
Inflammasome in ALS Skeletal Muscle: NLRP3 as a Potential Biomarker

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. NLRP3 Inflammasome Is Upregulated in Skeletal Muscle of SOD1G93A Mice
2.2. Increased Transcriptional Levels of Nlrp3 in Skeletal Muscle at Earlier Stages Correlated with a Higher Survival Rate in SOD1G93A Mice
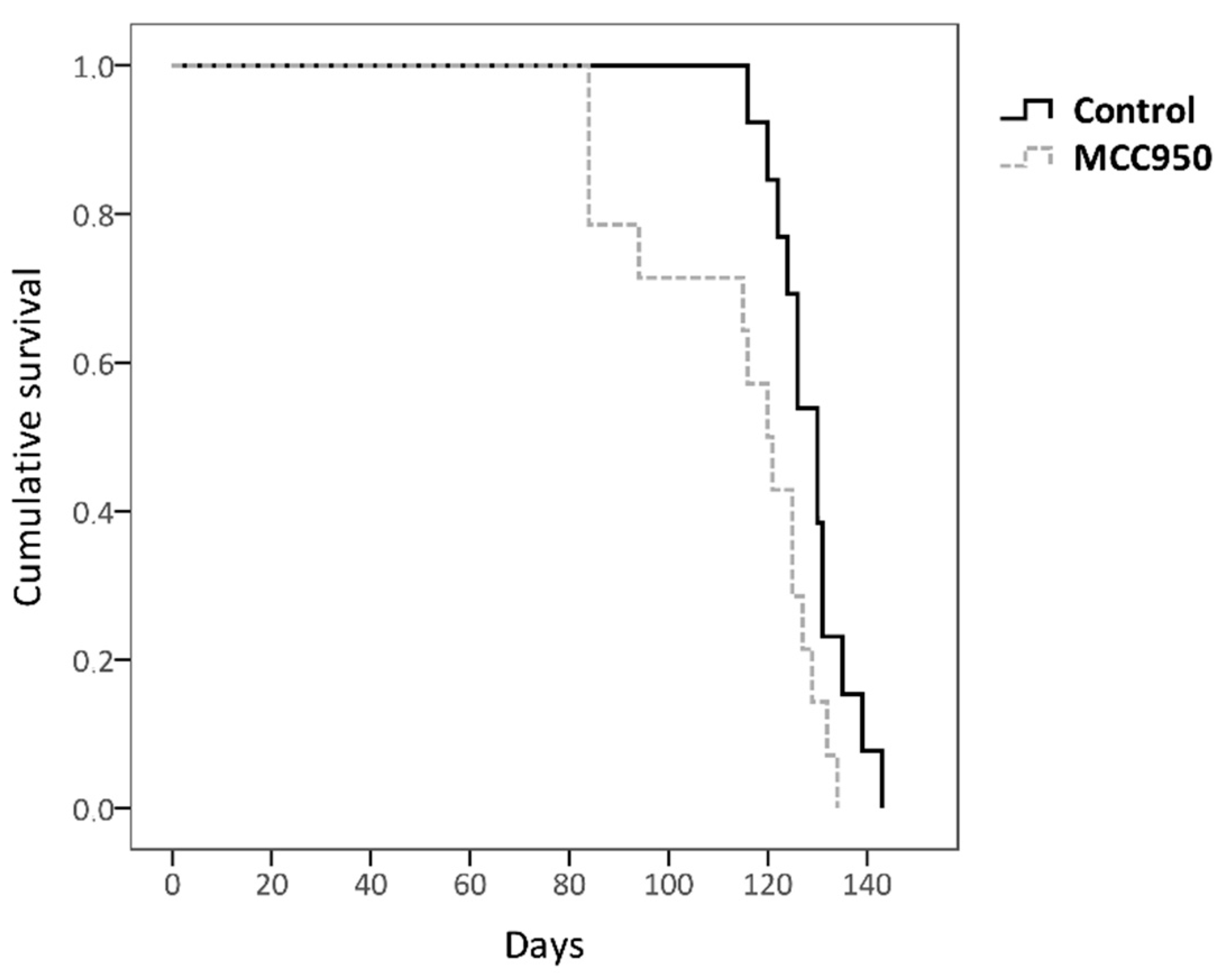
2.3. Inactivation of NLRP3 with MCC950 Exerted a Detrimental Effect on the Longevity of SOD1G93A Mice
2.4. Potential Support for Diagnosis Role of NLRP3 in Blood Samples from ALS Patients
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Tissue Collection
4.2.1. Extraction of Skeletal Muscle Samples in WT and Transgenic SOD1G93A Mice
4.2.2. Extraction of Biopsies from Skeletal Muscle of SOD1G93A Mice
4.3. Administration of MCC950
4.4. Patients
4.5. Blood Samples from Patients
4.6. Gene Expression
4.7. Protein Expression
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ALS | Amyotrophic lateral sclerosis |
| ASC | Apoptosis-associated Speck-like protein containing a CARD |
| DAMPs | Damage-associated molecular patterns |
| FALS | Familial amyotrophic lateral sclerosis |
| NLR | NOD-like receptors |
| NLRC4 | NLR family CARD domain containing 4 |
| NLRP3 | NLR Protein 3 |
| NOD | Nucleotide-binding oligomerization domain |
| OMP | Other myopathies patients |
| P2X7 | P2X purinoreceptor 7 |
| PAMPs | Pathogen-associated molecular patterns |
| PRRs | Pattern recognition receptors |
| SALS | Sporadic amyotrophic lateral sclerosis |
| SOD1 | Superoxide Dismutase 1 |
| TLR | Toll-like receptor |
References
- Calvo, A.C.; Manzano, R.; Mendonça, D.M.; Muñoz, M.J.; Zaragoza, P.; Osta, R. Amyotrophic Lateral Sclerosis: A Focus on Disease Progression. BioMed Res. Int. 2014, 2014, 925101. [Google Scholar] [CrossRef]
- Pradat, P.F.; Dib, M. Biomarkers in amyotrophic lateral sclerosis: Facts and future horizons. Mol. Diagn. Ther. 2009, 13, 115–125. [Google Scholar] [CrossRef]
- Blasco, H.; Vourc’h, P.; Pradat, P.F.; Gordon, P.H.; Andres, C.R.; Corcia, P. Further development of biomarkers in amyotrophic lateral sclerosis. Expert Rev. Mol. Diagn. 2016, 16, 853–868. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Beers, D.R.; Appel, S.H. Immune-mediated mechanisms in the pathoprogression of amyotrophic lateral sclerosis. J. Neuroimmune Pharmacol. 2013, 8, 888–899. [Google Scholar] [CrossRef]
- McCombe, P.A.; Henderson, R.D. The Role of immune and inflammatory mechanisms in ALS. Curr Mol. Med. 2011, 11, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Debye, B.; Schmülling, L.; Zhou, L.; Rune, G.; Beyer, C.; Johann, S. Neurodegeneration and NLRP3 inflammasome expression in the anterior thalamus of SOD1(G93A) ALS mice. Brain Pathol. 2018, 28, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Giacoppo, S.; Bramanti, P.; Mazzon, E. NLRP3 Inflammasome Activation in a Transgenic Amyotrophic Lateral Sclerosis Model. Inflammation 2018, 41, 93–103. [Google Scholar] [CrossRef]
- Kadhim, H.; Deltenre, P.; Martin, J.J.; Sébire, G. In-situ expression of Interleukin-18 and associated mediators in the human brain of sALS patients: Hypothesis for a role for immune-inflammatory mechanisms. Med. Hypotheses 2016, 86, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Johann, S.; Heitzer, M.; Kanagaratnam, M.; Goswami, A.; Rizo, T.; Weis, J.; Troost, D.; Beyer, C. NLRP3 inflammasome is expressed by astrocytes in the SOD1 mouse model of ALS and in human sporadic ALS patients. Glia 2015, 63, 2260–2273. [Google Scholar] [CrossRef]
- Wong, M.; Martin, L.J. Skeletal muscle-restricted expression of human SOD1 causes motor neuron degeneration in transgenic mice. Hum. Mol. Genet. 2010, 19, 2284–2302. [Google Scholar] [CrossRef] [Green Version]
- Coll, R.C.; Robertson, A.A.B.; Chae, J.J.; Higgins, S.C.; Muñoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A small molecule inhibitior of the NLRP3 inflammasome is a potential therapeutic for inflammatory diseases. Nat. Med. 2015, 21, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Abellán, A.; Angosto-Bazarra, D.; Martínez-Banaclocha, H.; de Torre-Minguela, C.; Cerón-Carrasco, J.P.; Pérez-Sánchez, H.; Arostegui, J.I.; Pelegrin, P. MCC950 closes the active conformation of NLRP3 to an inactive state. Nat. Chem. Biol. 2019, 15, 560–564. [Google Scholar] [CrossRef]
- Coll, R.C.; Hill, J.R.; Day, C.J.; Zamoshnikova, A.; Boucher, D.; Massey, N.L.; Chitty, J.L.; Fraser, J.A.; Jennings, M.P.; Robertson, A.A.B.; et al. MCC950 directly targets the NLRP3 ATP-hydrolysis motif for inflammasome inhibition. Nat. Chem. Biol. 2019, 15, 556–559. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, J.M.; Smit-Oistad, I.M.; Macrander, C.; Krakora, D.; Meyer, M.G.; Suzuki, M. Macrophage-mediated inflammation and glial response in the skeletal muscle of a rat model of familial amyotrophic lateral sclerosis (ALS). Exp. Neurol. 2016, 277, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, S.; Esch, E.; Hartmann, P.; Goswami, A.; Nikolin, S.; Weis, J.; Beyer, C.; Johann, S. Expression profile of pattern recognition receptors in skeletal muscle of SOD1(G93A) amyotrophic lateral sclerosis (ALS) mice and sporadic ALS patients. Neuropathol. Appl. Neurobiol. 2018, 44, 606–627. [Google Scholar] [CrossRef] [PubMed]
- Pillon, N.J.; Krook, A. Innate immune receptors in skeletal muscle metabolism. Exp. Cell Res. 2017, 360, 47–54. [Google Scholar] [CrossRef]
- Frost, R.A.; Lang, C.H. Regulation of muscle growth by pathogen-associated molecules. J. Anim Sci. 2008, 86, E84–E93. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Naito, S. Tissue-specific mRNA expression profiles of human toll-like receptors and related genes. Biol. Pharm. Bull. 2005, 28, 886–892. [Google Scholar] [CrossRef] [Green Version]
- Nalbandian, A.; Khan, A.A.; Srivastava, R.; Llewellyn, K.J.; Tan, B.; Shukr, N.; Fazli, Y.; Kimonis, V.E.; BenMohamed, L. Activation of the NLRP3 Inflammasome Is Associated with Valosin-Containing Protein Myopathy. Inflammation 2017, 40, 21–41. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, I.; Kratz, A.K.; Alexanderson, H.; Patarroyo, M. Decreased expression of interleukin-1α, interleukin-1β, and cell adhesion molecules in muscle tissue following corticosteroid treatment in patients with polymyositis and dermatomyositis. Arthritis Rheum. 2000, 43, 336–348. [Google Scholar] [CrossRef]
- McBride, M.J.; Foley, K.P.; D’Souza, D.M.; Li, Y.E.; Lau, T.C.; Hawke, T.J.; Schertzer, J.D. The NLRP3 inflammasome contributes to sarcopenia and lower muscle glycolytic potential in old mice. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E222–E232. [Google Scholar] [CrossRef] [Green Version]
- Boursereau, R.; Abou-Samra, M.; Lecompte, S.; Noel, L.; Brichard, S.M. Downregulation of the NLRP3 inflammasome by adiponectin rescues Duchenne muscular dystrophy. BMC Biol. 2018, 16, 33. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Niu, F.; Chen, J.; Cao, X.; Liu, Z.; Bao, X.; Xu, Y. Ghrelin improves muscle function in dystrophin-deficient mdx mice by inhibiting NLRP3 inflammasome activation. Life Sci. 2019, 232, 116654. [Google Scholar] [CrossRef]
- Huang, N.; Kny, M.; Riediger, F.; Busch, K.; Schmidt, S.; Luft, F.C.; Slevogt, H.; Fielitz, J. Deletion of Nlrp3 protects from inflammation-induced skeletal muscle atrophy. Intensive Care Med. Exp. 2017, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.C.; Manzano, R.; Atencia-Cibreiro, G.; Oliván, S.; Muñoz, M.J.; Zaragoza, P.; Cordero-Vázquez, P.; Esteban-Pérez, J.; García-Redondo, A.; Osta, R. Genetic biomarkers for ALS disease in transgenic SOD1(G93A) mice. PLoS ONE. 2012, 7, e32632. [Google Scholar] [CrossRef] [Green Version]
- Calvo, A.C.; Atencia-Cibreiro, G.; Torre Merino, P.; Roy, J.F.; Galiana, A.; Juárez Rufián, A.; Cano, J.M.; Martín, M.A.; Moreno, L.; Larrodé, P.; et al. Collagen XIX Alpha 1 Improves Prognosis in Amyotrophic Lateral Sclerosis. Aging Dis. 2019, 10, 278–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, R.C.; O’Neill, L.; Schroder, K. Questions and controversies in innate immune research: What is the physiological role of NLRP3? Cell Death Discov. 2016, 2, 16019. [Google Scholar] [CrossRef] [PubMed]
- Deora, V.; Lee, J.D.; Albornoz, E.A.; McAlary, L.; Jagaraj, C.J.; Robertson, A.A.B.; Atkin, J.D.; Cooper, M.A.; Schroder, K.; Yerbury, J.J.; et al. The microglial NLRP3 inflammasome is activated by amyotrophic lateral sclerosis proteins. Glia 2020, 68, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Meissner, F.; Molawi, K.; Zychlinsky, A. Mutant superoxide dismutase 1-induced IL-1beta accelerates ALS pathogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 13046–13050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apolloni, S.; Amadio, S.; Parisi, C.; Matteucci, A.; Potenza, R.L.; Armida, M.; Popoli, P.; D’Ambrosi, N.; Volonté, C. Spinal cord pathology is ameliorated by P2X7 antagonism in a SOD1-mutant mouse model of amyotrophic lateral sclerosis. Dis. Model. Mech. 2014, 7, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, R.; Sluyter, V.; Watson, D.; Sluyter, R.; Yerbury, J.J. P2X7 antagonism using Brilliant Blue G reduces body weight loss and prolongs survival in female SOD1 G93A amyotrophic lateral sclerosis mice. PeerJ 2017, 5, e3064. [Google Scholar] [CrossRef]
- Apolloni, S.; Amadio, S.; Montilli, C.; Volonté, C.; D’ambrosi, N. Ablation of p2X7 receptor exacerbates gliosis and motoneuron death in the SOD1-G93A mouse model of amyotrophic lateral sclerosis. Hum. Mol. Genet. 2013, 22, 4102–4116. [Google Scholar] [CrossRef]
- Tan, M.S.; Yu, J.T.; Jiang, T.; Zhu, X.C.; Tan, L. The NLRP3 inflammasome in Alzheimer’s disease. Mol. Neurobiol. 2013, 48, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Kolb, R.; Liu, G.H.; Janowski, A.M.; Sutterwala, F.S.; Zhang, W. Inflammasomes in cancer: A double-edged sword. Protein Cell 2014, 5, 12–20. [Google Scholar] [CrossRef]
- Wu, J.X.; Shi, K.H. NLRP3, a Double-edged Sword in Lung Injury Diseases. Inflammation 2015, 38, 1855–1856. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves Cambui, R.A.; Ferreira do Espírito Santo, G.; Pereira Fernandes, F.; Vinicius Nunes, C.L.; Borsatto Galera, B.; Palacio Fávaro, E.G.; Amorim Rizzo, L.; Elias, R.M.; Pontillo, A. Double-edged sword of inflammasome genetics in colorectal cancer prognosis. Clin. Immunol. 2020, 213, 108373. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Gao, W.; Loughran, P.; Shapiro, R.; Fan, J.; Billiar, T.R.; Scott, M.J. Caspase 1 activation is protective against hepatocyte cell death by up-regulating beclin 1 protein and mitochondrial autophagy in the setting of redox stress. J. Biol. Chem. 2013, 288, 15947–15958. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Rüegg, A.; Werner, S.; Beer, H.D. Active Caspase-1 Is a Regulator of Unconventional Protein Secretion. Cell 2008, 132, 818–831. [Google Scholar] [CrossRef] [Green Version]
- Zaki, M.H.; Boyd, K.L.; Vogel, P.; Kastan, M.B.; Lamkanfi, M.; Kanneganti, T.D. The NLRP3 Inflammasome Protects against Loss of Epithelial Integrity and Mortality during Experimental Colitis. Immunity 2010, 32, 379–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruchard, M.; Rebé, C.; Derangère, V.; Togbé, D.; Ryffel, B.; Boidot, R.; Humblin, E.; Hamman, A.; Chalmin, F.; Berger, H.; et al. The receptor NLRP3 is a transcriptional regulator of TH2 differentiation. Nat. Immunol. 2015, 16, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Ting, J.P.Y.; Harton, J.A. NLRP3 moonlights in TH2 polarization. Nat. Immunol. 2015, 16, 794–796. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, X.; Miao, Y.; Wang, Y.; Wang, H.; Cheng, Z.; Wang, X.; Jing, X.; Jia, L.; Dai, L.; et al. NLRP3 regulates macrophage M2 polarization through up-regulation of IL-4 in asthma. Biochem. J. 2018, 475, 1995–2008. [Google Scholar] [CrossRef]
- Bruchard, M.; Boidot, R.; Ghiringhelli, F.; Végran, F. Transcriptome analysis of TH2 CD4+ T cells differentiated from wild-type and NLRP3KO mice. Genom. Data 2015, 5, 314–315. [Google Scholar] [CrossRef] [Green Version]
- Kugelberg, E. T cell differentiation: NLRP3 goes beyond the inflammasome. Nat. Rev. Immunol. 2015, 15, 467. [Google Scholar] [CrossRef] [PubMed]
- Horsley, V.; Jansen, K.M.; Mills, S.T.; Pavlath, G.K. IL-4 Acts as a Myoblast Recruitment Factor during Mammalian Muscle Growth. Cell 2003, 113, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Zou, X.; Wu, R.; Zhong, R.; Zhu, D.; Zhang, Y. Accelerated regeneration of the skeletal muscle in RNF13-knockout mice is mediated by macrophage-secreted IL-4/IL-6. Protein Cell. 2014, 5, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondo, T.M.; Mooney, D.J. Functional muscle recovery with nanoparticle-directed M2 macrophage polarization in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 10648–10653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francos-Quijorna, I.; Amo-Aparicio, J.; Martinez-Muriana, A.; López-Vales, R. IL-4 Drives Microglia and Macrophages Toward a Phenotype Conducive for Tissue Repair and Functional Recovery After Spinal Cord Injury. Glia 2016, 64, 2079–2092. [Google Scholar] [CrossRef] [Green Version]
- Rossi, C.; Cusimano, M.; Zambito, M.; Finardi, A.; Capotondo, A.; Garcia-Manteiga, J.M.; Comi, G.; Furlan, R.; Martino, G.; Muzio, L. Interleukin 4 modulates microglia homeostasis and attenuates the early slowly progressive phase of amyotrophic lateral sclerosis. Cell Death Dis. 2018, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Fabbrizio, P.; Apolloni, S.; Bianchi, A.; Salvatori, I.; Valle, C.; Lanzuolo, C.; Bendotti, C.; Nardo, G.; Volonté, C. P2X7 activation enhances skeletal muscle metabolism and regeneration in SOD1G93A mouse model of amyotrophic lateral sclerosis. Brain Pathol. 2020, 30, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Franceschini, A.; Capece, M.; Chiozzi, P.; Falzoni, S.; Sanz, J.M.; Sarti, A.C.; Bonora, M.; Pinton, P.; Di Virgilio, F. The P2X7 receptor directly interacts with the NLRP3 inflammasome scaffold protein. FASEB J. 2015, 29, 2450–2461. [Google Scholar] [CrossRef]
- Di Virgilio, F. The Therapeutic Potential of Modifying Inflammasomes and NOD-Like Receptors. Pharm. Rev. 2013, 65, 872–905. [Google Scholar] [CrossRef] [PubMed]
- Grinstein, L.; Endter, K.; Hedrich, C.M.; Reinke, S.; Luksch, H.; Schulze, F.; Robertson, A.A.B.; Cooper, M.A.; Rösen-Wolff, A.; Winkler, S. An optimized whole blood assay measuring expression and activity of NLRP3, NLRC4 and AIM2 inflammasomes. Clin. Immunol. 2018, 191, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heitzer, M.; Kaiser, S.; Kanagaratnam, M.; Zendedel, A.; Hartmann, P.; Beyer, C.; Johann, S. Administration of 17β-Estradiol Improves Motoneuron Survival and Down-regulates Inflammasome Activation in Male SOD1(G93A) ALS Mice. Mol. Neurobiol. 2017, 54, 8429–8443. [Google Scholar] [CrossRef] [PubMed]
- Gurney, M.E. Transgenic-mouse model of amyotrophic lateral sclerosis. N. Engl. J. Med. 1994, 331, 1721–1722. [Google Scholar] [CrossRef]
- Oliván, S.; Calvo, A.C.; Rando, A.; Muñoz, M.J.; Zaragoza, P.; Osta, R. Comparative study of behavioural tests in the SOD1G93A mouse model of amyotrophic lateral sclerosis. Exp. Anim. 2015, 64, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Dempsey, C.; Rubio Araiz, A.; Bryson, K.J.; Finucane, O.; Larkin, C.; Mills, E.L.; Robertson, A.A.B.; Cooper, M.A.; O’Neill, L.A.J.; Lynch, M.A. Inhibiting the NLRP3 inflammasome with MCC950 promotes non-phlogistic clearance of amyloid-β and cognitive function in APP/PS1 mice. Brain Behav. Immun. 2017, 61, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients’ Characteristics | ALS Patients (n = 14) | OMP Patients (n = 14) | Control Subjects (n = 14) |
|---|---|---|---|
| Gender (n) | 9 males | 10 males | 6 males |
| 5 females | 4 females | 8 females | |
| Age at illness onset (mean ± SD) | 62 ± 8.26 | ||
| Disease duration, months (mean ± SD) | 27 ± 19.18 | ||
| Age at sampling (mean ± SD) | 65 ± 8.23 | 57 ± 13.61 | 58 ± 9.53 |
| Site at illness onset (n patients) Bulbar Upper limb Lower limb Generalized | |||
| 2 | |||
| 4 | |||
| 7 | |||
| 1 |
| Gene | Symbol | Organism | Assay ID |
|---|---|---|---|
| NLR family, pyrin domain containing 3 | Nlrp3 | Mus musculus | Mm00840904_m1 |
| PYD and CARD domain containing | Pycard | Mus musculus | Mm00445747_g1 |
| caspase 1 | Casp1 | Mus musculus | Mm00438023_m1 |
| Interleukin 1 beta | Il1β | Mus musculus | Mm00434228_m1 |
| Glyceraldehyde-3-phosphate dehydrogenase | Gapdh | Mus musculus | 4352932E |
| Actin, beta, cytoplasmic | ß-actin | Mus musculus | 4352933E |
| NLR family pyrin domain containing 3 | NLRP3 | Homo sapiens | Hs00918082_m1 |
| Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | Homo sapiens | Hs02786624_g1 |
| Hypoxanthine phosphoribosyltransferase 1 | HPRT1 | Homo sapiens | Hs02800695_m1 |
| TATA-box binding protein | TBP | Homo sapiens | Hs00427620_m1 |
| Antibody | Host | Dilution | Reference Number |
|---|---|---|---|
| Cryopyrin (H-66) | Rabbit | 1:250 | sc-66846 |
| ASC (N-15)-R | Rabbit | 1:250 | sc-22514-R |
| caspase-1 p20 (M-19) | Rabbit | 1:250 | sc-1218-R |
| IL-1ß (H-153) | Rabbit | 1:250 | sc-7884 |
| GAPDH (FL-335) | Rabbit | 1:1000 | sc-25778 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-García, L.; Miana-Mena, F.J.; Moreno-Martínez, L.; de la Torre, M.; Lunetta, C.; Tarlarini, C.; Zaragoza, P.; Calvo, A.C.; Osta, R. Inflammasome in ALS Skeletal Muscle: NLRP3 as a Potential Biomarker. Int. J. Mol. Sci. 2021, 22, 2523. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052523
Moreno-García L, Miana-Mena FJ, Moreno-Martínez L, de la Torre M, Lunetta C, Tarlarini C, Zaragoza P, Calvo AC, Osta R. Inflammasome in ALS Skeletal Muscle: NLRP3 as a Potential Biomarker. International Journal of Molecular Sciences. 2021; 22(5):2523. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052523
Chicago/Turabian StyleMoreno-García, Leticia, Francisco J. Miana-Mena, Laura Moreno-Martínez, Miriam de la Torre, Christian Lunetta, Claudia Tarlarini, Pilar Zaragoza, Ana Cristina Calvo, and Rosario Osta. 2021. "Inflammasome in ALS Skeletal Muscle: NLRP3 as a Potential Biomarker" International Journal of Molecular Sciences 22, no. 5: 2523. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052523