
Multiple Na,K-ATPase Subunits Colocalize in the Brush Border of Mouse Choroid Plexus Epithelial Cells

 ,
, 
Abstract
:1. Introduction
2. Results
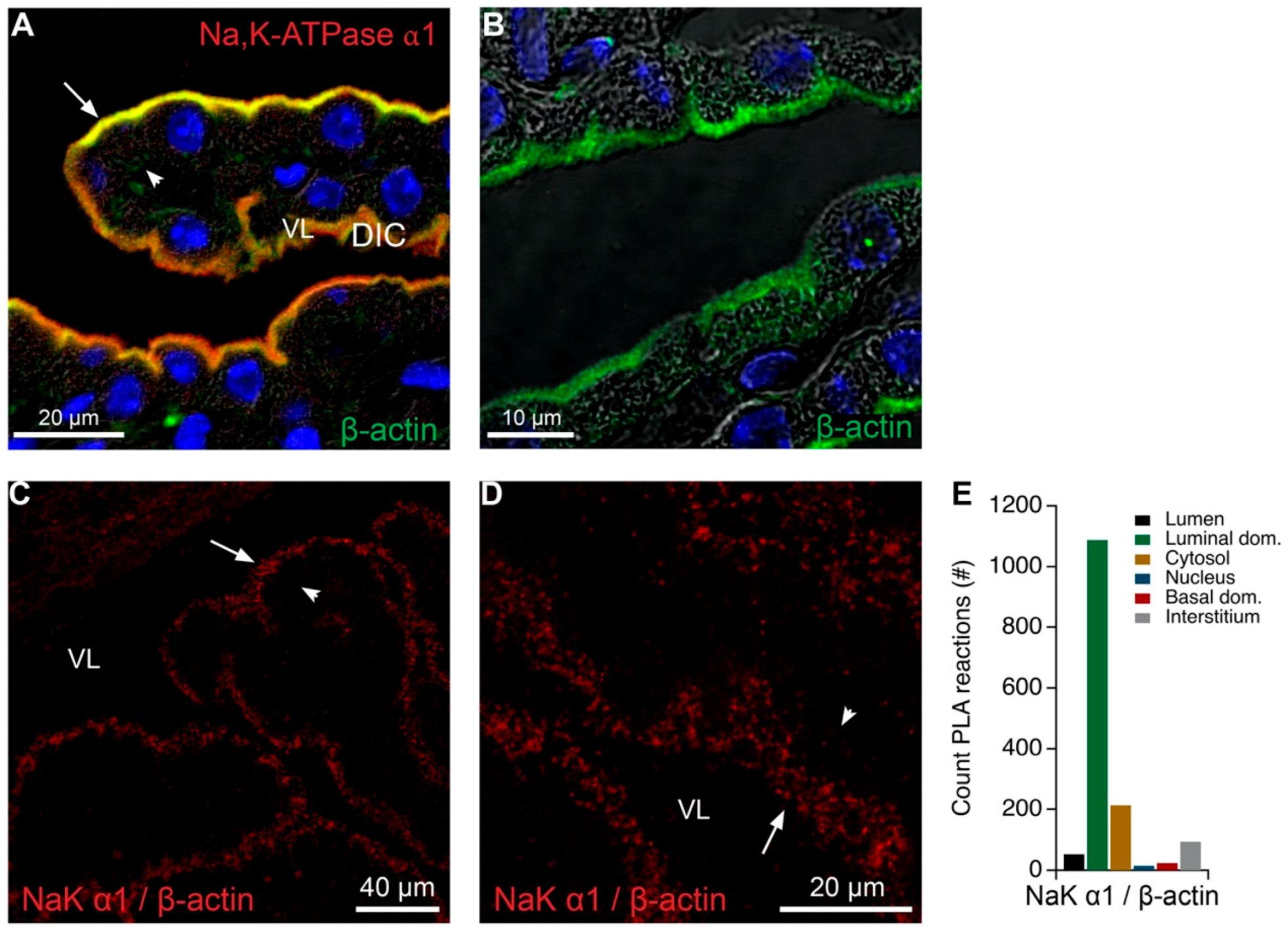
2.1. The CP Expresses Several Na,K-ATPase Subunits in the Brush Border
2.2. Na,K-ATPase α1 Immunoprecipitates Both Luminal and Basolateral Transport Proteins
2.3. Na,K-ATPases and AQP1 Exist in the Same Subcellular Fractions
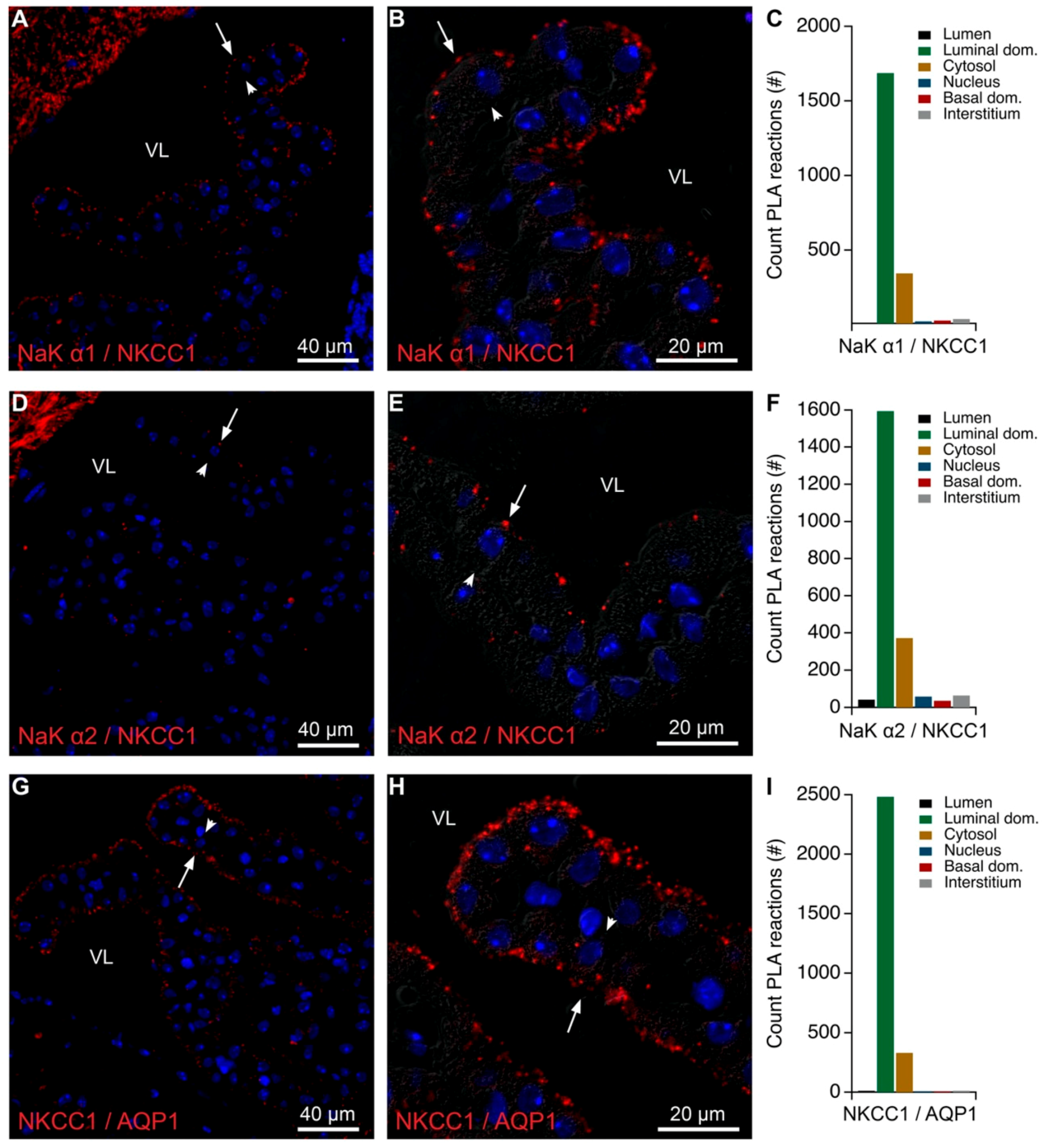
2.4. NKCC1, Ncbe, and AE2 Belong to the Same Subcellular Fractions
2.5. Intracellular Proximity of a Subset of Na,K-ATPase Subunits with AE2 and Ncbe
2.6. Brush Border Proximity of Na,K-ATPase α1 Subunits and Ankyrin-3 in the CP
2.7. Anchoring of the Brush Border Na,K-ATPase α1 Subunits May Not Involve the Spectrin Network
2.8. Partial Basolateral Accumulation of NaK-ATPase and AQP1 in NKCC1 ko CP
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Fluorescence-Activated Cell Sorting (FACS) of CPECs
4.3. RNA Purification and Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
4.4. Tissue Fixation and Immunohistochemistry (IHC)
4.5. Proximity Ligation Assay
4.6. Stimulated Emission Depletion (STED) Microscopy
4.7. Image Analysis
4.8. Co-Immunoprecipitation
4.9. Separation of Subcellular Elements by Differential Centrifugation
4.10. Sodium Dodecyl Sulphate (SDS) and Native Polyacrylamide Gel Electrophoresis (PAGE)
4.11. Immunoblotting
4.12. Mass Spectrometry Analysis
4.13. MS Data Analysis
4.14. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AE2 | Anion exchanger 2 |
| AQP1 | Aquaporin 1 |
| BSA | Bovine serum albumin |
| CP | Choroid plexus |
| CPE | Choroid plexus epithelium |
| CPEC | Choroid plexus epithelial cells |
| CSF | Cerebrospinal fluid |
| EDTA | Ethylenediaminetetraacetic acid |
| EGTA | (Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| FACS | Fluorescence-activated cell sorting |
| FXYD1 | Phospholemman |
| HEPES | 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid |
| HBS | Hepes buffered salt solution |
| IB | Immunoblotting |
| IHC | Immunohistochemistry |
| Kir 7.1 | Inward rectifier potassium channel 13 |
| NBCe2 | Electrogenic Na-bicarbonate cotransporter 2 |
| NBCn1 | Electroneutral Na-bicarbonate cotransporter 1 |
| Ncbe | Na-dependent chloride-bicarbonate exchanger |
| NHE1 | Na-hydrogen exchanger |
| NKCC1 | Na-K-2Cl cotransporter 1 |
| PAGE | Polyacrylamide gel electrophoresis |
| PBS | Phosphate buffered salt solution |
| PLA | Proximity ligation assay |
| RT-PCR | Reverse transcriptase polymerase chain reaction |
| STED | Stimulated emission depletion |
| TRPv4 | Transient receptor potential cation channel V4 |
References
- Damkier, H.H.; Brown, P.D.; Praetorius, J. Cerebrospinal Fluid Secretion by the Choroid Plexus. Physiol. Rev. 2013, 93, 1847–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, D.S.; Pease, D.C. The electron microscopy of the choroid plexus. J. Biophys. Biochem. Cytol. 1956, 2, 467–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cereijido, M.; Contreras, R.G.; Shoshani, L. Cell Adhesion, Polarity, and Epithelia in the Dawn of Metazoans. Physiol. Rev. 2004, 84, 1229–1262. [Google Scholar] [CrossRef]
- Zlokovic, B.V.; Mackic, J.B.; Wang, L.; McComb, J.G.; McDonough, A. Differential expression of Na,K-ATPase alpha and beta subunit isoforms at the blood-brain barrier and the choroid plexus. J. Biol. Chem. 1993, 268, 8019–8025. [Google Scholar] [CrossRef]
- Christensen, I.B.; Wu, Q.; Bohlbro, A.S.; Skals, M.G.; Damkier, H.H.; Hübner, C.A.; Fenton, R.A.; Praetorius, J. Genetic disruption of slc4a10 alters the capacity for cellular metabolism and vectorial ion transport in the choroid plexus epithelium. Fluids Barriers CNS 2020, 17, 2–18. [Google Scholar] [CrossRef]
- Damkier, H.H.; Christensen, H.L.; Christensen, I.B.; Wu, Q.; Fenton, R.A.; Praetorius, J. The murine choroid plexus epithelium expresses the 2Cl−/H+ exchanger ClC-7 and Na+/H+ exchanger NHE6 in the luminal membrane domain. Am. J. Physiol. Cell Physiol. 2018, 314, C439–C448. [Google Scholar] [CrossRef] [Green Version]
- Feschenko, M.S.; Donnet, C.; Wetzel, R.K.; Asinovski, N.K.; Jones, L.R.; Sweadner, K.J. Phospholemman, a Single-Span Membrane Protein, Is an Accessory Protein of Na,K-ATPase in Cerebellum and Choroid Plexus. J. Neurosci. 2003, 23, 2161–2169. [Google Scholar] [CrossRef]
- Gonzalez-Martinez, L.M.; Avila, J.; Marti, E.; Lecuona, E.; Martin-Vasallo, P. Expression of the beta-subunit isoforms of the Na,K-ATPase in rat embryo tissues, inner ear and choroid plexus. Biol. Cell 1994, 81, 215–222. [Google Scholar] [CrossRef]
- Watts, A.G.; Sanchez-Watts, G.; Emanuel, J.R.; Levenson, R. Cell-specific expression of mRNAs encoding Na+,K+-ATPase alpha- and beta-subunit isoforms within the rat central nervous system. Proc. Natl. Acad. Sci. USA 1991, 88, 7425–7429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagin, O.; Turdikulova, S.; Sachs, G. Recombinant addition of N-glycosylation sites to the basolateral Na,K-ATPase beta1 subunit results in its clustering in caveolae and apical sorting in HGT-1 cells. J. Biol. Chem. 2005, 280, 43159–43167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagin, O.; Tokhtaeva, E.; Yakubov, I.; Shevchenko, E.; Sachs, G. Inverse Correlation between the Extent of N-Glycan Branching and Intercellular Adhesion in Epithelia. J. Biol. Chem. 2008, 283, 2192–2202. [Google Scholar] [CrossRef] [Green Version]
- Morrow, J.S.; Cianci, C.D.; Ardito, T.; Mann, A.S.; Kashgarian, M. Ankyrin links fodrin to the alpha subunit of Na,K-ATPase in Madin-Darby canine kidney cells and in intact renal tubule cells. J. Cell Biol. 1989, 108, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.J.; Hammerton, R.W. A membrane-cytoskeletal complex containing Na+,K+-ATPase, ankyrin, and fodrin in Ma-din-Darby canine kidney (MDCK) cells: Implications for the biogenesis of epithelial cell polarity. J. Cell Biol. 1989, 108, 893–902. [Google Scholar] [CrossRef]
- Alper, S.L.; Stuart-Tilley, A.; Simmons, C.F.; Brown, D.; Drenckhahn, D. The fodrin-ankyrin cytoskeleton of choroid plexus preferentially colocalizes with apical Na+,K+-ATPase rather than with basolateral anion exchanger AE2. J. Clin. Investig. 1994, 93, 1430–1438. [Google Scholar] [CrossRef] [Green Version]
- Christensen, I.B.; Gyldenholm, T.; Damkier, H.; Praetorius, J. Polarization of membrane associated proteins in the choroid plexus epithelium from normal and slc4a10 knockout mice. Front. Physiol. 2013, 4, 344. [Google Scholar] [CrossRef] [Green Version]
- Marrs, J.A.; Napolitano, E.W.; Murphy-Erdosh, C.; Mays, R.W.; Reichardt, L.F.; Nelson, W.J. Distinguishing roles of the mem-brane-cytoskeleton and cadherin mediated cell-cell adhesion in generating different Na+,K+-ATPase distributions in po-larized epithelia. J. Cell Biol. 1993, 123, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Masuzawa, T.; Ohta, T.; Kawamura, M.; Nakahara, N.; Sato, F. Immunohistochemical localization of Na+,K+-ATPase in the choroid plexus. Brain Res. 1984, 302, 357–362. [Google Scholar] [CrossRef]
- Zeuthen, T.; Wright, E.M. An electrogenic Na+/K+ pump in the choroid plexus. Biochim. Biophys. Acta 1978, 511, 517–522. [Google Scholar] [CrossRef]
- Crambert, G.; Füzesi, M.; Garty, H.; Karlish, S.; Geering, K. Phospholemman (FXYD1) associates with Na,K-ATPase and regulates its transport properties. Proc. Natl. Acad. Sci. USA 2002, 99, 11476–11481. [Google Scholar] [CrossRef] [Green Version]
- Ernst, S.A.; Palacios, J.R.; Siegel, G.J. Immunocytochemical localization of Na+,K+-ATPase catalytic polypeptide in mouse choroid plexus. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1986, 34, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, G.J.; Holm, C.; Schreiber, J.H.; Desmond, T.; Ernst, S.A. Purification of mouse brain (Na+ + K+)-ATPase catalytic unit, characterization of antiserum, and immunocytochemical localization in cerebellum, choroid plexus, and kidney. J. Histochem. Cytochem. 1984, 32, 1309–1318. [Google Scholar] [CrossRef]
- Barwe, S.P.; Anilkumar, G.; Moon, S.Y.; Zheng, Y.; Whitelegge, J.P.; Rajasekaran, S.A.; Rajasekaran, A.K. Novel Role for Na,K-ATPase in Phosphatidylinositol 3-Kinase Signaling and Suppression of Cell Motility. Mol. Biol. Cell 2005, 16, 1082–1094. [Google Scholar] [CrossRef] [PubMed]
- Brignone, M.S.; Lanciotti, A.; Macioce, P.; Macchia, G.; Gaetani, M.; Aloisi, F.; Petrucci, T.C.; Ambrosini, E. The β1 subunit of the Na,K-ATPase pump interacts with megalencephalic leucoencephalopathy with subcortical cysts protein 1 (MLC1) in brain astrocytes: New insights into MLC pathogenesis. Hum. Mol. Genet. 2010, 20, 90–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damkier, H.H.; Prasad, V.; Hubner, C.A.; Praetorius, J. Nhe1 is a luminal Na+/H+ exchanger in mouse choroid plexus and is targeted to the basolateral membrane in Ncbe/Nbcn2-null mice. Am. J. Physiol. Cell Physiol. 2009, 296, C1291–C1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damkier, H.; Praetorius, J. Genetic ablation of Slc4a10 alters the expression pattern of transporters involved in solute movement in the mouse choroid plexus. Am. J. Physiol. Cell Physiol. 2012, 302, C1452–C1459. [Google Scholar] [CrossRef] [Green Version]
- Christensen, H.L.; Barbuskaite, D.; Rojek, A.; Malte, H.; Christensen, I.B.; Füchtbauer, A.C.; Füchtbauer, E.-M.; Wang, T.; Praetorius, J.; Damkier, H. The choroid plexus sodium-bicarbonate cotransporter NBCe2 regulates mouse cerebrospinal fluid pH. J. Physiol. 2018, 596, 4709–4728. [Google Scholar] [CrossRef] [Green Version]
- Flagella, M.; Clarke, L.L.; Miller, M.L.; Erway, L.C.; Giannella, R.A.; Andringa, A.; Gawenis, L.R.; Kramer, J.; Duffy, J.J.; Doetschman, T.; et al. Mice Lacking the Basolateral Na-K-2Cl Cotransporter Have Impaired Epithelial Chloride Secretion and Are Profoundly Deaf. J. Biol. Chem. 1999, 274, 26946–26955. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Yang, B.; Gillespie, A.; Carlson, E.J.; Epstein, C.J.; Verkman, A. Severely Impaired Urinary Concentrating Ability in Transgenic Mice Lacking Aquaporin-1 Water Channels. J. Biol. Chem. 1998, 273, 4296–4299. [Google Scholar] [CrossRef] [Green Version]
- Boedtkjer, E.; Praetorius, J.; Matchkov, V.V.; Stankevicius, E.; Mogensen, S.; Füchtbauer, A.C.; Simonsen, U.; Füchtbauer, E.-M.; Aalkjaer, C. Disruption of Na+,HCO3− Cotransporter NBCn1 (slc4a7) Inhibits NO-Mediated Vasorelaxation, Smooth Muscle Ca2+ Sensitivity, and Hypertension Development in Mice. Circulation 2011, 124, 1819–1829. [Google Scholar] [CrossRef] [Green Version]
- Weber, P.; Bartsch, U.; Schachner, M.; Montag, D. Na,K-ATPase subunit beta1 knock-in prevents lethality of beta2 deficiency in mice. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 9192–9203. [Google Scholar] [CrossRef] [Green Version]
- Kashgarian, M.; Biemesderfer, D.; Caplan, M.; Forbush, B., 3rd. Monoclonal antibody to Na,K-ATPase: Immunocytochemical lo-calization along nephron segments. Kidney Int. 1985, 28, 899–913. [Google Scholar] [CrossRef] [Green Version]
- Kurihara, K.; Moore-Hoon, M.L.; Saitoh, M.; Turner, R.J. Characterization of a phosphorylation event resulting in upregulation of the salivary Na+-K+-2Cl− cotransporter. Am. J. Physiol. 1999, 277, C1184–C1193. [Google Scholar] [CrossRef] [PubMed]
- Sykaras, A.G.; Demenis, C.; Cheng, L.; Pisitkun, T.; McLaughlin, J.T.; Fenton, R.A.; Smith, C.P. Duodenal CCK Cells from Male Mice Express Multiple Hormones Including Ghrelin. Endocrinology 2014, 155, 3339–3351. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Description | Abbreviation | Max. Cover. | Uniq. Pept. | n |
|---|---|---|---|---|---|
| 21450277 | Sodium/potassium-transp. ATPase a-1 | Na,K-ATPase α1 | 25.71 | 18 | 5 |
| 6753138 | Sodium/potassium-transp. ATPase b-1 | Na,K-ATPase β1 | 20.72 | 5 | 5 |
| 569003445 | Solute carrier family 12 member 2 | NKCC1 | 19.66 | 17 | 2 |
| 22165384 | Tubulin beta-4B chain | β-tubulin | 17.08 | 3 | 1 |
| 6671509 | Actin, cytoplasmic 1 | β-actin | 14.40 | 4 | 3 |
| 34996479 | Phospholemman isoform b | FXYD1 | 13.04 | 1 | 1 |
| 6678469 | Tubulin alpha-1C chain | α-tubulin | 11.80 | 3 | 3 |
| 30409956 | Sodium/potassium-transp. ATPase a-2 | Na,K-ATPase α2 | 11.18 | 2 | 1 |
| 568960685 | Sodium/potassium-transp. ATPase b-3 | Na,K-ATPase β3 | 10.74 | 2 | 1 |
| 7106439 | Tubulin beta-5 chain | β-tubulin | 9.91 | 1 | 2 |
| 226958351 | Sodium/potassium-transp. ATPase a-4 | Na,K-ATPase α4 | 8.72 | 1 | 1 |
| 6680710 | Aquaporin-1 | AQP1 | 7.06 | 2 | 4 |
| 6996913 | Annexin A2 | Annexin-2 | 5.31 | 1 | 2 |
| 7242138 | Sodium/potassium-transp. ATPase b-2 | Na,K-ATPase β2 | 4.14 | 1 | 1 |
| 568933073 | Anion exchange protein 2 | AE2 | 3.76 | 3 | 1 |
| 568971845 | Junction plakoglobin | γ-catenin | 3.57 | 2 | 2 |
| 7710096 | Beta-1-syntrophin | β1-syntrophin | 3.17 | 1 | 1 |
| 6678059 | Beta-2-syntrophin | β2-syntrophin | 2.50 | 1 | 1 |
| 158966733 | Inward rectifier potassium channel 13 | Kir7.1 | 2.50 | 1 | 1 |
| 295054271 | Spectrin alpha chain, non-erythroc. 1 | α2-spectrin | 2.12 | 4 | 1 |
| 568978640 | Alpha-actinin-1 | α-actinin | 1.79 | 1 | 1 |
| 334688858 | Sodium-driven chloride bicarb. exch. | Ncbe | 1.66 | 1 | 1 |
| Accession | Description | Abbreviation | Max. Coverage | Uniq. Pept. | n |
|---|---|---|---|---|---|
| 6671509 | Actin, cytoplasmic 1 | β-actin | 33.33 | 4 | 1 |
| 6678469 | Tubulin alpha-1C chain | α-tubulin | 29.84 | 3 | 2 |
| 568907654 | Tubulin alpha-4A chain | α-tubulin | 26.10 | 2 | 1 |
| 22165384 | Tubulin beta-4B chain | β-tubulin | 27.19 | 11 | 1 |
| 6680710 | Aquaporin-1 | AQP1 | 17.84 | 5 | 2 |
| 158966733 | Inward rectifier potassium channel 13 | Kir7.1 | 8.61 | 3 | 1 |
| 21450277 | Sodium/potassium-transp. ATPase a-1 | Na,K-ATPase α1 | 54.06 | 39 | 2 |
| 30409956 | Sodium/potassium-transp. ATPase a-2 | Na,K-ATPase α2 | 48.92 | 24 | 2 |
| 594190942 | Sodium/potassium-transp. ATPase a-3 | Na,K-ATPase α3 | 43.83 | 19 | 1 |
| 226958351 | Sodium/potassium-transp. ATPase a-4 | Na,K-ATPase α4 | 13.57 | 3 | 1 |
| 6753138 | Sodium/potassium-transp. ATPase b-1 | Na,K-ATPase β1 | 38.82 | 15 | 2 |
| 7242138 | Sodium/potassium-transp. ATPase b-2 | Na,K-ATPase β2 | 16.21 | 5 | 2 |
| 568960685 | Sodium/potassium-transp. ATPase b-3 | Na,K-ATPase β3 | 21.90 | 5 | 1 |
| 37577140 | Phospholemman | FXYD1 | 17.39 | 2 | 2 |
| Parameter | NKA α1 AQP1 | NKA β1 NKA β1 | NKA α1 NKA α2 | NKA α1 NKA β1 | NKA β1 α2-Spec |
|---|---|---|---|---|---|
| % colocalized material A thresh. | 79.46 | 41.87 | 56.1525 | 31.31 | 30 |
| % colocalized material B thresh. | 39.81 | 50.34 | 72.5 | 45.47 | 33.93 |
| Pearson’s coefficient in volume | 0.659 | 0.591 | 0.79953 | 0.575 | 0.493 |
| Pearson’s coefficient in ROI | 0.659 | 0.591 | 0.79953 | 0.575 | 0.493 |
| Pearson’s coefficient in volume | 0.131 | 0.224 | 0.321 | 0.21 | 0.457 |
| Thresholded Mander’s coefficient | 0.227 | 0.204 | 0.24873 | 0.127 | 0.09 |
| Accession | Description | Abbrev. | Max. Coverage | Uniq. Pept. | n |
|---|---|---|---|---|---|
| 6671509 | Actin, cytoplasmic 1 | β-actin | 4.53 | 2 | 1 |
| 922959903 | Tubulin alpha-4A chain | α-tubulin | 6.50 | 1 | 1 |
| 568933073 | Anion exchange protein 2 | AE2 | 8.42 | 10 | 4 |
| 334688858 | Sodium-driven chloride bicarb. exch. | Ncbe | 8.37 | 9 | 4 |
| 124517716 | Solute carrier family 12 member 2 | NKCC1 | 25.37 | 27 | 4 |
| 755513830 | Transient receptor pot. cation chan. V4 | TRPv4 | 3.33 | 3 | 4 |
| 295054266 | Spectrin alpha chain, non-erythrocytic 1 | α2-spectrin | 11.18 | 16 | 3 |
| 117938334 | Spectrin beta chain, non-erythrocytic 1 | β2-spectrin | 3.76 | 5 | 3 |
| Accession | Description | Abbrev. | Max. Coverage | Uniq. Pept. | n |
|---|---|---|---|---|---|
| 927028891 | Actin, cytoplasmic 2 | β-actin | 8.30 | 1 | 1 |
| 568933067 | Anion exchange protein 2 | AE2 | 15.90 | 11 | 4 |
| 334688858 | Sodium-driven chloride bicarb. exch. | Ncbe | 4.32 | 3 | 3 |
| 755513830 | Transient receptor pot. cation chan. V4 | TRPv4 | 4.71 | 2 | 2 |
| 569003445 | Solute carrier family 12 member 2 | NKCC1 | 3.28 | 2 | 1 |
| 295054271 | Spectrin alpha chain, non-erythrocytic 1 | α2-spectrin | 0.85 | 1 | 1 |
| 117938334 | Spectrin beta chain, non-erythrocytic 1 | β2-spectrin | 3.85 | 5 | 2 |
| Target cDNA | Forward Primer | Reverse Primer |
|---|---|---|
| Na,K-ATPase α1 | ATCATCGTAGCCAACGTGCCAG | TGCACTTTAAGAGCGCCGACTC |
| Na,K-ATPase α2 | ACTGCCAGGGGCATTGTGATTG | ACATGTGAGCCACTGTCATGCG |
| Na,K-ATPase α3 | ACGCCCATCGCCATTGAGATTG | TCGCATCAGCTTTACGGAACCC |
| Na,K-ATPase α4 | AGGAGCAAACCACGGGGAAAAC | AAGAAGCAGAACCCCAGCACAC |
| Na,K-ATPase β1 | AAAAGCCAAGGAGGAAGGCAGC | TCGGTTTGAAGCCCAACACTCG |
| Na,K-ATPase β2 | AAAGAGAAGAAGAGCTGCGGGC | TGAACTGGCAGGCACGTTTTGG |
| Na,K-ATPase β3 | TTTTCCCAAACCGCAGACTGCC | ACGTCCCAAGAACTTGTCACGC |
| Na,K-ATPase β4 | GTGGCGGAAATTGCAGATCGTG | ACGGAAGCCTACAATCCGGTTC) |
| α1-Spectrin | AAGGAAGAGATTGCTGCTCGCC | AAGCCTTCTGCCTCTGAAAGCC |
| α2-Spectrin | AGCTACCAAACGCAAGCACCAG | TTGACATTCTCCGTTTCGCGGC |
| β1-Spectrin | AGTGAACCTTGCCGCCAACAAC | TCTCAACATCTGCTTGCCGCTG |
| β2-Spectrin | AACAACCGCTGGGATGTGGATG | ATCTGGCACCACAGCAGCAATG |
| β3-Spectrin | CCATTGGTGCGAAGTGCAGAAC | TTGTTTGCGGCACAGCATCC |
| β4-Spectrin | AACGTGGGGTCACATGACATCG | AAACTTGACGGGCTTCTCCAGC |
| β5-Spectrin | TGCCTCGCCAAAGATGTGGAAG | TGCTCAATGCGACTGCTGAACC |
| Target | Antibody | Host | Application | Source |
|---|---|---|---|---|
| Na,K-ATPase α1 | 56-0/3B-0 | Mouse | IB, IHC, PLA, STED | Gift from Forbush, B., 3rd [31] |
| Na,K-ATPase α1 | 05-369 | Rabbit | IP | Millipore |
| Na,K-ATPase α2 | 16836-1 | Rabbit | IB, IHC, PLA, STED | Proteintec |
| Na,K-ATPase β1 | MA3-930 | Mouse | IB, IHC, STED | Thermo Scientific |
| Na,K-ATPase β1 | SpET1 | Rabbit | PLA, STED | Gift from P Martín-Vasallo [8] |
| Na,K-ATPase β2 | SpET2 | Rabbit | IHC | Gift from P Martín-Vasallo [8] |
| Na,K-ATPase β2 | 17369 AP | Rabbit | IHC | Genscript |
| AQP1 | 2343 AP | Rabbit | IB, IHC, PLA, STED | Genscript |
| NKCC1 Nt | BSC2-Nt | Rabbit | IB, IHC, PLA, STED | Gift from J Turner [32] |
| β-actin | B1040 | Rabbit | IHC, PLA, STED | Life Sciences |
| α2-Spectrin | sc-46696 | Mouse | IHC, PLA, STED | Santa Cruz |
| Ankyrin-3 | sc-28561 | Rabbit | IB, IP, IHC, PLA | Santa Cruz |
| 20S Proteasome | sc-67339 | Rabbit | IP | Santa Cruz |
| AE2 | c-terminal | Rabbit | IB, IHC, PLA | Gift from A Stuart-Tilley [14] |
| NCBE | 1139 AP | Rabbit | IB, IHC, PLA | Genscript |
| NBCe2 | 9949 AP | Rabbit | IB, IHC, PLA | Genscript |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baasch Christensen, I.; Cheng, L.; Brewer, J.R.; Bartsch, U.; Fenton, R.A.; Damkier, H.H.; Praetorius, J. Multiple Na,K-ATPase Subunits Colocalize in the Brush Border of Mouse Choroid Plexus Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 1569. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041569
Baasch Christensen I, Cheng L, Brewer JR, Bartsch U, Fenton RA, Damkier HH, Praetorius J. Multiple Na,K-ATPase Subunits Colocalize in the Brush Border of Mouse Choroid Plexus Epithelial Cells. International Journal of Molecular Sciences. 2021; 22(4):1569. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041569
Chicago/Turabian StyleBaasch Christensen, Inga, Lei Cheng, Jonathan R. Brewer, Udo Bartsch, Robert A. Fenton, Helle H. Damkier, and Jeppe Praetorius. 2021. "Multiple Na,K-ATPase Subunits Colocalize in the Brush Border of Mouse Choroid Plexus Epithelial Cells" International Journal of Molecular Sciences 22, no. 4: 1569. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041569