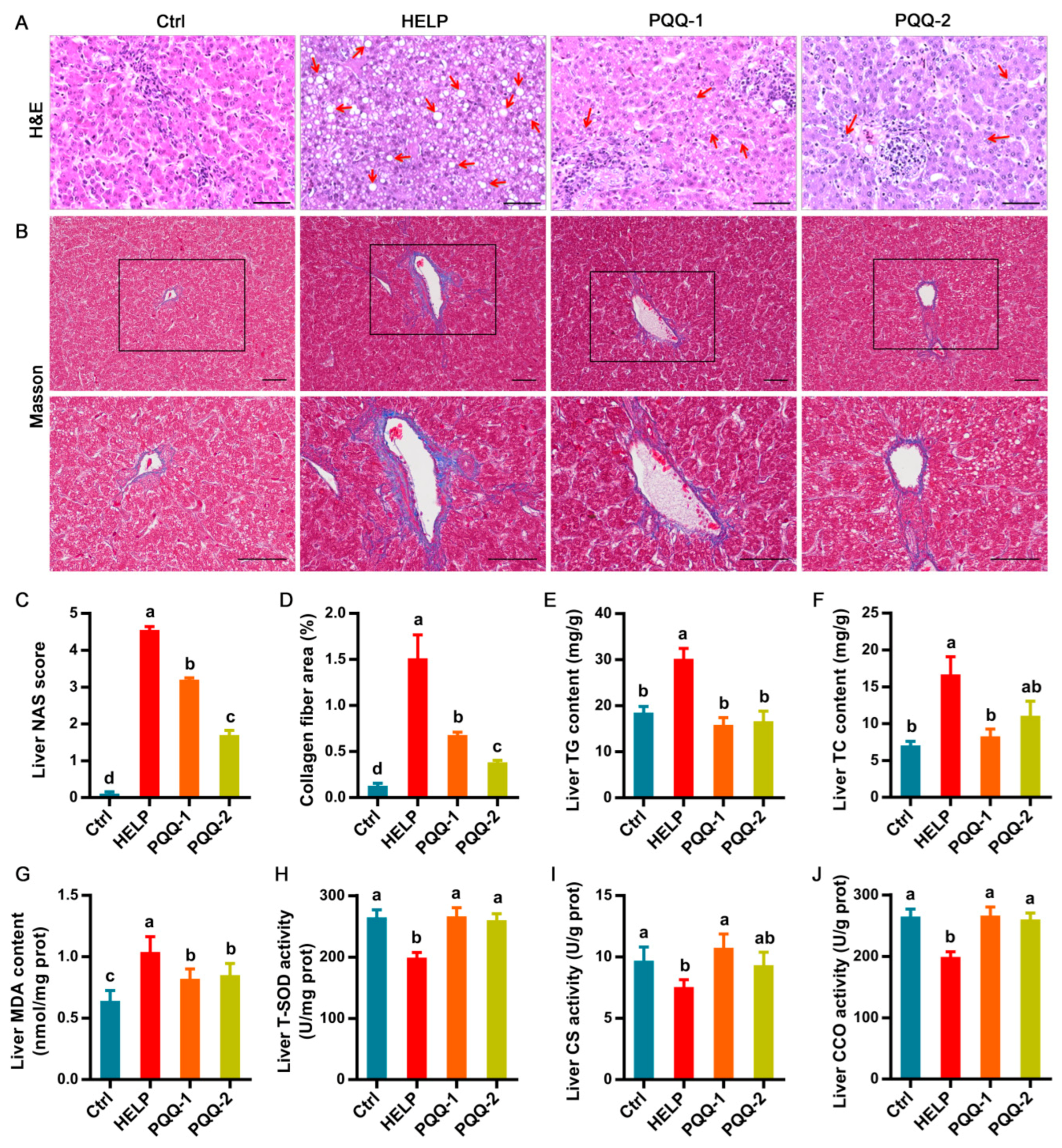
2.2. Effects of Dietary PQQ.Na2 on Liver Function of Laying Hens
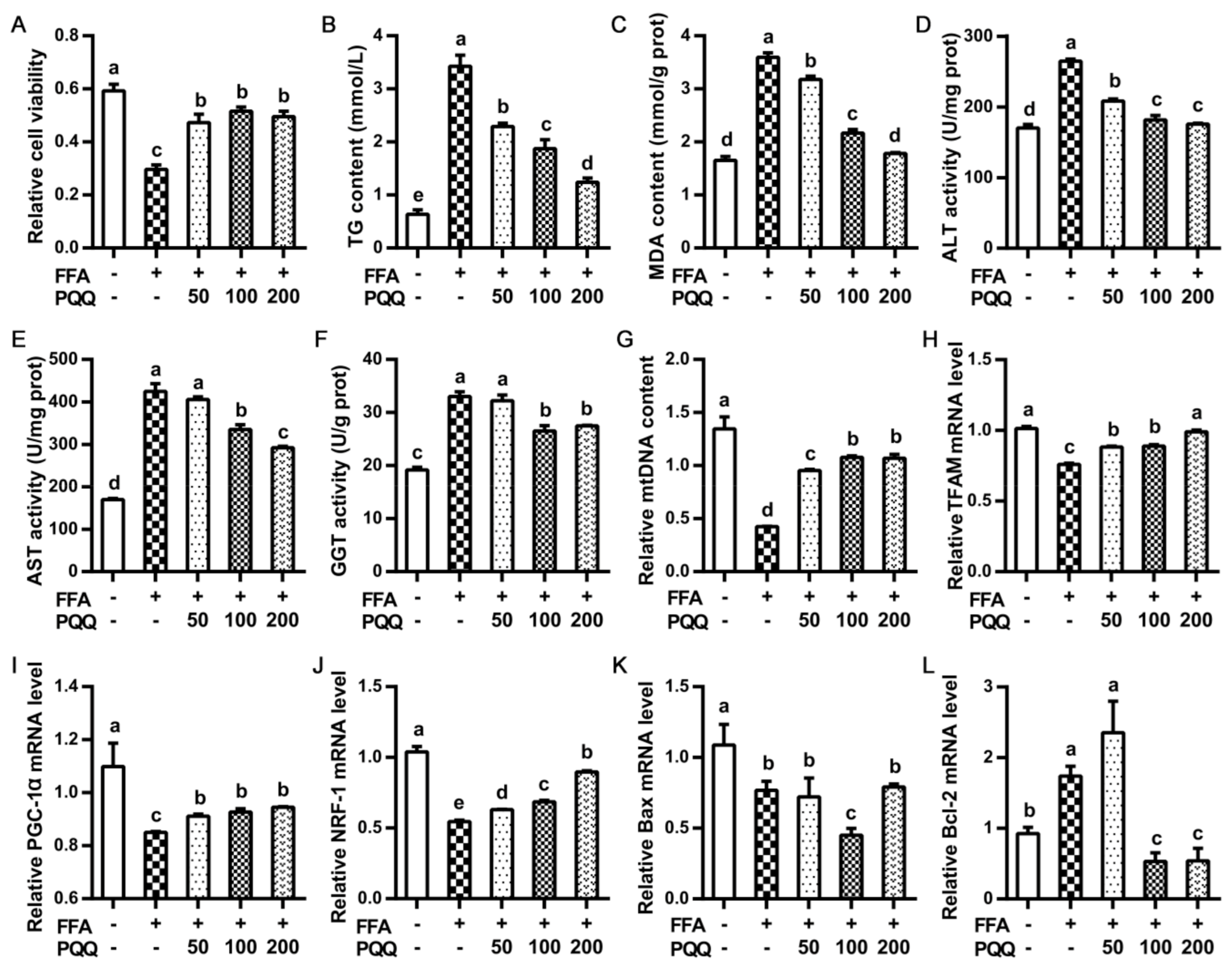
2.3. Effects of PQQ.Na2 on the FFA-Induced Steatosis of Primary Hepatocytes
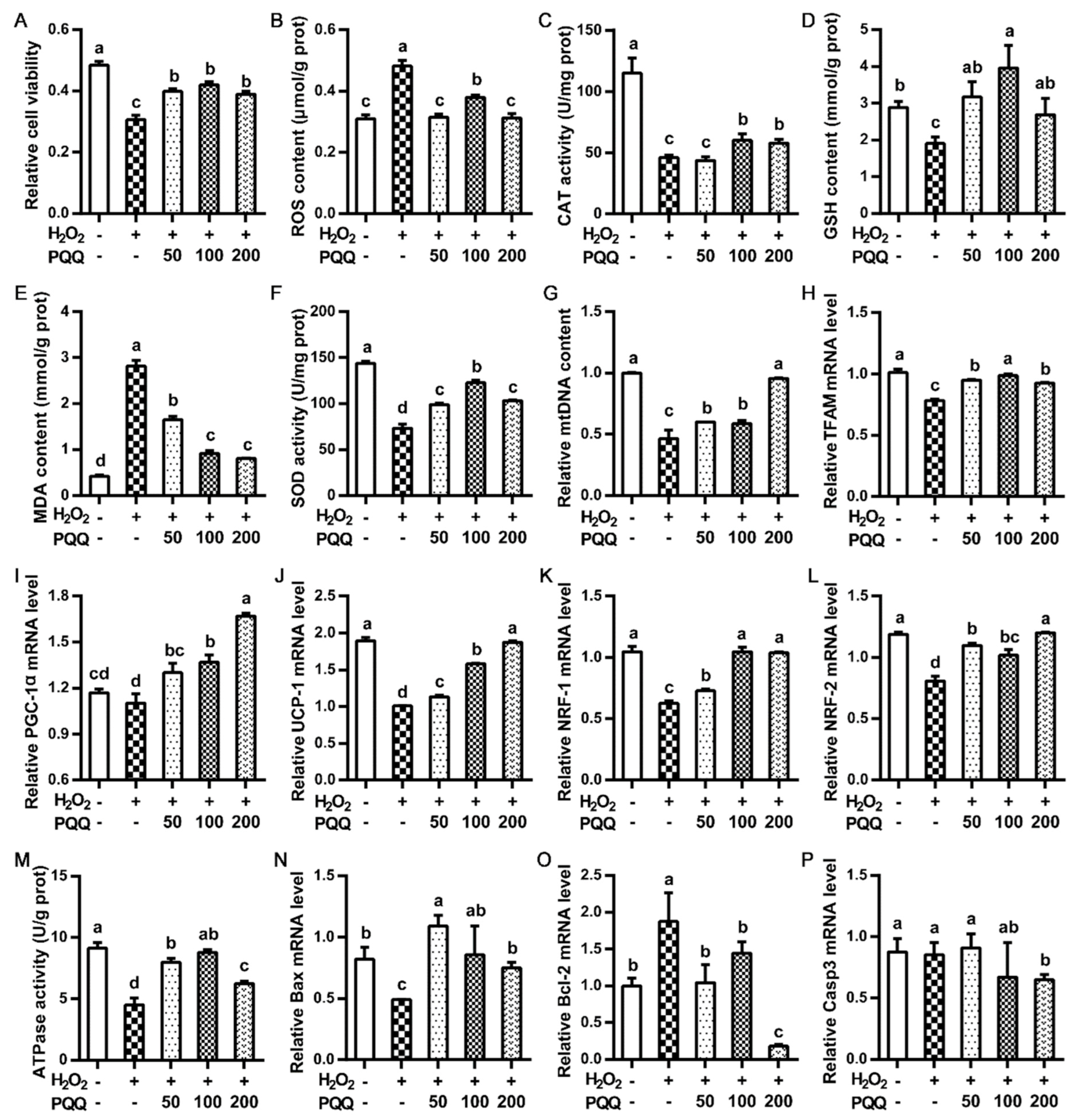
2.4. Effects of PQQ.Na2 on the Primary Hepatocytes upon Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Bird Management and Diet Allocation
4.3. Sample Collection of Animal Experiment
4.4. Histological Analysis of the Liver
4.5. Egg Quality Determination
4.6. Primary Chicken Hepatocytes Isolation and Culture
4.7. Cell Treatment and Sample Collection
4.8. Cell Viability Assay
4.9. Quantitative Real-Time PCR Analysis
4.10. Chemical Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Targher, G. Concordance between MAFLD and NAFLD diagnostic criteria in ‘real-world’ data. Liver Int. 2020, 40, 2879–2880. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Eslam, M.; Kawaguchi, T.; Tsutsumi, T.; Nakano, D.; Yoshinaga, S.; Takahashi, H.; Anzai, K.; George, J.; Torimura, T. MAFLD identifies patients with significant hepatic fibrosis better than NAFLD. Liver Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.E. Nonalcoholic fatty liver disease: An expanding health care epidemic. Clin. Liver Dis. 2018, 22, xiii–xxiv. [Google Scholar] [CrossRef]
- Erickson, S.K. Nonalcoholic fatty liver disease. J. Lipid Res. 2009, 50, S412–S416. [Google Scholar] [CrossRef] [Green Version]
- Jou, J.; Choi, S.S.; Diehl, A.M. Mechanisms of disease progression in nonalcoholic fatty liver disease. Semin. Liver Dis. 2008, 28, 370–379. [Google Scholar] [CrossRef]
- Xu, X.; Lu, L.; Dong, Q.; Li, X.; Zhang, N.; Xin, Y.; Xuan, S. Research advances in the relationship between nonalcoholic fatty liver disease and atherosclerosis. Lipids Health Dis. 2015, 14, 158. [Google Scholar] [CrossRef] [Green Version]
- Pierantonelli, I.; Svegliati-Baroni, G. Nonalcoholic fatty liver disease: Basic pathogenetic mechanisms in the progression from NAFLD to NASH. Transplantation 2019, 103, e1–e13. [Google Scholar] [CrossRef]
- Begriche, K.; Massart, J.; Robin, M.-A.; Bonnet, F.; Fromenty, B. Mitochondrial adaptations and dysfunctions in nonalcoholic fatty liver disease. Hepatology 2013, 58, 1497–1507. [Google Scholar] [CrossRef]
- Gusdon, A.M.; Song, K.-X.; Qu, S. Nonalcoholic fatty liver disease: Pathogenesis and therapeutics from a mitochondria-centric perspective. Oxid. Med. Cell. Longev. 2014, 2014, 637027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Park, J.S.; Roh, Y.S. Molecular insights into the role of mitochondria in non-alcoholic fatty liver disease. Arch. Pharm. Res. 2019, 42, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Grattagliano, I.; Montezinho, L.P.; Oliveira, P.J.; Frühbeck, G.; Gómez-Ambrosi, J.; Montecucco, F.; Carbone, F.; Wieckowski, M.R.; Wang, D.Q.H.; Portincasa, P. Targeting mitochondria to oppose the progression of nonalcoholic fatty liver disease. Biochem. Pharmacol. 2019, 160, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Simões, I.C.M.; Fontes, A.; Pinton, P.; Zischka, H.; Wieckowski, M.R. Mitochondria in non-alcoholic fatty liver disease. Int. J. Biochem. Cell Biol. 2018, 95, 93–99. [Google Scholar] [CrossRef]
- Spahis, S.; Delvin, E.; Borys, J.-M.; Levy, E. Oxidative stress as a critical factor in nonalcoholic fatty liver disease pathogenesis. Antioxid. Redox Signal. 2017, 26, 519–541. [Google Scholar] [CrossRef]
- Farzanegi, P.; Dana, A.; Ebrahimpoor, Z.; Asadi, M.; Azarbayjani, M.A. Mechanisms of beneficial effects of exercise training on non-alcoholic fatty liver disease (NAFLD): Roles of oxidative stress and inflammation. Eur. J. Sport Sci. 2019, 19, 994–1003. [Google Scholar] [CrossRef]
- Sunny, N.E.; Bril, F.; Cusi, K. Mitochondrial adaptation in nonalcoholic fatty liver disease: Novel mechanisms and treatment strategies. Trends Endocrinol. Metab. 2017, 28, 250–260. [Google Scholar] [CrossRef]
- Koliaki, C.; Szendroedi, J.; Kaul, K.; Jelenik, T.; Nowotny, P.; Jankowiak, F.; Herder, C.; Carstensen, M.; Krausch, M.; Knoefel, W.T.; et al. Adaptation of hepatic mitochondrial function in humans with non-alcoholic fatty liver is lost in steatohepatitis. Cell Metab. 2015, 21, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Salisbury, S.A.; Forrest, H.S.; Cruse, W.B.; Kennard, O. A novel coenzyme from bacterial primary alcohol dehydrogenases. Nature 1979, 280, 843–844. [Google Scholar] [CrossRef]
- Kumazawa, T.; Sato, K.; Seno, H.; Ishii, A.; Suzuki, O. Levels of pyrroloquinoline quinone in various foods. Biochem. J. 1995, 307 Pt 2, 331–333. [Google Scholar] [CrossRef] [Green Version]
- Misra, H.S.; Khairnar, N.P.; Barik, A.; Priyadarsini, K.I.; Mohan, H.; Apte, S.K. Pyrroloquinoline-quinone: A reactive oxygen species scavenger in bacteria. FEBS Lett. 2004, 578, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saihara, K.; Kamikubo, R.; Ikemoto, K.; Uchida, K.; Akagawa, M. Pyrroloquinoline quinone, a redox-active o-quinone, stimulates mitochondrial biogenesis by activating the SIRT1/PGC-1α signaling pathway. Biochemistry 2017, 56, 6615–6625. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.S.; Machek, S.B.; Cardaci, T.D.; Wilburn, D.T.; Kim, C.S.; Suezaki, E.S.; Willoughby, D.S. Effects of pyrroloquinoline quinone (PQQ) supplementation on aerobic exercise performance and indices of mitochondrial biogenesis in untrained men. J. Am. Coll. Nutr. 2020, 39, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Bauerly, K.; Harris, C.; Chowanadisai, W.; Graham, J.; Havel, P.J.; Tchaparian, E.; Satre, M.; Karliner, J.S.; Rucker, R.B. Altering pyrroloquinoline quinone nutritional status modulates mitochondrial, lipid, and energy metabolism in rats. PLoS ONE 2011, 6, e21779. [Google Scholar] [CrossRef] [PubMed]
- Jonscher, K.R.; Stewart, M.S.; Alfonso-Garcia, A.; Defelice, B.C.; Wang, X.X.; Luo, Y.; Levi, M.; Heerwagen, M.J.R.; Janssen, R.C.; De, L.H.; et al. Early PQQ supplementation has persistent long-term protective effects on developmental programming of hepatic lipotoxicity and inflammation in obese mice. FASEB J. 2017, 31, 1434–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galton, D.J. Lipogenesis in human adipose tissue. J. Lipid Res. 1968, 9, 19–26. [Google Scholar] [CrossRef]
- Hamid, H.; Zhang, J.Y.; Li, W.X.; Liu, C.; Li, M.L.; Zhao, L.H.; Ji, C.; Ma, Q.G. Interactions between the cecal microbiota and non-alcoholic steatohepatitis using laying hens as the model. Poult. Sci. 2019, 98, 2509–2521. [Google Scholar] [CrossRef]
- Tsai, M.T.; Chen, Y.J.; Chen, C.Y.; Tsai, M.H.; Han, C.L.; Chen, Y.J.; Mersmann, H.J.; Ding, S.T. Identification of potential plasma biomarkers for nonalcoholic fatty liver disease by integrating transcriptomics and proteomics in laying hens. J. Nutr. 2017, 147, 293–303. [Google Scholar] [CrossRef]
- Chen, W.L.; Wei, H.W.; Chiu, W.Z.; Kang, C.H.; Lin, T.H.; Hung, C.C.; Chen, M.C.; Shieh, M.S.; Lee, C.C.; Lee, H.M. Metformin regulates hepatic lipid metabolism through activating AMP-activated protein kinase and inducing ATGL in laying hens. Eur. J. Pharmacol. 2011, 671, 107–112. [Google Scholar] [CrossRef]
- Dajani, A.; AbuHammour, A. Treatment of nonalcoholic fatty liver disease: Where do we stand? An overview. Saudi J. Gastroenterol. 2016, 22, 91–105. [Google Scholar]
- Rozenboim, I.; Mahato, J.; Cohen, N.A.; Tirosh, O. Low protein and high-energy diet: A possible natural cause of fatty liver hemorrhagic syndrome in caged White Leghorn laying hens. Poult. Sci. 2016, 95, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Ruan, J.; Luo, J.; Wang, T.; Yang, F.; Cao, H.; Huang, J.; Hu, G. Abnormal histopathology, fat percent and hepatic apolipoprotein A I and apolipoprotein B100 mRNA expression in fatty liver hemorrhagic syndrome and their improvement by soybean lecithin. Poult. Sci. 2017, 96, 3559–3563. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ruan, J.; Wang, T.; Luo, J.; Cao, H.; Song, Y.; Huang, J.; Hu, G. Improving effect of dietary soybean phospholipids supplement on hepatic and serum indexes relevant to fatty liver hemorrhagic syndrome in laying hens. Anim. Sci. J. 2017, 88, 1860–1869. [Google Scholar] [CrossRef] [PubMed]
- Cherian, G. Nutrition and metabolism in poultry: Role of lipids in early diet. J. Anim. Sci. Biotechnol. 2015, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, F.M.; Gershwin, M.E.; Rucker, R.B. Dietary pyrroloquinoline quinone: Growth and immune response in BALB/c mice. J. Nutr. 1994, 124, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, H.J.; Xu, L.; Long, C.; Samuel, K.G.; Yue, H.Y.; Sun, L.L.; Wu, S.G.; Qi, G.H. Dietary supplementation of pyrroloquinoline quinone disodium protects against oxidative stress and liver damage in laying hens fed an oxidized sunflower oil-added diet. Animal 2016, 10, 1129–1136. [Google Scholar] [CrossRef]
- Nassir, F.; Adewole, O.L.; Brunt, E.M.; Abumrad, N.A. CD36 deletion reduces VLDL secretion, modulates liver prostaglandins, and exacerbates hepatic steatosis in ob/ob mice. J. Lipid Res. 2013, 54, 2988–2997. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, T.; Xu, C.; Wang, D.; Ren, J.; Li, Y.; Tian, Y.; Wang, Y.; Jiao, Y.; Kang, X.; et al. Transcriptome profile of liver at different physiological stages reveals potential mode for lipid metabolism in laying hens. BMC Genom. 2015, 16, 763. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.K.; Kim, J.S.; Ahn, H.J.; Hwang, J.H.; Kim, J.M.; Lee, H.T.; An, B.K.; Kang, C.W. Changes in hepatic lipid parameters and hepatic messenger ribonucleic acid expression following estradiol administration in laying hens (Gallus domesticus). Poult. Sci. 2010, 89, 2660–2667. [Google Scholar] [CrossRef]
- Jiang, S.; Cheng, H.W.; Cui, L.Y.; Zhou, Z.L.; Hou, J.F. Changes of blood parameters associated with bone remodeling following experimentally induced fatty liver disorder in laying hens. Poult. Sci. 2013, 92, 1443–1453. [Google Scholar] [CrossRef]
- Trott, K.A.; Giannitti, F.; Rimoldi, G.; Hill, A.; Woods, L.; Barr, B.; Anderson, M.; Mete, A. Fatty liver hemorrhagic syndrome in the backyard chicken: A retrospective histopathologic case series. Vet. Pathol. 2014, 51, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.-H.; Zhao, Y.; Ge, X.; Yuan, S.-J.; Wang, J.-H.; Zhi, J.-J.; Yang, Y.-X.; Du, B.-H.; Guo, W.-J.; Wang, S.-S.; et al. Production and radioprotective effects of pyrroloquinoline quinone. Int. J. Mol. Sci. 2011, 12, 8913–8923. [Google Scholar] [CrossRef] [PubMed]
- Neto, S.A.; Suda, E.L.; Xu, S.; Meredith, M.T.; De Andrade, A.R.; Minteer, S.D. Direct electron transfer-based bioanodes for ethanol biofuel cells using PQQ-dependent alcohol and aldehyde dehydrogenases. Electrochim. Acta 2013, 87, 323–329. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, N.; Miao, D. Radioprotective effects of pyrroloquinoline quinone on parotid glands in C57BL/6J mice. Exp. Ther. Med. 2016, 12, 3685–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Chen, N.; Miao, D. Biological effects of pyrroloquinoline quinone on liver damage in Bmi-1 knockout mice. Exp. Ther. Med. 2015, 10, 451–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasakura, H.; Moribe, H.; Nakano, M.; Ikemoto, K.; Takeuchi, K.; Mori, I. Lifespan extension by peroxidase and dual oxidase-mediated ROS signaling through pyrroloquinoline quinone in. J. Cell Sci. 2017, 130, 2631–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Sun, T.; Wu, J.; Kalionis, B.; Zhang, C.; Yuan, D.; Huang, J.; Cai, W.; Fang, H.; Xia, S. Icariin intervenes in cardiac inflammaging through upregulation of SIRT6 enzyme activity and inhibition of the NF-kappa B pathway. Biomed Res. Int. 2015, 2015, 895976. [Google Scholar] [CrossRef]
- Harris, C.B.; Chowanadisai, W.; Mishchuk, D.O.; Satre, M.A.; Slupsky, C.M.; Rucker, R.B. Dietary pyrroloquinoline quinone (PQQ) alters indicators of inflammation and mitochondrial-related metabolism in human subjects. J. Nutr. Biochem. 2013, 24, 2076–2084. [Google Scholar] [CrossRef] [Green Version]
- Senior, J.R. Alanine aminotransferase: A clinical and regulatory tool for detecting liver injury-past, present, and future. Clin. Pharmacol. Ther. 2012, 92, 332–339. [Google Scholar] [CrossRef]
- Shim, J.-Y.; Kim, M.-H.; Kim, H.-D.; Ahn, J.-Y.; Yun, Y.-S.; Song, J.-Y. Protective action of the immunomodulator ginsan against carbon tetrachloride-induced liver injury via control of oxidative stress and the inflammatory response. Toxicol. Appl. Pharmacol. 2010, 242, 318–325. [Google Scholar] [CrossRef]
- Vergani, L. Fatty acids and effects on in vitro and in vivo models of liver steatosis. Curr. Med. Chem. 2019, 26, 3439–3456. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Tian, K.; Zhang, T.; Fan, C.-H.; Zhou, P.; Zeng, D.; Zhao, S.; Li, L.-S.; Smith, H.S.; Li, J.; et al. Cyanate induces oxidative stress injury and abnormal lipid metabolism in Liver through Nrf2/HO-1. Molecules 2019, 24, 3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Han, N.; Zhao, K.; Li, Y.; Chi, Y.; Wang, B. Protective effects of pyrroloquinoline quinine against oxidative stress-induced cellular senescence and inflammation in human renal tubular epithelial cells via Keap1/Nrf2 signaling pathway. Int. Immunopharmacol. 2019, 72, 445–453. [Google Scholar] [CrossRef]
- Jiang, C.; Jiang, L.; Li, Q.; Liu, X.; Zhang, T.; Yang, G.; Zhang, C.; Wang, N.; Sun, X.; Jiang, L. Pyrroloquinoline quinine ameliorates doxorubicin-induced autophagy-dependent apoptosis via lysosomal-mitochondrial axis in vascular endothelial cells. Toxicology 2019, 425, 152238. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wang, X.; Sun, H. The role of mitochondria in cellular toxicity as a potential drug target. Cell Biol. Toxicol. 2018, 34, 87–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedgwick, B.; Riches, K.; Bageghni, S.A.; O’Regan, D.J.; Porter, K.E.; Turner, N.A. Investigating inherent functional differences between human cardiac fibroblasts cultured from nondiabetic and Type 2 diabetic donors. Cardiovasc. Pathol. 2014, 23, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, K.; Anjaneyulu, M.; Choi, J.; Kumar, P.; Salimian, M.; Ho, C.-Y.; Russell, J.W. Role of mitochondria in diabetic peripheral neuropathy: Influencing the NAD-dependent SIRT1-PGC-1α-TFAM pathway. Int. Rev. Neurobiol. 2019, 145, 177–209. [Google Scholar]
- Que, F.G.; Gores, G.J. Cell death by apoptosis: Basic concepts and disease relevance for the gastroenterologist. Gastroenterology 1996, 110, 1238–1243. [Google Scholar] [CrossRef]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Zhang, T.; Zhang, R.; Liu, R.; Chen, Y. Molecular mechanism on cadmium-induced activity changes of catalase and superoxide dismutase. Int. J. Biol. Macromol. 2015, 77, 59–67. [Google Scholar] [CrossRef]
- Boysen, G. The glutathione conundrum: Stoichiometric disconnect between its formation and oxidative stress. Chem. Res. Toxicol. 2017, 30, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.H.; Kim, K.S.; Choi, S.M.; Yang, H.S.; Yoon, Y. The effects of uncoupling protein-1 genotype on lipoprotein cholesterol level in Korean obese subjects. Metabolism 2004, 53, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Pandey, S.K.; Kumar, G.N. Pyrroloquinoline quinone-secreting probiotic Escherichia coli Nissle 1917 ameliorates ethanol-induced oxidative damage and hyperlipidemia in rats. Alcohol. Clin. Exp. Res. 2014, 38, 2127–2137. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Sun, J.; Wu, J.; Chen, L. Pyrroloquinoline quinone prevents knee osteoarthritis by inhibiting oxidative stress and chondrocyte senescence. Am. J. Transl. Res. 2019, 11, 1460–1472. [Google Scholar]
- Geng, Q.; Gao, H.; Yang, R.; Guo, K.; Miao, D. Pyrroloquinoline quinone prevents estrogen deficiency-induced osteoporosis by inhibiting oxidative stress and osteocyte senescence. Int. J. Biol. Sci. 2019, 15, 58–68. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Wang, Y.; Zhao, K.; Chi, Y.; Wang, B. Pyrroloquinoline quinine protects HK-2cells against high glucose-induced oxidative stress and apoptosis through Sirt3 and PI3K/Akt/FoxO3a signaling pathway. Biochem. Biophys. Res. Commun. 2019, 508, 398–404. [Google Scholar] [CrossRef]
- Killgore, J.; Smidt, C.; Duich, L.; Romero-Chapman, N.; Tinker, D.; Reiser, K.; Melko, M.; Hyde, D.; Rucker, R. Nutritional importance of pyrrolo quinoline quinone. Science 1989, 245, 850–852. [Google Scholar] [CrossRef]
- Steinberg, F.; Stites, T.E.; Anderson, P.; Storms, D.; Rucker, R. Pyrroloquinoline quinone improves growth and reproductive performance in mice fed chemically defined diets. Exp. Biol. Med. 2003, 228, 160–166. [Google Scholar] [CrossRef]
- Bauerly, K.A.; Storms, D.H.; Harris, C.B.; Hajizadeh, S.; Sun, M.Y.; Cheung, C.P.; Satre, M.A.; Fascetti, A.J.; Tchaparian, E.; Rucker, R.B. Pyrroloquinoline quinone nutritional status alters lysine metabolism and modulates mitochondrial DNA content in the mouse and rat. Biochim. Biophys. Acta 2006, 1760, 1741–1748. [Google Scholar] [CrossRef]
- Chowanadisai, W.; Bauerly, K.A.; Tchaparian, E.; Wong, A.; Cortopassi, G.A.; Rucker, R.B. Pyrroloquinoline quinone stimulates mitochondrial biogenesis through cAMP response element-binding protein phosphorylation and increased PGC-1alpha expression. J. Biol. Chem. 2010, 285, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Tchaparian, E.; Marshal, L.; Cutler, G.; Bauerly, K.; Chowanadisai, W.; Satre, M.; Harris, C.; Rucker, R. Identification of transcriptional networks responding to pyrroloquinoline quinone dietary supplementation and their influence on thioredoxin expression, and the JAK/STAT and MAPK pathways. Biochem. J. 2010, 429, 515–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tracy, S.; David, S.; Kathryn, B.; James, M.; Calliandra, H.; Andrea, F.; Quinton, R.; Eskouhie, T.; Michael, S.; Rucker, R.B. Pyrroloquinoline quinone modulates mitochondrial quantity and function in mice. J. Nutr. 2006, 136, 390–396. [Google Scholar]
- Fraslin, J.M.; Touquette, L.; Douaire, M.; Menezo, Y.; Guillemot, J.C.; Mallard, J. Isolation and long-term maintenance of differentiated adult chicken hepatocytes in primary culture. In Vitro Cell. Dev. Biol. 1992, 28A, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Mi, J.; Lu, L.; Zeng, X.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef]
- Zheng, T.; Jia, R.; Cao, L.; Du, J.; Gu, Z.; He, Q.; Xu, P.; Yin, G. Effects of chronic glyphosate exposure on antioxdative status, metabolism and immune response in tilapia (GIFT, Oreochromis niloticus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 239, 108878. [Google Scholar] [CrossRef]
- Du, J.; Jia, R.; Cao, L.P.; Ding, W.; Xu, P.; Yin, G. Effects of Rhizoma Alismatis extract on biochemical indices and adipose gene expression in oleic acid-induced hepatocyte injury in Jian carp (Cyprinus carpio var. Jian). Fish Physiol. Biochem. 2018, 44, 747–768. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Ctrl | HELP | PQQ-1 | PQQ-2 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Performance | ||||||
| Laying rate, % | 91.02 | 91.04 | 91.36 | 92.71 | 0.58 | 0.71 |
| Average egg weight, g | 58.87 | 59.33 | 58.41 | 59.22 | 0.25 | 0.59 |
| Average daily feed intake, g | 117.72 b | 132.67 a | 118.20 b | 122.38 b | 1.99 | 0.03 |
| Feed/egg | 2.20 b | 2.43 a | 2.35 ab | 2.37 ab | 0.04 | 0.17 |
| Egg quality | ||||||
| Eggshell strength, N/m2 | 43.76 | 43.65 | 44.97 | 40.42 | 0.77 | 0.19 |
| Eggshell thickness, mm | 0.43 | 0.43 | 0.44 | 0.44 | 0.002 | 0.77 |
| Egg shape index | 1.30 | 1.30 | 1.31 | 1.30 | 0.005 | 0.86 |
| Albumen height, mm | 6.65 a | 5.64 b | 6.57 a | 6.51 a | 0.11 | <0.01 |
| Yolk color | 6.36 a | 5.57 b | 5.78 ab | 5.89 ab | 0.12 | 0.02 |
| Haugh unit | 78.96 a | 72.44 b | 79.38 a | 78.26 a | 0.74 | <0.01 |
| Items | Ctrl | HELP | PQQ-1 | PQQ-2 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Triglyceride (TG), mmol/L | 11.22 b | 15.38 a | 8.25 b | 10.63 b | 0.78 | <0.01 |
| Total cholesterol (TC), mmol/L | 3.03 b | 3.94 a | 2.77 b | 3.19 b | 0.23 | <0.01 |
| Alanine transaminase (ALT), U/L | 72.47 a | 84.66 a | 50.33 b | 54.26 b | 3.93 | <0.01 |
| Aspartate aminotransferase (AST), U/L | 7.47 | 8.66 | 8.61 | 7.65 | 0.58 | 0.86 |
| Cholinesterase (ChE), U/L | 62.81 b | 85.69 a | 87.84 a | 78.75 a | 3.53 | <0.01 |
| Lactate dehydrogenase (LDH), U/μL | 4.06 b | 5.13 a | 3.64 b | 3.46 b | 0.17 | <0.01 |
| Malondialdehyde (MDA), nmol/mL | 6.40 b | 9.09 a | 6.74 b | 8.76 a | 0.37 | <0.01 |
| Superoxide dismutase (SOD), U/mL | 274.49 a | 234.35 b | 274.33 a | 269.56 a | 6.21 | 0.04 |
| High-density lipoprotein cholesterol (HDL-C), mmol/L | 1.57 | 1.54 | 1.55 | 1.58 | 0.03 | 0.97 |
| Low-density lipoprotein cholesterol (LDL-C), mmol/L | 1.72 b | 2.68 a | 1.40 b | 1.61b | 0.13 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, K.; Zhao, Q.; Wang, J.; Qi, G.-H.; Wu, S.-G.; Zhang, H.-J. Effects of Pyrroloquinoline Quinone on Lipid Metabolism and Anti-Oxidative Capacity in a High-Fat-Diet Metabolic Dysfunction-Associated Fatty Liver Disease Chick Model. Int. J. Mol. Sci. 2021, 22, 1458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031458
Qiu K, Zhao Q, Wang J, Qi G-H, Wu S-G, Zhang H-J. Effects of Pyrroloquinoline Quinone on Lipid Metabolism and Anti-Oxidative Capacity in a High-Fat-Diet Metabolic Dysfunction-Associated Fatty Liver Disease Chick Model. International Journal of Molecular Sciences. 2021; 22(3):1458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031458
Chicago/Turabian StyleQiu, Kai, Qin Zhao, Jing Wang, Guang-Hai Qi, Shu-Geng Wu, and Hai-Jun Zhang. 2021. "Effects of Pyrroloquinoline Quinone on Lipid Metabolism and Anti-Oxidative Capacity in a High-Fat-Diet Metabolic Dysfunction-Associated Fatty Liver Disease Chick Model" International Journal of Molecular Sciences 22, no. 3: 1458. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031458