
Reticular Basement Membrane Thickness Is Associated with Growth- and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Patient Description and Basic Laboratory Tests
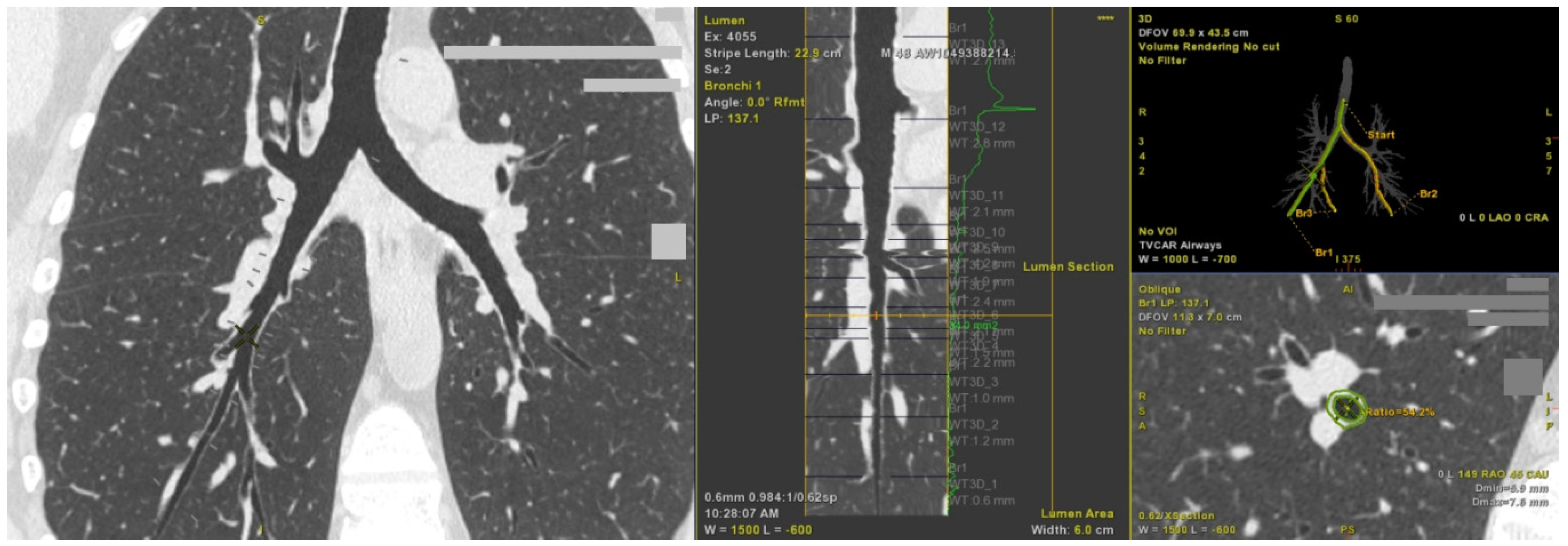
2.2. Computed Tomography Imaging Shows Airway Wall Thickening in Asthma Patients with Persistent Airflow Limitation
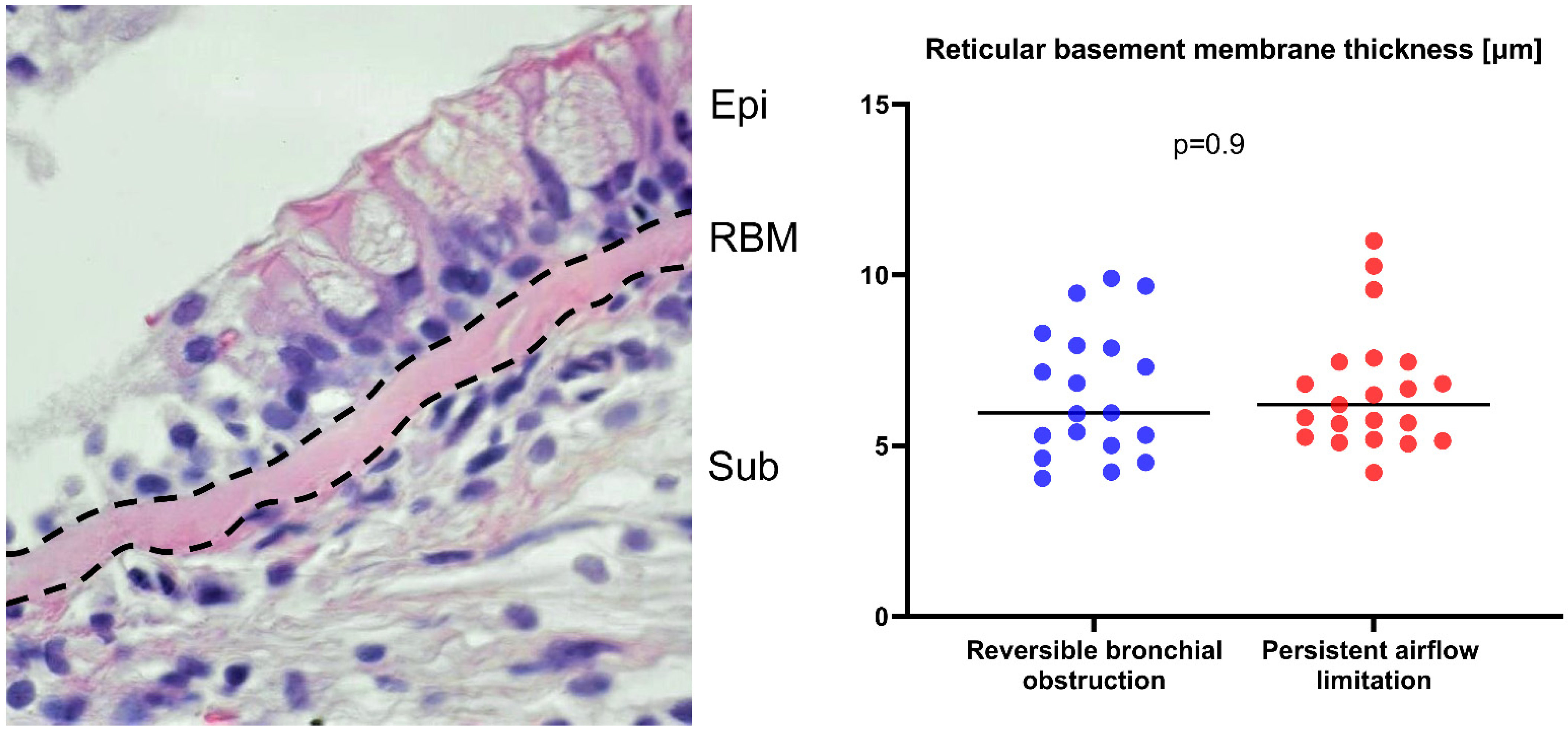
2.3. Reticular Basement Membrane Thickness Is Not Increased in Asthma Patients with Persistent Airflow Limitation
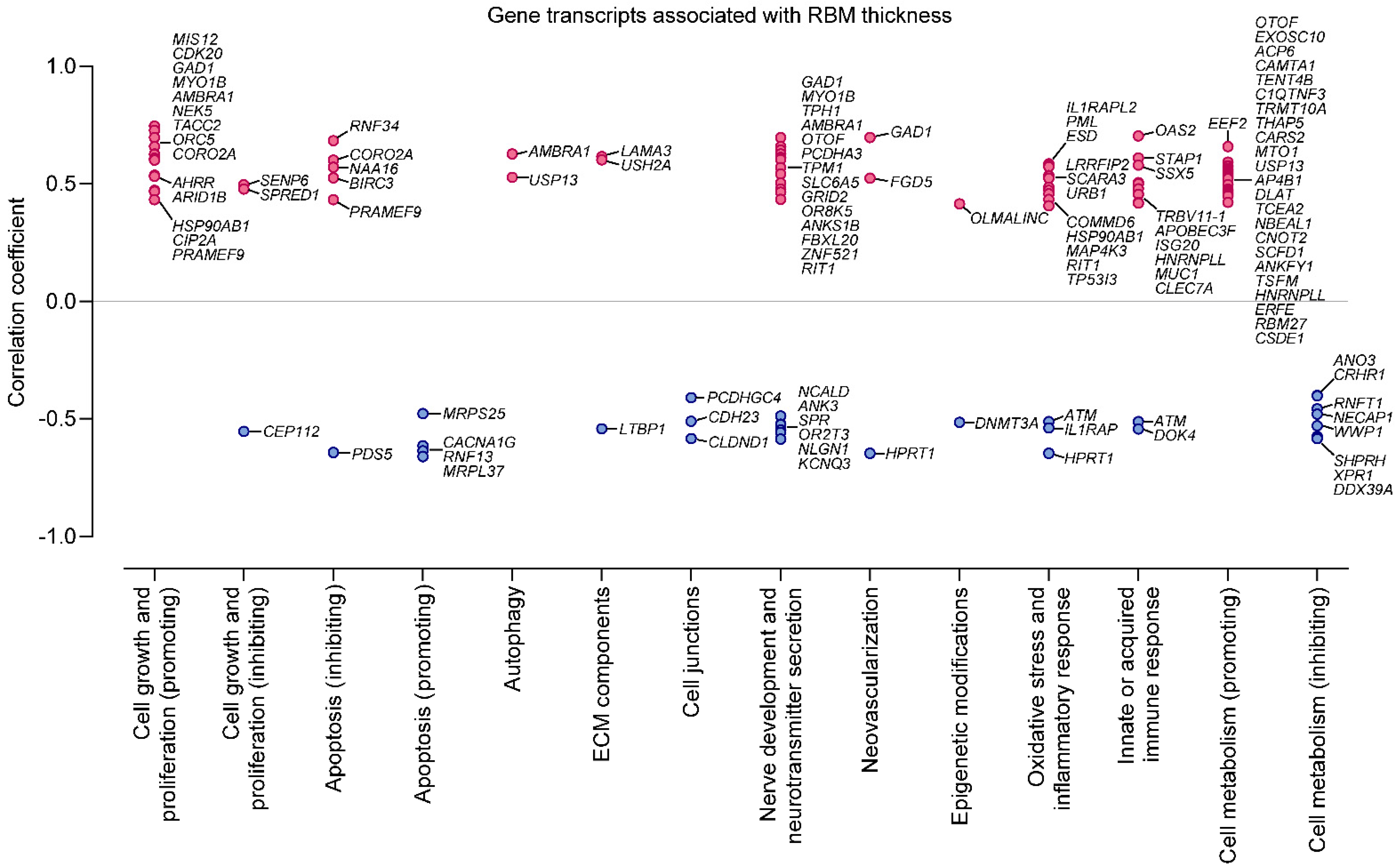
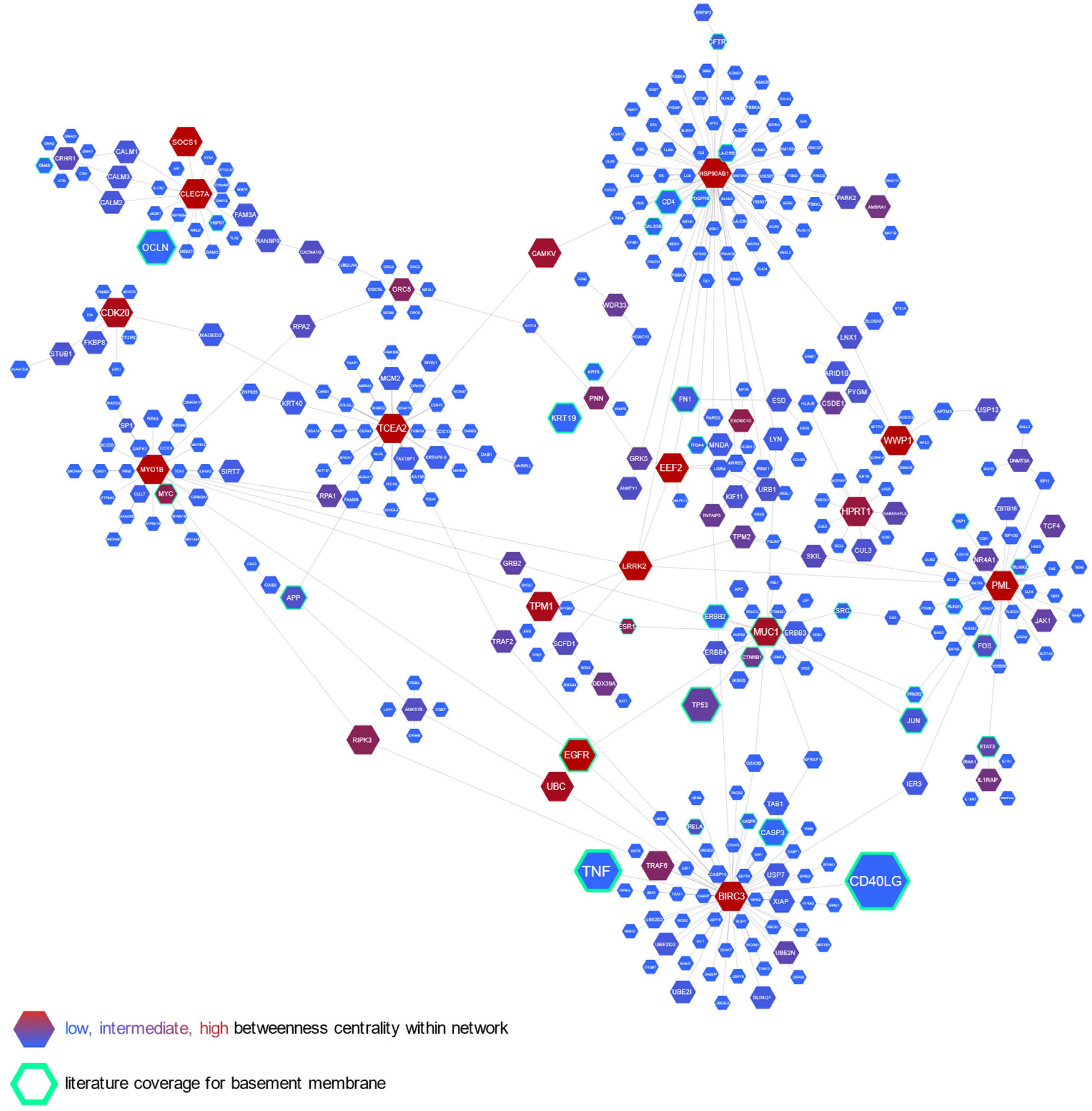
2.4. Increased Reticular Basement Membrane Thickness Is Associated with Fibrosis- and Growth-Promoting Pattern of Gene Expression in Bronchial Epithelium
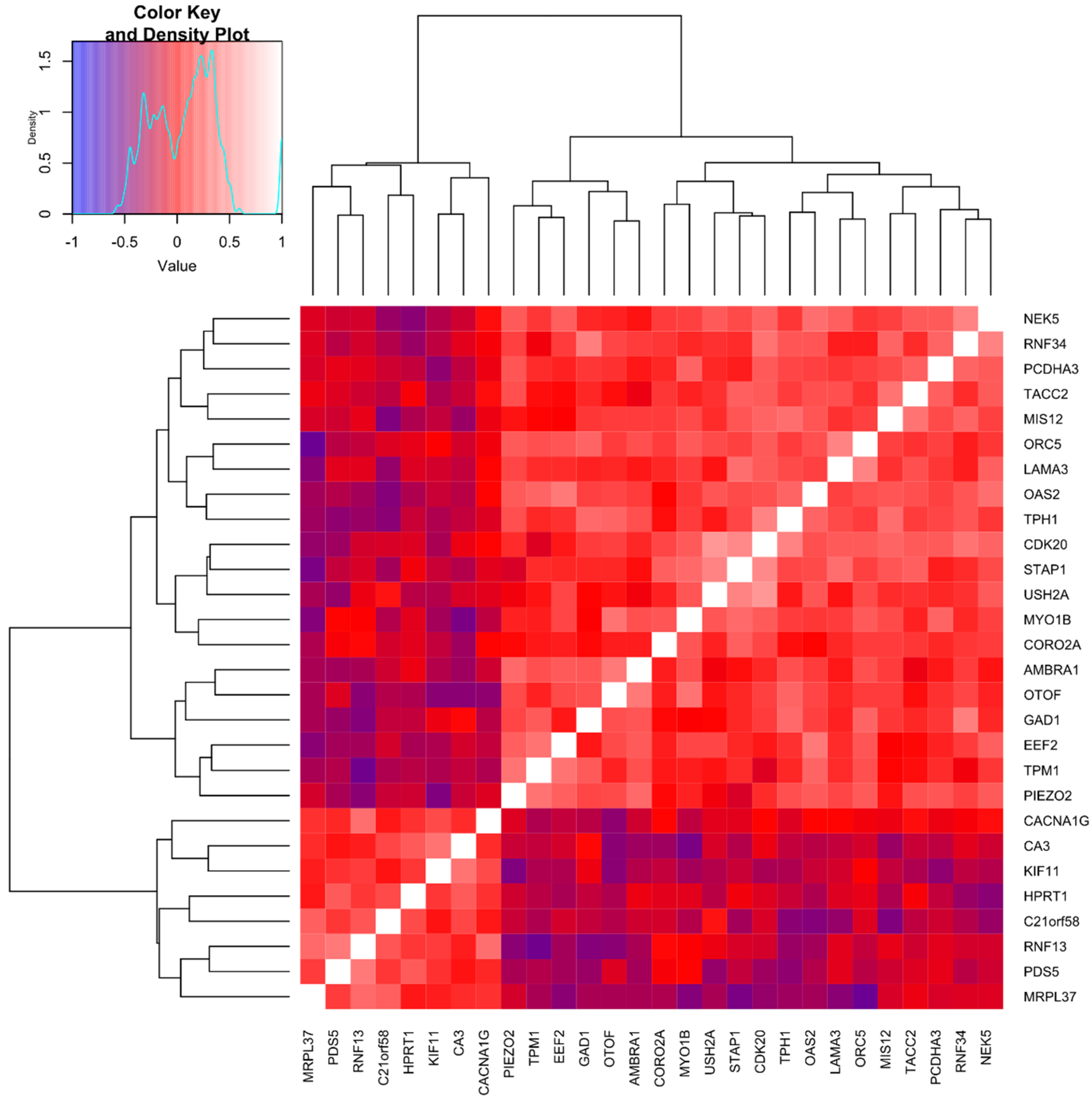
2.5. Cluster Analysis Integrating Molecular and Remodeling Data Reveals Subgroups of Asthma Patients with Unique Bronchial Epithelial Gene Expression Patterns
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Lung CT
4.3. Bronchofiberoscopy, Endobronchial Biopsy, and Bronchial Brush Biopsy
4.4. Bronchoalveolar Lavage Fluid Analysis
4.5. Blood Laboratory Investigations
4.6. Histologic Examination
4.7. RNA Preparation for Microarray Analysis
4.8. Microarray Data Retrieval and Bioconductor
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hough, K.P.; Curtiss, M.L.; Blain, T.J.; Liu, R.M.; Trevor, J.; Deshane, J.S.; Thannickal, V.J. Airway Remodeling in Asthma. Front. Med. 2020, 7, 191. [Google Scholar] [CrossRef]
- Busse, W.W.; Lemanske, R.F. Asthma. N. Engl. J. Med. 2001, 344, 350–362. [Google Scholar] [CrossRef]
- Trejo Bittar, H.E.; Yousem, S.A.; Wenzel, S.E. Pathobiology of Severe Asthma. Annu. Rev. Pathol. Mech. Dis. 2015, 10, 511–545. [Google Scholar] [CrossRef] [Green Version]
- Bazan-Socha, S.; Bukiej, A.; Marcinkiewicz, C.; Musial, J. Integrins in Pulmonary Inflammatory Diseases. Curr. Pharm. Des. 2005, 11. [Google Scholar] [CrossRef]
- Laulajainen-Hongisto, A.; Toppila-Salmi, S.K.; Luukkainen, A.; Kern, R. Airway Epithelial Dynamics in Allergy and Related Chronic Inflammatory Airway Diseases. Front. Cell Dev. Biol. 2020, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Lambrecht, B.N.; Hammad, H. The Airway Epithelium in Asthma. Nat. Med. 2012, 18, 684–692. [Google Scholar] [CrossRef]
- Hellings, P.W.; Steelant, B. Epithelial Barriers in Allergy and Asthma. J. Allergy Clin. Immunol. 2020, 145, 1499–1509. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, C.; Galaktionov, K. P42, a Novel Cyclin-Dependent Kinase-Activating Kinase in Mammalian Cells. J. Biol. Chem. 2004, 279, 4507–4514. [Google Scholar] [CrossRef] [Green Version]
- Goshima, G.; Kiyomitsu, T.; Yoda, K.; Yanagida, M. Human Centromere Chromatin Protein HMis12, Essential for Equal Segregation, Is Independent of CENP-A Loading Pathway. J. Cell Biol. 2003, 160, 25–39. [Google Scholar] [CrossRef]
- Carretero, M.; Ruiz-Torres, M.; Rodríguez-Corsino, M.; Barthelemy, I.; Losada, A. Pds5B Is Required for Cohesion Establishment and Aurora B Accumulation at Centromeres. EMBO J. 2013, 32, 2938–2949. [Google Scholar] [CrossRef] [Green Version]
- Åbrink, M.; Aveskogh, M.; Hellman, L. Isolation of CDNA Clones for 42 Different Krüppel-Related Zinc Finger Proteins Expressed in the Human Monoblast Cell Line U-937. DNA Cell Biol. 1995, 14, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Matz, A.; Lee, S.J.; Schwedhelm-Domeyer, N.; Zanini, D.; Holubowska, A.; Kannan, M.; Farnworth, M.; Jahn, O.; Göpfert, M.C.; Stegmüller, J. Regulation of Neuronal Survival and Morphology by the E3 Ubiquitin Ligase RNF157. Cell Death Differ. 2015, 22, 626–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faccio, L.; Fusco, C.; Chen, A.; Martinotti, S.; Bonventre, J.V.; Zervos, A.S. Characterization of a Novel Human Serine Protease That Has Extensive Homology to Bacterial Heat Shock Endoprotease HtrA and Is Regulated by Kidney Ischemia. J. Biol. Chem. 2000, 275, 2581–2588. [Google Scholar] [CrossRef] [Green Version]
- Giunta, C.; Elçioglu, N.H.; Albrecht, B.; Eich, G.; Chambaz, C.; Janecke, A.R.; Yeowell, H.; Weis, M.A.; Eyre, D.R.; Kraenzlin, M.; et al. Spondylocheiro Dysplastic Form of the Ehlers-Danlos Syndrome—An Autosomal-Recessive Entity Caused by Mutations in the Zinc Transporter Gene SLC39A13. Am. J. Hum. Genet. 2008, 82, 1290–1305. [Google Scholar] [CrossRef] [Green Version]
- Bonen, A.; Heynen, M.; Hatta, H. Distribution of Monocarboxylate Transporters MCT1-MCT8 in Rat Tissues and Human Skeletal Muscle. Appl. Physiol. Nutr. Metab. 2006, 31, 31–39. [Google Scholar] [CrossRef]
- Mingot, J.M.; Kostka, S.; Kraft, R.; Hartmann, E.; Görlich, D. Importin 13: A Novel Mediator of Nuclear Import and Export. EMBO J. 2001, 20, 3685–3694. [Google Scholar] [CrossRef] [Green Version]
- Cloning of the LamA3 Gene Encoding the Alpha 3 Chain of the Adhesive Ligand Epiligrin. Expression in Wound Repair—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/8077230/ (accessed on 30 October 2020).
- Weston, M.D.; Eudy, J.D.; Fujita, S.; Yao, S.F.; Usami, S.; Cremers, C.; Greenburg, J.; Ramesar, R.; Martini, A.; Moller, C.; et al. Genomic Structure and Identification of Novel Mutations in Usherin, the Gene Responsible for Usher Syndrome Type IIa. Am. J. Hum. Genet. 2000, 66, 1199–1210. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.G.; Czabotar, P.E.; Policheni, A.N.; Caminschi, I.; San Wan, S.; Kitsoulis, S.; Tullett, K.M.; Robin, A.Y.; Brammananth, R.; van Delft, M.F.; et al. The Dendritic Cell Receptor Clec9A Binds Damaged Cells via Exposed Actin Filaments. Immunity 2012, 36, 646–657. [Google Scholar] [CrossRef] [Green Version]
- Kristiansen, H.; Gad, H.H.; Eskildsen-Larsen, S.; Despres, P.; Hartmann, R. The Oligoadenylate Synthetase Family: An Ancient Protein Family with Multiple Antiviral Activities. J. Interferon Cytokine Res. 2011, 31, 41–47. [Google Scholar] [CrossRef]
- Kawano, H.; Nakatani, T.; Mori, T.; Ueno, S.; Fukaya, M.; Abe, A.; Kobayashi, M.; Toda, F.; Watanabe, M.; Matsuoka, I. Identification and Characterization of Novel Developmentally Regulated Neural-Specific Proteins, BRINP Family. Mol. Brain Res. 2004, 125, 60–75. [Google Scholar] [CrossRef]
- Ha, G.H.; Kim, J.L.; Breuer, E.K.Y. Transforming Acidic Coiled-Coil Proteins (TACCs) in Human Cancer. Cancer Lett. 2013, 336, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Mallampalli, R.K.; Li, X.; Jang, J.H.; Kaminski, T.; Hoji, A.; Coon, T.; Chandra, D.; Welty, S.; Teng, Y.; Sembrat, J.; et al. Cigarette Smoke Exposure Enhances Transforming Acidic Coiled-Coil-Containing Protein 2 Turnover and Thereby Promotes Emphysema. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Zhang, J.; Yang, X.; Wu, Z.; Sun, C.; Wang, Z.; Wang, B. NEK5 Promotes Breast Cancer Cell Proliferation through Up-Regulation of Cyclin A2. Mol. Carcinog. 2019, 58, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, P.; Nassar, J.; Peiffer, I.; Stanojcic, S.; Sterkers, Y.; Delamarre, A.; Bocquet, S.; Méchali, M. The ORC Ubiquitin Ligase OBI1 Promotes DNA Replication Origin Firing. Nat. Commun. 2019, 10, 2426. [Google Scholar] [CrossRef] [PubMed]
- Maria Fimia, G.; Stoykova, A.; Romagnoli, A.; Giunta, L.; Di Bartolomeo, S.; Nardacci, R.; Corazzari, M.; Fuoco, C.; Ucar, A.; Schwartz, P.; et al. Ambra1 Regulates Autophagy and Development of the Nervous System. Nature 2007, 447, 1121–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallos, G.; Townsend, E.; Yim, P.; Virag, L.; Zhang, Y.; Xu, D.; Bacchetta, M.; Emala, C.W. Airway Epithelium Is a Predominant Source of Endogenous Airway GABA and Contributes to Relaxation of Airway Smooth Muscle Tone. Am. J. Physiol. Cell. Mol. Physiol. 2013, 304, L191–L197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafaei, S.; Kazemnejad, A.; Azimzadeh Jamalkandi, S.; Amirhashchi, S.; Donnelly, S.C.; Armstrong, M.E.; Doroudian, M. Identification of Novel Genes in Human Airway Epithelial Cells Associated with Chronic Obstructive Pulmonary Disease (COPD) Using Machine-Based Learning Algorithms. Sci. Rep. 2018, 8, 15775. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.R.; Lai, S.Y.; Huang, L.J.; Zhang, Z.F.; Liu, J.; Zheng, S.R.; Ding, K.; Bai, X.; Zhou, J.Y. Myosin 1b Promotes Cell Proliferation, Migration, and Invasion in Cervical Cancer. Gynecol. Oncol. 2018, 149, 188–197. [Google Scholar] [CrossRef]
- Marshall, T.W.; Aloor, H.L.; Bear, J.E. Coronin 2A Regulates a Subset of Focal-Adhesion-Turnover Events through the Cofilin Pathway. J. Cell Sci. 2009, 122, 3061–3069. [Google Scholar] [CrossRef] [Green Version]
- Janco, M.; Bonello, T.T.; Byun, A.; Coster, A.C.F.; Lebhar, H.; Dedova, I.; Gunning, P.W.; Böcking, T. The Impact of Tropomyosins on Actin Filament Assembly Is Isoform Specific. Bioarchitecture 2016, 6, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, N.A.; Drescher, M.J.; Drescher, D.G. Direct Interaction of Otoferlin with Syntaxin 1A, SNAP-25, and the L-Type Voltage-Gated Calcium Channel Ca v1.3. J. Biol. Chem. 2009, 284, 1364–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Erlandsen, H.; Haavik, J.; Knappskog, P.M.; Stevens, R.C. Three-Dimensional Structure of Human Tryptophan Hydroxylase and Its Implications for the Biosynthesis of the Neurotransmitters Serotonin and Melatonin. Biochemistry 2002, 41, 12569–12574. [Google Scholar] [CrossRef] [PubMed]
- Dürk, T.; Duerschmied, D.; Müller, T.; Grimm, M.; Reuter, S.; Vieira, R.P.; Ayata, K.; Cicko, S.; Sorichter, S.; Walther, D.J.; et al. Production of Serotonin by Tryptophan Hydroxylase 1 and Release via Platelets Contribute to Allergic Airway Inflammation. Am. J. Respir. Crit. Care Med. 2013, 187, 476–485. [Google Scholar] [CrossRef] [PubMed]
- McDonald, E.R.; El-Deiry, W.S. Suppression of Caspase-8- and -10-Associated RING Proteins Results in Sensitization to Death Ligands and Inhibition of Tumor Cell Growth. Proc. Natl. Acad. Sci. USA 2004, 101, 6170–6175. [Google Scholar] [CrossRef] [Green Version]
- Davydova, E.; Ho, A.Y.Y.; Malecki, J.; Moen, A.; Enserink, J.M.; Jakobsson, M.E.; Loenarz, C.; Falnes, P. Identification and Characterization of a Novel Evolutionarily Conserved Lysine-Specific Methyltransferase Targeting Eukaryotic Translation Elongation Factor 2 (EEF2). J. Biol. Chem. 2014, 289, 30499–30510. [Google Scholar] [CrossRef] [Green Version]
- Ohya, K.I.; Kajigaya, S.; Kitanaka, A.; Yoshida, K.; Miyazato, A.; Yamashita, Y.; Yamanaka, T.; Ikeda, U.; Shimada, K.; Ozawa, K.; et al. Molecular Cloning of a Docking Protein, BRDG1, That Acts Downstream of the Tec Tyrosine Kinase. Proc. Natl. Acad. Sci. USA 1999, 96, 11976–11981. [Google Scholar] [CrossRef] [Green Version]
- Maghsoudloo, M.; Azimzadeh Jamalkandi, S.; Najafi, A.; Masoudi-Nejad, A. Identification of Biomarkers in Common Chronic Lung Diseases by Co-Expression Networks and Drug-Target Interactions Analysis. Mol. Med. 2020, 26. [Google Scholar] [CrossRef]
- Yagi, T.; Takeichi, M. Cadherin Superfamily Genes: Functions, Genomic Organization, and Neurologic Diversity. Genes Dev. 2000, 14, 1169–1180. [Google Scholar] [CrossRef]
- Wu, J.; Bond, C.; Chen, P.; Chen, M.; Li, Y.; Shohet, R.V.; Wright, G. HIF-1α in the Heart: Remodeling Nucleotide Metabolism. J. Mol. Cell. Cardiol. 2015, 82, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.-H.; Han, L.; Ouyang, X.; Kahn, A.M.; Kanellis, J.; Li, P.; Feng, L.; Nakagawa, T.; Watanabe, S.; Hosoyamada, M.; et al. Uric Acid Causes Vascular Smooth Muscle Cell Proliferation by Entering Cells via a Functional Urate Transporter. Am. J. Nephrol. 2005, 25, 425–433. [Google Scholar] [CrossRef]
- Li, S.; Sun, Z.; Zhang, Y.; Ruan, Y.; Chen, Q.; Gong, W.; Yu, J.; Xia, W.; He, J.C.J.; Huang, S.; et al. COX-2/MPGES-1/PGE2 Cascade Activation Mediates Uric Acidinduced Mesangial Cell Proliferation. Oncotarget 2017, 8, 10185–10198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshad, M.; Ye, Z.; Gu, X.; Wong, C.K.; Liu, Y.; Li, D.; Zhou, L.; Zhang, Y.; Bay, W.P.; Yu, V.C.; et al. RNF13, a RING Finger Protein, Mediates Endoplasmic Reticulum Stress-Induced Apoptosis through the Inositol-Requiring Enzyme (IRE1α)/c-Jun NH 2-Terminal Kinase Pathway. J. Biol. Chem. 2013, 288, 8726–8736. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.; Rathore, S.; Kimanius, D.; Aibara, S.; Bai, X.C.; Rorbach, J.; Amunts, A.; Ramakrishnan, V. Structures of the Human Mitochondrial Ribosome in Native States of Assembly. Nat. Struct. Mol. Biol. 2017, 24, 866–869. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Maiti, P.; Barrientos, A. Mitochondrial Ribosomes in Cancer. Semin. Cancer Biol. 2017, 47, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Monteil, A.; Chemin, J.; Bourinet, E.; Mennessier, G.; Lory, P.; Nargeot, J. Molecular and Functional Properties of the Human α(1G) Subunit That Forms T-Type Calcium Channels. J. Biol. Chem. 2000, 275, 6090–6100. [Google Scholar] [CrossRef] [Green Version]
- Nonomura, K.; Woo, S.H.; Chang, R.B.; Gillich, A.; Qiu, Z.; Francisco, A.G.; Ranade, S.S.; Liberles, S.D.; Patapoutian, A. Piezo2 Senses Airway Stretch and Mediates Lung Inflation-Induced Apnoea. Nature 2017, 541, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Okuyan, D.; Aydogan Turkoglu, S.; Kockar, F. Carbonic Anhydrase III Is a New Target of HIF1α in Prostate Cancer Model. Gene 2020, 762, 145034. [Google Scholar] [CrossRef]
- Kim, T.H.; Shin, S.W.; Park, J.S.; Park, C.S. Genome Wide Identification and Expression Profile in Epithelial Cells Exposed to TiO2 Particles. Environ. Toxicol. 2015, 30, 293–300. [Google Scholar] [CrossRef]
- Weissbach, A.; Siegesmund, K.; Brüggemann, N.; Schmidt, A.; Kasten, M.; Pichler, I.; Muhle, H.; Lohmann, E.; Lohnau, T.; Schwinger, E.; et al. Exome Sequencing in a Family with Restless Legs Syndrome. Mov. Disord. 2012, 27, 1686–1689. [Google Scholar] [CrossRef]
- Payne, D.N.R.; Rogers, A.V.; Ädelroth, E.; Bandi, V.; Guntupalli, K.K.; Bush, A.; Jeffery, P.K. Early Thickening of the Reticular Basement Membrane in Children with Difficult Asthma. Am. J. Respir. Crit. Care Med. 2003, 167, 78–82. [Google Scholar] [CrossRef]
- Saglani, S.; Payne, D.N.; Zhu, J.; Wang, Z.; Nicholson, A.G.; Bush, A.; Jeffery, P.K. Early Detection of Airway Wall Remodeling and Eosinophilic Inflammation in Preschool Wheezers. Am. J. Respir. Crit. Care Med. 2007, 176, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Van Rijt, L.S.; Vos, N.; Willart, M.; Muskens, F.; Tak, P.P.; Van Der Horst, C.; Hoogsteden, H.C.; Lambrecht, B.N. Persistent Activation of Dendritic Cells after Resolution of Allergic Airway Inflammation Breaks Tolerance to Inhaled Allergens in Mice. Am. J. Respir. Crit. Care Med. 2011, 184, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Robertson, I.B.; Horiguchi, M.; Zilberberg, L.; Dabovic, B.; Hadjiolova, K.; Rifkin, D.B. Latent TGF-β-Binding Proteins. Matrix Biol. 2015, 47, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Alpatov, R.; Munguba, G.C.; Caton, P.; Joo, J.H.; Shi, Y.; Shi, Y.; Hunt, M.E.; Sugrue, S.P. Nuclear Speckle-Associated Protein Pnn/DRS Binds to the Transcriptional Corepressor CtBP and Relieves CtBP-Mediated Repression of the E-Cadherin Gene. Mol. Cell. Biol. 2004, 24, 10223–10235. [Google Scholar] [CrossRef] [Green Version]
- Khurana, N.; Bhattacharyya, S. Hsp90, the Concertmaster: Tuning Transcription. Front. Oncol. 2015, 5, 100. [Google Scholar] [CrossRef] [Green Version]
- Han, H.J.; Tokino, T.; Nakamura, Y. CSR, a Scavenger Receptor-like Protein with a Protective Role against Cellular Damage Caused by UV Irradiation and Oxidative Stress. Hum. Mol. Genet. 1998, 7, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.H.; Liu, M.; Zhou, X.E.; Liang, G.; Zhao, G.; Xu, H.E.; Melcher, K.; Jones, P.A. Structure of Nucleosome-Bound DNA Methyltransferases DNMT3A and DNMT3B. Nature 2020, 586, 151–155. [Google Scholar] [CrossRef]
- Kurogane, Y.; Miyata, M.; Kubo, Y.; Nagamatsu, Y.; Kundu, R.K.; Uemura, A.; Ishida, T.; Quertermous, T.; Hirata, K.I.; Rikitake, Y. FGD5 Mediates Proangiogenic Action of Vascular Endothelial Growth Factor in Human Vascular Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 988–996. [Google Scholar] [CrossRef] [Green Version]
- Favre, C.; Gérard, A.; Clauzier, E.; Pontarotti, P.; Olive, D.; Nunès, J.A. DOK4 and DOK5: New Dok-Related Genes Expressed in Human T Cells. Genes Immun. 2003, 4, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Kinlough, C.L.; Poland, P.A.; Bruns, J.B.; Harkleroad, K.L.; Hughey, R.P. MUC1 Membrane Trafficking Is Modulated by Multiple Interactions. J. Biol. Chem. 2004, 279, 53071–53077. [Google Scholar] [CrossRef] [Green Version]
- Global Initiative for Asthma—Global Initiative for Asthma—GINA. Available online: https://ginasthma.org/ (accessed on 26 September 2018).
- Culver, B.H.; Graham, B.L.; Coates, A.L.; Wanger, J.; Berry, C.E.; Clarke, P.K.; Hallstrand, T.S.; Hankinson, J.L.; Kaminsky, D.A.; MacIntyre, N.R.; et al. ATS Committee on Proficiency Standards for Pulmonary Function Laboratories. Recommendations for a Standardized Pulmonary Function Report. An Official American Thoracic Society Technical Statement. Am. J. Respir. Crit. Care Med. 2017, 196, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Sokolowski, J.W.; Burgher, L.W.; Jones, F.L.; Patterson, J.R.; Selecky, P.A. Position Paper on Guidelines for Fiberoptic Bronchoscopy in Adults. Am. Rev. Respir. Dis. 1987, 136, 1066. [Google Scholar] [CrossRef]
- Kozlik, P.; Zuk, J.; Bartyzel, S.; Zarychta, J.; Okon, K.; Zareba, L.; Bazan, J.G.; Kosalka, J.; Soja, J.; Musial, J.; et al. The Relationship of Airway Structural Changes to Blood and Bronchoalveolar Lavage Biomarkers, and Lung Function Abnormalities in Asthma. Clin. Exp. Allergy 2020, 50, 15–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrando, R.E.; Nyengaard, J.R.; Hays, S.R.; Fahy, J.V.; Woodruff, P.G. Applying Stereology to Measure Thickness of the Basement Membrane Zone in Bronchial Biopsy Specimens. J. Allergy Clin. Immunol. 2003, 112, 1243–1245. [Google Scholar] [CrossRef] [PubMed]
- Matas, J.; Chum, O.; Urban, M.; Pajdla, T. Robust Wide Baseline Stereo from Maximally Stable Extremal Regions. Proc. Br. Mach. Vis. Conf. 2002, 22, 384–396. [Google Scholar]
- Tsai, M.Y.; Chen, T.B.; Lu, H.H.S. Microarray Image and Data Analysis; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No-Persistent Airflow Limitation, n = 19 | Persistent Airflow Limitation, n = 21 | p-Value | |
|---|---|---|---|
| Demographic data | |||
| Age, years | 52 (42–63) | 56 (53–68) | 0.07 |
| Male gender, n (%) | 5 (26) | 3 (14) | 0.58 |
| Height, m | 1.62 ± 0.08 | 1.63 ± 0.09 | 0.8 |
| Body mass index, kg/m2 | 28.2 (24.9–31.3) | 25.7 (23.9–31.2) | 0.26 |
| Past smoking, n (%) | 5 (26) | 5 (24) | 0.86 |
| Pack-years of smoking, n | 0 (0–0) | 0 (0–0) | 0.46 |
| Living primarily in inner-city environments, n (%) | 9 (47) | 12 (57) | 0.76 |
| Asthma-Related Variables | |||
| Atopy, n (%) | 11 (58) | 9 (43) | 0.95 |
| Asthma duration, years | 11 (5–20) | 8.5 (5.5–18) | 0.75 |
| Asthma severity (GINA): | |||
| persistent moderate, n (%) | 8 (42) | 7 (33) | |
| persistent severe, n (%) | 11 (58) | 14 (67) | 0.57 |
| Asthma symptom control §: | |||
| well-controlled asthma, n (%) | 2 (11) | 2 (10) | |
| not-well controlled asthma, n (%) | 4 (21) | 5 (24) | |
| very poorly controlled asthma, n (%) | 13 (68) | 14 (67) | 0.97 |
| Oral glucocorticosteroids use, n (%) | 5 (26) | 5 (24) | 0.86 |
| FEV1 before bronchodilator, L | 2.58 ± 0.84 | 1.67 ± 0.8 | <0.001 |
| FEV1 before bronchodilator, % of the predicted value | 101 (85.9–110) | 68.05 (48.3–79) | <0.001 |
| FEV1 after bronchodilator, L | 2.61 ± 0.81 | 1.92 ± 0.89 | 0.02 |
| FEV1 after bronchodilator, % of the predicted value | 103.8 (93–114.5) | 73.7 (61.6–88.2) | <0.001 |
| Change in FEV1 after bronchodilator, L | 0.16 (0.03–0.29) | 0.23 (0.15–0.34) | 0.23 |
| FEV1/VC (before bronchodilator) | 74.1 (71.6–76.3) | 56.7 (49–61.1) | <0.001 |
| FEV1/VC (after bronchodilator) | 77.8 (72–79.7) | 62.3 (54.7–67.8) | <0.001 |
| No-Persistent Airflow Limitation, n = 19 | Persistent Airflow Limitation, n = 21 | p-Value | |
|---|---|---|---|
| Blood Count | |||
| Hemoglobin, g/dL | 13.4 (12.9–13.9) | 13.7 (12.6–14.4) | 0.67 |
| Red blood cells, 106/µL | 4.62 (4.3–4.84) | 4.71 (4.17–5.16) | 0.71 |
| White blood cells, 103/µL | 5.88 (4.95–7.51) | 7.45 (6.51–9.25) | 0.008 |
| Neutrophils, 103/µL | 3.5 (2.7–4.2) | 3.54 (2.7–4.2) | 0.73 |
| Lymphocytes, 103/µL | 1.9 (1.5–2.4) | 2.2 (1.97–2.6) | 0.13 |
| Monocytes, 103/µL | 0.51 (0.48–0.68) | 0.68 (0.53–1.1) | 0.02 |
| Blood platelets, 103/µL | 218 (177–246) | 224 (187–294) | 0.26 |
| Asthma and Inflammatory Biomarkers in Peripheral Blood | |||
| Eosinophilia/µL | 130 (70–290) | 565 (380–1340) | <0.001 |
| Immunoglobulin E, IU/mL | 43 (22.7–133) | 152 (32–586) | 0.19 |
| C-reactive protein, mg/L | 4.24 (0.45–8.67) | 6.71 (0.58–42) | 0.9 |
| Interleukin 6, pg/mL | 0.58 (0.43–1.04) | 1.12 (0.5–3.21) | 0.08 |
| Interleukin 10, pg/mL | 0.44 (0.005–0.76) | 0.67 (0.41–1.62) | 0.05 |
| Periostin, ng/mL | 0.3 (0.27–0.36) | 0.38 (0.31–0.51) | 0.06 |
| A disintegrin and metalloproteinase domain-containing protein 33, ng/mL | 1.2 (0.2–2.29) | 1.76 (1.26–2.77) | 0.11 |
| Reticular Basement Membrane Thickness | |||
| The thickness of the reticular basement membrane of the bronchial mucosa, µm | 5.96 (5–7.96) | 6.2 (5.24–7.45) | 0.9 |
| Bronchoalveolar Lavage Cellularity and Biomarkers | |||
| Macrophages, % | 86.8 (74.5–93) | 83 (56.3–91) | 0.47 |
| Lymphocytes, % | 7 (4.5–15.5) | 7 (3–10.5) | 0.33 |
| Neutrophils, % | 3 (2–4.5) | 4 (2–12.5) | 0.34 |
| Eosinophils, % | 0.1 (0–1) | 2.3 (0.5-7) | 0.004 |
| Periostin, ng/mL | 0.85 (0.76–0.96) | 0.81 (0.72–0.95) | 0.6 |
| Interleukin 6, pg/ml | 0.99 (0.67–1.14) | 0.2 (0.005–0.86) | 0.01 |
| Interleukin 10, pg/ml | 0.01 (0.005–0.01) | 0.01 (0.005–0.01) | 0.81 |
| No-Persistent Airflow Limitation, n = 19 | Persistent Airflow Limitation, n = 21 | p-Value | p-Value (Adjusted) | |
|---|---|---|---|---|
| The Right Upper Lobe Apical Segmental Bronchus (RB1) | ||||
| Lumen diameter, mm | 3.95 (3.6–4.5) | 4.05 (3.7–5.3) | 0.58 | 0.23 |
| Airway diameter, mm | 7.4 (6.8–8.4) | 8.3 (7.3–9.3) | 0.13 | 0.76 |
| Wall thickness, mm | 1.8 (1.6–1.9) | 1.95 (1.8–2.4) | 0.02 | 0.16 |
| Wall thickness ratio | 23.75 (21.3–24.6) | 24.75 (23.1–26.9) | 0.07 | 0.01 |
| Lumen area, mm2 | 12.3 (10–16) | 12.8 (10.5–22) | 0.65 | 0.2 |
| Wall area, mm2 | 30.8 (25.9–39.3) | 39.4 (31.1–48.1) | 0.1 | 0.78 |
| Wall area ratio | 72.55 (67.3–75.1) | 74.8 (71.3–78.7) | 0.08 | 0.01 |
| The Right Lower Lobe Basal Posterior Bronchus (RB10) | ||||
| Lumen diameter, mm | 4.35 (3.7–4.7) | 3.8 (3–4.5) | 0.17 | 0.02 |
| Airway diameter, mm | 7.9 (6.3–8.5) | 7.6 (6.2–8.4) | 0.36 | 0.03 |
| Wall thickness, mm | 1.8 (1.6–1.9) | 1.8 (1.5–1.9) | 0.98 | 0.54 |
| Wall thickness ratio | 23.1 (19.8–24.9) | 24 (22–26.4) | 0.16 | 0.07 |
| Lumen area, mm2 | 14 (8.3–17) | 11.1 (7.3–16.2) | 0.31 | 0.04 |
| Wall area, mm2 | 33.6 (22.4–42.4) | 31.7 (21.9–40.5 | 0.5 | 0.08 |
| Wall area ratio | 71.3 (63.6–74.8) | 73.3 (68.5–78.5) | 0.18 | 0.06 |
| Gene | Alias for Gene | Correlation Coefficient | Potential Linkage with Airway and Asthma Pathology |
|---|---|---|---|
| MIS12 | MIS12 kinetochore complex component | 0.745 | Plays an essential role in chromosome segregation [9]. The higher expression suggests increased proliferation potency of airway epithelial cells. |
| CDK20 | Cyclin-dependent kinase 20 | 0.727 | The encoded kinase may activate airway epithelial cell growth and proliferation [8]. |
| TACC2 | Transforming acidic coiled-coil protein 2 | 0.608 | A centrosome- and microtubule-interacting protein, involved in homologous recombination repair [22], attenuates DNA damage and cytotoxicity in immortalized human bronchial epithelial cells [23]. |
| NEK5 | NIMA related kinase 5 | 0.625 | Regulates centrosome integrity and is involved in cell proliferation [24]. The higher expression suggests increased proliferation potency of airway epithelial cells. |
| ORC5 | The origin recognition complex subunit 5 | 0.603 | Promotes DNA replication [25]. The higher expression might indicate increased proliferation potency of airway epithelial cells. |
| AMBRA1 | Autophagy and beclin 1 regulator 1 | 0.626 | Regulates autophagy, cell survival, and proliferation [26]. A positive correlation with reticular basement membrane (RBM) thickness suggests that autophagy mechanisms are likely involved in airway remodeling pathology. |
| GAD1 | Glutamate decarboxylase 1 | 0.696 | An enzyme catalyzing gamma-aminobutyric acid (GABA) production. GABA may be produced by human epithelial cells, likely relaxing airway smooth muscle [27]. Higher expression of GAD1 in airway transcriptome has also been documented in chronic obstructive pulmonary disease (COPD) patients [28]. |
| MYO1B | Myosin I B | 0.658 | A motor protein related to the actin filament-based cell organization and movement [29]. Higher airway epithelial cell expression might reflect epithelial-to-mesenchymal transition, a structural modification, whereby epithelial cells lose cell–cell polarity and display mesenchymal features, such as migratory properties [5]. |
| CORO2A | Coronin 2A | 0.6 | An actin-binding protein involved in cell cycle progression, signal transduction, cell motility, apoptosis, and gene expression [30]. Higher expression in airway epithelium may also reflect the epithelial-to-mesenchymal transition. |
| TPM1 | Tropomyosin alpha-1 chain | 0.603 | An actin-binding protein involved in the contractile cell system [31]. The higher expression may also reflect the epithelial-to-mesenchymal transition. |
| OTOF | Otoferlin | 0.612 | A protein involved in the vesicular release. Pathological mutations in OTOF are related to deafness. Although its exact role in asthma pathology is unknown, it may be involved in airway mucus secretion [32]. |
| TPH1 | Tryptophan hydrolase 1 | 0.643 | A hydroxylase that catalyzes the first rate-limiting step in the biosynthesis of serotonin [33], an important inflammatory mediator in asthma. Serotonin modulates the function of various inflammatory cells, such as mast cells, eosinophils, dendritic cells, and lung epithelium. Higher levels have been shown in asthma patients’ airways; however, its exact source in the lungs is unknown [34]. |
| RNF34 | Ring finger protein 34 | 0.683 | A ring finger containing protein, involved in protein–protein and protein–DNA interactions. It likely protects epithelial cells from premature apoptosis [35]. |
| EEF2 | Eukaryotic translation elongation factor 2 | 0.658 | An essential factor in cell protein biosynthesis. The higher expression suggests increased protein translation potency in asthma patients with thicker RBM [36]. |
| STAP1 | Signal transducing adaptor family member 1 | 0.609 | Positively regulates gene expression and signal transduction. The exact role in asthma pathology is unknown; however, it is abundant in activated human B cells [37]. |
| OAS2 | 2′-5′-oligoadenylate synthetase 2 | 0.703 | An interferon-induced antiviral ribonuclease L that destabilizes double-stranded viral RNA. It plays a critical role in the cellular innate antiviral response. A positive correlation with RBM thickness suggests that viral infections, essential asthma exacerbation triggers, might also be involved in airway remodeling pathology. The encoded protein also regulates cell apoptosis, growth, differentiation, and gene expression [20]. |
| USH2A | Usherin | 0.601 | An extracellular matrix protein, found in many tissues’ basement membrane, including the lungs [18]. |
| LAMA3 | Laminin subunit α3 | 0.615 | A protein belonging to the laminin family. Laminins are essential for the formation and function of the basement membrane [17]. Higher airway expression has been linked with COPD and idiopathic pulmonary fibrosis patients [38]. |
| PCDHA3 | Protocadherin alpha 3 | 0.609 | A neural cadherin-like cell adhesion protein. Its role in airways is unknown; however, it might be related to the neural network and synaptic organization in airway remodeling pathology [39]. |
| PDS5 | Cohesin associated factor B | −0.643 | A protein that negatively regulates cell proliferation [10]. The lower expression suggests increased proliferation potency of airway epithelial cells. |
| HPRT1 | Hypoxanthine phosphoribosyltransferase 1 | −0.647 | A transferase engaged in purine salvage pathway. Its deficiency might be associated with increased local uric acid production [40]. Soluble uric acid is an endogenous danger signal, which may stimulate neovascularization [41] and cell proliferation [42]. Furthermore, hypoxia-inducible factor 1 α (HIF1-α) strongly upregulates HPRT1; therefore, a lower expression of HPRT1 might indicate a worse response to oxidative stress in airway epithelium [40]. |
| RNF13 | Ring finger protein 13 | −0.636 | A ring zinc finger containing protein, promoting cell apoptosis [43]. The lower expression might indicate the anti-apoptotic potency of airway epithelial cells. |
| MRPL37 | Mitochondrial ribosomal protein L37 | −0.66 | A large subunit of mitochondrial ribosomal proteins, which catalyzes protein production within the mitochondrion. Recent investigations have uncovered this protein’s pro-apoptotic role [44,45]; therefore, its lower expression might indicate anti-apoptotic potency of airway epithelial cells. |
| CACNA1G | Calcium voltage-gated channel subunit α1 G | −0.615 | A transmembrane voltage-dependent calcium channel promoting cell death. The decreased expression suggests lower apoptotic activity [46]. |
| PIEZO2 | Piezo type mechanosensitive ion channel component 2 | −0.6 | A stretch-gated ion channel that senses airway stretches [47]. Lower expression in those with a thicker RBM might indicate a weaker response to epithelial cells’ mechanical stimuli. Lower PIEZO2 expression has been shown in COPD patients [28]. |
| CA3 | Carbonic anhydrase 3 | −0.606 | A carbonic anhydrase that facilitates the reversible hydrating of carbon dioxide. Lower expression in those with thicker RBM might indicate a possible weaker contraction ability or a worse response to oxidative stress in airway epithelium because CA3 is upregulated by HIF1-α [48]. |
| KIF11 | Kinesin family member 11 | −0.645 | A motor cell protein involved i.a. in cilia beating and cell division. The lower expression might suggest impaired cilia function in those with a thicker RBM [49]. |
| C21orf58 | Chromosome 21 open reading frame 58 | −0.629 | Exact biological roles are unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazan-Socha, S.; Buregwa-Czuma, S.; Jakiela, B.; Zareba, L.; Zawlik, I.; Myszka, A.; Soja, J.; Okon, K.; Zarychta, J.; Kozlik, P.; et al. Reticular Basement Membrane Thickness Is Associated with Growth- and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients. Int. J. Mol. Sci. 2021, 22, 998. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22030998
Bazan-Socha S, Buregwa-Czuma S, Jakiela B, Zareba L, Zawlik I, Myszka A, Soja J, Okon K, Zarychta J, Kozlik P, et al. Reticular Basement Membrane Thickness Is Associated with Growth- and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients. International Journal of Molecular Sciences. 2021; 22(3):998. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22030998
Chicago/Turabian StyleBazan-Socha, Stanislawa, Sylwia Buregwa-Czuma, Bogdan Jakiela, Lech Zareba, Izabela Zawlik, Aleksander Myszka, Jerzy Soja, Krzysztof Okon, Jacek Zarychta, Paweł Kozlik, and et al. 2021. "Reticular Basement Membrane Thickness Is Associated with Growth- and Fibrosis-Promoting Airway Transcriptome Profile-Study in Asthma Patients" International Journal of Molecular Sciences 22, no. 3: 998. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22030998