
Inspiratory Off-Switch Mediated by Optogenetic Activation of Inhibitory Neurons in the preBötzinger Complex In Vivo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
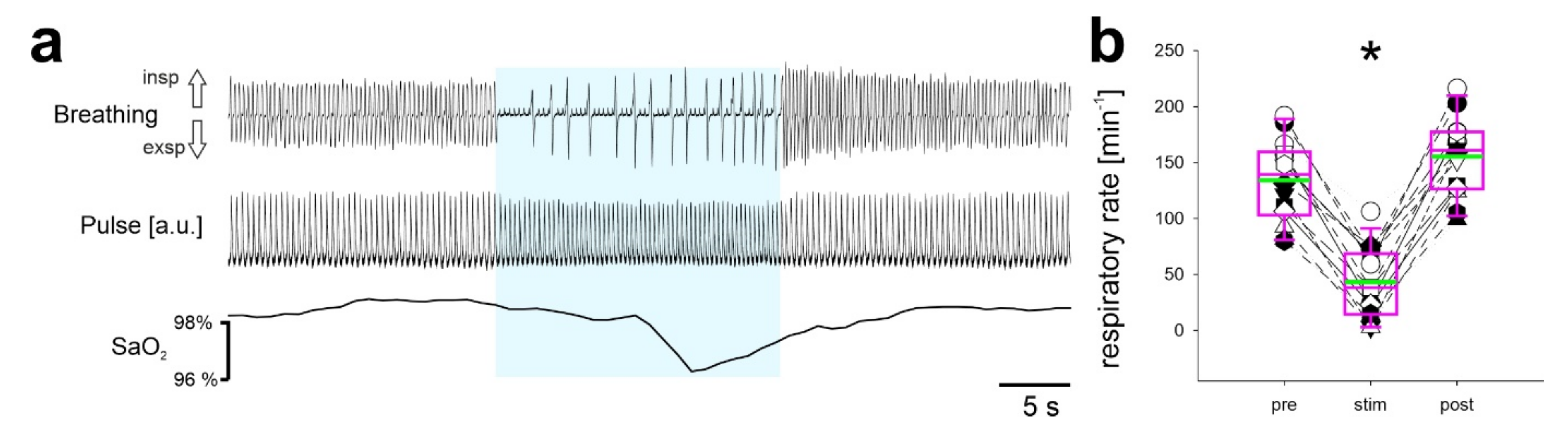
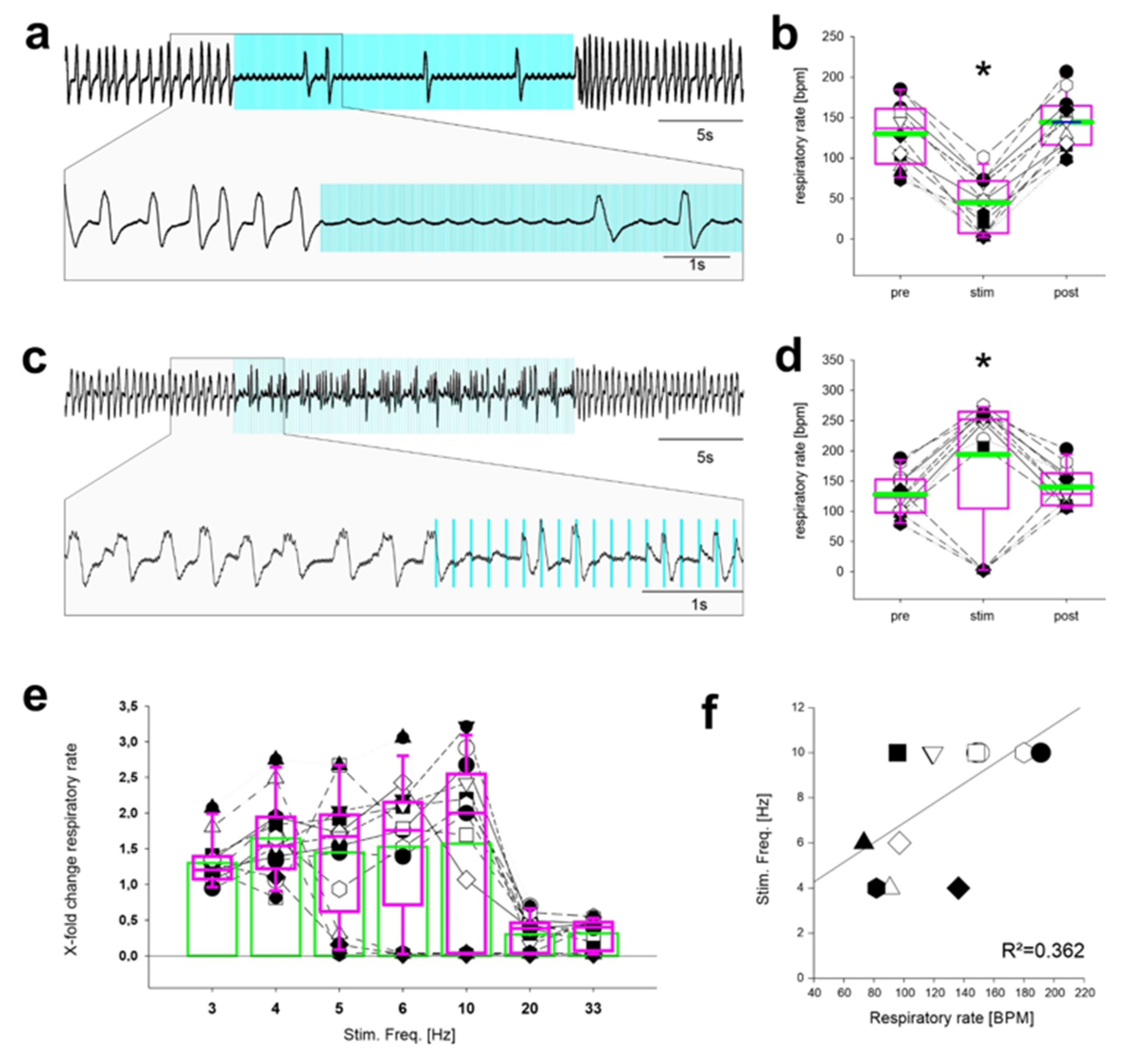
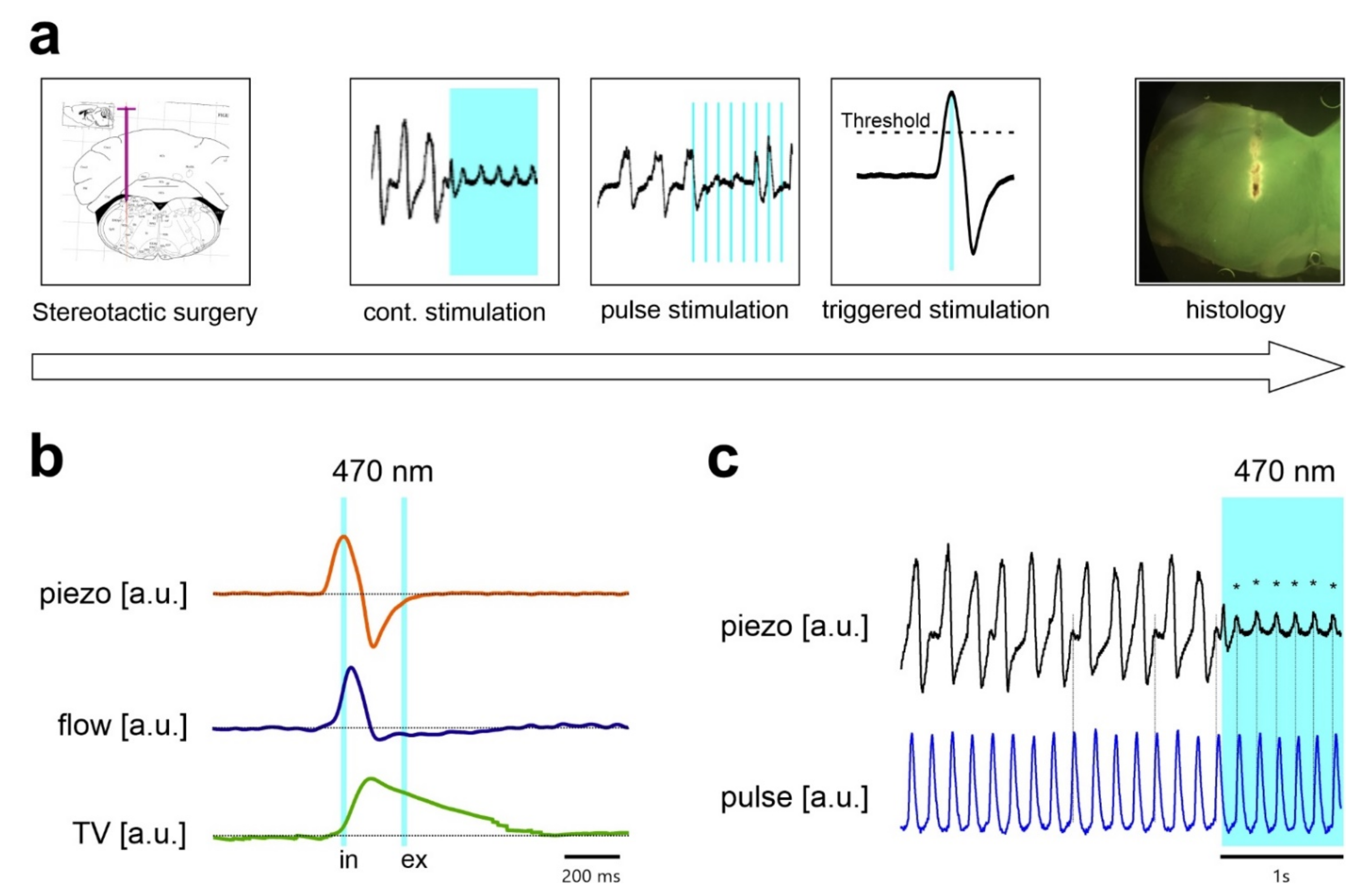
2.1. Continuous Stimulation of Inhibitory Neurons in the preBötC in Anesthetized Mice
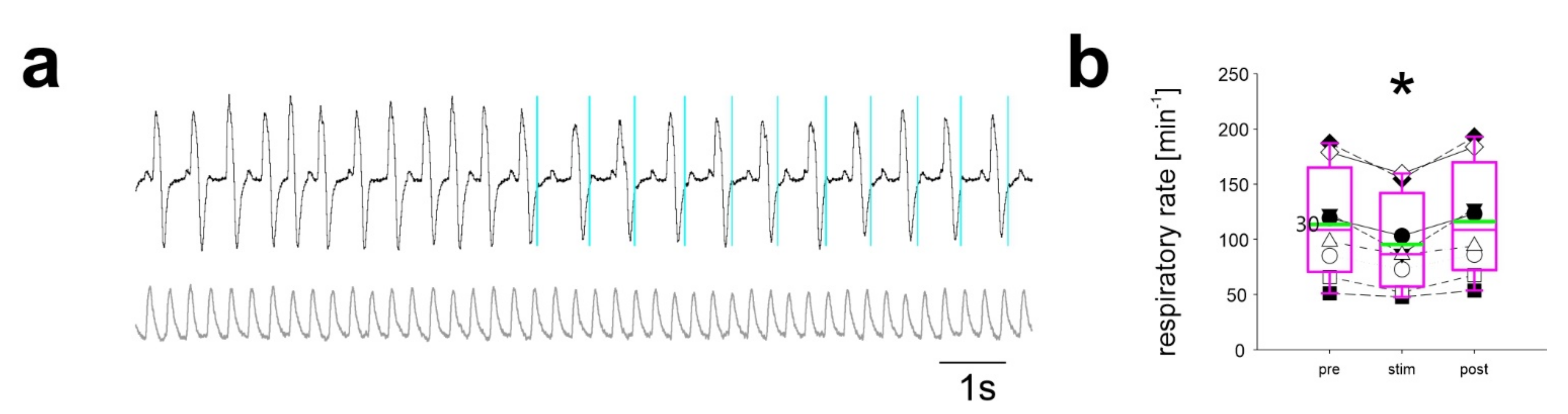
2.2. Stimulation Using Discontinuous Light Pulses with Different Frequencies
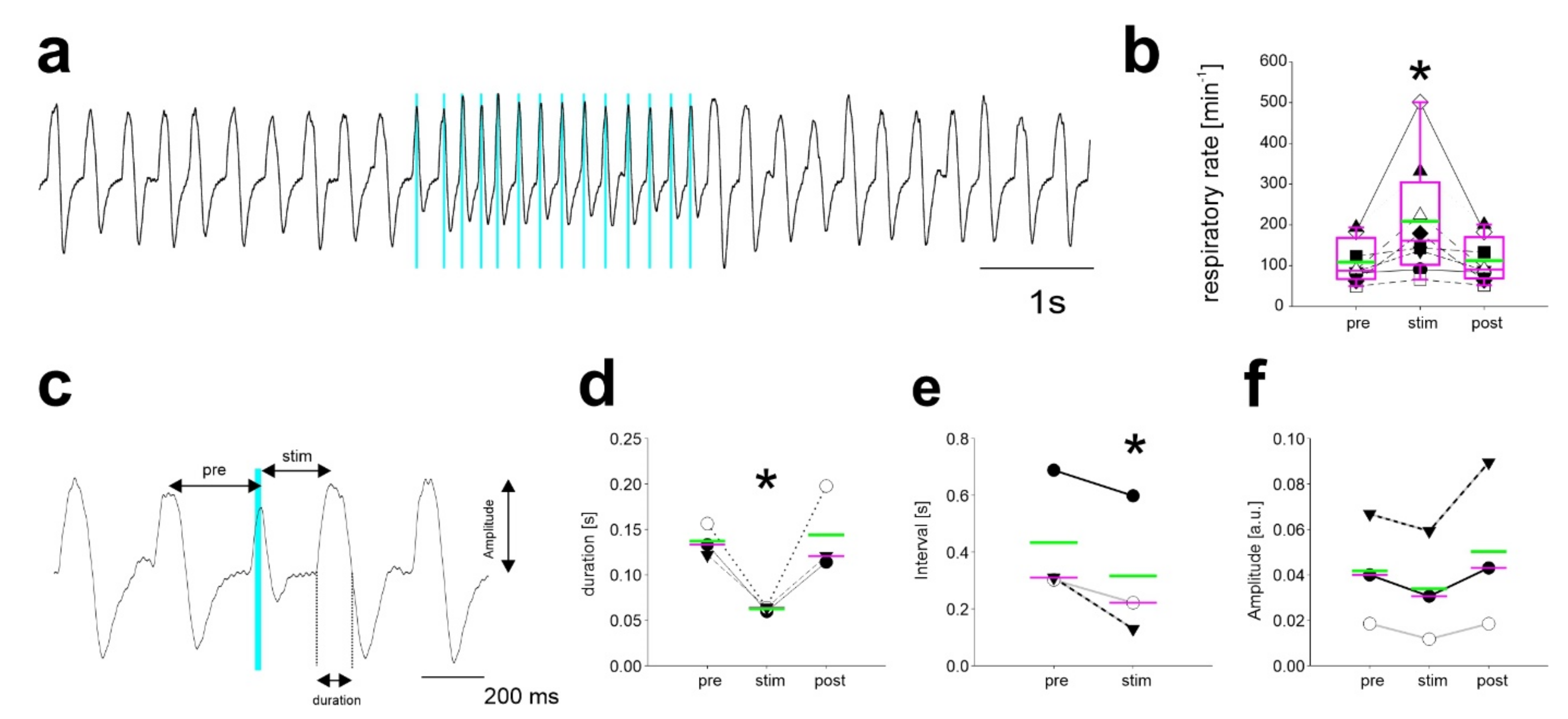
2.3. Repetitive Stimulation of Inhibitory Neurons during Inspiration Increases the Respiratory Rate
3. Discussion
4. Methods
4.1. Ethics and Animal Handling
4.2. Monitoring of Breathing
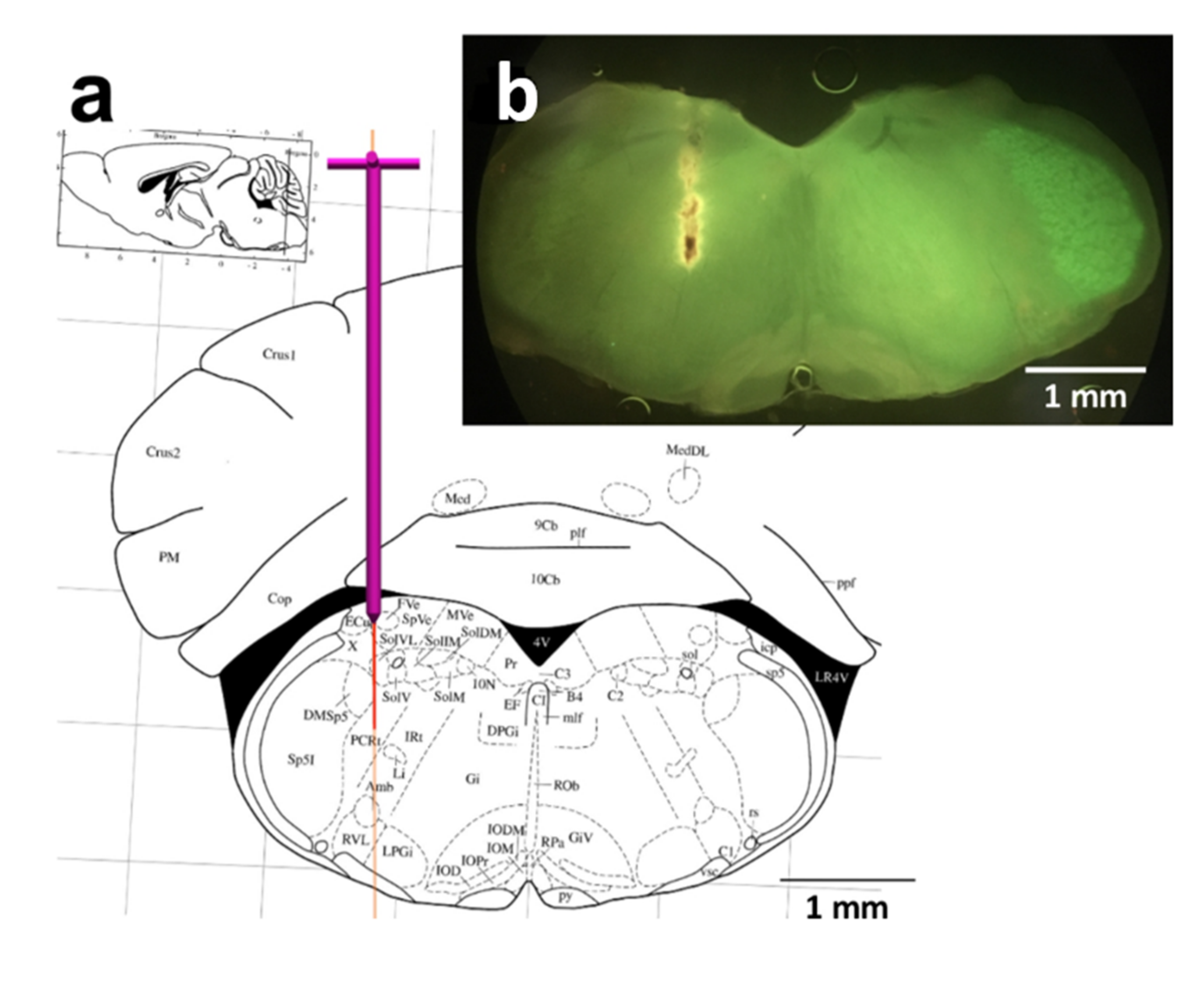
4.3. Stereotactic Implantation of Optical Fiber
4.4. Transcardial Perfusion for Tissue Fixation
4.5. Microscopy and Image Acquisition
4.6. Data Analysis and Presentation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richter, D.W.; Smith, J.C. Respiratory rhythm generation in vivo. Physiology 2014, 29, 58–71. [Google Scholar] [CrossRef] [Green Version]
- Del Negro, C.A.; Funk, G.D.; Feldman, J.L. Breathing matters. Nat. Rev. Neurosci. 2018, 19, 351–367. [Google Scholar] [CrossRef]
- Vann, N.C.; Pham, F.D.; Hayes, J.A.; Kottick, A.; Del Negro, C.A. Transient Suppression of Dbx1 PreBötzinger Interneurons Disrupts Breathing in Adult Mice. PLoS ONE 2016, 11, e0162418. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Janczewski, W.A.; Yang, P.; Shao, X.M.; Callaway, E.M.; Feldman, J.L. Silencing preBotzinger Complex somatostatin-expressing neurons induces persistent apnea in awake rat. Nat. Neurosci. 2008, 11, 538–540. [Google Scholar] [CrossRef] [Green Version]
- Gray, P.A.; Janczewski, W.A.; Mellen, N.; McCrimmon, D.R.; Feldman, J.L. Normal breathing requires preBotzinger complex neurokinin-1 receptor-expressing neurons. Nat. Neurosci. 2001, 4, 927–930. [Google Scholar] [CrossRef]
- Smith, J.C.; Ellenberger, H.H.; Ballanyi, K.; Richter, D.W.; Feldman, J.L. Pre-Botzinger complex: A brainstem region that may generate respiratory rhythm in mammals. Science 1991, 254, 726–729. [Google Scholar] [CrossRef]
- Bautista, T.G.; Pitts, T.E.; Pilowsky, P.M.; Morris, K.F. The Brainstem Respiratory Network. In Neuronal Networks in Brain Function, CNS Disorders, and Therapeutics; Academic Press: Cambridge, MA, USA, 2014; pp. 235–245. [Google Scholar]
- Smith, J.C.; Abdala, A.P.; Borgmann, A.; Rybak, I.A.; Paton, J.F. Brainstem respiratory networks: Building blocks and microcircuits. Trends Neurosci. 2013, 36, 152–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ausborn, J.; Koizumi, H.; Barnett, W.H.; John, T.T.; Zhang, R.; Molkov, Y.I.; Smith, J.C.; Rybak, I.A. Organization of the core respiratory network: Insights from optogenetic and modeling studies. PLoS Comput. Biol. 2018, 14, e1006148. [Google Scholar] [CrossRef] [Green Version]
- Molkov, Y.I.; Rubin, J.E.; Rybak, I.A.; Smith, J.C. Computational models of the neural control of breathing. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierrefiche, O.; Schwarzacher, S.W.; Bischoff, A.M.; Richter, D.W. Blockade of synaptic inhibition within the pre-Botzinger complex in the cat suppresses respiratory rhythm generation in vivo. J. Physiol. 1998, 509 Pt 1, 245–254. [Google Scholar] [CrossRef]
- Janczewski, W.A.; Tashima, A.; Hsu, P.; Cui, Y.; Feldman, J.L. Role of inhibition in respiratory pattern generation. J. Neurosci. 2013, 33, 5454–5465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortuna, M.G.; Kugler, S.; Hulsmann, S. Probing the function of glycinergic neurons in the mouse respiratory network using optogenetics. Respir. Physiol. Neurobiol. 2019, 265, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Baertsch, N.A.; Baertsch, H.C.; Ramirez, J.M. The interdependence of excitation and inhibition for the control of dynamic breathing rhythms. Nat. Commun. 2018, 9, 843. [Google Scholar] [CrossRef]
- Cregg, J.M.; Chu, K.A.; Dick, T.E.; Landmesser, L.T.; Silver, J. Phasic inhibition as a mechanism for generation of rapid respiratory rhythms. Proc. Natl. Acad. Sci. USA 2017, 114, 12815–12820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, D.; Worrell, J.W.; Cui, Y.; Feldman, J.L. Optogenetic perturbation of preBotzinger complex inhibitory neurons modulates respiratory pattern. Nat. Neurosci. 2015, 18, 408–414. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Kam, K.; Sherman, D.; Janczewski, W.A.; Zheng, Y.; Feldman, J.L. Defining preBotzinger Complex Rhythm- and Pattern-Generating Neural Microcircuits In Vivo. Neuron 2016, 91, 602–614. [Google Scholar] [CrossRef] [Green Version]
- Dick, T.E.; Dutschmann, M.; Feldman, J.L.; Fong, A.Y.; Hulsmann, S.; Morris, K.M.; Ramirez, J.M.; Smith, J.C.; Respiratory Neurobiology Consortium. Facts and challenges in respiratory neurobiology. Respir. Physiol. Neurobiol. 2018, 258, 104–107. [Google Scholar] [CrossRef]
- Hirrlinger, J.; Marx, G.; Besser, S.; Sicker, M.; Kohler, S.; Hirrlinger, P.G.; Wojcik, S.M.; Eulenburg, V.; Winkler, U.; Hulsmann, S. GABA-Glycine Cotransmitting Neurons in the Ventrolateral Medulla: Development and Functional Relevance for Breathing. Front. Cell. Neurosci. 2019, 13, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, J.; Besser, S.; Schnell, C.; Eulenburg, V.; Hirrlinger, J.; Wojcik, S.M.; Hulsmann, S. Genetic ablation of VIAAT in glycinergic neurons causes a severe respiratory phenotype and perinatal death. Brain Struct. Funct. 2015, 220, 2835–2849. [Google Scholar] [CrossRef]
- Euler, C.V.; Trippenbach, T. Cyclic Excitability Changes of the Inspiratory ‘Off-switch’ Mechanism. Acta Physiol. Scand. 1975, 93, 560–562. [Google Scholar] [CrossRef]
- Richter, D.W. Generation and maintenance of the respiratory rhythm. J. Exp. Biol. 1982, 100, 93–107. [Google Scholar]
- Kottick, A.; Del Negro, C.A. Synaptic Depression Influences Inspiratory-Expiratory Phase Transition in Dbx1 Interneurons of the preBotzinger Complex in Neonatal Mice. J. Neurosci. 2015, 35, 11606–11611. [Google Scholar] [CrossRef] [Green Version]
- Morgado-Valle, C.; Fernandez-Ruiz, J.; Lopez-Meraz, L.; Beltran-Parrazal, L. Substitution of extracellular Ca2+ by Sr2+ prolongs inspiratory burst in pre-Botzinger complex inspiratory neurons. J. Neurophysiol. 2015, 113, 1175–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumsden, T. The regulation of respiration: Part I. J. Physiol. 1923, 58, 81. [Google Scholar] [CrossRef] [Green Version]
- Ezure, K.; Tanaka, I. GABA, in some cases together with glycine, is used as the inhibitory transmitter by pump cells in the Hering-Breuer reflex pathway of the rat. Neuroscience 2004, 127, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Kubin, L.; Alheid, G.F.; Zuperku, E.J.; McCrimmon, D.R. Central pathways of pulmonary and lower airway vagal afferents. J. Appl. Physiol. 2006, 101, 618–627. [Google Scholar] [CrossRef]
- Oku, Y.; Hulsmann, S. A computational model of the respiratory network challenged and optimized by data from optogenetic manipulation of glycinergic neurons. Neuroscience 2017, 347, 111–122. [Google Scholar] [CrossRef]
- Smith, J.C.; Abdala, A.P.; Rybak, I.A.; Paton, J.F. Structural and functional architecture of respiratory networks in the mammalian brainstem. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2577–2587. [Google Scholar] [CrossRef] [PubMed]
- Dutschmann, M.; Herbert, H. The Kolliker-Fuse nucleus gates the postinspiratory phase of the respiratory cycle to control inspiratory off-switch and upper airway resistance in rat. Eur. J. Neurosci. 2006, 24, 1071–1084. [Google Scholar] [CrossRef]
- Vogelgesang, S. Identifying Target Genes related to Respiratory Network Dysfunction in a Mouse Model for the Rett Syndrome. Ph.D. Thesis, Georg-August-Universität Göttingen, Göttingen, Germany, 2013. [Google Scholar]
- Hartley, C.J.; Taffet, G.E.; Michael, L.H.; Pham, T.T.; Entman, M.L. Noninvasive determination of pulse-wave velocity in mice. Am. J. Physiol. Heart Circ. Physiol. 1997, 273, H494–H500. [Google Scholar] [CrossRef]
- Ishihara, N.; Armsen, W.; Papadopoulos, T.; Betz, H.; Eulenburg, V. Generation of a mouse line expressing Cre recombinase in glycinergic interneurons. Genesis 2010, 48, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Madisen, L.; Mao, T.; Koch, H.; Zhuo, J.-M.; Berenyi, A.; Fujisawa, S.; Hsu, Y.-W.A.; Garcia, A.J.; Gu, X.; Zanella, S.; et al. A toolbox of Cre-dependent optogenetic transgenic mice for light-induced activation and silencing. Nat. Neurosci. 2012, 15, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Drorbaugh, J.E.; Fenn, W.O. A barometric method for measuring ventilation in newborn infants. Pediatrics 1955, 16, 81–87. [Google Scholar] [PubMed]
- Hulsmann, S.; Mesuret, G.; Dannenberg, J.; Arnoldt, M.; Niebert, M. GlyT2-Dependent Preservation of MECP2-Expression in Inhibitory Neurons Improves Early Respiratory Symptoms but Does Not Rescue Survival in a Mouse Model of Rett Syndrome. Front. Physiol. 2016, 7, 385. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Franklin, K.B.J.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hülsmann, S.; Hagos, L.; Eulenburg, V.; Hirrlinger, J. Inspiratory Off-Switch Mediated by Optogenetic Activation of Inhibitory Neurons in the preBötzinger Complex In Vivo. Int. J. Mol. Sci. 2021, 22, 2019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042019
Hülsmann S, Hagos L, Eulenburg V, Hirrlinger J. Inspiratory Off-Switch Mediated by Optogenetic Activation of Inhibitory Neurons in the preBötzinger Complex In Vivo. International Journal of Molecular Sciences. 2021; 22(4):2019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042019
Chicago/Turabian StyleHülsmann, Swen, Liya Hagos, Volker Eulenburg, and Johannes Hirrlinger. 2021. "Inspiratory Off-Switch Mediated by Optogenetic Activation of Inhibitory Neurons in the preBötzinger Complex In Vivo" International Journal of Molecular Sciences 22, no. 4: 2019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042019